Methanolic Extracts of Solieria robusta Inhibits Proliferation of Oral Cancer Ca9-22 Cells via Apoptosis and Oxidative Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cell Viability of MESR-Incubated Ca9-22 Cells
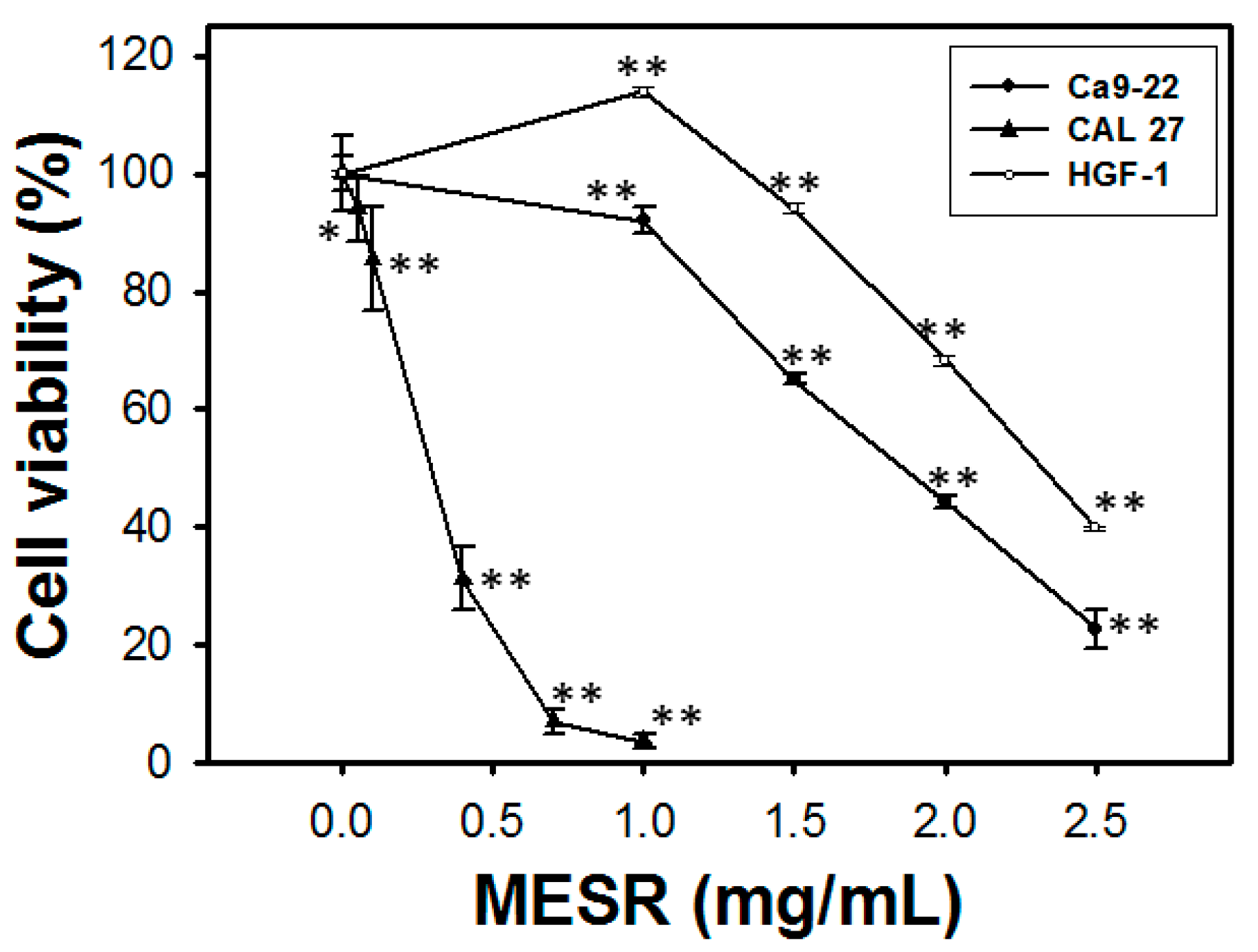
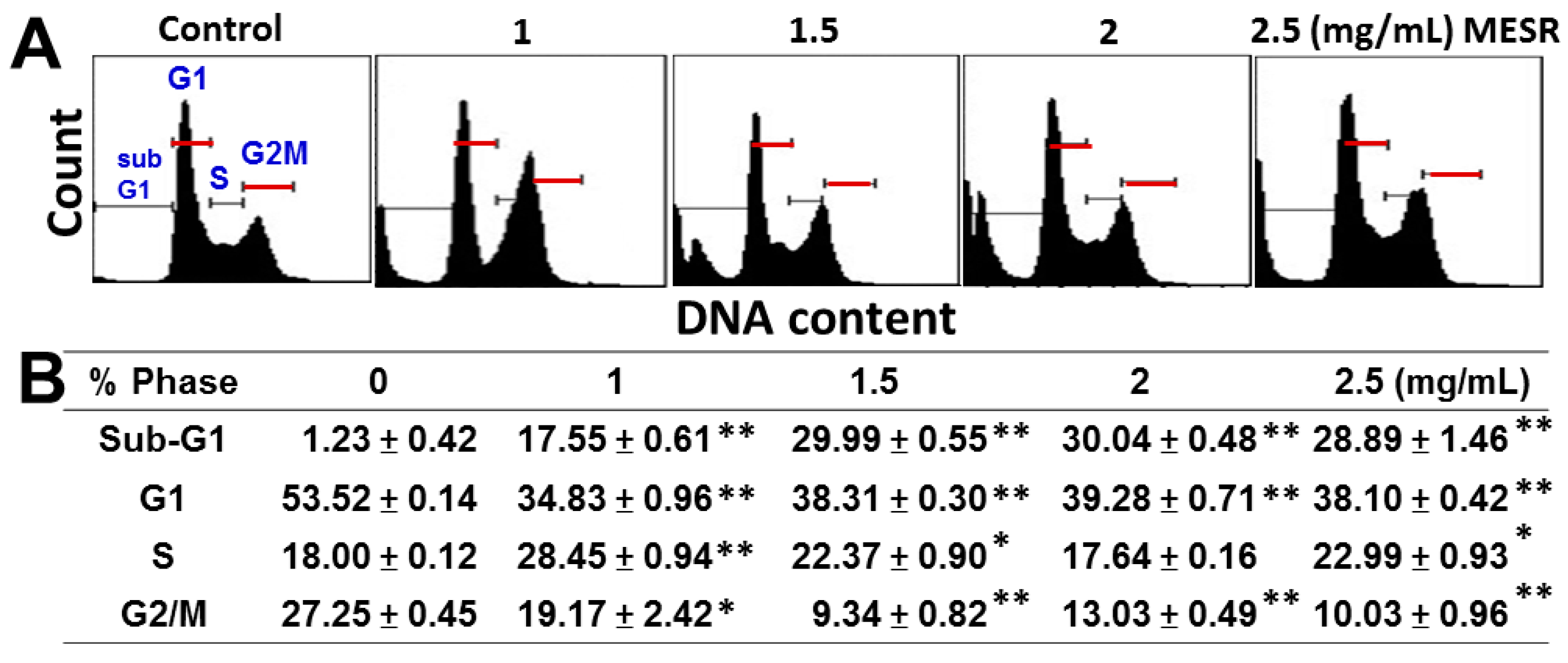
2.2. Cell Cycle Distribution of MESR-Incubated Ca9-22 Cells

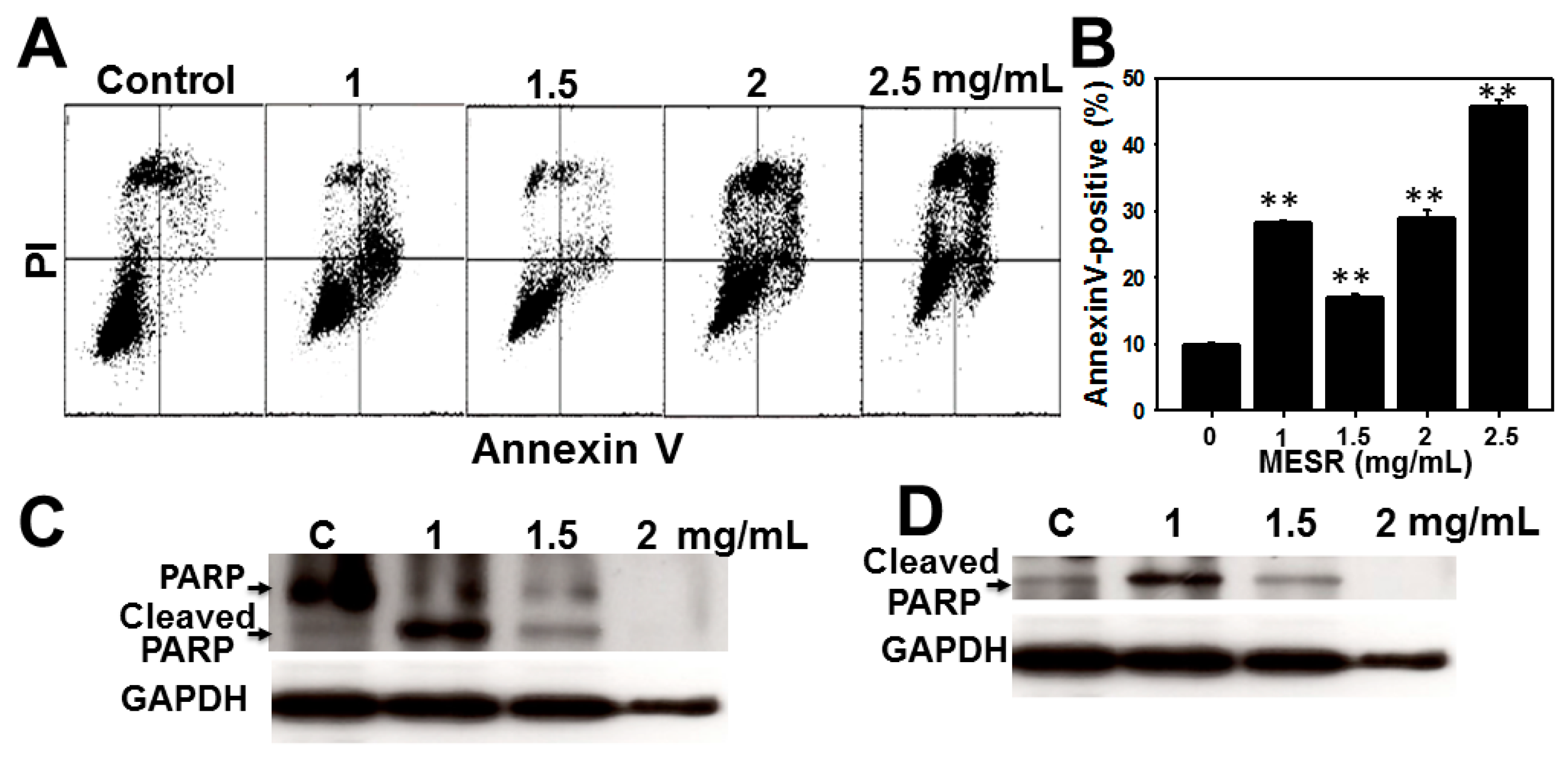
2.3. Apoptosis Analysis of MESR-Incubated Ca9-22 Cells

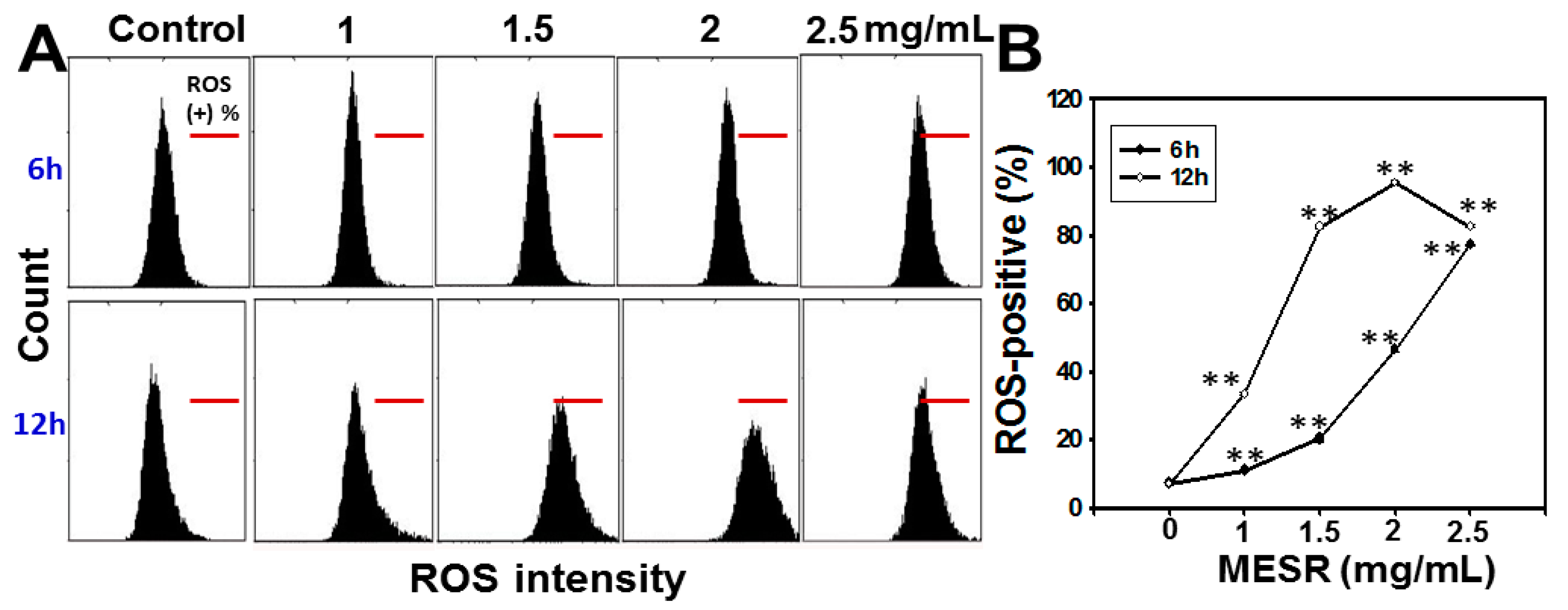
2.4. ROS Changes of MESR-Incubated Ca9-22 Cells

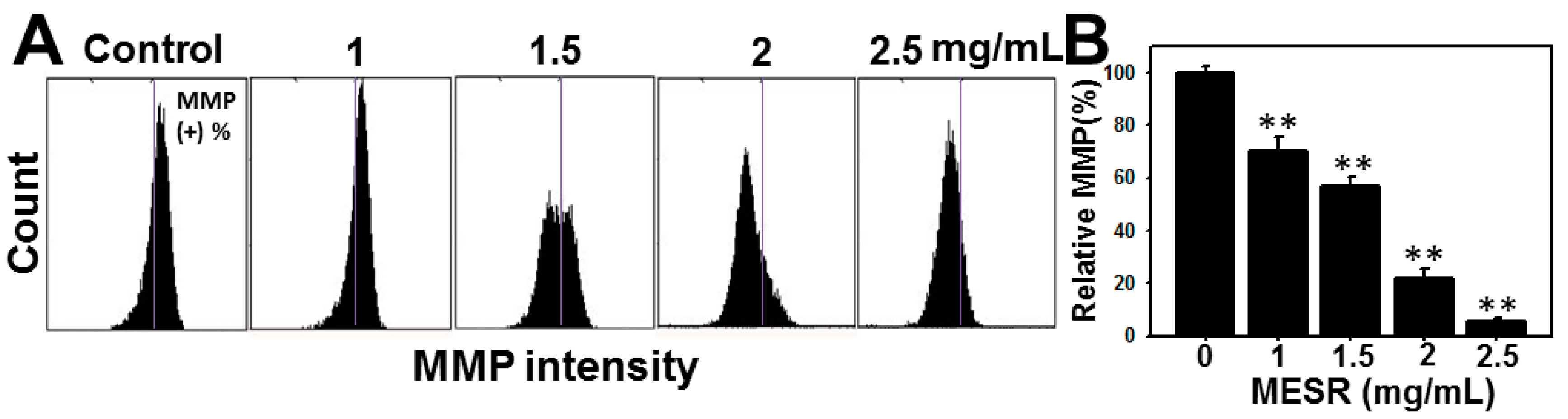
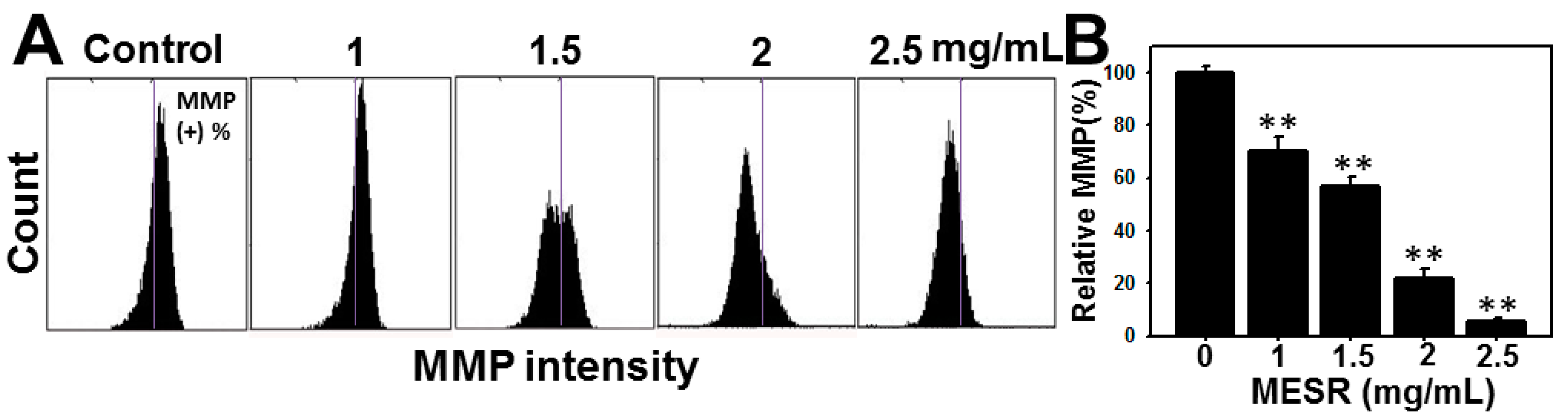
2.5. MMP Changes of MESR-Incubated Ca9-22 Cells
2.6. Discussion
3. Experimental Section
3.1. Cell Cultures and Methanolic Extracts of S. robusta
3.2. Cell Viability
3.3. Cell Cycle Distribution
3.4. Apoptosis Analysis
3.5. Intracellular ROS Level
3.6. Mitochondrial Membrane Potential (MMP)
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yen, C.Y.; Chen, C.H.; Chang, C.H.; Tseng, H.F.; Liu, S.Y.; Chuang, L.Y.; Wen, C.H.; Chang, H.W. Matrix metalloproteinases (MMP) 1 and MMP10 but not MMP12 are potential oral cancer markers. Biomarkers 2009, 14, 244–249. [Google Scholar] [CrossRef]
- Yen, C.Y.; Huang, C.Y.; Hou, M.F.; Yang, Y.H.; Chang, C.H.; Huang, H.W.; Chen, C.H.; Chang, H.W. Evaluating the performance of fibronectin 1 (FN1), integrin alpha4beta1 (ITGA4), syndecan-2 (SDC2), and glycoprotein CD44 as the potential biomarkers of oral squamous cell carcinoma (OSCC). Biomarkers 2013, 18, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Okui, T.; Shimo, T.; Fukazawa, T.; Hassan, N.M.; Honami, T.; Ibaragi, S.; Takaoka, M.; Naomoto, Y.; Sasaki, A. Novel HSP90 inhibitor NVP-AUY922 enhances the anti-tumor effect of temsirolimus against oral squamous cell carcinoma. Curr. Cancer Drug Targets 2012, 13, 289–299. [Google Scholar] [CrossRef]
- Hu, Y.; McIntosh, G.H.; le Leu, R.K.; Nyskohus, L.S.; Woodman, R.J.; Young, G.P. Combination of selenium and green tea improves the efficacy of chemoprevention in a rat colorectal cancer model by modulating genetic and epigenetic biomarkers. PLoS One 2013, 8, e64362. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Lim, C.W.; Kim, Y.K.; Yoon, H.D.; Lee, K.J. Immunostimulating and anticancer activities of hot water extract from Capsosiphon fulvescens. J. Kor. Soc. Appl. Biol. Chem. 2006, 49, 343–348. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Telles, C.B.; Dantas-Santos, N.; Camara, R.B.; Cordeiro, S.L.; Costa, M.S.; Almeida-Lima, J.; Melo-Silveira, R.F.; Oliveira, R.M.; et al. Antioxidant and antiproliferative activities of heterofucans from the seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952–966. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Bhadouria, P.; Bisen, P.S. Nutritional and therapeutic potential of Spirulina. Curr. Pharm. Biotechnol. 2005, 6, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Ahmadzadeh, S.; Tajbakhsh, S.; Rastian, Z.; Yousefi, F.; Farshadpour, F.; Sartavi, K. Anticancer activity of Sargassum oligocystum water extract against human cancer cell lines. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 669–673. [Google Scholar] [PubMed]
- Renju, G.L.; Muraleedhara Kurup, G.; Bandugula, V.R. Effect of lycopene isolated from Chlorella marina on proliferation and apoptosis in human prostate cancer cell line PC-3. Tumour Biol. 2014. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamad, R.; Baharara, J.; Zafar-Balanejad, S.; Fargahi, F.; Rahman, H.S. Antioxidant, antiproliferative, and antiangiogenesis effects of polyphenol-rich seaweed (Sargassum muticum). Biomed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Konickova, R.; Vankova, K.; Vanikova, J.; Vanova, K.; Muchova, L.; Subhanova, I.; Zadinova, M.; Zelenka, J.; Dvorak, A.; Kolar, M.; et al. Anti-cancer effects of blue-green alga Spirulina platensis, a natural source of bilirubin-like tetrapyrrolic compounds. Ann. Hepatol. 2014, 13, 273–283. [Google Scholar]
- Yeh, C.C.; Tseng, C.N.; Yang, J.I.; Huang, H.W.; Fang, Y.; Tang, J.Y.; Chang, F.R.; Chang, H.W. Antiproliferation and induction of apoptosis in Ca9-22 oral cancer cells by ethanolic extract of Gracilaria tenuistipitata. Molecules 2012, 17, 10916–10927. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.C.; Yang, J.I.; Lee, J.C.; Tseng, C.N.; Chan, Y.C.; Hseu, Y.C.; Tang, J.Y.; Chuang, L.Y.; Huang, H.W.; Chang, F.R.; et al. Anti-proliferative effect of methanolic extract of Gracilaria tenuistipitata on oral cancer cells involves apoptosis, DNA damage, and oxidative stress. BMC Complement. Altern. Med. 2012, 12, 142. [Google Scholar]
- Kim, J.Y.; Yoon, M.Y.; Cha, M.R.; Hwang, J.H.; Park, E.; Choi, S.U.; Park, H.R.; Hwang, Y.I. Methanolic extracts of Plocamium telfairiae induce cytotoxicity and caspase-dependent apoptosis in HT-29 human colon carcinoma cells. J. Med. Food 2007, 10, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Ara, J.; Sultana, V.; Qasim, R.; Ahmad, V.U. Hypolipidaemic activity of seaweed from Karachi coast. Phytother. Res. 2002, 16, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Barrett, T.; Benson, D.A.; Bryant, S.H.; Canese, K.; Chetvernin, V.; Church, D.M.; DiCuccio, M.; Edgar, R.; Federhen, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2009, 37, D5–D15. [Google Scholar]
- Kamenarska, Z.; Taniguchi, T.; Ohsawa, N.; Hiraoka, M.; Itoh, N. A vanadium-dependent bromoperoxidase in the marine red alga Kappaphycus alvarezii (Doty) Doty displays clear substrate specificity. Phytochemistry 2007, 68, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Pooja, S. Algae used as medicine and food-a short review. J. Pharm. Sci. Res. 2014, 6, 33–35. [Google Scholar]
- O’Brien, M.A.; Moravec, R.A.; Riss, T.L. Poly (ADP-ribose) polymerase cleavage monitored in situ in apoptotic cells. Biotechniques 2001, 30, 886–891. [Google Scholar] [PubMed]
- Zakaria, N.A.; Ibrahim, D.; Sulaiman, S.F.; Supardy, N.A. Assessment of antioxidant activity, total phenol content and in vitro toxicity of Malaysian red seaweed. J. Chem. Pharm. Res. 2011, 3, 182–191. [Google Scholar]
- Wang, T.; Jónsdóttir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extract from Icelandic seaweed. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Mukhtar, H. Cancer chemoprevention through dietary antioxidants: Progress and promise. Antioxid. Redox Signal. 2008, 10, 475–510. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.I.; Yeh, C.C.; Lee, J.C.; Yi, S.C.; Huang, H.W.; Tseng, C.N.; Chang, H.W. Aqueous extracts of the edible Gracilaria tenuistipitata are protective against H2O2-induced DNA damage, growth inhibition, and cell cycle arrest. Molecules 2012, 17, 7241–7254. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.Y.; Ou, N.; Lu, Q.B. Antioxidant induces DNA damage, cell death and mutagenicity in human lung and skin normal cells. Sci. Rep. 2013, 3, 3169. [Google Scholar] [PubMed]
- Fox, J.T.; Sakamuru, S.; Huang, R.; Teneva, N.; Simmons, S.O.; Xia, M.; Tice, R.R.; Austin, C.P.; Myung, K. High-throughput genotoxicity assay identifies antioxidants as inducers of DNA damage response and cell death. Proc. Natl. Acad. Sci. USA 2012, 109, 5423–5428. [Google Scholar] [CrossRef] [PubMed]
- Dellai, A.; Deghrigue, M.; Bouraoui, A. Evaluation of the antiproliferative activity of methanol extract and its fractions from the mediterranean seaweed, Halurus equisetifolius. Int. J. Pharm. Pharm. Sci. 2013, 5, 148–152. [Google Scholar]
- Yang, Y.; Zhang, Z.; Li, S.; Ye, X.; Li, X.; He, K. Synergy effects of herb extracts: Pharmacokinetics and pharmacodynamic basis. Fitoterapia 2014, 92, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H. Synergy research: Approaching a new generation of phytopharmaceuticals. Fitoterapia 2011, 82, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nat. Rev. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012. [Google Scholar] [CrossRef]
- Kurt, O.; Ozdal-Kurt, F.; Tuglu, M.; Akcora, C. The cytotoxic, neurotoxic, apoptotic and antiproliferative activities of extracts of some marine algae on the MCF-7 cell line. Biotech. Histochem 2014, 89, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Kajstura, M.; Halicka, H.D.; Pryjma, J.; Darzynkiewicz, Z. Discontinuous fragmentation of nuclear DNA during apoptosis revealed by discrete “sub-G1” peaks on DNA content histograms. Cytometry A 2007, 71, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Haung, J.W.; Chang, F.R.; Huang, K.J.; Huang, H.M.; Huang, H.W.; Chou, C.K.; Wu, Y.C.; Chang, H.W. Golden berry-derived 4beta-hydroxywithanolide E for selectively killing oral cancer cells by generating ROS, DNA damage, and apoptotic pathways. PLoS One 2013, 8, e64739. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Chang, H.W.; Huang, H.M.; Chong, I.W.; Chen, J.S.; Chen, C.Y.; Wang, H.M. (−)-Anonaine induces DNA damage and inhibits growth and migration of human lung carcinoma h1299 cells. J. Agric. Food Chem. 2011, 59, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Liu, P.L.; Huang, K.J.; Wang, H.M.; Chang, K.F.; Chou, C.K.; Chang, F.R.; Chong, I.W.; Fang, K.; Chen, J.S.; et al. Goniothalamin inhibits growth of human lung cancer cells through DNA damage, apoptosis, and reduced migration ability. J. Agric. Food Chem. 2011, 59, 4288–4293. [Google Scholar]
- Huang, H.W.; Chung, Y.A.; Chang, H.S.; Tang, J.Y.; Chen, I.S.; Chang, H.W. Antiproliferative effects of methanolic extracts of Cryptocarya concinna Hance roots on oral cancer Ca9-22 and CAL 27 cell lines involving apoptosis, ROS induction, and mitochondrial depolarization. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef]
- Yen, C.Y.; Chiu, C.C.; Haung, R.W.; Yeh, C.C.; Huang, K.J.; Chang, K.F.; Hseu, Y.C.; Chang, F.R.; Chang, H.W.; Wu, Y.C. Antiproliferative effects of goniothalamin on Ca9-22 oral cancer cells through apoptosis; DNA damage and ROS induction. Mutat. Res. 2012, 747, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, Y.-H.; Farooqi, A.A.; Li, K.-T.; Butt, G.; Tang, J.-Y.; Wu, C.-Y.; Cheng, Y.-B.; Hou, M.-F.; Chang, H.-W. Methanolic Extracts of Solieria robusta Inhibits Proliferation of Oral Cancer Ca9-22 Cells via Apoptosis and Oxidative Stress. Molecules 2014, 19, 18721-18732. https://doi.org/10.3390/molecules191118721
Yen Y-H, Farooqi AA, Li K-T, Butt G, Tang J-Y, Wu C-Y, Cheng Y-B, Hou M-F, Chang H-W. Methanolic Extracts of Solieria robusta Inhibits Proliferation of Oral Cancer Ca9-22 Cells via Apoptosis and Oxidative Stress. Molecules. 2014; 19(11):18721-18732. https://doi.org/10.3390/molecules191118721
Chicago/Turabian StyleYen, Yii-Huei, Ammad Ahmad Farooqi, Kun-Tzu Li, Ghazala Butt, Jen-Yang Tang, Chang-Yi Wu, Yuan-Bin Cheng, Ming-Feng Hou, and Hsueh-Wei Chang. 2014. "Methanolic Extracts of Solieria robusta Inhibits Proliferation of Oral Cancer Ca9-22 Cells via Apoptosis and Oxidative Stress" Molecules 19, no. 11: 18721-18732. https://doi.org/10.3390/molecules191118721