5-Azacytidine Induces Anoikis, Inhibits Mammosphere Formation and Reduces Metalloproteinase 9 Activity in MCF-7 Human Breast Cancer Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
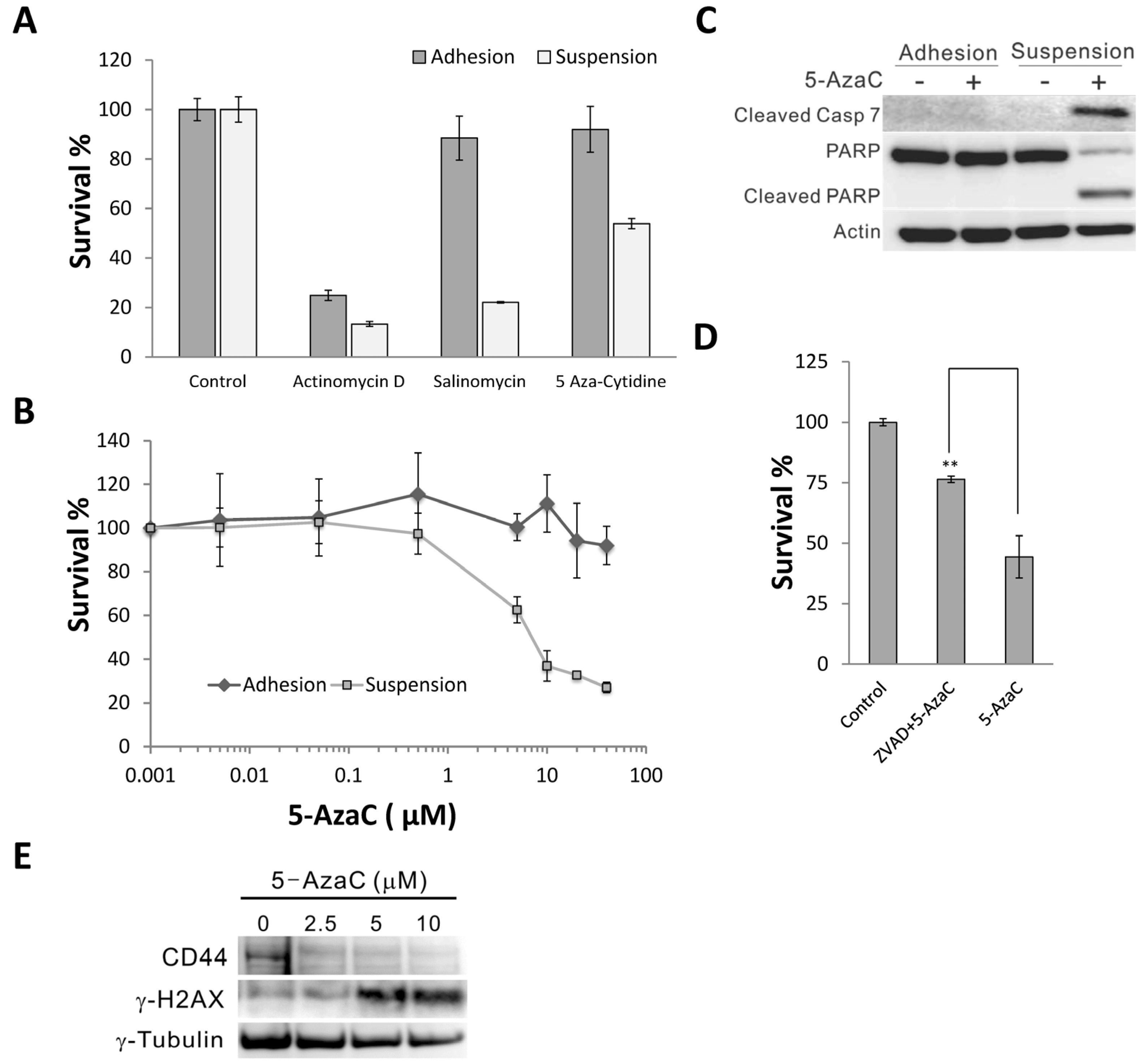
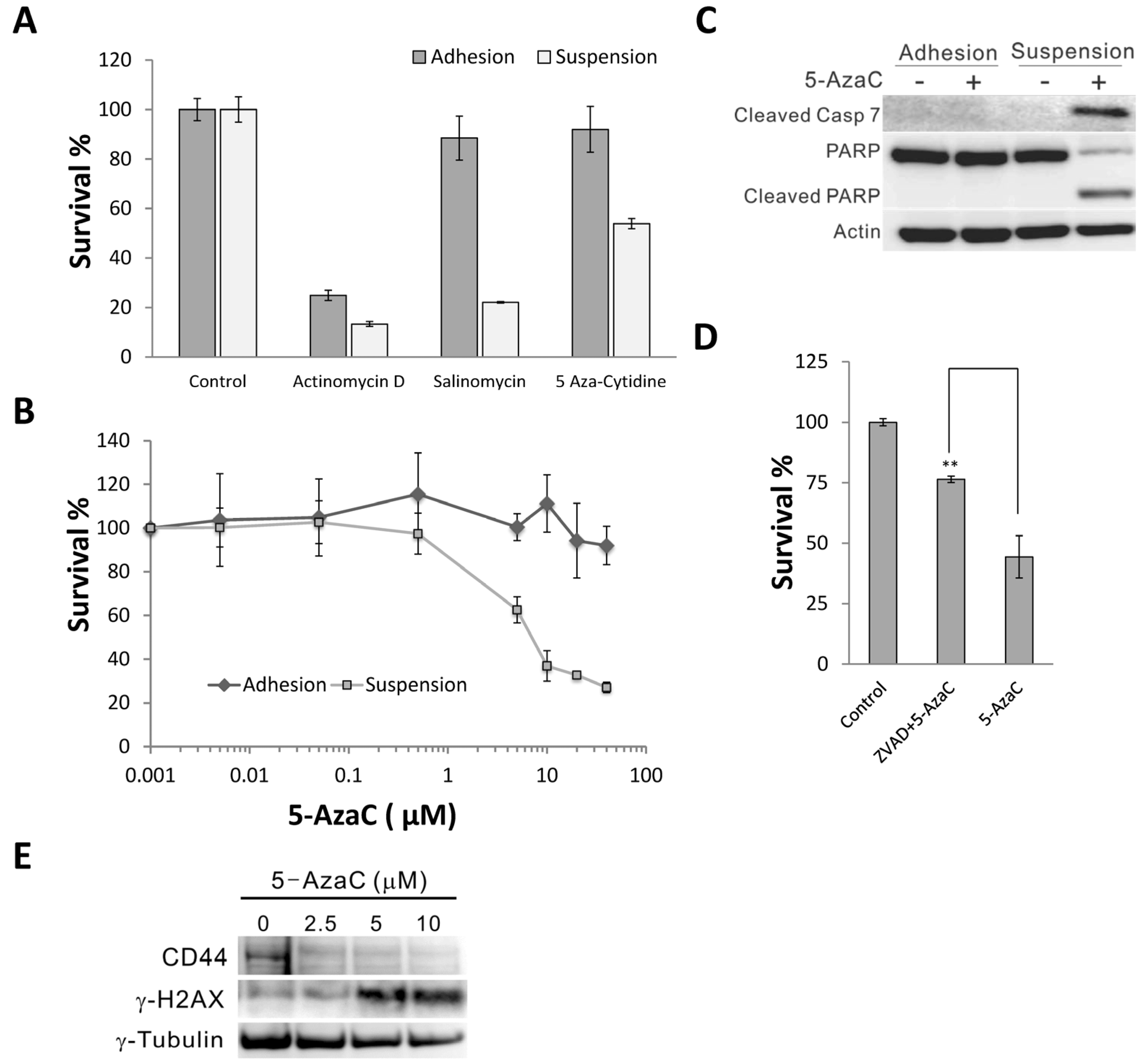
2.1. 5-Azacytidine Sensitizes MCF-7 Cells to Anoikis
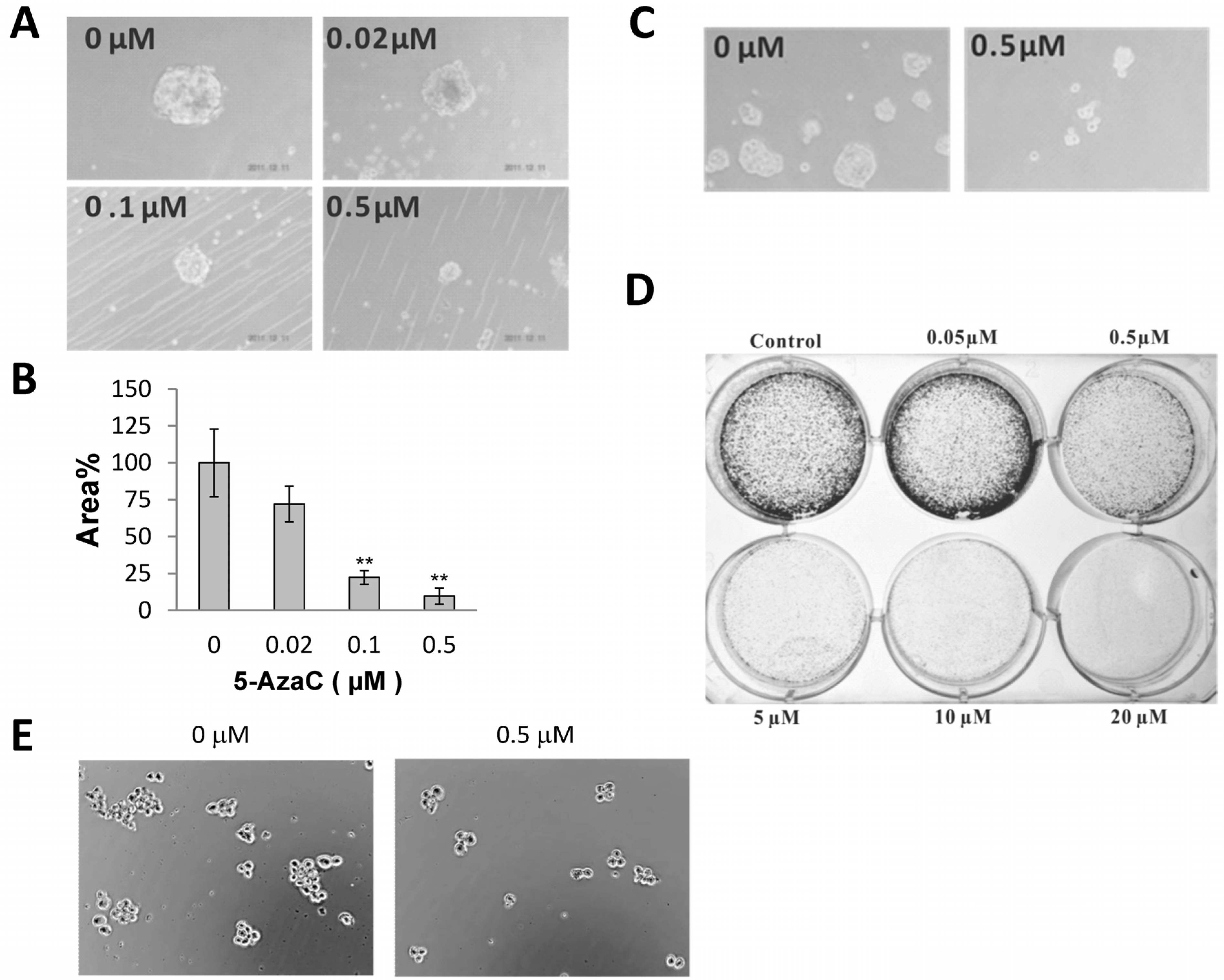
2.2. 5-AzaC Reduces the Clonogenicity of MCF-7 Cells

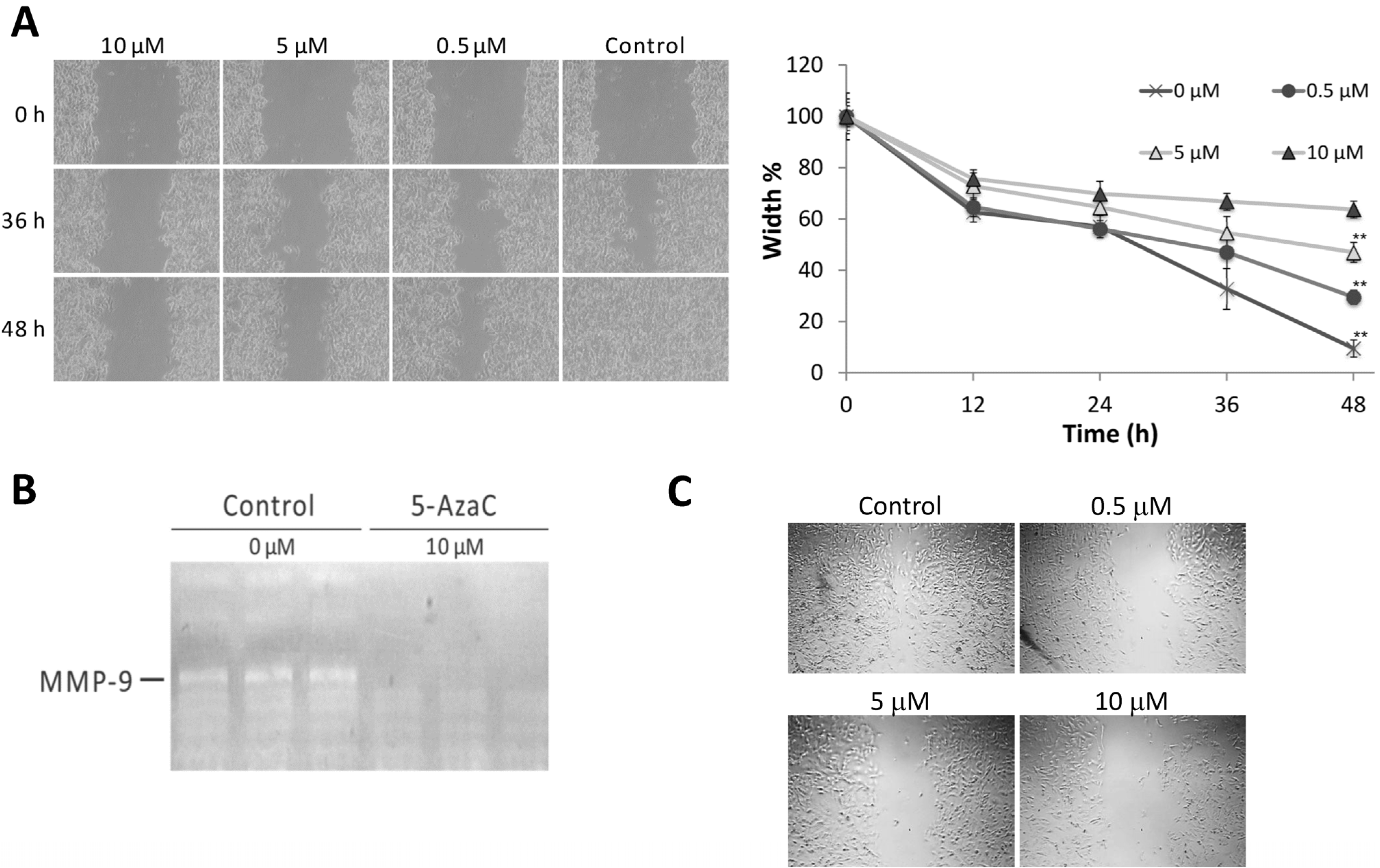
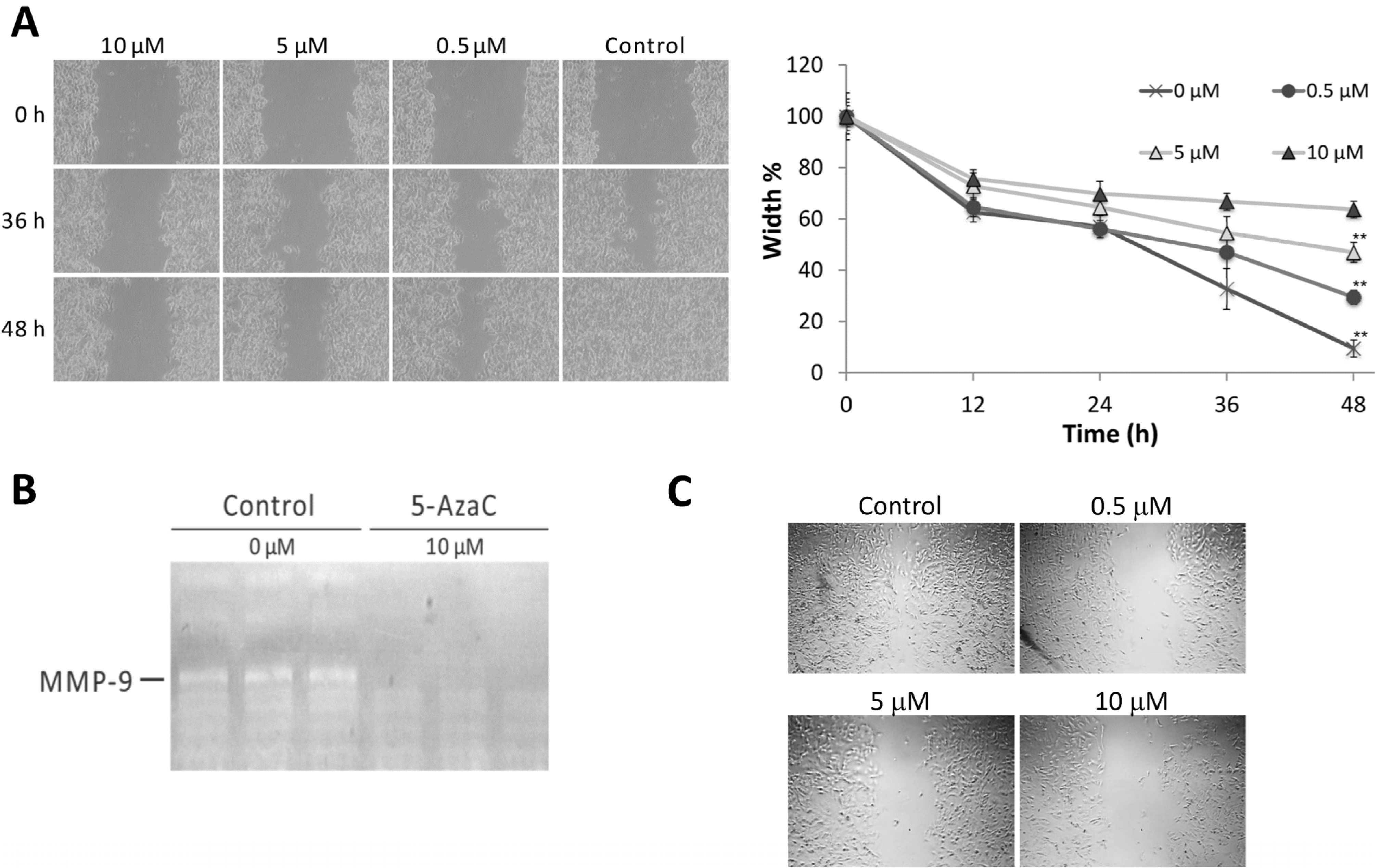
2.3. 5-AzaC Inhibits the Migration and Metalloproteinase 9 Activity of MCF-7 Cells
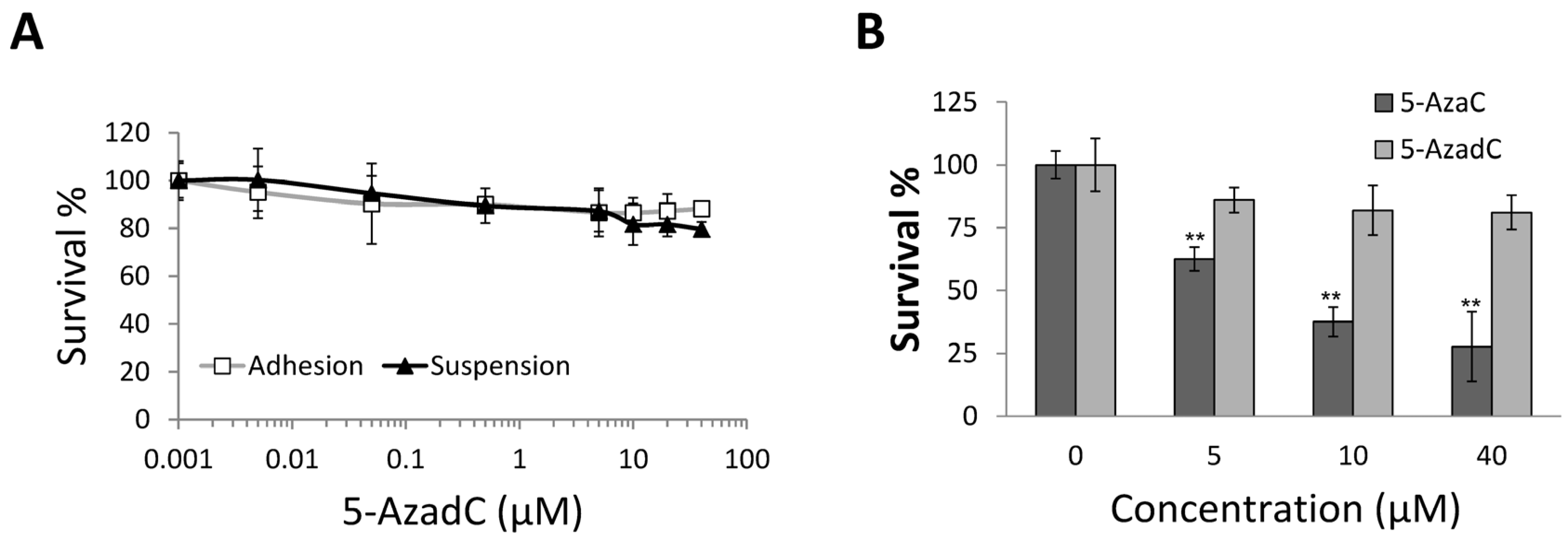
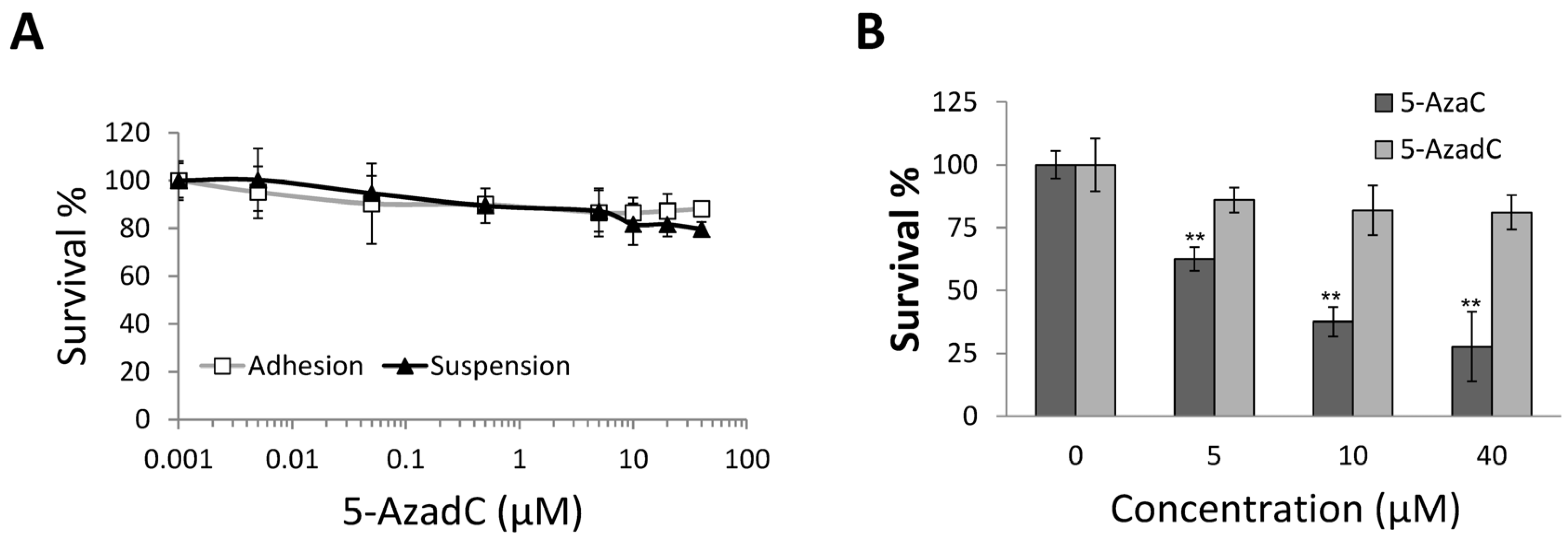
2.4. 5-AzadC Is Not Effective in Inducing MCF-7 Anoikis
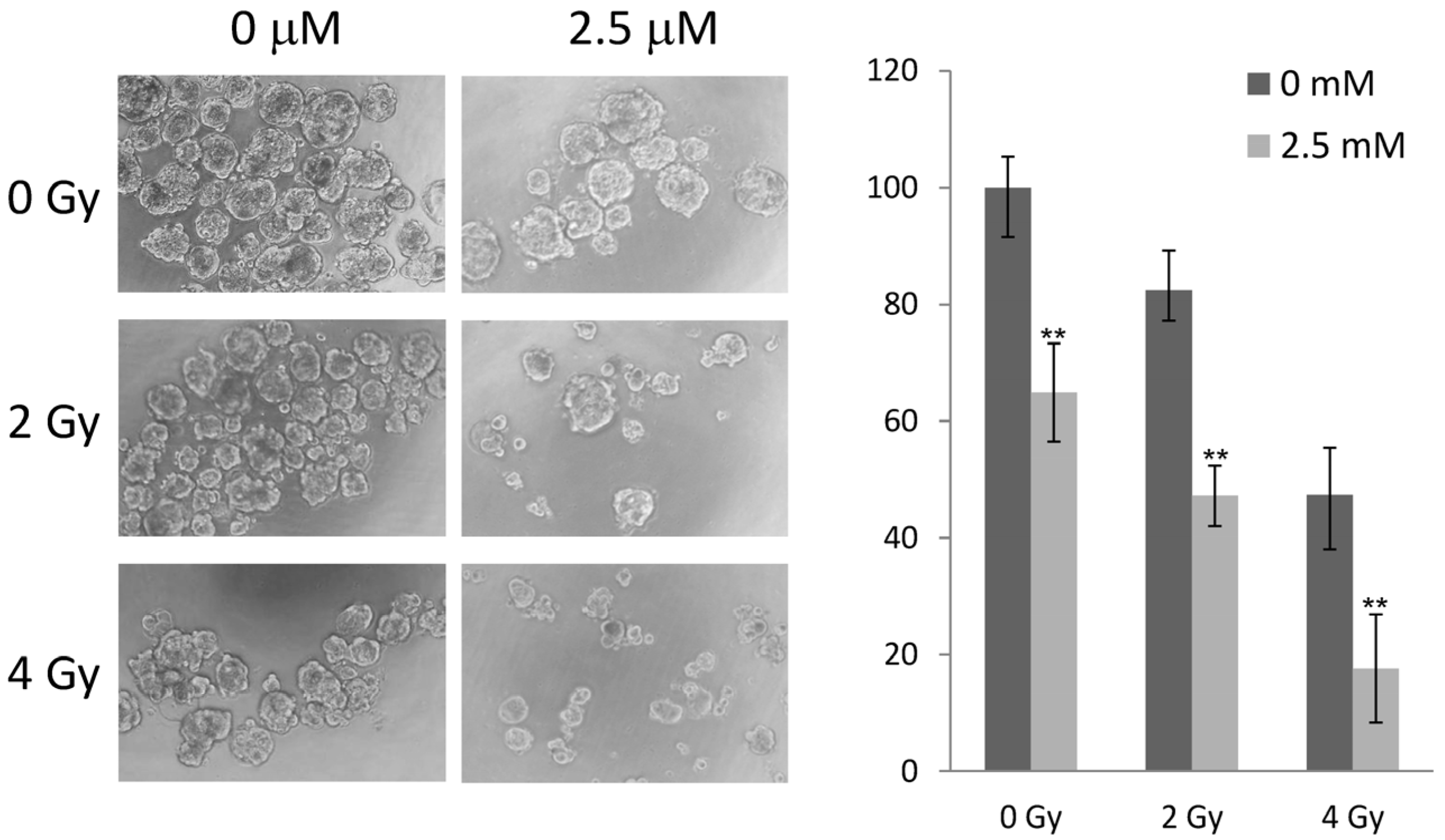
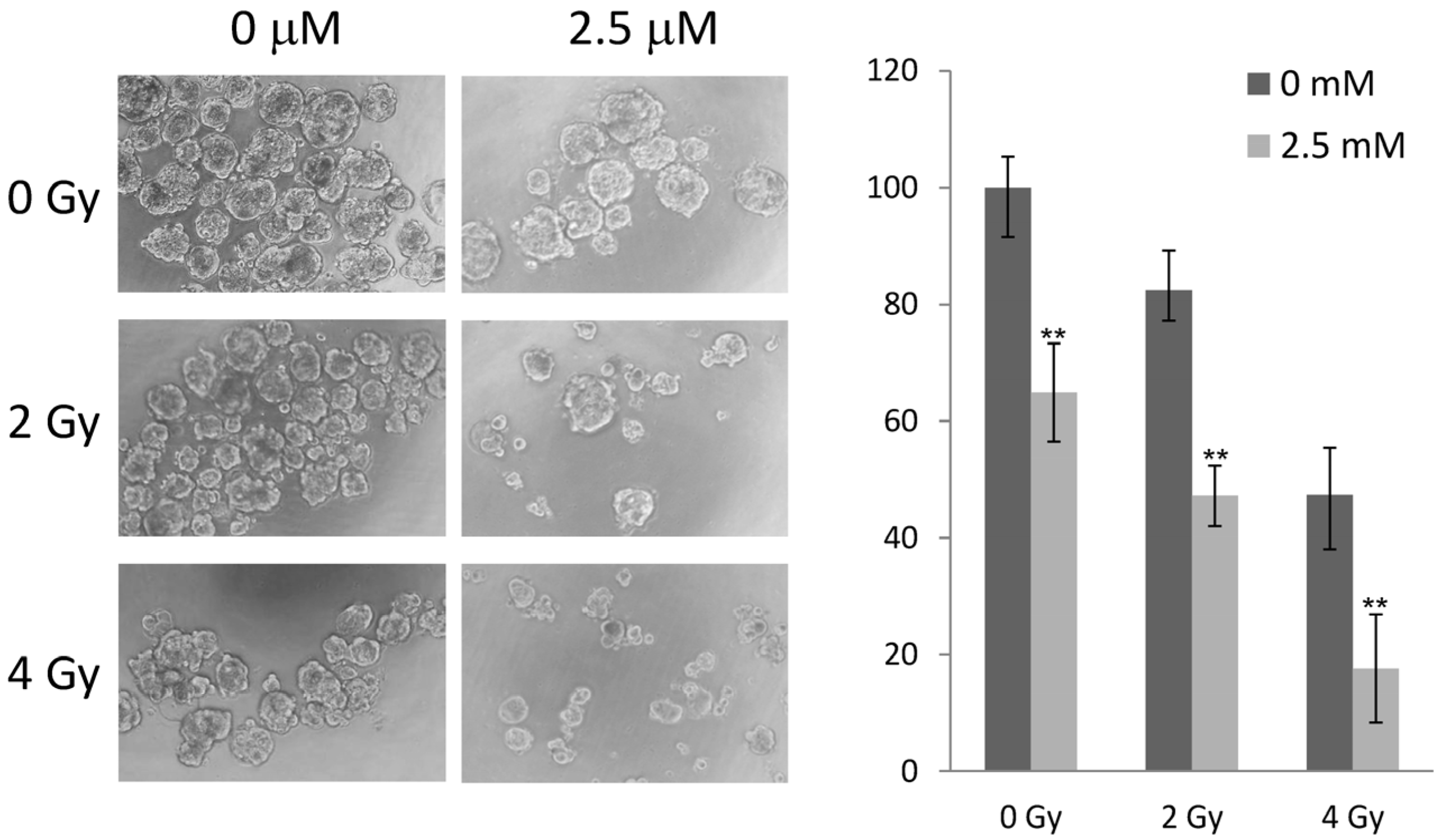
2.5. 5-AzaC and Radiation Therapy Collaboratively Reduced the Growth of MCF-7 Tumorspheres
3. Experimental
3.1. Cell Culture
3.2. Assessment of Cellular Viability by Trypan Blue
3.3. WST-1 Assay
3.4. Tumorsphere Formation Assay
3.5. Soft Agar Colony Formation Assay
3.6. Clonogenic Assay
3.7. Wound-Healing Motility Assay
3.8. Gelatin Zymography
3.9. Western Blotting
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflictts of Interest
References
- Liu, S.; Dontu, G.; Wicha, M.S. Mammary stem cells, self-renewal pathways, and carcinogenesis. Breast Cancer Res. 2005, 7, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Korkaya, H.; Paulson, A.; Charafe-Jauffret, E.; Ginestier, C.; Brown, M.; Dutcher, J.; Clouthier, S.G.; Wicha, M.S. Regulation of mammary stem/progenitor cells by PTEN/Akt/beta-catenin signaling. PLoS Biol. 2009, 7, e1000121. [Google Scholar] [CrossRef] [Green Version]
- Wicha, M.S.; Liu, S.; Dontu, G. Cancer stem cells: An old idea--a paradigm shift. Cancer Res. 2006, 66, 1883–1890; Discussion 1895–1896. [Google Scholar] [CrossRef]
- Soltysova, A.; Altanerova, V.; Altaner, C. Cancer stem cells. Neoplasma 2005, 52, 435–440. [Google Scholar]
- Ailles, L.E.; Weissman, I.L. Cancer stem cells in solid tumors. Curr. Opin. Biotechnol. 2007, 18, 460–466. [Google Scholar] [CrossRef]
- Polyak, K.; Weinberg, R.A. Transitions between epithelial and mesenchymal states: Acquisition of malignant and stem cell traits. Nat. Rev. Cancer 2009, 9, 265–273. [Google Scholar] [CrossRef]
- Ponti, D.; Costa, A.; Zaffaroni, N.; Pratesi, G.; Petrangolini, G.; Coradini, D.; Pilotti, S.; Pierotti, M.A.; Daidone, M.G. Isolation and in vitro propagation of tumorigenic breast cancer cells with stem/progenitor cell properties. Cancer Res. 2005, 65, 5506–5511. [Google Scholar] [CrossRef]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef]
- Cioce, M.; Gherardi, S.; Viglietto, G.; Strano, S.; Blandino, G.; Muti, P.; Ciliberto, G. Mammosphere-forming cells from breast cancer cell lines as a tool for the identification of CSC-like- and early progenitor-targeting drugs. Cell Cycle 2010, 9, 2878–2887. [Google Scholar]
- Rappa, G.; Lorico, A. Phenotypic characterization of mammosphere-forming cells from the human MA-11 breast carcinoma cell line. Exp. Cell Res. 2010, 316, 1576–1586. [Google Scholar] [CrossRef]
- Gupta, P.B.; Onder, T.T.; Jiang, G.; Tao, K.; Kuperwasser, C.; Weinberg, R.A.; Lander, E.S. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 2009, 138, 645–659. [Google Scholar] [CrossRef]
- Tseng, C.N.; Huang, C.F.; Cho, C.L.; Chang, H.W.; Huang, C.W.; Chiu, C.C.; Chang, Y.F. Brefeldin a effectively inhibits cancer stem cell-like properties and MMP-9 activity in human colorectal cancer colo 205 cells. Molecules 2013, 18, 10242–10253. [Google Scholar] [CrossRef]
- Lu, S.; Labhasetwar, V. Drug resistant breast cancer cell line displays cancer stem cell phenotype and responds sensitively to epigenetic drug SAHA. Drug Deliv. Transl. Res. 2013, 3, 183–194. [Google Scholar] [CrossRef]
- Nicolini, A.; Ferrari, P.; Fini, M.; Borsari, V.; Fallahi, P.; Antonelli, A.; Berti, P.; Carpi, A.; Miccoli, P. Stem cells: Their role in breast cancer development and resistance to treatment. Curr. Pharm. Biotechnol. 2011, 12, 196–205. [Google Scholar] [CrossRef]
- Lawson, J.C.; Blatch, G.L.; Edkins, A.L. Cancer stem cells in breast cancer and metastasis. Breast Cancer Res. Treat 2009, 118, 241–254. [Google Scholar] [CrossRef]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; Dewhirst, M.W.; Bigner, D.D.; Rich, J.N. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006, 444, 756–760. [Google Scholar] [CrossRef]
- Diehn, M.; Cho, R.W.; Lobo, N.A.; Kalisky, T.; Dorie, M.J.; Kulp, A.N.; Qian, D.; Lam, J.S.; Ailles, L.E.; Wong, M.; et al. Association of reactive oxygen species levels and radioresistance in cancer stem cells. Nature 2009, 458, 780–783. [Google Scholar] [CrossRef]
- Wang, J.L.; Sun, Y.; Wu, S. Gamma-irradiation induces matrix metalloproteinase II expression in a p53-dependent manner. Mol. Carcinog. 2000, 27, 252–258. [Google Scholar] [CrossRef]
- Kargiotis, O.; Geka, A.; Rao, J.; Kyritsis, A. Effects of irradiation on tumor cell survival, invasion and angiogenesis. J. NeuroOncol. 2010, 100, 323–338. [Google Scholar] [CrossRef]
- Cheng, J.C.H.; Chou, C.H.; Kuo, M.L.; Hsieh, C.Y. Radiation-enhanced hepatocellular carcinoma cell invasion with MMP-9 expression through PI3K//Akt//NF-[kappa]B signal transduction pathway. Oncogene 2006, 25, 7009–7018. [Google Scholar] [CrossRef]
- Zhou, Y.-C.; Liu, J.-Y.; Li, J.; Zhang, J.; Xu, Y.-Q.; Zhang, H.-W.; Qiu, L.-B.; Ding, G.-R.; Su, X.-M.; Mei, S.; et al. Ionizing radiation promotes migration and invasion of cancer cells through transforming growth factor-beta–mediated epithelial–mesenchymal transition. Int. J. Radiat. Oncol. Biol. Phys. 2011, 81, 1530–1537. [Google Scholar] [CrossRef]
- Trainer, D.L.; Kline, T.; Mallon, F.; Greig, R.; Poste, G. Effect of 5-azacytidine on DNA methylation and the malignant properties of B16 melanoma cells. Cancer Res. 1985, 45, 6124–6130. [Google Scholar]
- Kaminskas, E.; Farrell, A.; Abraham, S.; Baird, A.; Hsieh, L.S.; Lee, S.L.; Leighton, J.K.; Patel, H.; Rahman, A.; Sridhara, R.; et al. Fda Approval summary: Azacitidine for treatment of myelodysplastic syndrome subtypes. Clin. Cancer Res. 2005, 11, 3604–3608. [Google Scholar] [CrossRef]
- Qiu, X.; Hother, C.; Ralfkiaer, U.M.; Sogaard, A.; Lu, Q.; Workman, C.T.; Liang, G.; Jones, P.A.; Gronbaek, K. Equitoxic doses of 5-azacytidine and 5-aza-2'deoxycytidine induce diverse immediate and overlapping heritable changes in the transcriptome. PLoS One 2010, 5, e12994. [Google Scholar] [CrossRef]
- Snyder, R.D.; Lachmann, P.J. Differential effects of 5-azacytidine and 5-azadeoxycytidine on cytotoxicity, DNA-strand breaking and repair of X-ray-induced DNA damage in HeLa cells. Mutat. Res. 1989, 226, 185–190. [Google Scholar] [CrossRef]
- De Carvalho, D.D.; You, J.S.; Jones, P.A. DNA methylation and cellular reprogramming. Trends Cell Biol. 2010, 20, 609–617. [Google Scholar] [CrossRef]
- Mikkelsen, T.S.; Hanna, J.; Zhang, X.; Ku, M.; Wernig, M.; Schorderet, P.; Bernstein, B.E.; Jaenisch, R.; Lander, E.S.; Meissner, A. Dissecting direct reprogramming through integrative genomic analysis. Nature 2008, 454, 49–55. [Google Scholar] [CrossRef]
- Charafe-Jauffret, E.; Ginestier, C.; Iovino, F.; Tarpin, C.; Diebel, M.; Esterni, B.; Houvenaeghel, G.; Extra, J.M.; Bertucci, F.; Jacquemier, J.; et al. Aldehyde dehydrogenase 1-positive cancer stem cells mediate metastasis and poor clinical outcome in inflammatory breast cancer. Clin. Cancer Res. 2010, 16, 45–55. [Google Scholar] [CrossRef]
- Hiraga, T.; Ito, S.; Nakamura, H. Cancer stem-like cell marker CD44 promotes bone metastases by enhancing tumorigenicity, cell motility, and hyaluronan production. Cancer Res. 2013, 73, 4112–4122. [Google Scholar] [CrossRef]
- Yen, C.Y.; Chiu, C.C.; Chang, F.R.; Chen, J.Y.; Hwang, C.C.; Hseu, Y.C.; Yang, H.L.; Lee, A.Y.; Tsai, M.T.; Guo, Z.L.; et al. 4beta-Hydroxywithanolide E from Physalis peruviana (golden berry) inhibits growth of human lung cancer cells through DNA damage, apoptosis and G2/M arrest. BMC Cancer 2010, 10. [Google Scholar] [CrossRef]
- Chang, Y.F.; Chou, H.J.; Yen, Y.C.; Chang, H.W.; Hong, Y.R.; Huang, H.W.; Tseng, C.N. Agrin induces association of Chrna1 mRNA and nicotinic acetylcholine receptor in C2C12 myotubes. FEBS Lett. 2012, 586, 3111–3116. [Google Scholar] [CrossRef]
- Sample Availability: 5-AzaC (A2385) and 5-AzadC (A3656) are available from Sigma-Aldrich (St. Louis, MO, USA).
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, H.-W.; Wang, H.-C.; Chen, C.-Y.; Hung, T.-W.; Hou, M.-F.; Yuan, S.-S.F.; Huang, C.-J.; Tseng, C.-N. 5-Azacytidine Induces Anoikis, Inhibits Mammosphere Formation and Reduces Metalloproteinase 9 Activity in MCF-7 Human Breast Cancer Cells. Molecules 2014, 19, 3149-3159. https://doi.org/10.3390/molecules19033149
Chang H-W, Wang H-C, Chen C-Y, Hung T-W, Hou M-F, Yuan S-SF, Huang C-J, Tseng C-N. 5-Azacytidine Induces Anoikis, Inhibits Mammosphere Formation and Reduces Metalloproteinase 9 Activity in MCF-7 Human Breast Cancer Cells. Molecules. 2014; 19(3):3149-3159. https://doi.org/10.3390/molecules19033149
Chicago/Turabian StyleChang, Hsueh-Wei, Hui-Chun Wang, Chiau-Yi Chen, Ting-Wei Hung, Ming-Feng Hou, Shyng-Shiou F. Yuan, Chih-Jen Huang, and Chao-Neng Tseng. 2014. "5-Azacytidine Induces Anoikis, Inhibits Mammosphere Formation and Reduces Metalloproteinase 9 Activity in MCF-7 Human Breast Cancer Cells" Molecules 19, no. 3: 3149-3159. https://doi.org/10.3390/molecules19033149
APA StyleChang, H.-W., Wang, H.-C., Chen, C.-Y., Hung, T.-W., Hou, M.-F., Yuan, S.-S. F., Huang, C.-J., & Tseng, C.-N. (2014). 5-Azacytidine Induces Anoikis, Inhibits Mammosphere Formation and Reduces Metalloproteinase 9 Activity in MCF-7 Human Breast Cancer Cells. Molecules, 19(3), 3149-3159. https://doi.org/10.3390/molecules19033149