2.1. Effect of Exogenous Spermidine on Seed Germination Characteristics
With the increase of PEG concentration, germination percentage, germination vigor and germination index gradually declined and mean germination time gradually increased in seeds primed with spermidine (Spd) or water, but exogenous Spd apparently improved white clover seed germination and shortened the mean germination time of seeds under water stress (
Table 1). Seeds primed with Spd were able to maintain a higher germination percentage, vigor and index than seeds primed with water for the same PEG concentration. When the PEG concentration was 15%, seeds primed with Spd had 8.0%, 8.7% and 19.5% higher germination percentage, vigor and index than seeds primed with water, respectively. Mean germination time in seeds primed with Spd was shortened from 1.11 to 1.04 days and from 1.54 to 1.33 days under 0% and 15% PEG conditions, as compared with seeds primed with water, respectively (
Table 1).
Table 1.
The effect of seed priming with water or spermidine (Spd) on seed germination characteristics in white clover under 7 days of different water stress conditions. Values are mean ± SE (n = 6). Different letters in a vertical column indicate a significant difference between each treatment under different PEG concentration. The asterisk indicates a significant difference exists between seed priming with water or Spd. LSD (p ≤ 0.05).
Table 1.
The effect of seed priming with water or spermidine (Spd) on seed germination characteristics in white clover under 7 days of different water stress conditions. Values are mean ± SE (n = 6). Different letters in a vertical column indicate a significant difference between each treatment under different PEG concentration. The asterisk indicates a significant difference exists between seed priming with water or Spd. LSD (p ≤ 0.05).
| PEG (%) | Germination Percentage (%) | Germination Vigor (%) | Germination Index | Mean Germination Time (d) |
|---|
| Water | Spd | Water | Spd | Water | Spd | Water | Spd |
|---|
| 0 | 95.5 ± 2.5 a | 97.5 ± 1.0 a | 95.5 ± 2.5 a | 97.5 ± 5.1 a | 45.38 ± 1.65 a | 47.88 ± 0.48 a * | 1.11 ± 0.05 d | 1.04 ± 0.03 d * |
| 10 | 92.0 ± 1.6 a | 96.5 ± 1.0 a * | 87.3 ± 8.3 a | 94.7 ± 2.3 a | 39.62 ± 4.15 b | 43.56 ± 1.80 b | 1.41 ± 0.32 c | 1.27 ± 0.12 c |
| 15 | 78.7 ± 3.1 b | 86.7 ± 3.6 b * | 74.7 ± 3.1 b | 83.3 ± 5.0 b * | 31.93 ± 2.20 c | 38.16 ± 1.97 c * | 1.54 ± 0.11 b c | 1.33 ± 0.04 c * |
| 18 | 54.7 ± 8.1 c | 59.5 ± 4.4 c | 53.3 ± 4.1 c | 58.7 ± 4.2 c | 16.53 ± 0.55 d | 21.98 ± 1.21 d * | 1.72 ± 0.18 b a | 1.70 ± 0.11 b |
| 20 | 23.5 ± 6.6 d | 26.0 ± 7.1 d | 23.0 ± 7.0 d | 25.0 ± 8.2 d | 6.36 ± 1.87 e | 6.86 ± 1.90 e | 1.89 ± 0.10 a | 1.90 ± 0.12 a |
Under different water stress conditions (0% to 20% PEG), seedling fresh weight in both treatments declined significantly and exogenous Spd significantly increased seedling fresh weight when the PEG concentration was 10% (
Table 2). In addition, exogenous Spd treatment apparently simulated growth of roots, and led to a significant increase in seedlings dry weight under water stress. Seedlings dry weight of exogenous Spd treatment increased by about 38% and 19% over non-Spd treatment at 10% and 15% PEG, respectively. A significantly higher seeds vigour index also was observed in seeds priming with Spd than that in seeds priming with water at 15% PEG stress (
Table 2).
Table 2.
The effect of seed priming with water or spermidine (Spd) on fresh weight, dry weight, root length and seed vigour index in seeds of white clover after 7 days of germination under different water stress. Values are mean ± SE (n = 6). Different letters in a vertical column indicate a significant difference in each treatment under different PEG concentration conditions. The asterisk indicates a significant difference exists between seed priming with water or Spd. LSD (p ≤ 0.05).
Table 2.
The effect of seed priming with water or spermidine (Spd) on fresh weight, dry weight, root length and seed vigour index in seeds of white clover after 7 days of germination under different water stress. Values are mean ± SE (n = 6). Different letters in a vertical column indicate a significant difference in each treatment under different PEG concentration conditions. The asterisk indicates a significant difference exists between seed priming with water or Spd. LSD (p ≤ 0.05).
| PEG (%) | Seedling Fresh Weight (mg·10 Seedling−1) | Seedling Dry Weight (mg·10 Seedling−1) | Root Length (cm) | Seed Vigour Index |
|---|
| Water | Spd | Water | Spd | Water | Spd | Water | Spd |
|---|
| 0 | 86.7 ± 4.8 a | 86.1 ± 4.4 a | 5.5 ± 0.4 b | 5.6 ± 0.07 c | 2.97 ± 0.15 b | 3.46 ± 0.15 a * | 4.02 ± 0.48 a | 4.01 ± 0.17 a |
| 10 | 55.3 ± 6.3 b | 65.1 ± 2.4 b * | 5.7 ± 0.6 b a | 8.0 ± 0.10 a * | 3.20 ± 0.03 a | 3.45 ± 0.21 a * | 2.26 ± 0.45 b | 2.84 ± 0.10 b |
| 15 | 47.4 ± 1.1 c | 50.7 ± 3.6 c | 6.0 ± 0.06 b a | 7.2 ± 0.03 b a * | 2.69 ± 0.10 c | 2.88 ± 0.12 b | 1.52 ± 0.14 c | 1.91 ± 0.09 c * |
| 18 | 37.4 ± 2.9 d | 40.5 ± 1.4 d | 5.9 ± 0.09 b a | 6.5 ± 0.10 b | 2.23 ± 0.22 d | 2.37 ± 0.08 c | 0.70 ± 0.18 d | 0.88 ± 0.01 d |
| 20 | 24.2 ± 4.5 e | 24.4 ± 1.2 e | 6.2 ± 0.02 a | 6.7 ± 0.03 b | 1.47 ± 0.05 e | 1.60 ± 0.14 d | 0.16 ± 0.07 e | 0.23 ± 0.02 e |
2.2. Effect of Exogenous Spermidine on Carbohydrate Levels
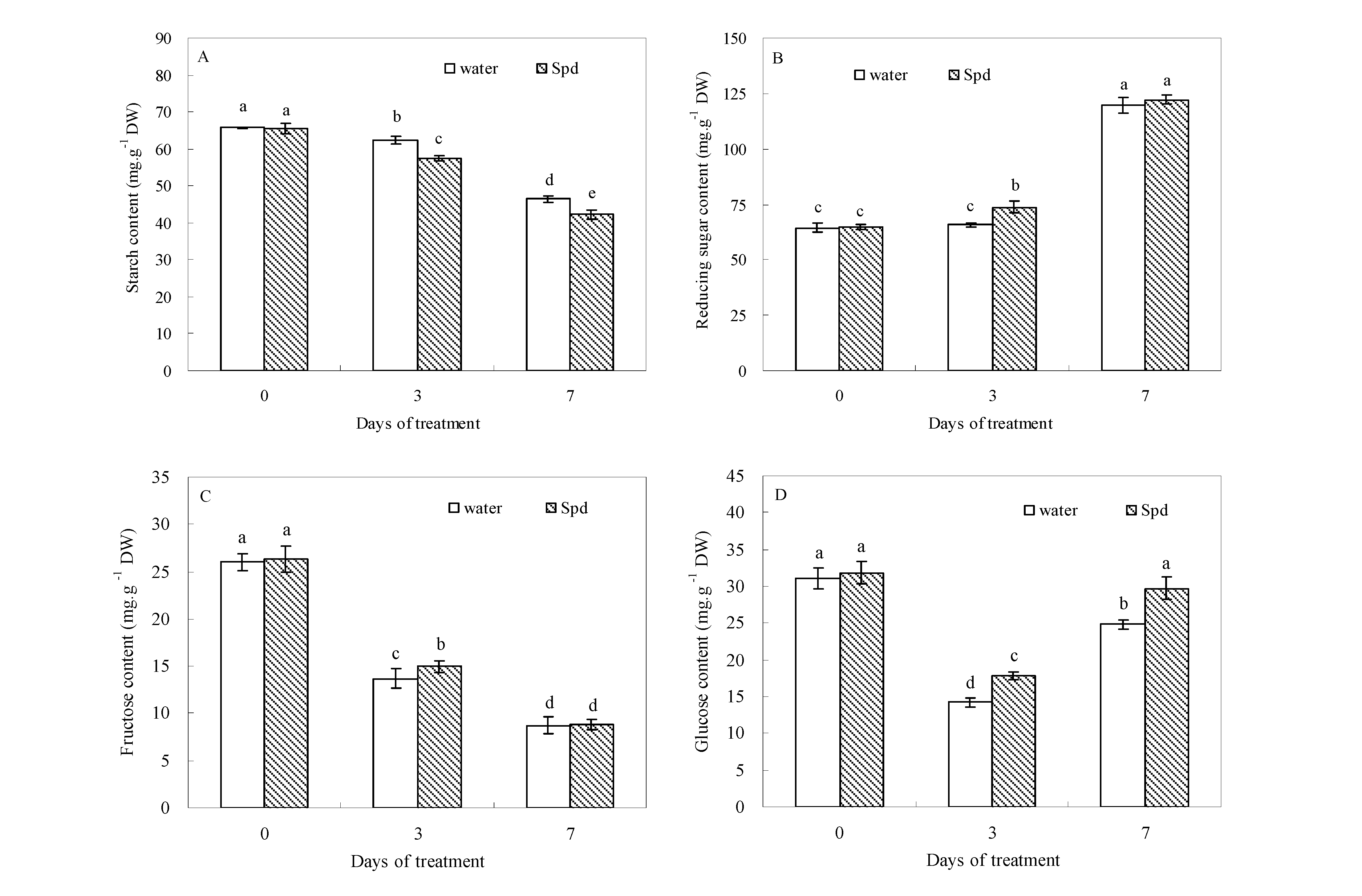
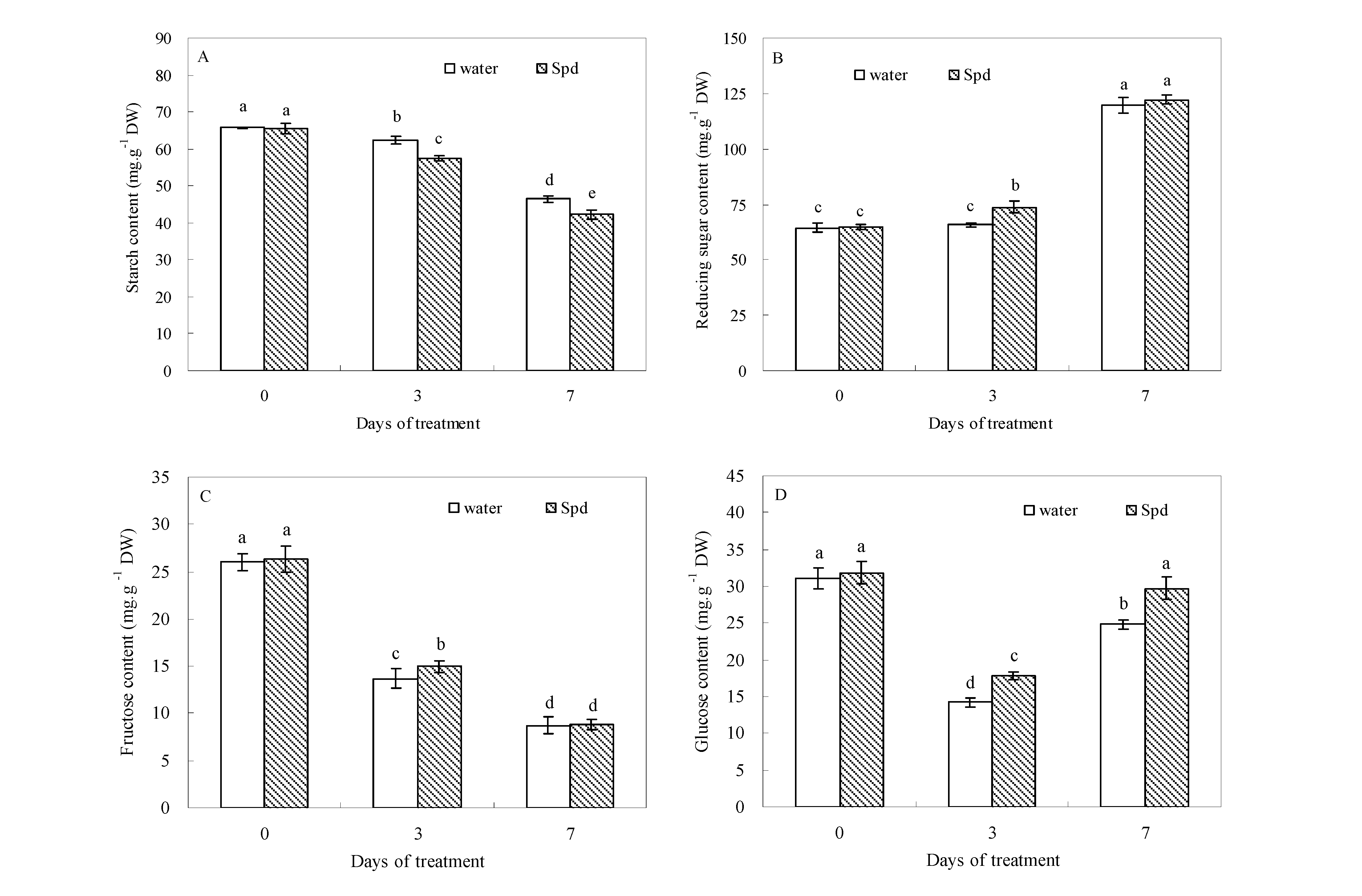
Four organic carbohydrates were detected in the seeds of both treatments. At the beginning of germination (0 day), the contents of starch, reducing sugar, fructose and glucose were not significantly different between the two treatments. Seed starch content decreased, whereas reducing sugar content increased in white clover during the process of germination (
Figure 1A,B).
Figure 1.
The effect of seed soaking with water or spermidine (Spd) on (A) starch content; (B) reducing sugar content; (C) fructose content and (D) glucose content in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 1.
The effect of seed soaking with water or spermidine (Spd) on (A) starch content; (B) reducing sugar content; (C) fructose content and (D) glucose content in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Under water stress, seeds primed with Spd showed further decreased seed starch content at 3 days of germination. In contrast, seeds primed with Spd showed a significantly increased reducing sugar content in germinating seeds. At 3 days of germination, reducing sugar content in exogenous Spd treatment was about 12.2% higher than that in the control (seeds primed with water,
Figure 1B). Fructose content began to decline from the beginning to the end of germination in both treatments, but compared to seeds primed with water, exogenous Spd significantly enhanced fructose accumulation after 3 days of germination (
Figure 1C). The difference in glucose content was significantly greater in seeds primed with Spd than primed with water under different germination time conditions. Accordingly, Spd treatment resulted in 18.6% and 16.5% higher glucose content than water treatment at 3 and 7 days of germination under water stress (
Figure 1D).
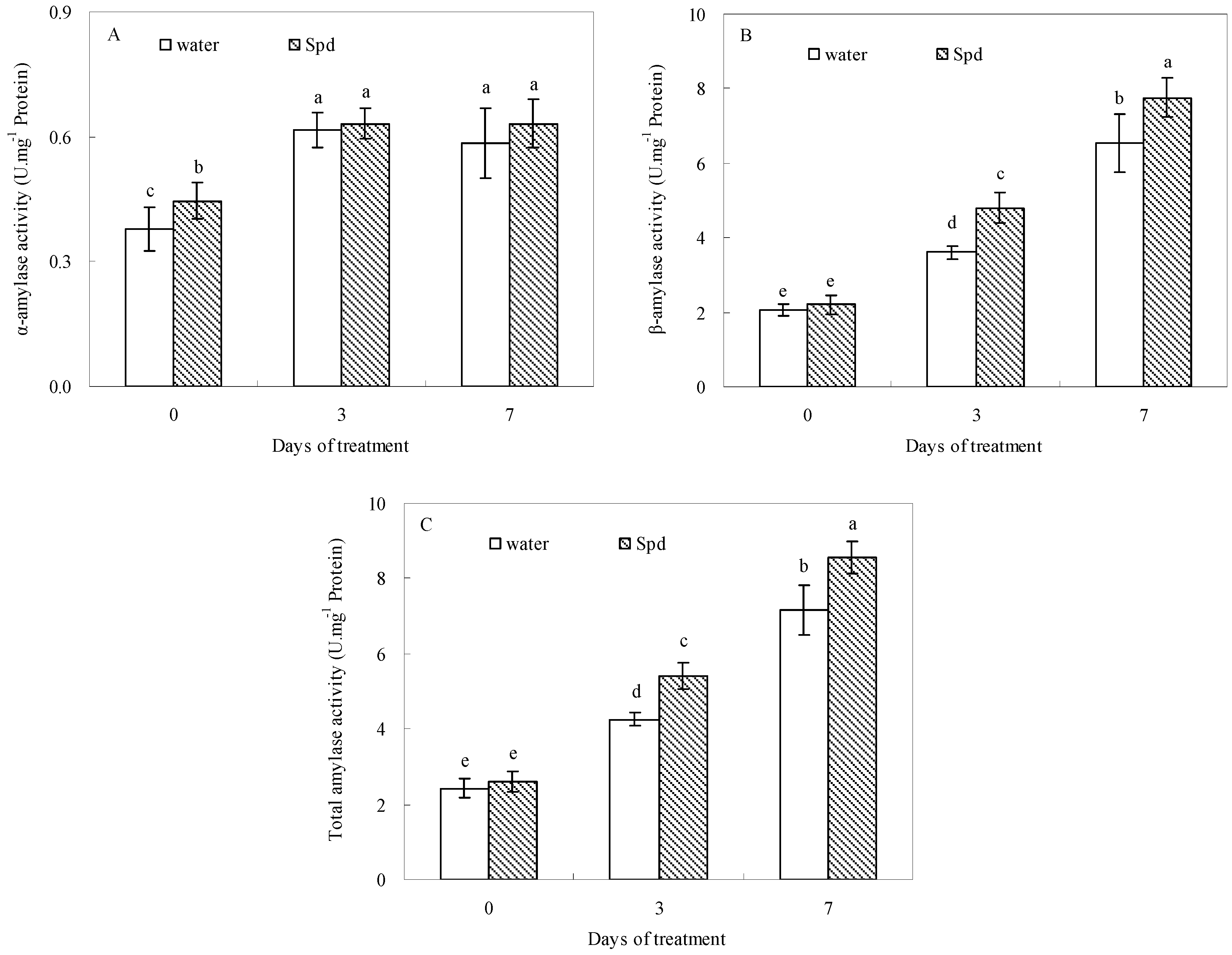
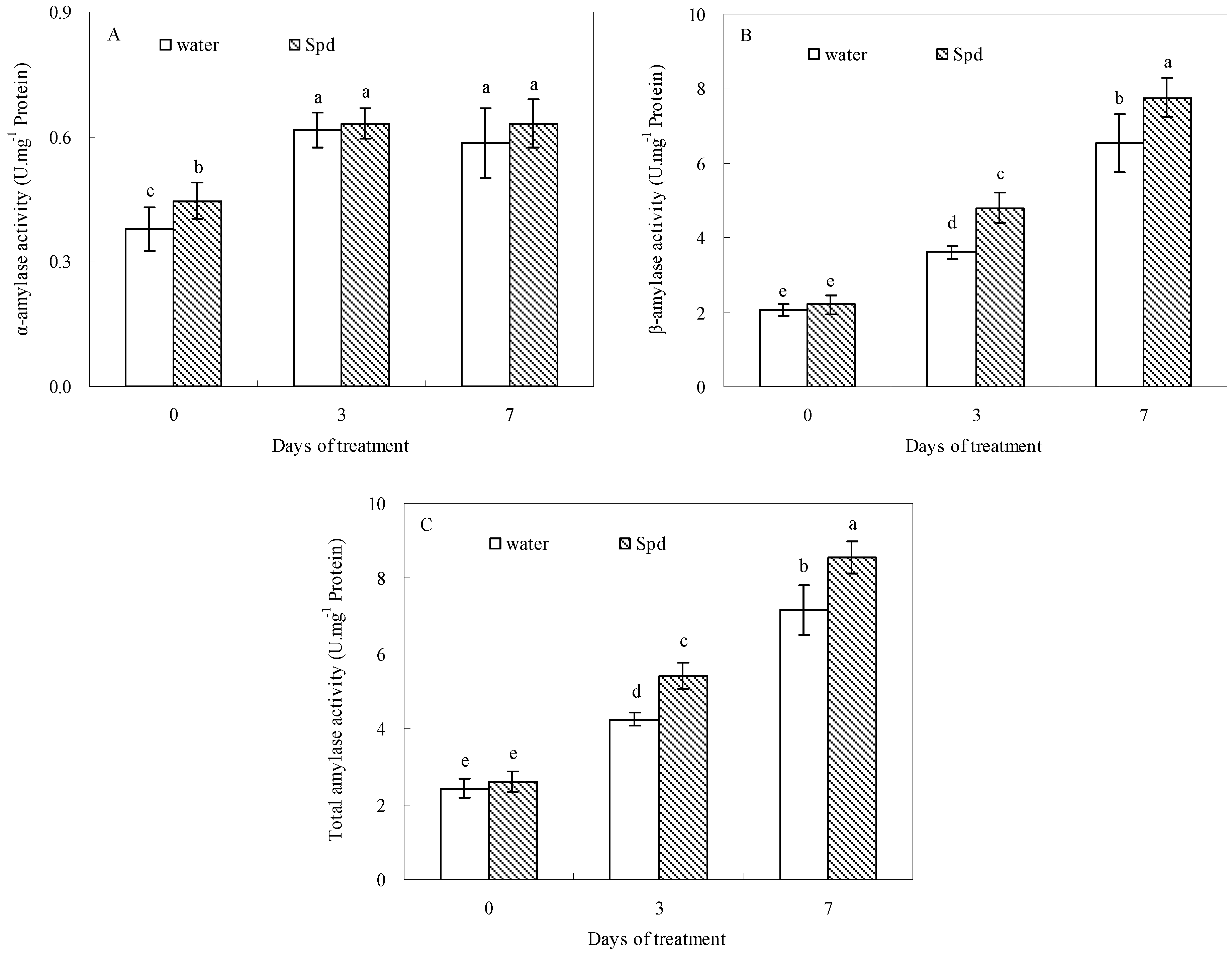
2.3. Effect of Exogenous Spermidine on Amylase Activities and Gene Relative Expression
Progressive germination induced a significant increase of α-amylase activities, so that α-amylase activities of both treatments reached their maximum after 3 d of water stress. Compared to control (seeds primed with water), exogenous Spd significantly enhanced α-amylase activities at the beginning of the germination progress (
Figure 2A). The β-amylase and total amylase activities were also significantly impacted by exogenous Spd, and accordingly a significantly higher β-amylase and total amylase activities in exogenous Spd treatment was observed than that in control at the same level of water stress and germination time (
Figure 2B,C).
Figure 2.
The effect of seed soaking with water or spermidine (Spd) on (A) α-amylase activity; (B) β-amylase activity and (C) total amylase activity in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 2.
The effect of seed soaking with water or spermidine (Spd) on (A) α-amylase activity; (B) β-amylase activity and (C) total amylase activity in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
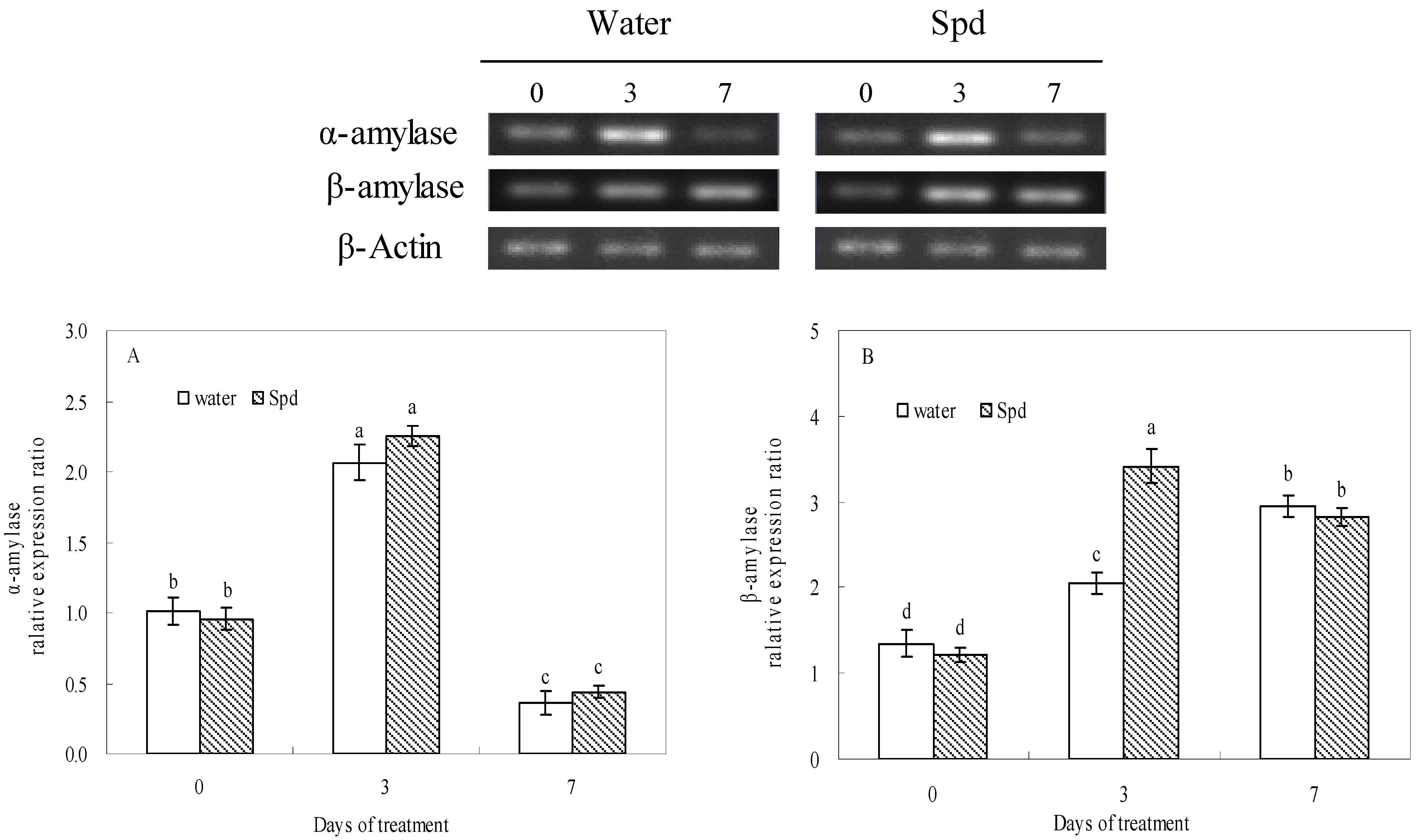
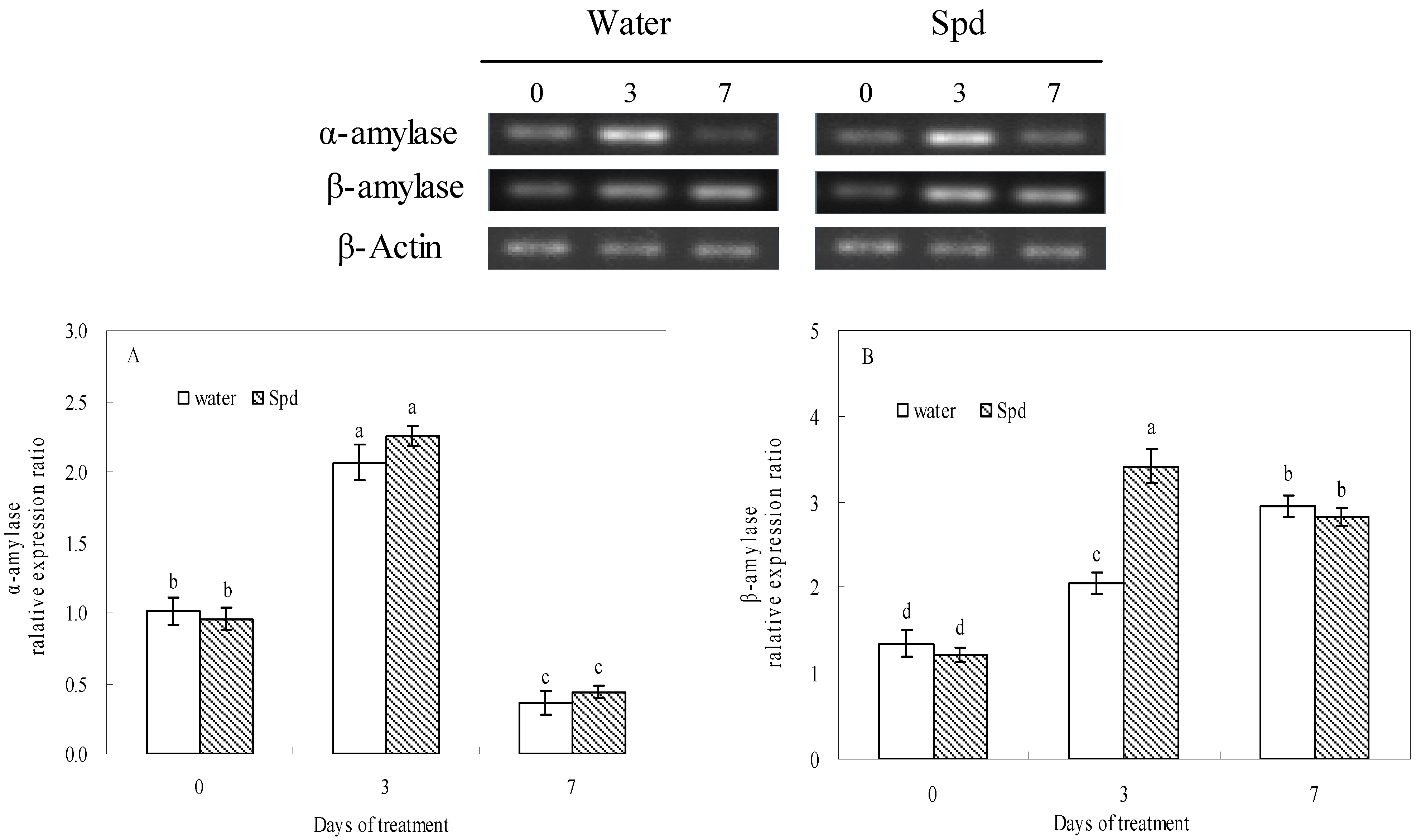
The transcript level of α-amylase gene increased rapidly at 3 days of germinating under water stress a thereafter decreased deeply in both treatments, but there was no significant difference between the two treatments (
Figure 3A). The transcript level of β-amylase gene gradually changed with the increase in germination time, and the transcript level of β-amylase gene of exogenous Spd treatment was 1.7 times as high as that of control at 3 d of germination, and the differences were statistically significant (
Figure 3B).
Figure 3.
The effect of seed soaking with water or spermidine (Spd) on (A) α-amylase gene and (B) β-amylase gene relative expression ratio in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 4). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 3.
The effect of seed soaking with water or spermidine (Spd) on (A) α-amylase gene and (B) β-amylase gene relative expression ratio in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 4). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
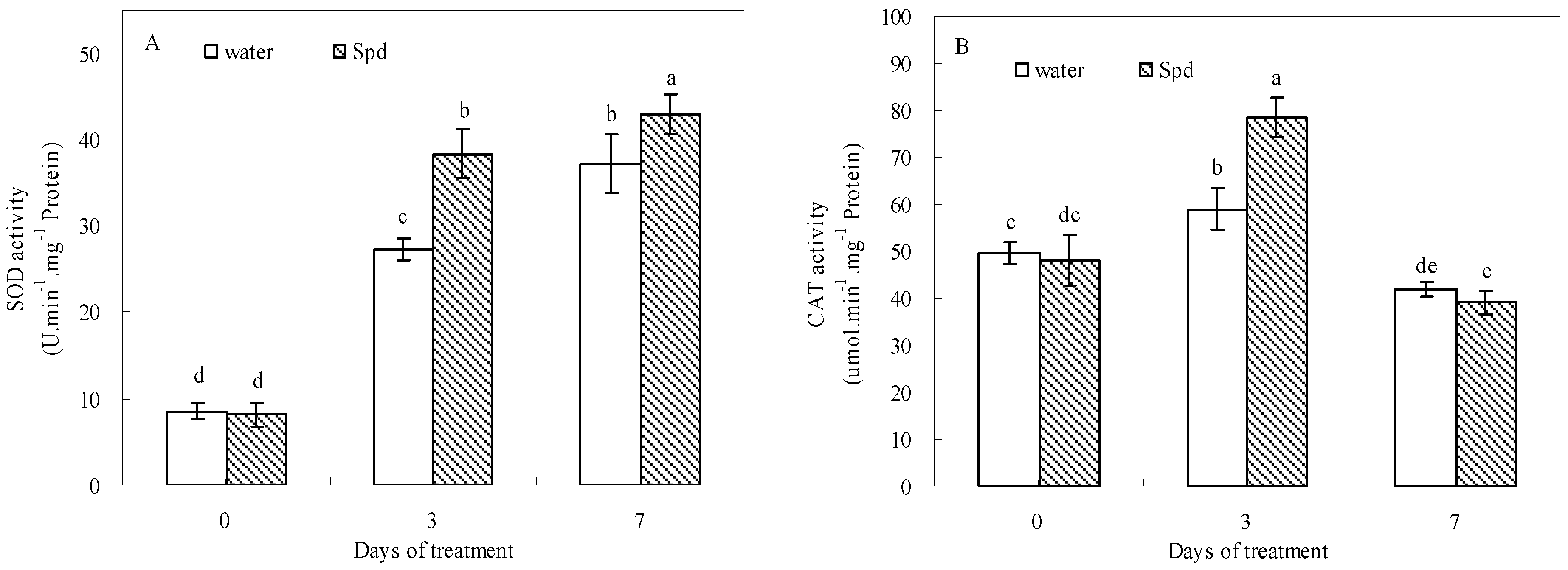
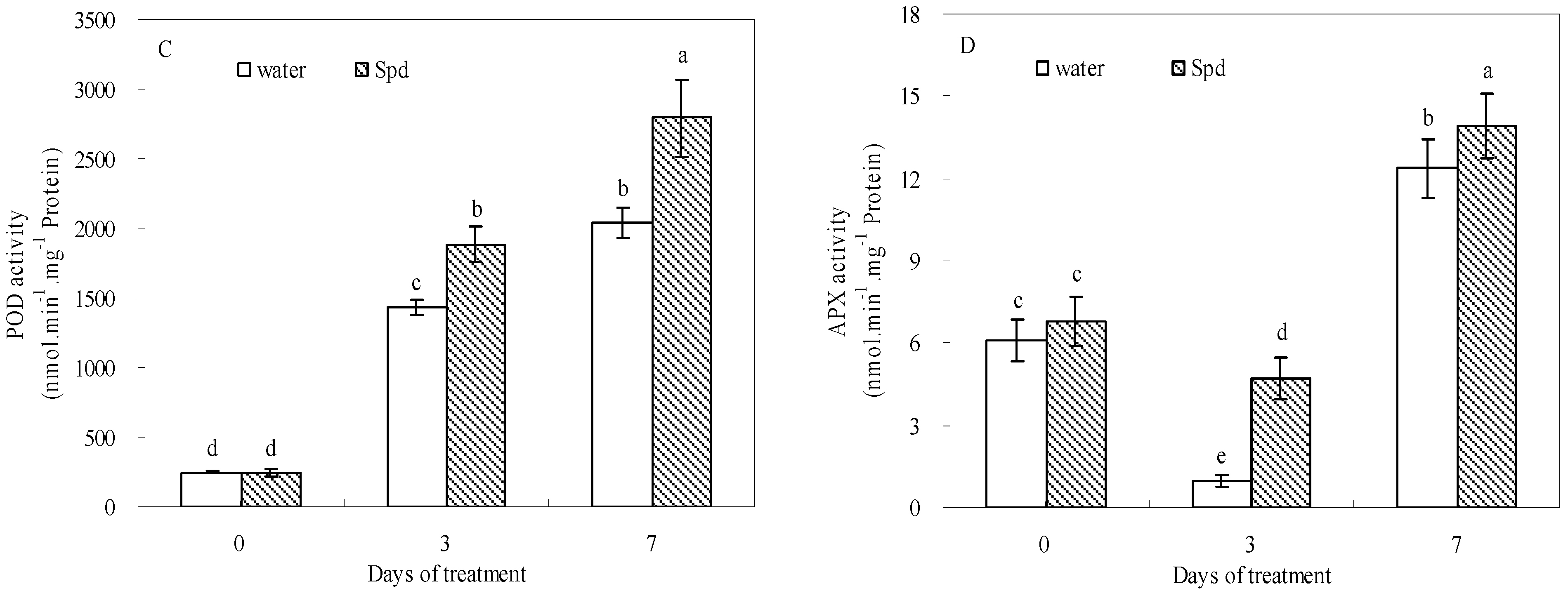
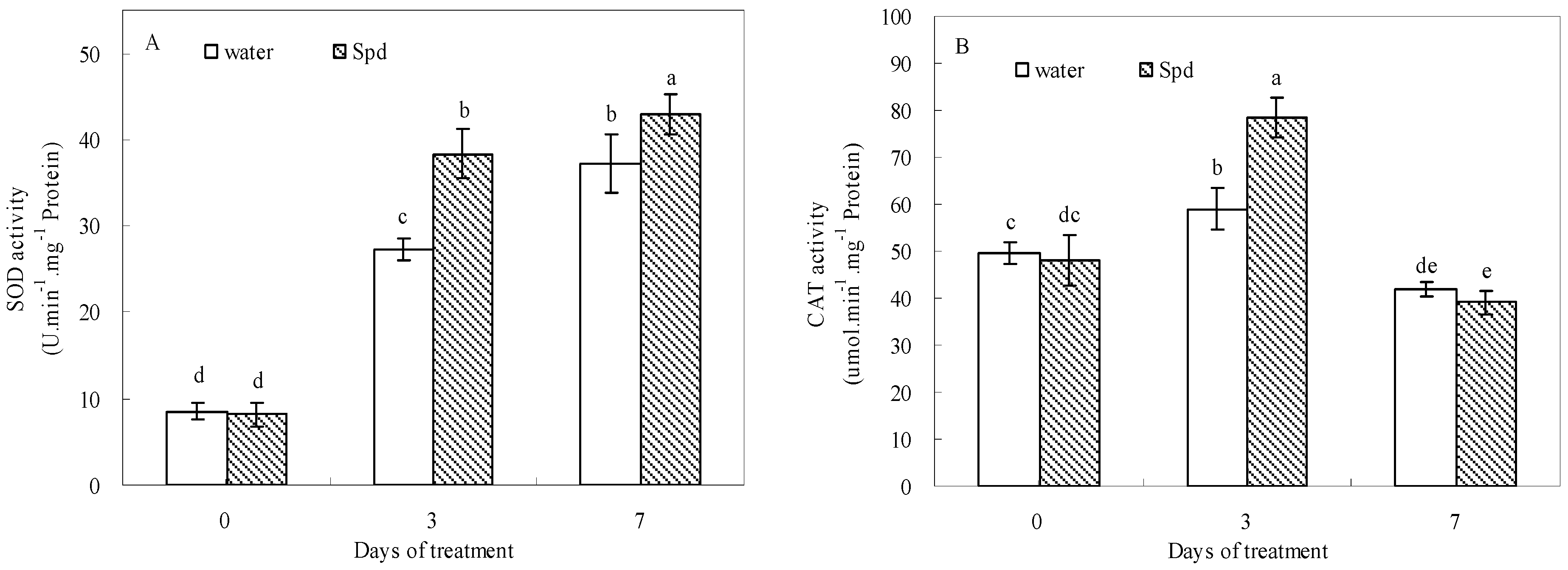
2.5. Effect of Exogenous Spermidine on Antioxidant Enzyme Activities and Gene Relative Expression
Exogenous Spd had stimulative effects on SOD, POD, CAT and APX activities during germination under water stress conditions (
Figure 5). In response to water stress, SOD and POD activities gradually increased and then reached a peak value at the last day of germination in both treatments. Moreover, SOD and POD activities after Spd treatment were significantly higher than those of control at 3 days and 7 days of germination (
Figure 5A,C).
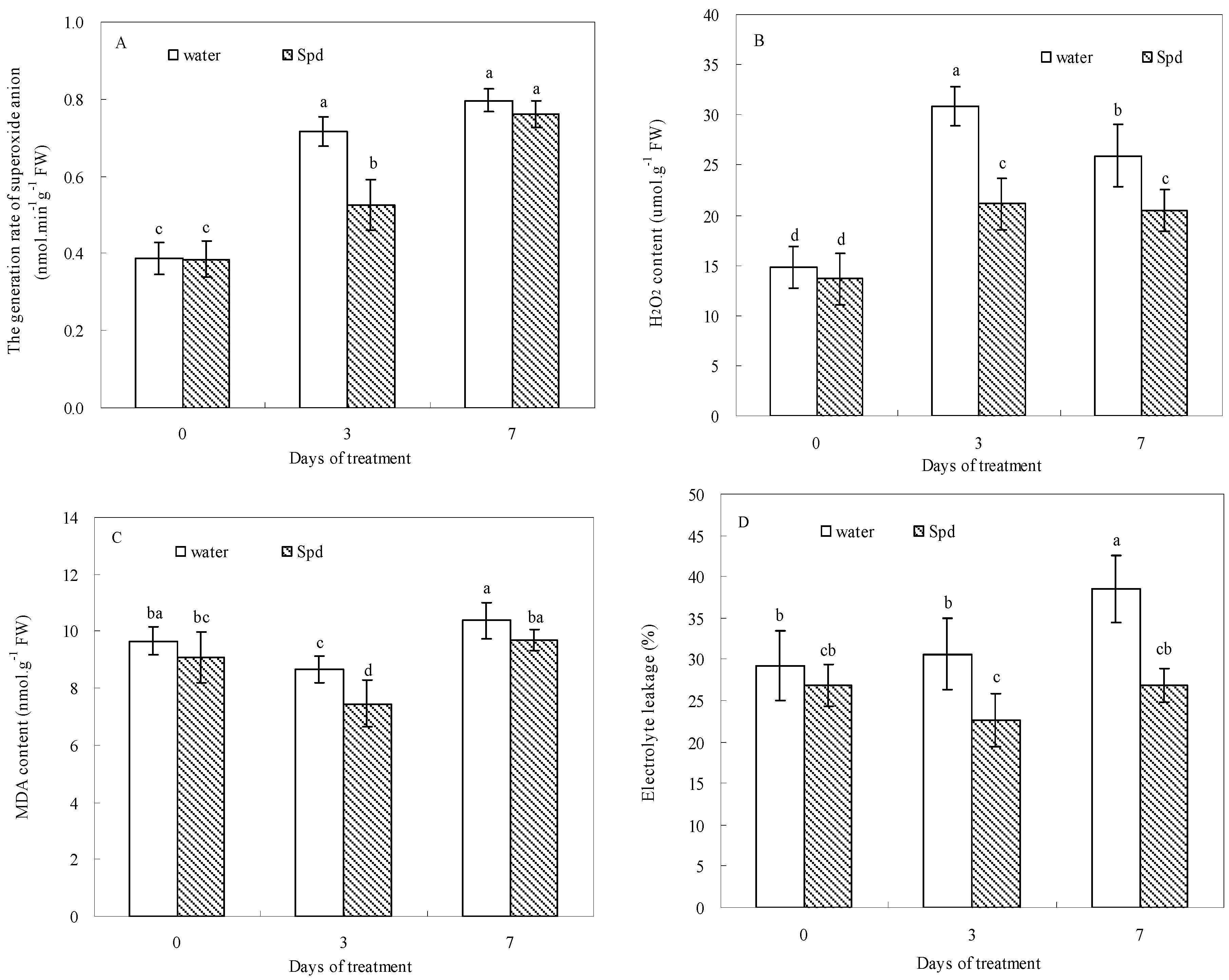
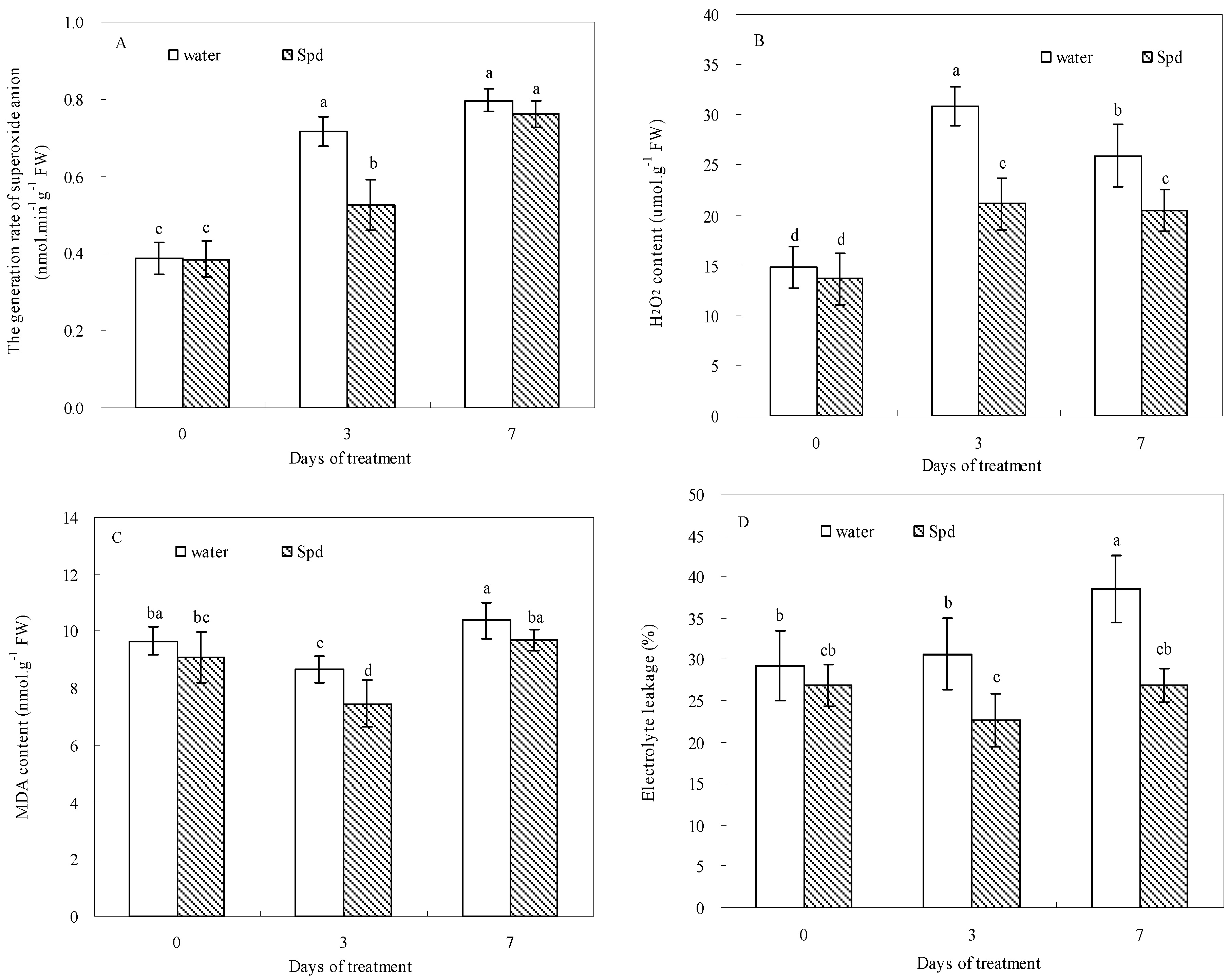
Figure 4.
The effect of seed soaking with water or spermidine (Spd) on (A) the generation rate of superoxide anion (O2−); (B) H2O2 content; (C) MDA content and (D) electrolyte leakage (EL) in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 4.
The effect of seed soaking with water or spermidine (Spd) on (A) the generation rate of superoxide anion (O2−); (B) H2O2 content; (C) MDA content and (D) electrolyte leakage (EL) in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 5.
The effect of seed soaking with water or spermidine (Spd) on (A) superoxide dismutase (SOD) activity; (B) catalase (CAT) activity; (C) guaiacol peroxidase (POD) activity and (D) ascorbate peroxidase (APX) activity in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 5.
The effect of seed soaking with water or spermidine (Spd) on (A) superoxide dismutase (SOD) activity; (B) catalase (CAT) activity; (C) guaiacol peroxidase (POD) activity and (D) ascorbate peroxidase (APX) activity in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 6). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
At the beginning of germination, CAT activities showed a gradual increase for both treatments and then started to decline following aggravating stress, but CAT activity was elevated significantly with Spd treatment as compared with that in control at 3 d (
Figure 5B). The APX activity of Spd treatment also was significantly higher than that of the control, and the difference was most pronounced at 3 d of germination under water stress (
Figure 5D).
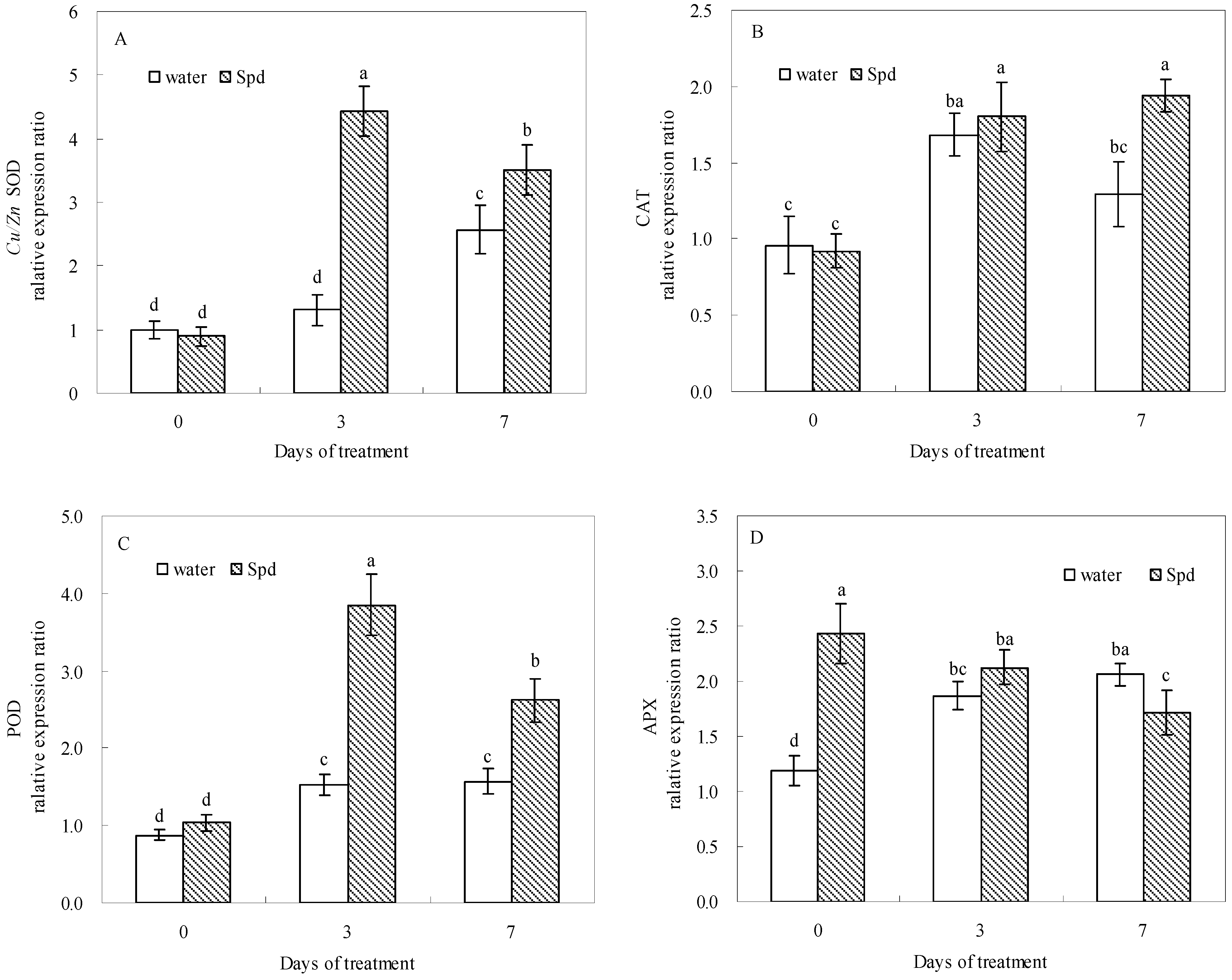
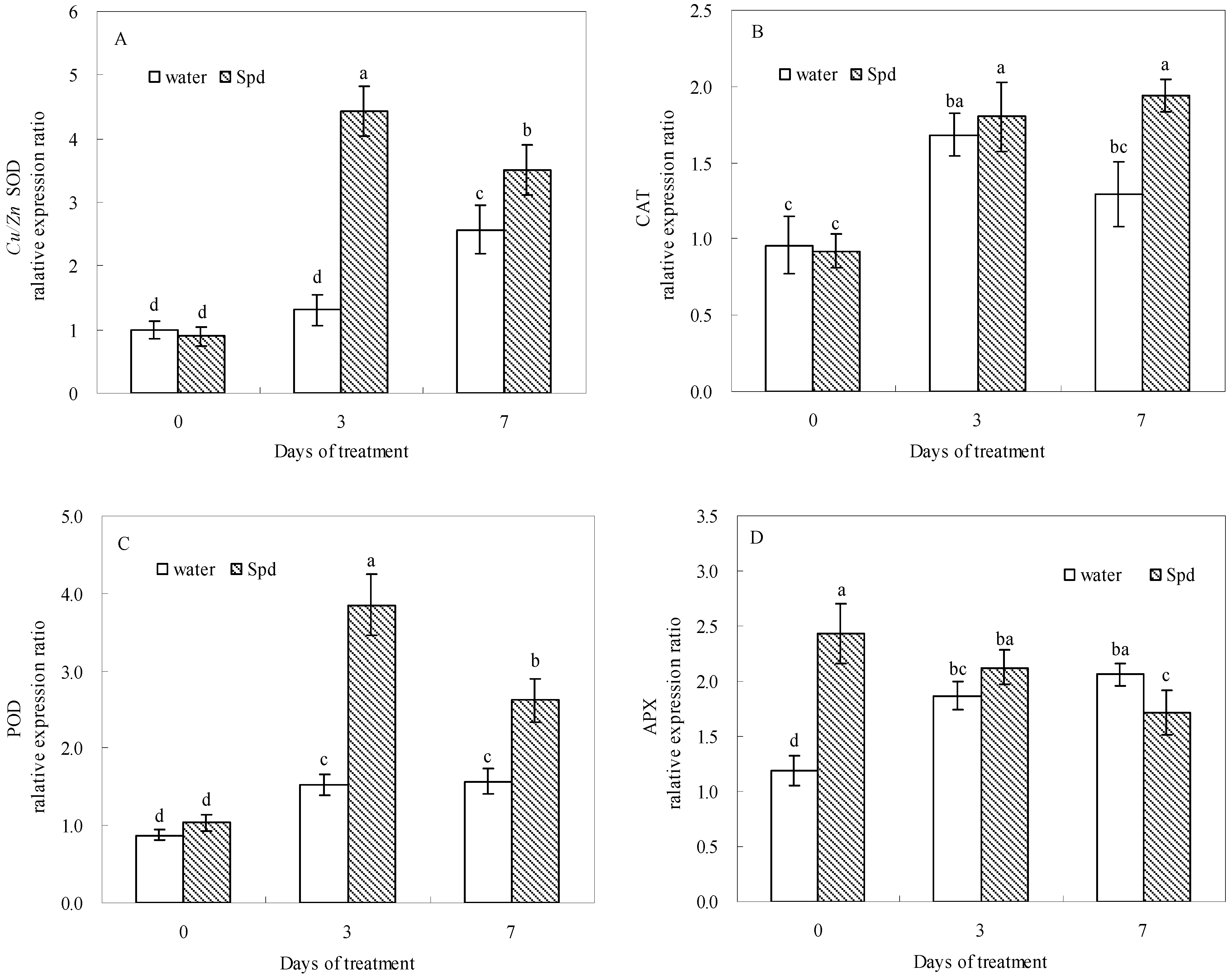
Under water stress, the exogenous Spd-treated seeds showed higher transcript levels of SOD, CAT, POD and APX genes compared with control during germination (
Figure 6). At 0 day of germination, seeds SOD, CAT and POD gene transcript levels in both treatments kept the similar level without statistically significant differences, whereas SOD, POD and CAT gene transcript levels after Spd treatment is 3.4 and 2.5 times at 3 days of germination and 1.5 times at 7 days of germination that of control under water stress, respectively (
Figure 6A–C).
Figure 6.
The effect of seed soaking with water or spermidine (Spd) on (A) superoxide dismutase (SOD) gene; (B) catalase (CAT) gene; (C) guaiacol peroxidase (POD) gene and (D) ascorbate peroxidase (APX) gene relative expression ratio in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 4). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
Figure 6.
The effect of seed soaking with water or spermidine (Spd) on (A) superoxide dismutase (SOD) gene; (B) catalase (CAT) gene; (C) guaiacol peroxidase (POD) gene and (D) ascorbate peroxidase (APX) gene relative expression ratio in the process of seeds germination (7 days) under water stress induced by 15% PEG. Vertical bars indicate ± SE of mean (n = 4). Different letters above columns indicate significant difference. LSD (p ≤ 0.05).
A significantly higher APX genes relative expression ratio was observed in seeds priming with Spd compared to that in seeds priming with water after 3 h of soaking (0 day). After that, the transcript level of APX genes began to decline in Spd treatment and increase in control, and showed different expression patterns (
Figure 6D).
Table 3.
The relationship between the enzyme activities and gene expressions of enzymes; α-amylase, β-amylase, Cu/ZnSOD, CAT, POD and APX in the process of seeds germination, submitted to water stress induced by 15% PEG for 0 day, 3 days and 7 days. “+” induced; “—” suppressed; “ns”, not significantly changed as compared with seeds priming with water (control) according to SE at p ≤ 0.05.
Table 3.
The relationship between the enzyme activities and gene expressions of enzymes; α-amylase, β-amylase, Cu/ZnSOD, CAT, POD and APX in the process of seeds germination, submitted to water stress induced by 15% PEG for 0 day, 3 days and 7 days. “+” induced; “—” suppressed; “ns”, not significantly changed as compared with seeds priming with water (control) according to SE at p ≤ 0.05.
| | Enzyme Activity | Gene Expression |
|---|
| 0 day | 3 days | 7 days | 0 day | 3 days | 7 days |
|---|
| α-amylase | + | ns | ns | ns | ns | ns |
| β-amylase | ns | + | + | ns | + | ns |
| Cu/ZnSOD | ns | + | + | ns | + | + |
| CAT | ns | + | ns | ns | ns | + |
| POD | ns | + | + | ns | + | + |
| APX | ns | + | + | + | ns | — |
The relations between gene expressions and enzyme activities were examined (
Table 3). Under water stress, a positive correlation between the biochemical and the molecular levels of enzyme-gene expression was found in seedlings of Spd-treated white clover at 3 d of treatment for β-amylase, SOD and POD, whereas a negative correlation was found at 7 d of treatment for APX. For α-amylase and CAT, a parallel induction of genes and enzymatic activities was observed as response to water stress at 3 and 7 d of treatment (
Table 4).
Table 4.
The effect of seed priming with water or spermidine (Spd) on ascorbic acid (AsA) and dehydroascorbic acid (DAsA) content in seeds of white clover after 7 days of germination under 15% PEG. Values are mean ± SE (n = 6). Different letters in a vertical column indicate significant difference between two treatments. LSD (p ≤ 0.05).
Table 4.
The effect of seed priming with water or spermidine (Spd) on ascorbic acid (AsA) and dehydroascorbic acid (DAsA) content in seeds of white clover after 7 days of germination under 15% PEG. Values are mean ± SE (n = 6). Different letters in a vertical column indicate significant difference between two treatments. LSD (p ≤ 0.05).
| Treatment | Content (umol.g−1 FW) |
|---|
| AsA | DAsA | AsA + DAsA | AsA/DAsA |
|---|
| Water | 18.77 ± 0.66 b | 3.45 ± 0.46 a | 20.97 ± 1.31 b | 5.21 ± 1.14 b |
| Spd | 21.42 ± 1.46 a | 1.52 ± 0.20 b | 23.59 ± 1.04 a | 14.59 ± 1.33 a |
2.6. Effect of Exogenous Spermidine on Ascorbic Acid and Glutathione Content
Total ascorbic acid, ascorbic acid (AsA), dehydroascorbic acid (DAsA) and the DAsA/AsA concentrations are shown in
Table 5. Exogenous Spd improved significantly the ascorbic acid (AsA), total ascorbic acid (AsA + DAsA) and the DAsA/AsA levels in white clover seedlings under water stress. On the contrary, dehydroascorbic acid (DAsA) content decreased by 56% with Spd treatment compared with that in control when exposed to 7 days of germination and showed a statistically significant difference (
Table 5). There were significant differences on reduced glutathione (GSH), total glutathione (GSH+GSSG) content and GSH/GSSG between two treatments at 3 days of germination (
Table 5). The 43% increase of GSH/GSSG in Spd treatment was observed relative to control and that indicated that the GSH turnover rate remained higher in Spd treatment when seeds germination of white clover under water stress.
Table 5.
The effect of seed priming with water or spermidine (Spd) on reduced glutathione (GSH) and glutathione disulfide (GSSG) content in seeds of white clover after 7 days of germination under 15% PEG. Values are mean ± SE (n = 6). Different letters in a vertical column indicate significant difference between two treatments. LSD (p ≤ 0.05).
Table 5.
The effect of seed priming with water or spermidine (Spd) on reduced glutathione (GSH) and glutathione disulfide (GSSG) content in seeds of white clover after 7 days of germination under 15% PEG. Values are mean ± SE (n = 6). Different letters in a vertical column indicate significant difference between two treatments. LSD (p ≤ 0.05).
| Treatment | Content (umol.g−1 FW) |
|---|
| GSH | GSSG | GSH+GSSG | GSH/GSSG |
|---|
| water | 1.37 ± 0.423 b | 2.01 ± 0.21 a | 3.29 ± 0.27 b | 0.75 ± 0.21 b |
| Spd | 2.29 ± 0.21 a | 1.86 ± 0.11 a | 4.01 ± 0.30 a | 1.07 ± 0.19 a |
2.8. Discussion
Seed germination is a complicated physiological process modulated by phytohormones or physiological activator such as abscisic acid [
29], nitric oxide [
30] or polyamines [
31]. Vigorous seedlings were receptive to effective seed priming with polyamines (PAs), and were evidently better able to resist the adverse effects of drought [
25]. Recently, it has been postulated that PAs can improve the cell water status in rice, thereby producing better growth under limited water supply conditions [
32]. Although the effect of all the PAs was stimulatory, spermidine (Spd) was statistically effective in most of the attribute studies [
18,
33]. The results of our study revealed that seeds primed with Spd not only showed improved germination percentages and shortened mean germination times but also displayed significantly enhanced seed vigor as indicated by longer root length, seedling fresh and dry weights compared with control. The result is consistent with an earlier study of Sinska and Lewandowska
et al. [
33] about the effect of exogenous Spd on apple seeds. Farooq
et al., [
2] found a strong correlation between amylase and soluble sugars which supported the assumption that speedy starch metabolism contributed towards the early emergence of seeds and vigorous seedling growth induced by priming with Spd. Metabolites of starch such as glucose are critical for seed germination as they are involved as osmolytes for cellular turgor maintenance and energy sources. Under water deficit, seed priming with Spd improved starch metabolism as indicated in this study by elevated α- and β-amylase activities in agreement with the results of Farooq
et al., [
2]. The data revealed that Spd plays an important role in accelerating starch metabolism during seed germination in response to water stress.
One of intricate effects of drought is loss of integrity of biological membranes due to oxidative damage; accordingly, maintenance of balance between the generation and quenching of reactive oxygen species (ROS) in plants is crucial to survive water stress [
34,
35]. Many previous studies have showed that PAs reduced ROS levels, for instance, under osmotic stress exogenous PAs inhibited the accumulation of O
2− and H
2O
2 in wheat seedlings and barley leaves [
36,
37]. Transgenic pear exhibited more accumulation of Spd when exposed to osmotic stress and displayed higher SOD and APX activities, less MDA and H
2O
2 content than the wild type, implying it suffered from less injury [
38,
39]; on the contrary, antisense inhibition of a Spd synthase gene reduced the antioxidant system significantly in pear accompanied by a greater accumulation of MDA compared with wild-type. Growth inhibition, Spd level, and antioxidant system were significantly ameliorated by exogenous Spd application in pear shoots [
40]. In current study, exogenous Spd significantly promoted activities of SOD, POD, CAT and APX relative to untreated seeds at different stress and germination time. The same result was obtained in study of Li [
41] about effect of exogenous Spd on wheat seedlings under osmotic stress. The ascorbate-glutathione cycle (AsA-GSH cycle) is the major non-enzymatic antioxidant process scavenging H
2O
2 in different cellular compartments [
42]. Significantly higher AsA/DAsA and GSH/GSSG ratios in seeds primed with Spd were observed in this study, which suggested that Spd accelerated the regeneration of AsA and GSH when seeds were germinated under water deficit. As a result, enhanced antioxidant enzyme activities and AsA-GSH cycle in Spd-treated seeds resulted in lower the generation rate of O
2−, H
2O
2 and MDA content and improved cell membrane stability, as demonstrated by lower electrolyte leakage (EL). This indicates that Spd is able to influence the antioxidant defense system to moderate the oxidative stress intensity induced by water stress during seed germination. These results further confirm the action of Spd acts as ROS scavengers.
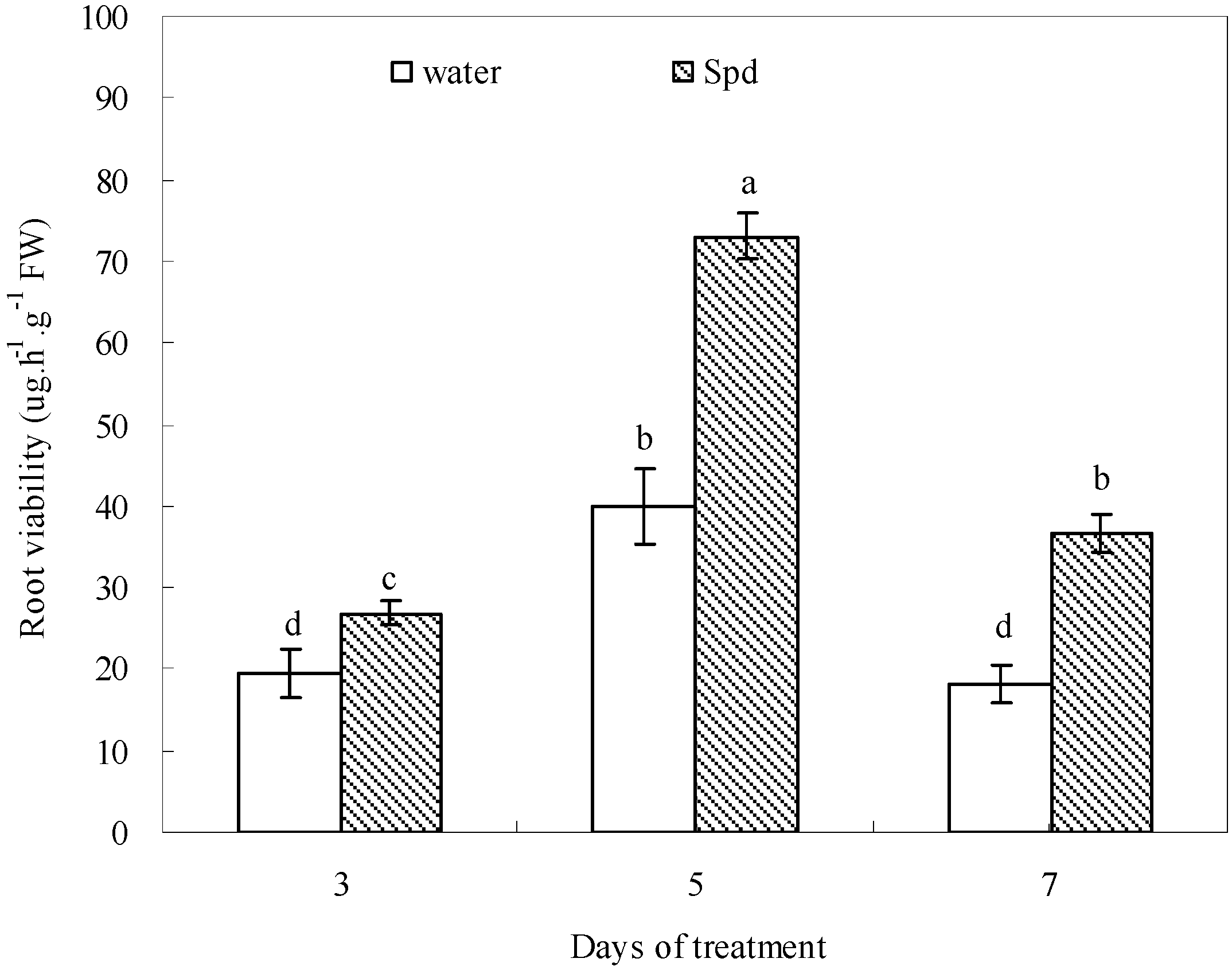
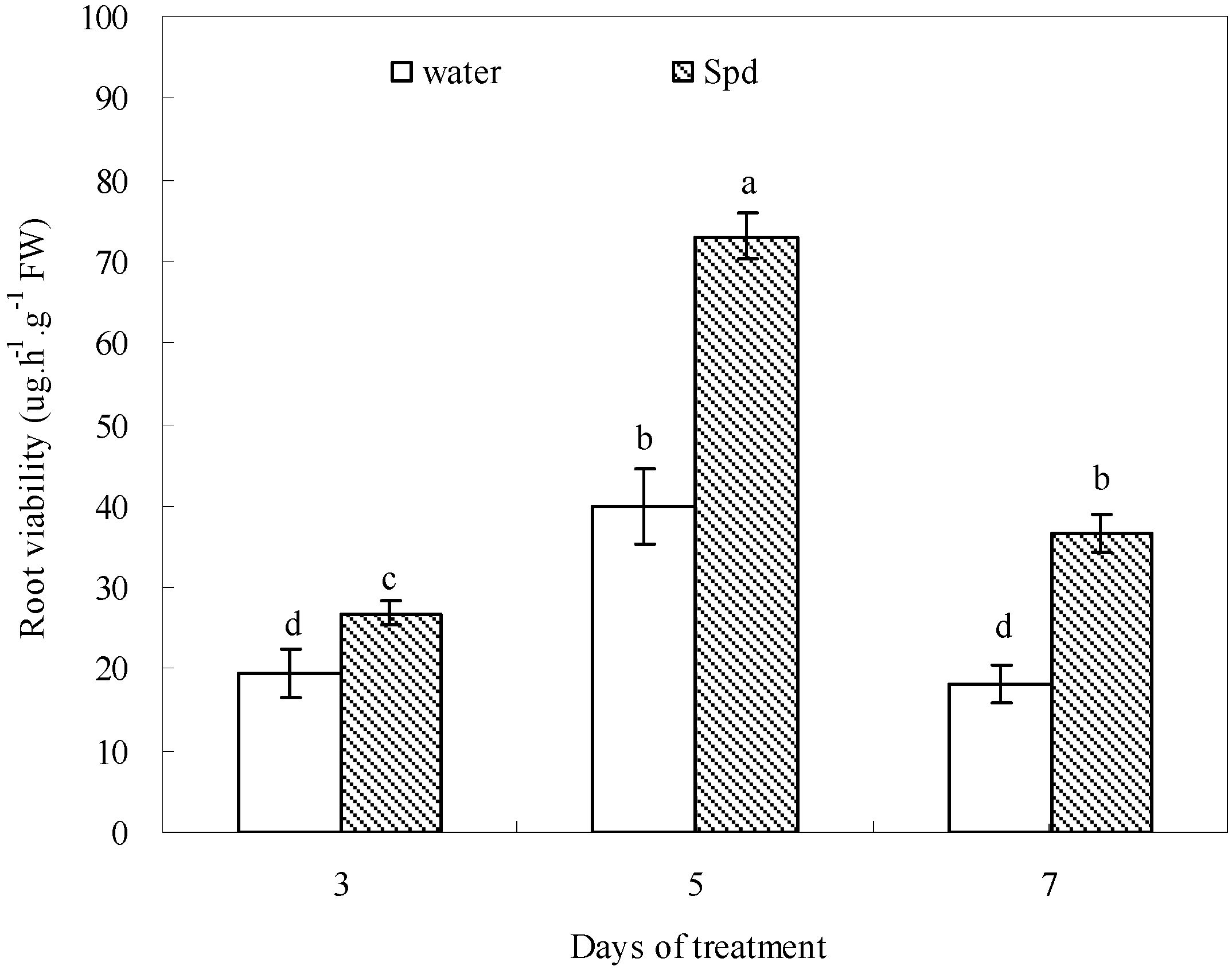
The role of PAs in root mortality and senescence caused by water deficit remains less well-documented. It has been reported that a drought-tolerant white clover cultivar exhibited significantly higher root viability than a drought-sensitive one under drought-stressed conditions [
43]. Our results showed that Spd may be involved in regulating root mortality, since seeds primed with exogenous Spd had significantly increased root viability and length during germination under water stress. According to the report of Xu
et al. [
44], root was more easily affected by Put than shoot during seed germination in chilling-stressed tobacco. High Spd level was characteristic of meristematic cells and might be essential for the development of globular structure and root length [
45,
46]. It is possible that the increase in antioxidant defense and the maintenance of higher starch metabolism in Spd-treated seeds under water stress could have stimulated root antioxidants to reduce root lipid peroxidation to maintain root survival and transported adequate carbon to the roots to maintain root growth. However, the accurate mechanism deserves more investigation, especially the effects of Spd on root tip viability when seed germination was subjected to water deficit.
PAs can regulate many enzyme activities by bonding with the enzyme protein or participation in the process of phosphorylation of the enzyme protein [
47,
48]. It was suggested that PAs were implicated in directly regulating antioxidant enzyme activities, thereby reducing oxidative stress [
49]. Apart from such direct stress protective roles, PAs may also play a role as a signaling molecule in plant responses to stress [
28]. Spd and Spm in particular promote gene expression and increase the DNA-binding activity of transcription factors [
50,
51]. The results from RT-PCR analysis in the present study showed that exogenous spermidine strongly affected the expression of genes encoding antioxidant enzymes and β-amylase during white clover seed germination subjected to water deficit, but there was no significant influence on the transcript level of α-amylase. However, exogenous Spd significantly enhanced α-amylase activity under the same water stress condition and germination time. As far as we know, the α-amylase is synthesized
de novo in the aleurone layer surrounding the endosperm in the process of seed germination. In contrast, a precursor form of β-amylase is present in seeds in the dry state as a proenzyme devoid of enzyme activity, which is activated during germination after cleavage of a peptide sequence at the C-terminal part of the enzyme [
12]. These results suggested that Spd might not only enhance the activities of α-amylase and β-amylase, but also induce the
de novo β-amylase synthesis by elevating β-amylase gene expression at the early stage of white clover seed germination under water stress. Moreover, the better germination of Spd-treated seeds under water stress could be associated with increased expression of specific genes encoding antioxidant enzymes, which partly influenced or improved antioxidant enzyme activities under stress. For antioxidant enzymes, the gene expression pattern did not always go along with their activities changes. The discrepancy between gene expression and enzyme activity indicates enzyme activity changes were not only caused by mRNA levels, but also regulated at the post-transcriptional level and influenced by cellular metabolism [
44,
52]. The current research also supports the notion that the function of Spd is involved in both sides as a stress-protecting compound and signaling molecule.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}