Metabolism of Tanshinone IIA, Cryptotanshinone and Tanshinone I from Radix Salvia Miltiorrhiza in Zebrafish

Abstract
:1. Introduction
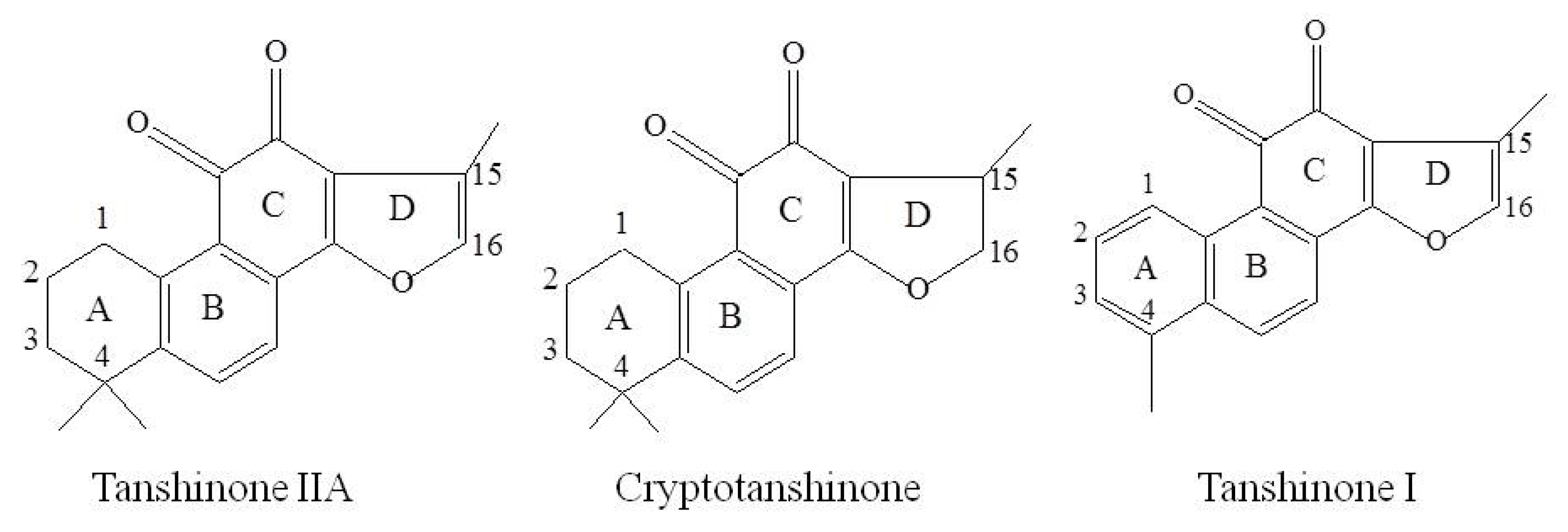
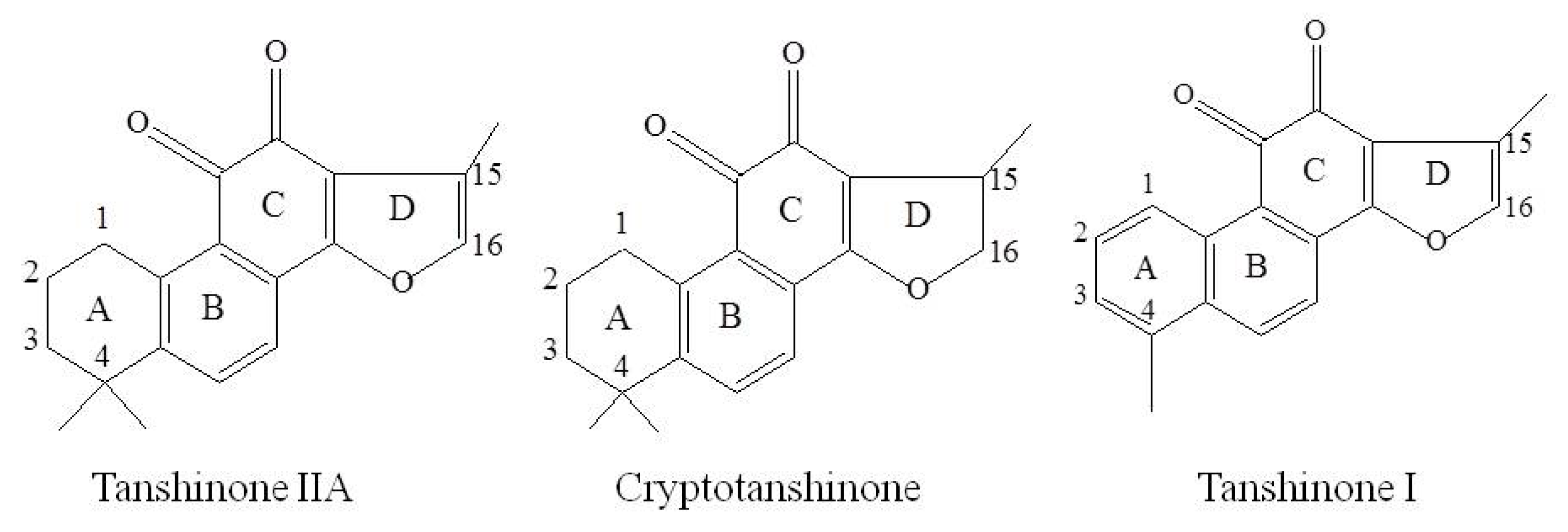
2. Results and Discussion
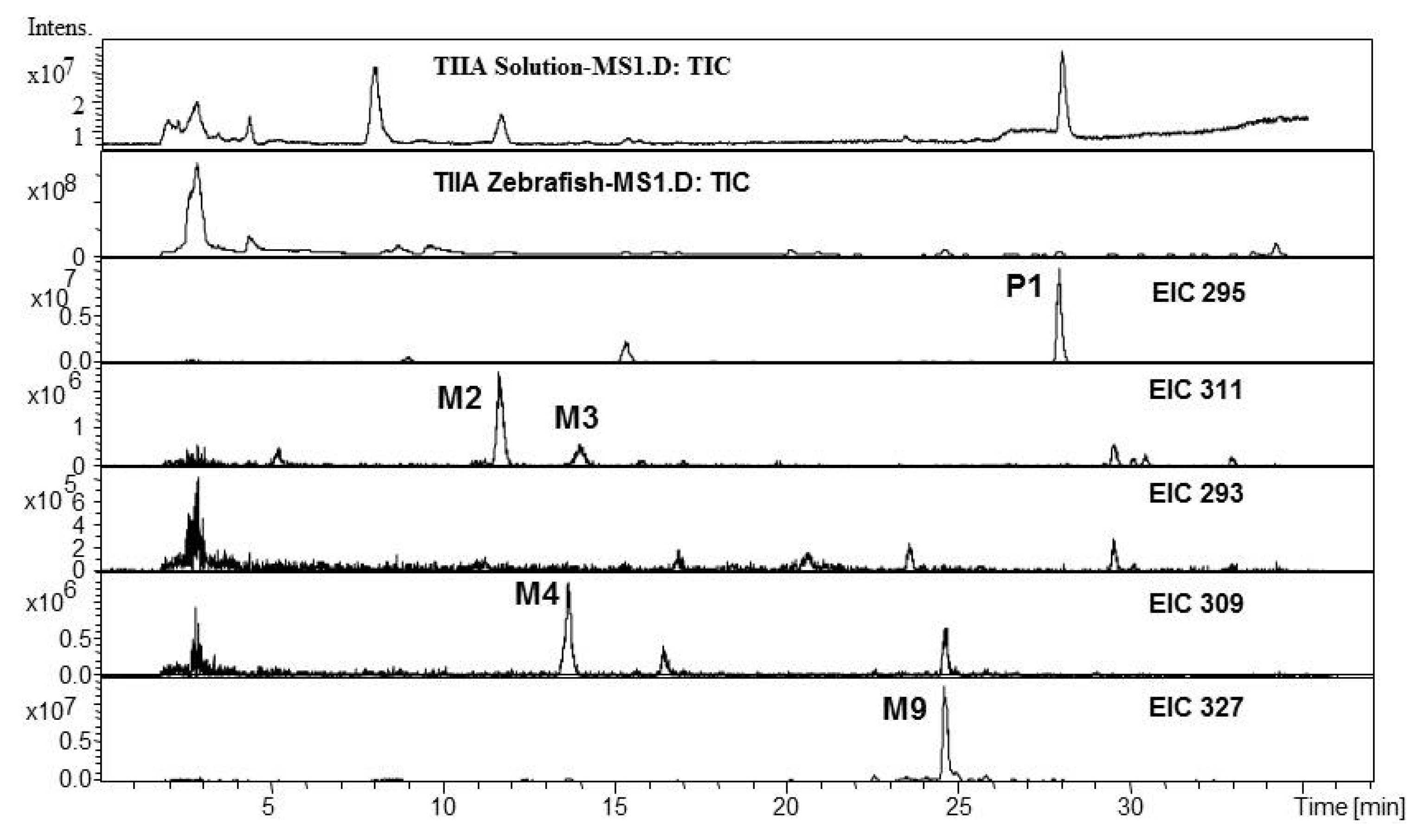
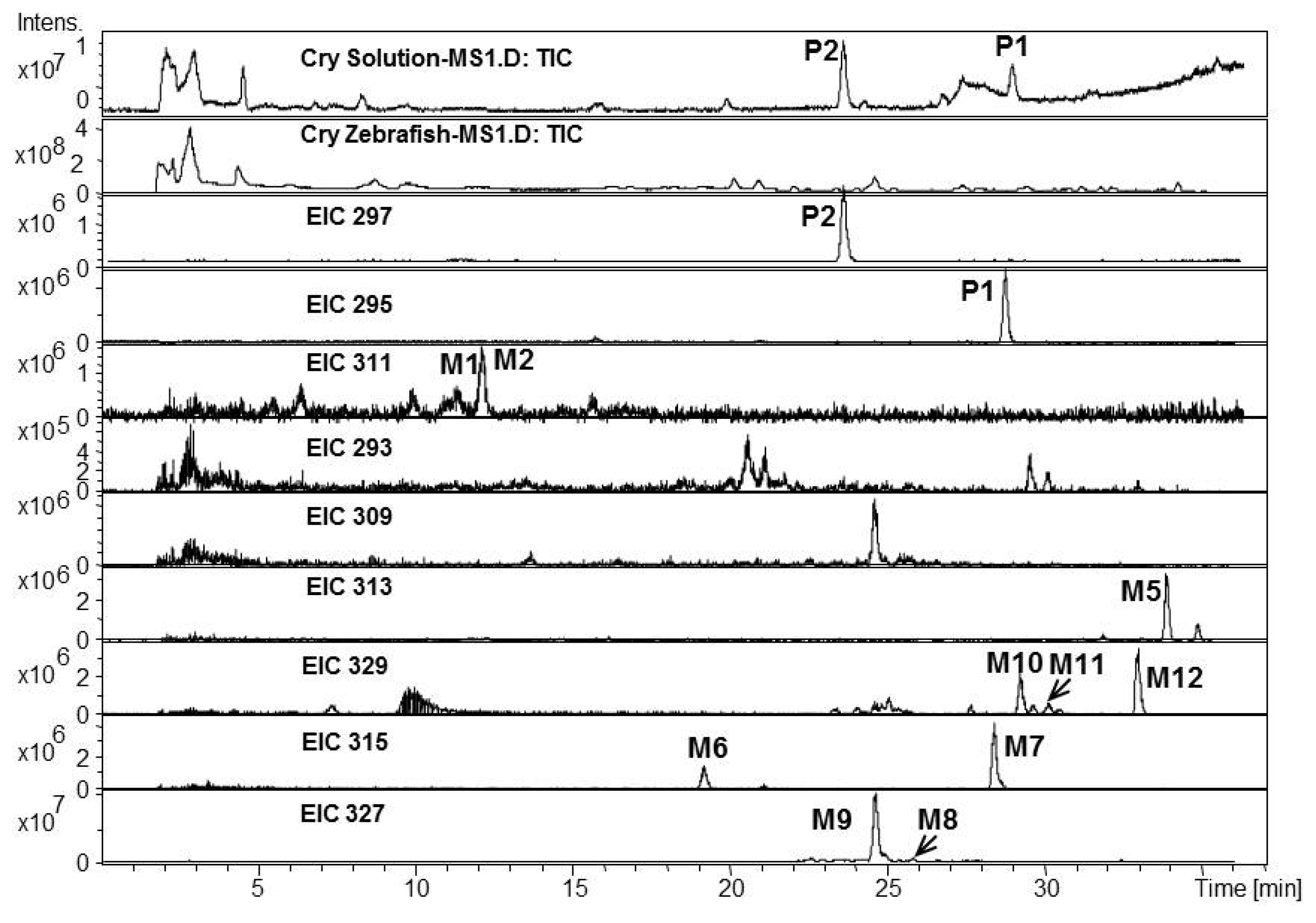
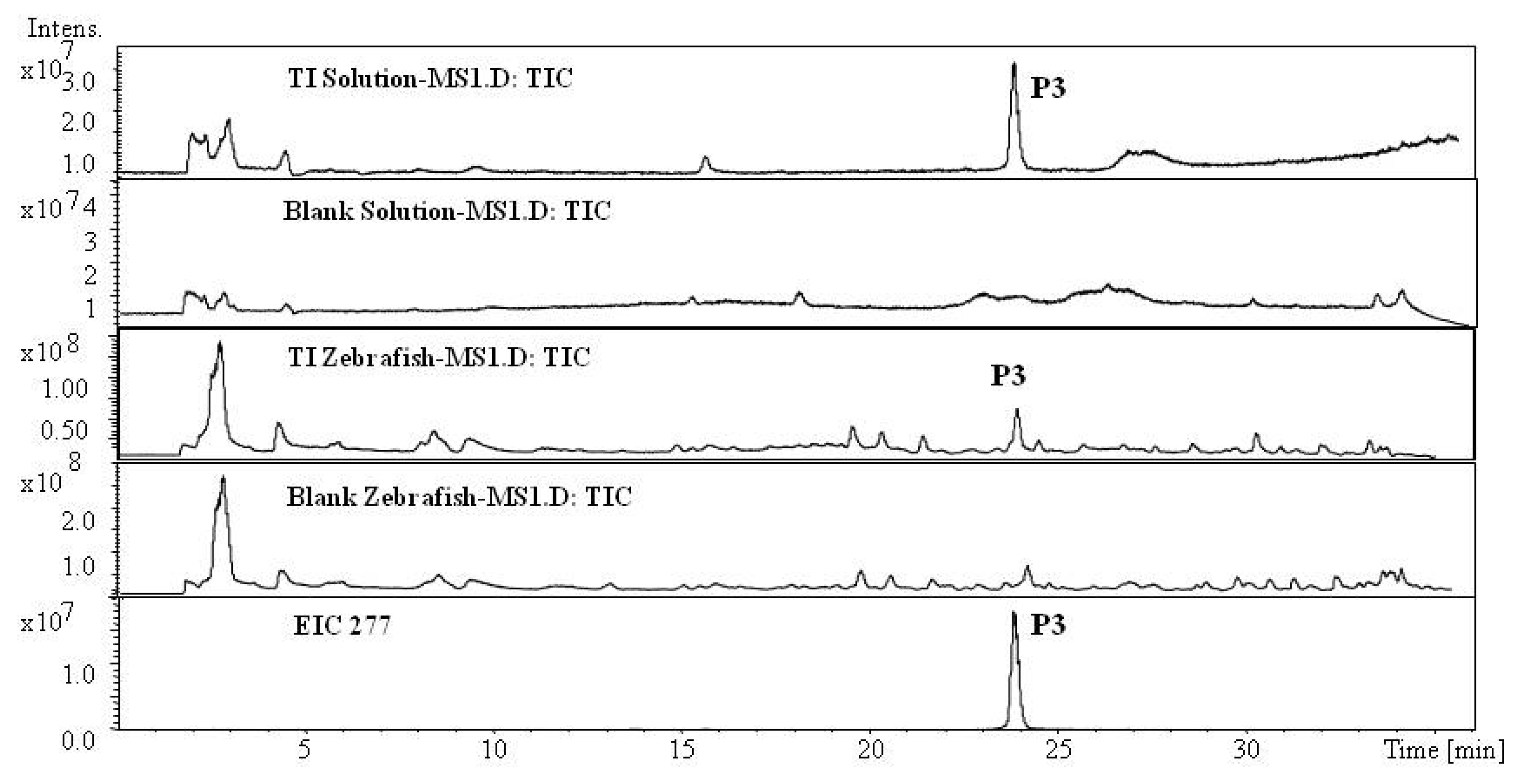
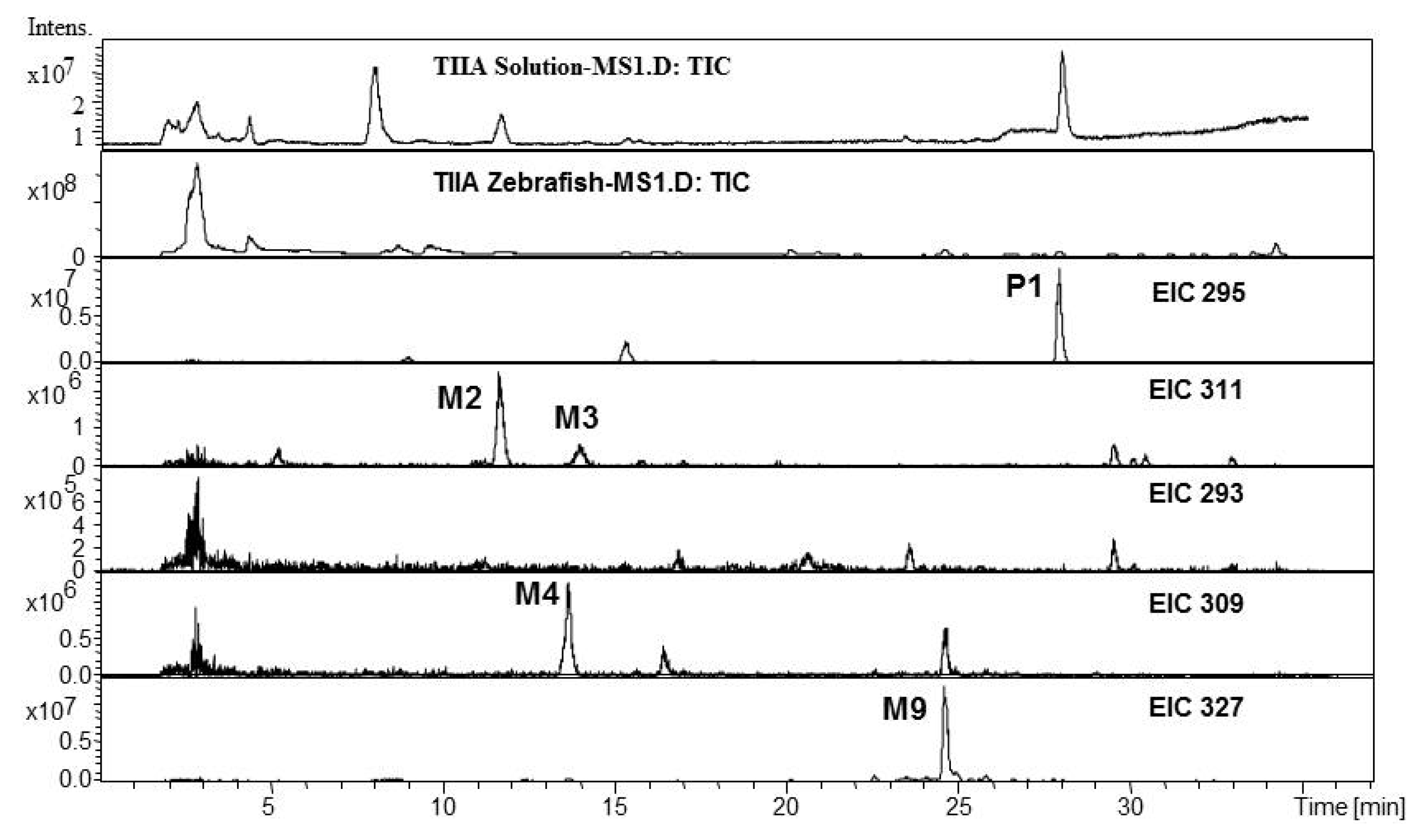
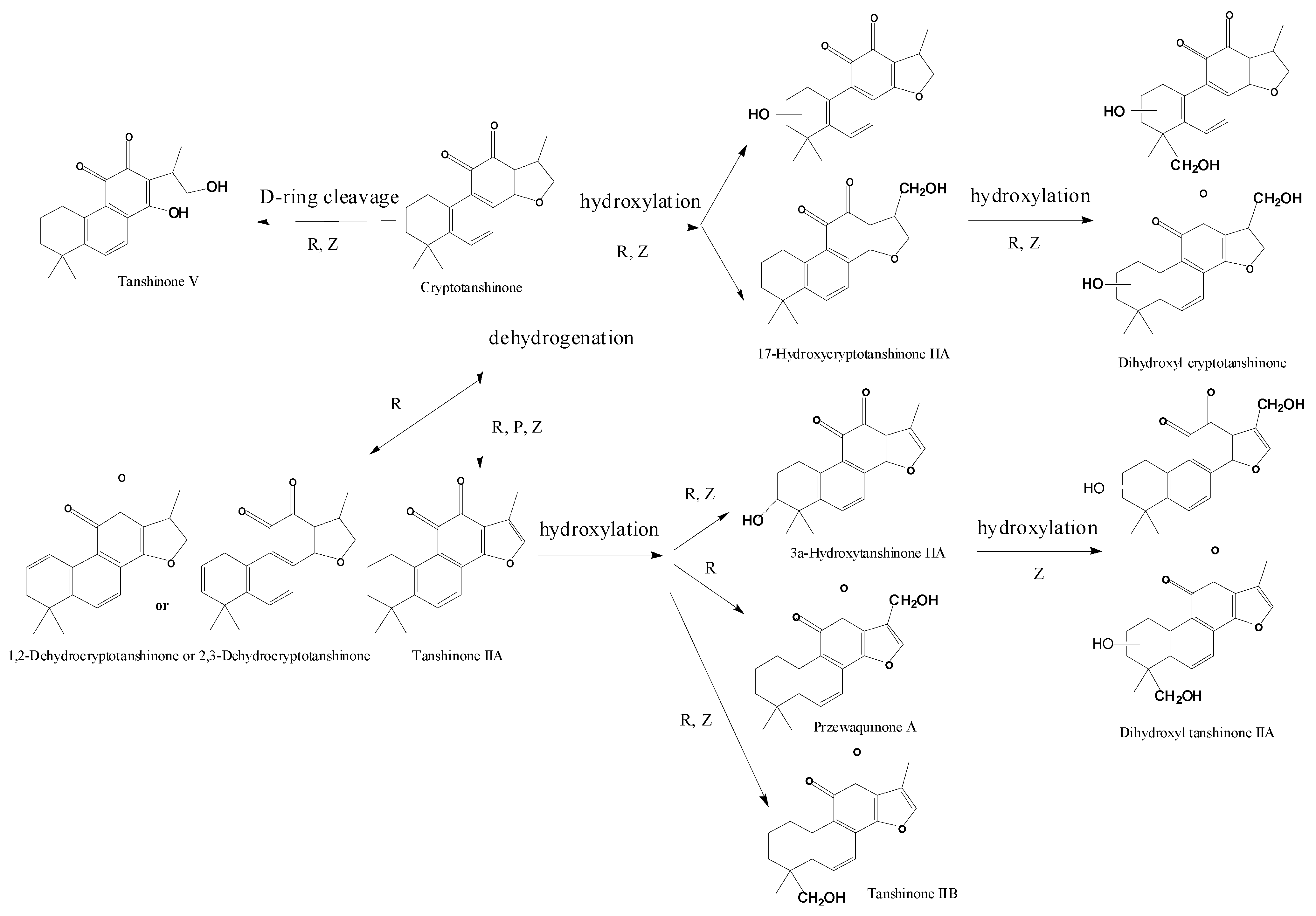
2.1. Analysis of Metabolic Components of TIIA, Cry and TI after Zebrafish Exposure by HPLC/IT-MSn

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
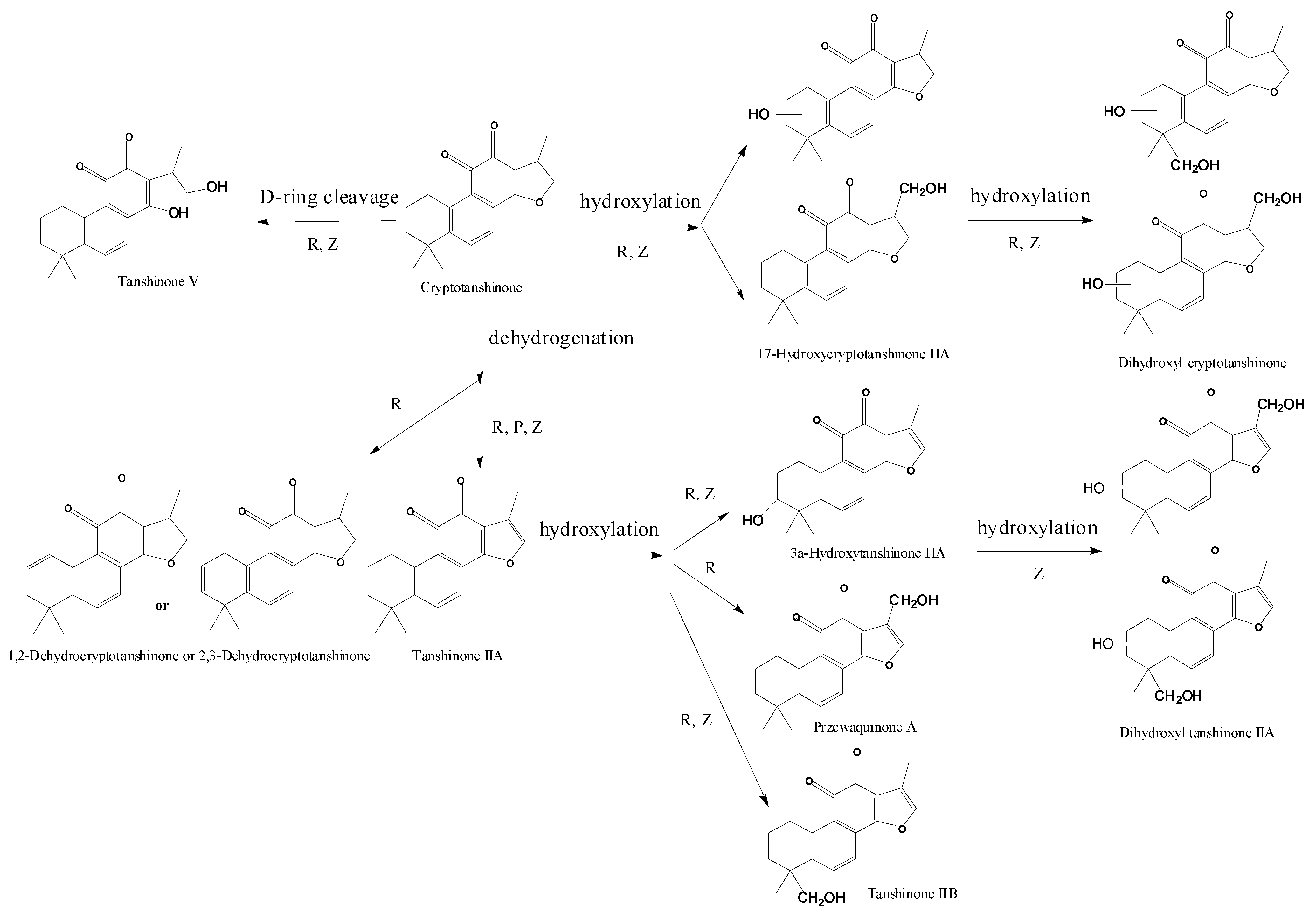
| Compounds | Retention Time (min) | Quasi-Molecular Ions Peak | HPLC/ESI-MSn Or fragment Ions | MW | Metabolite presumed | TIIA group | Cry group | TI group | Current Metabolism Reference | |
|---|---|---|---|---|---|---|---|---|---|---|
| [M+H]+ | [M+Na]+ | |||||||||
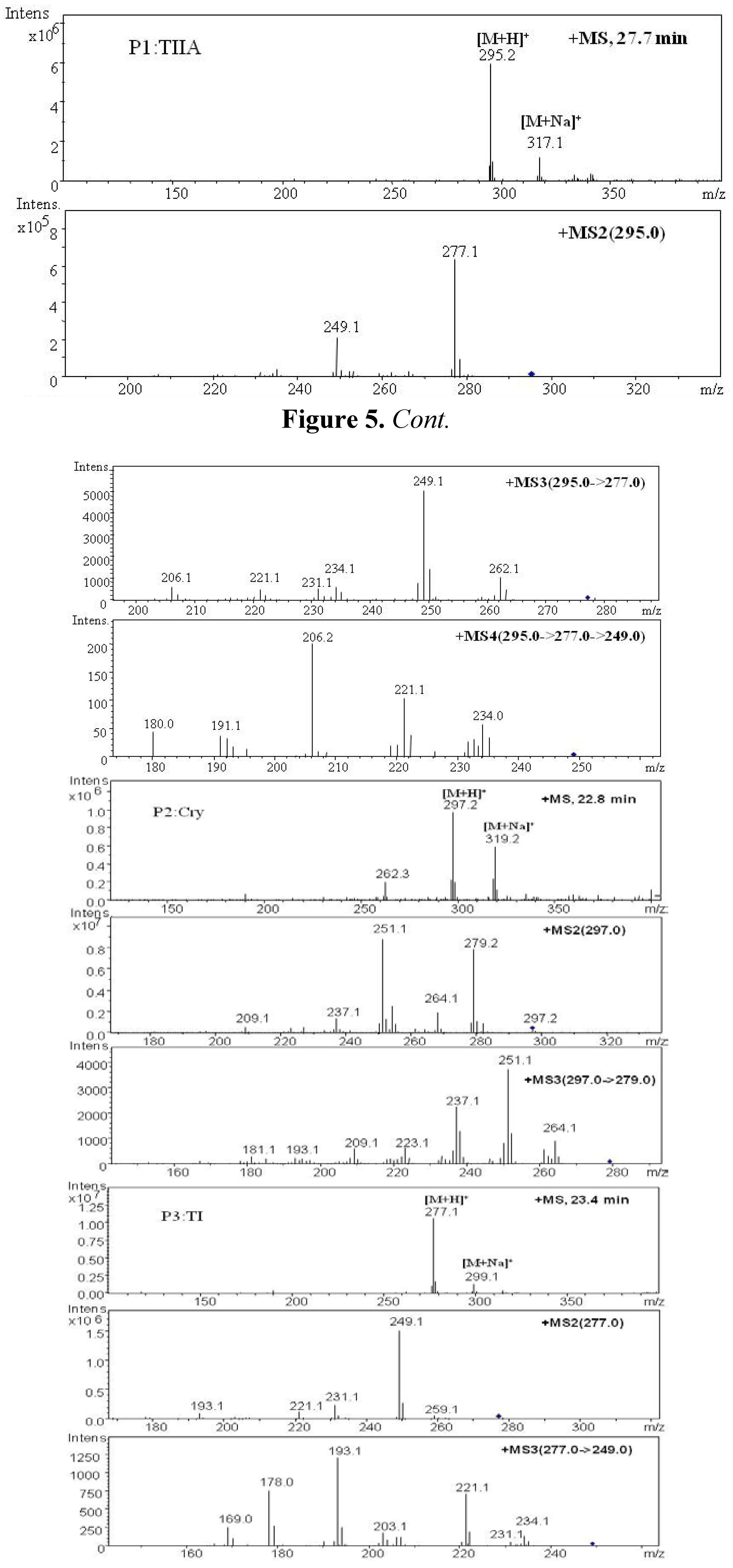
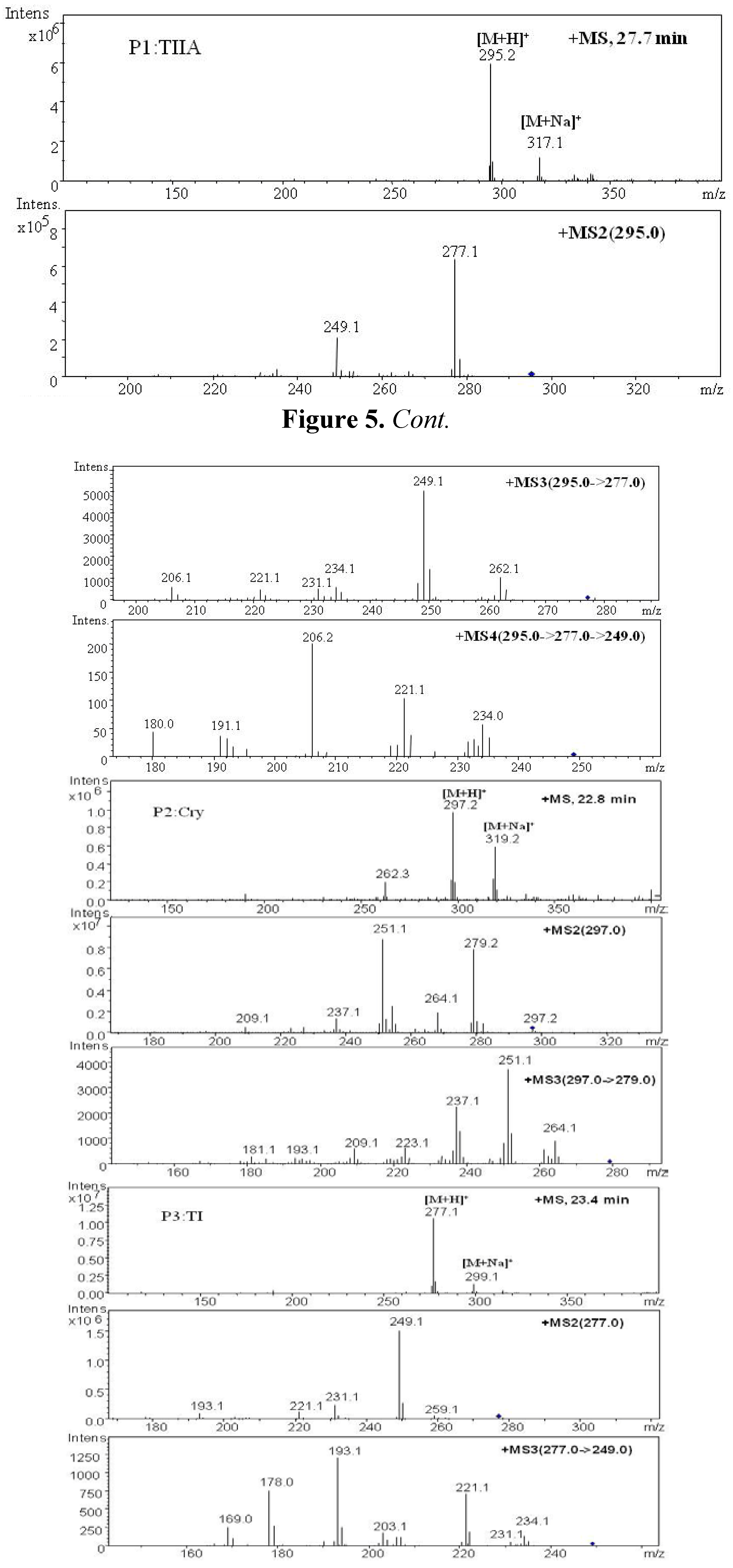
| P1 | 27.7 | 295.2 | 317.1 | MS2[295]: 277.1[M+H-H2O]+, 249.1[M+H-H2O-CO]+ | 294.2 | Tanshinone IIA | + | + | [ 29,32,34,38] | |
| MS3[295→277]: 262.1[M+H-CH3]+, 249.1[M+H-H2O-CO]+, 231.1[M+H-2H2O-CO]+, 221.1[M+H-H2O-2CO]+, 206.1[M+H-H2O-2CO-CH3]+ | ||||||||||
| MS4[295→277→249]: 234[M+H-H2O-CO-CH3]+, 221.1,206.2, 191.2[M+H-H2O-2CO-2CH3]+ | ||||||||||
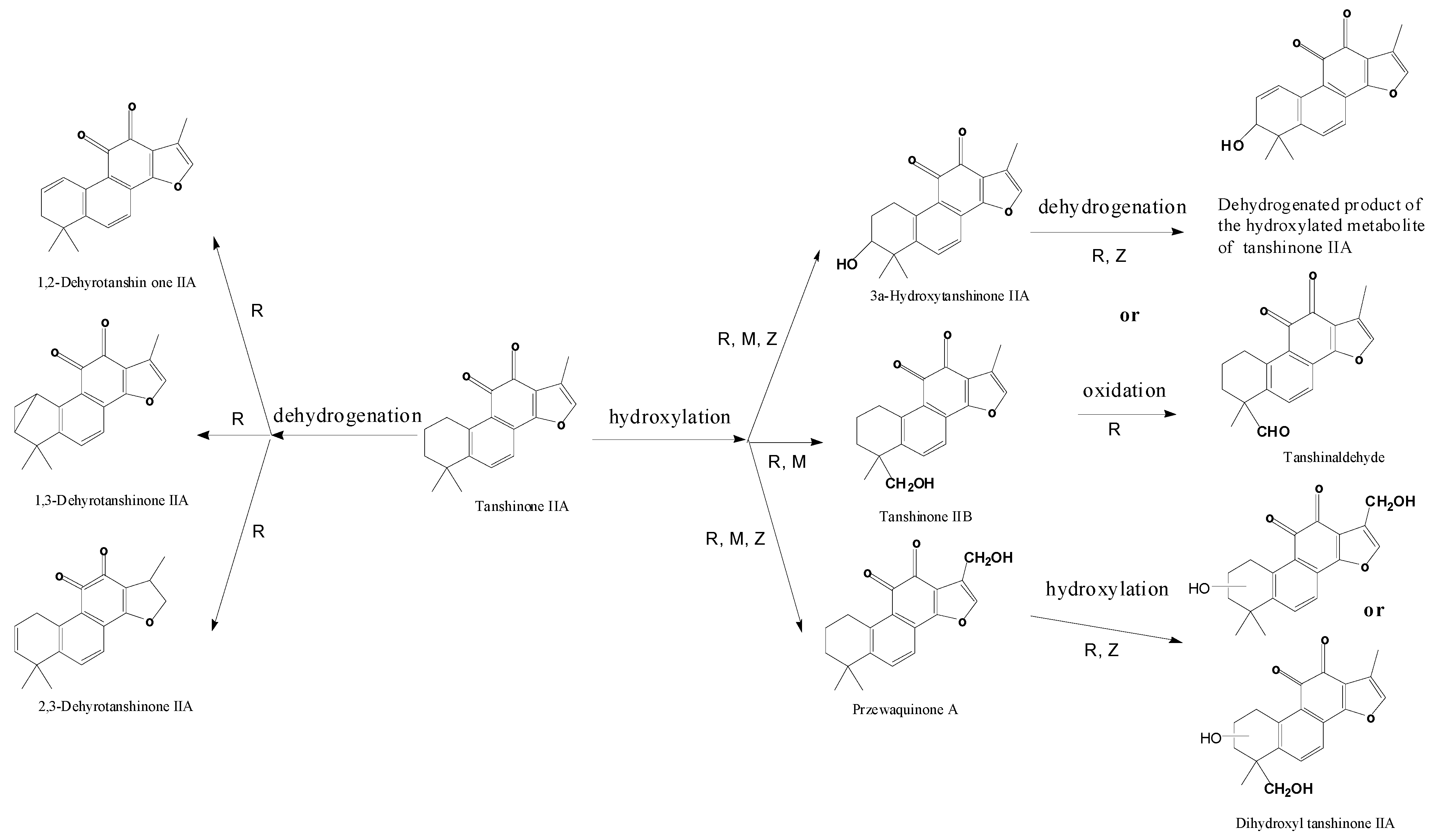
| P2 | 22.8 | 297.2 | 319.2 | MS2[297]: 297.2[M+H]+, 279.2[M+H-H2O]+, 264.1[M+H-H2O-CH3]+, 251.1[M+H-H2O-CO]+, 237.1[M+H-CO-CH3OH]+, 209.1[M+H-2CO-CH3OH]+ | 296.2 | Cryptotanshinone | + | [ 34,38] | ||
| MS3[297→279]: 264.1,251.1, 237.1,209.1 | ||||||||||
| P3 | 23.4 | 277.1 | 299.1 | MS2[277]: 259.1[M+H-H2O]+, 249.1[M+H-CO]+, 231.1[M+H-2H2O-CO]+, 221.1[M+H-H2O-2CO]+ | 276.1 | Tanshinone I | + | [ 34] | ||
| MS3[277→249]: 234.1[M+H-CO-CH3]+, 231.1[M+H-CO-H2O]+, 221.1[M+H-2CO]+, 203[M+H-2CO-H2O]+, 193.1[M+H-3CO]+, 178[M+H-3CO-CH3]+ | ||||||||||
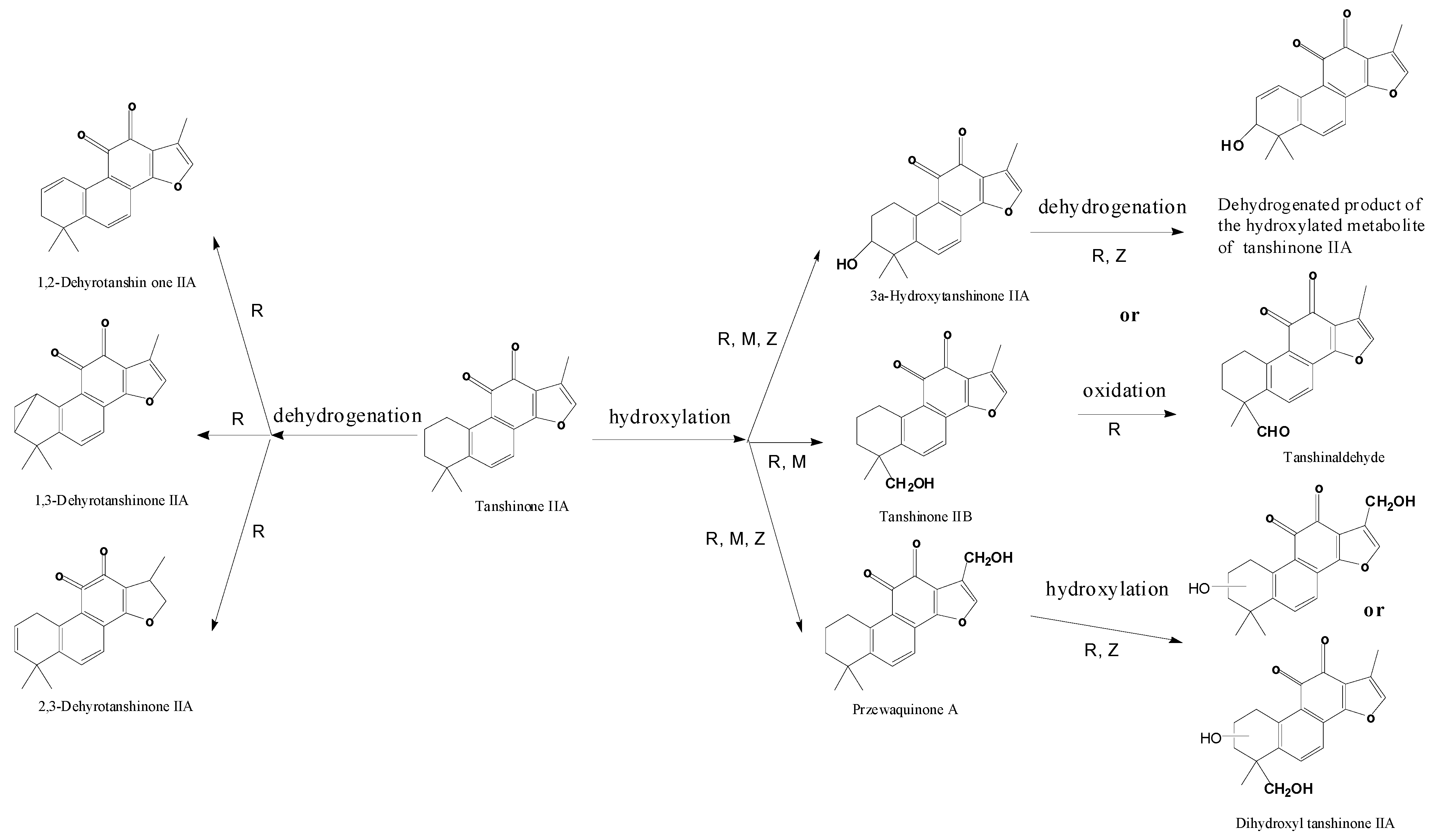
| M1 | 10.9 | 311.2 | 333.2 | 310.2 | Tanshinone IIB | + | [ 29,32,34,38] | |||
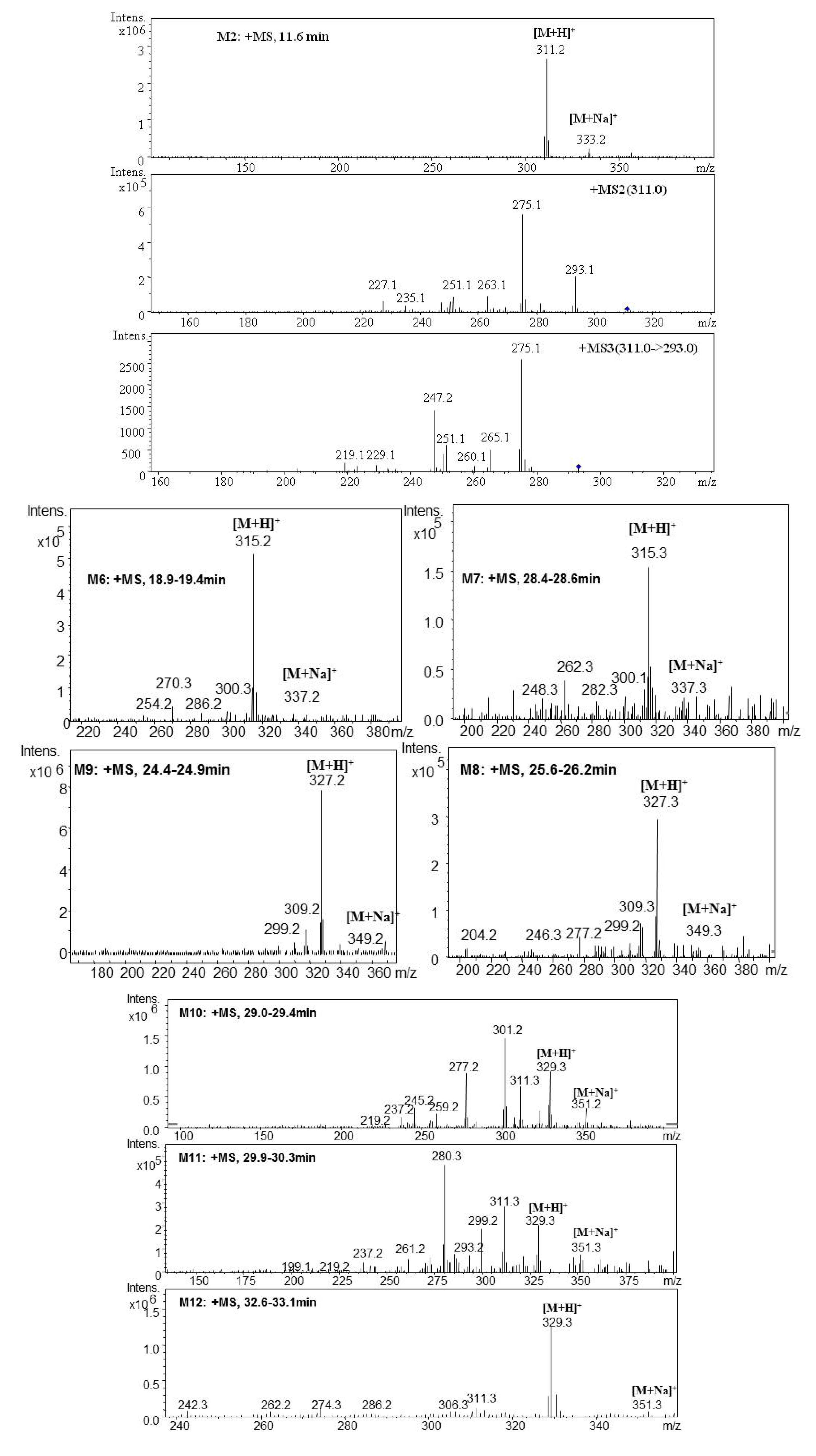
| M2 | 11.6 | 311.2 | 333.2 | MS2[311]: 293.1[M+H-H2O]+, 275.1[M+H-2H2O]+, 263.1[M+H-H2O-2CH3]+, 251.1[M+H-CO-CH3OH]+, 235.1[M+H-H2O-2CH3-CO]+ | 310.2 | 3α-Hydroxytanshinone IIA | + | + | [ 29,32,34,38] | |
| MS3[311→293]: 275.1[M+H-2H2O]+, 265.3[M+H-CO-H2O]+, 251.1[M+H-CO-CH3OH]+, 247.2 [M+H-CO-2H2O]+, 229.1[M+H-CO-3H2O]+, 219.1[M+H-2CO-2H2O]+ | ||||||||||
| M3 | 13.9 | 311.2 | 333.3 | 310.2 | Przewaquinone A | + | [ 29,32,34,38] | |||
| M4 | 13.6 | 309.1 | MS2[309]: 291.1[M+H-H2O]+, 277.2[M+H-CH3OH]+, 265.2[M+H-2CH3-CH3OH+H2O]+, 247.0[M+H-2CH3-CH3OH]+, 235.2[M+H-4CH3-CH3OH+H2O]+ | 308.1 | Dehydrogenated product ofthe hydroxylated metaboliteof tanshinone IIA | + | [ 34,38] | |||
| MS3[309→291]: 291.2[M+H-H2O]+, 273.2[M+H-2H2O]+, 261.2[M+H-H2O-2CH3]+, 245.1[M+H-2H2O-CO]+ | ||||||||||
| MS3[309→281]: 263.1,253.1[M+H-2CO]+, 239.1[M+H-2CO-CH3OH+H2O]+, 211.0[M+H-3CO-CH3OH+H2O]+, 201.2[M+H-4CO-CH3OH+2H2O]+, 183.0[M+H-4CO-CH3OH+H2O]+ | ||||||||||
| M5 | 34.0 | 313.3 | 335.3 | 256.3[M+H-CH3-CH3OH-CO+H2O]+, 230.3[M+H-CH3-CH3OH-2H2O]+, | 312.3 | Hydroxyl cryptotanshinone | + | [ 34,38] | ||
| M6 | 19.1 | 315.3 | 337.3 | 300.3[M+H-CH3]+, 286.2[M+H-CH3-CH3OH+H2O]+, 270.3[M+H-3CH3]+ | 314.3 | Tanshinone V | + | [ 34,38] | ||
| M7 | 28.5 | 315.3 | 337.3 | 300.1[M+H-CH3]+, 282.3[M+H-CH3-H2O]+, 262.3[M+H-2CO-CH3+H2O]+, | 314.3 | Tanshinone V isomer | + | [ 34,38] | ||
| M8 | 25.9 | 327.3 | 349.3 | 309.3[M+H-H2O]+, 299.2[M+H-CO]+, 277.2[M+H-H2O-CH3OH]+ | 326.3 | Dihydroxyl tanshinone IIA | + | [ 32] | ||
| M9 | 24.6 | 327.2 | 349.2 | 309.2[M+H-H2O]+, 299.2[M+H-CO]+ | 326.2 | Dihydroxyl tanshinone IIA | + | + | [ 32] | |
| M10 | 29.2 | 329.3 | 351.2 | 311.3[M+H-H2O]+, 301.2[M+H-CO]+ | 328.3 | Dihydroxyl cryptotanshinone | + | [ 33] | ||
| M11 | 30.1 | 329.3 | 351.3 | 311.3[M+H-H2O]+, 299.2[M+H-2CH3]+, 293.2[M+H-2H2O]+, 261.2[M+H-2H2O-CH3OH]+, 237.2[M+H-2H2O-2CO]+, 219.2[M+H-3H2O-2CO]+, 199.1[M+H-2H2O-3CO]+ | 328.3 | Dihydroxyl cryptotanshinone | + | [ 34] | ||
| M12 | 32.8 | 329.3 | 351.2 | 311.3[M+H-H2O]+, 286.2[M+H-CO-CH3]+ | 328.3 | Dihydroxyl cryptotanshinone | + | [ 34] | ||



2.2. Rationality and Advantages of Metabolic Study with Zebrafish Compared to Existing Models
3. Experimental
3.1. Chemicals and Reagents
3.2. Animals
3.3. Instruments
3.4. Biological Sample Collection
3.5. Sample Preparation
3.6. Analysis Condition
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Goldsmith, P. Zebrafish as a pharmacological tool: The how, why and when. Curr. Opin. Pharm. 2004, 4, 504–512. [Google Scholar] [CrossRef]
- Kari, G.; Rodeck, U.; Dicker, A.P. Zebrafish: An emerging model system for human disease and drug discovery. Clin. Pharmacol. Ther. 2007, 82, 70–80. [Google Scholar] [CrossRef]
- Den Hertog, J. Chemical genetics: Drug screens in zebrafish. Biosci. Rep. 2005, 25, 289–297. [Google Scholar] [CrossRef]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef]
- Rubinstein, A.L. Zebrafish assays for drug toxicity screening. Exp. Opin. Drug Metab. Toxicol. 2006, 2, 231–240. [Google Scholar] [CrossRef]
- Liang, A.H. Zebrafish—Useful model for pharmacodynamics and toxicity screening of traditional Chinese medicine. China J. Chin. Mater. Med. 2009, 34, 2839–2842. [Google Scholar]
- Thompson, E.D.; Burwinkel, K.E.; Chava, A.K.; Notch, E.G.; Mayer, G.D. Activity of Phase I and Phase II enzymes of the benzo[a]pyrene transformation pathway in zebrafish (Danio rerio) following waterborne exposure to arsenite. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2010, 152, 371–378. [Google Scholar]
- Almeida, D.V.; Nornberg, B.F.; Geracitano, L.A.; Barros, D.M.; Monserrat, J.M.; Marins, L.F. Induction of phase II enzymes and hsp70 genes by copper sulfate through the electrophile-responsive element (EpRE): Insights obtained from a transgenic zebrafish model carrying an orthologous EpRE sequence of mammalian origin. Fish Physiol. Biochem. 2010, 36, 347–353. [Google Scholar] [CrossRef]
- Meier, M.; Tokarz, J.; Haller, F.; Mindnich, R.; Adamski, J. Human and zebrafish hydroxysteroid dehydrogenase like 1 (HSDL1) proteins are inactive enzymes but conserved among species. Chem. Biol. Interact. 2009, 178, 197–205. [Google Scholar] [CrossRef]
- Liu, T.A.; Bhuiyan, S.; Liu, M.Y.; Sugahara, T.; Sakakibara, Y.; Suiko, M.; Yasuda, S.; Kakuta, Y.; Kimura, M.; Williams, F.E.; et al. Zebrafish as a model for the study of the phase II cytosolic sulfotransferases. Curr. Drug Metab. 2010, 11, 538–546. [Google Scholar] [CrossRef]
- Scornaienchi, M.L.; Thornton, C.; Willett, K.L.; Wilson, J.Y. Functional differences in the cytochrome P450 1 family enzymes from zebrafish (Danio rerio) using heterologously expressed proteins. Arch. Biochem. Biophys. 2010, 502, 17–22. [Google Scholar]
- Donnarumma, L.; de Angelis, G.; Gramenzi, F.; Vittozzi, L. Xenobiotic metabolizing enzyme systems in test fish. III. Comparative studies of liver cytosolic glutathione S-transferases. Ecotoxicol. Environ. Saf. 1988, 16, 180–186. [Google Scholar]
- Buhler, D.R.; Wang-Buhler, J.L. Rainbow trout cytochrome P450s: Purification, molecular aspects, metabolic activity, induction and role in environmental monitoring. Comp. Biochem. Physiol. Pharmacol. Toxicol. Endocrinol. 1998, 121, 107–137. [Google Scholar] [CrossRef]
- Morcillo, Y.; Janer, G.; O’Hara, S.C.; Livingstone, D.R.; Porte, C. Interaction of tributyltin with hepatic cytochrome P450 and uridine diphosphate-glucuronosyl transferase systems of fish: In vitro studies. Environ. Toxicol. Chem. 2004, 23, 990–996. [Google Scholar]
- Stuchal, L.D.; Kleinow, K.M.; Stegeman, J.J.; James, M.O. Demethylation of the pesticide methoxychlor in liver and intestine from untreated, methoxychlor-treated, and 3-methylcholanthrene-treated channel catfish (Ictalurus punctatus): Evidence for roles of CYP1 and CYP3A family isozymes. Drug Metab. Dispos. 2006, 34, 932–938. [Google Scholar]
- Wiegand, C.; Pflugmacher, S.; Giese, M.; Frank, H.; Steinberg, C. Uptake, toxicity, and effects on detoxication enzymes of atrazine and trifluoroacetate in embryos of zebrafish. Ecotoxicol. Environ. Saf. 2000, 45, 122–131. [Google Scholar]
- Fitzsimmons, P.; Lien, G.; Nichols, J. A compilation of in vitro rate and affinity values for xenobiotic biotransformation in fish, measured under physiological conditions. Comp. Biochem. Physiol. 2007, 145, 485–506. [Google Scholar]
- Gonzàlez, J.; Reimschuessel, R.; Shaikh, B.; Kane, A. Kinetics of hepatic phase I and II biotransformation reactions in eight finfish species. Mar. Environ. Res. 2009, 67, 183–188. [Google Scholar]
- Hu, G.; Siu, S.O.; Li, S.; Chu, I.K.; Kwan, Y.W.; Chan, S.W.; Leung, G.P.; Yan, R.; Lee, S.M. Metabolism of calycosin, an isoflavone from Astragali Radix, in zebrafish larvae. Xenobiotica 2012, 42, 294–303. [Google Scholar] [CrossRef]
- Jones, H.S.; Trollope, H.T.; Hutchinson, T.H.; Panter, G.H.; Chipman, J.K. Metabolism of ibuprofen in zebrafish larvae. Xenobiotic 2012. [Google Scholar] [CrossRef]
- Lindholst, C.; Wynne, P.M.; Marriott, P.; Pedersen, S.N.; Bjerregaard, P. Metabolism of bisphenol A in zebrafish (Danio rerio) and rainbow trout (Oncorhynchus mykiss) in relation to estrogenic response. Comp. Biochem. Physiol. Toxicol. Pharmacol. 2003, 135, 169–177. [Google Scholar]
- Chen, Q.X.; Zeng, S. Research progress of zebrafish used in drug metabolism. Yao Xue Xue Bao 2011, 46, 1026–1031. [Google Scholar]
- Wei, Y.J.; Li, P.; Fan, H.W.; Peng, Y.R.; Wang, C.M.; Shu, L.; Jia, X.B. Metabolism study of notoginsenoside R1, ginsenoside Rg1 and ginsenoside Rb1 of Radix Panax notoginseng in zebrafish. Molecules 2011, 16, 6621–6633. [Google Scholar] [CrossRef]
- Ho, S.Y.; Pack, M.; Farber, S.A. Analysis of small molecule metabolism in zebrafish. Methods Enzymol. 2003, 364, 408–426. [Google Scholar]
- Wei, Y.J.; Li, P.; Fan, H.W.; Sun, E.; Wang, C.M.; Shu, L.; Liu, W.; Xue, X.L.; Qian, Q.; Jia, X.B. Metabolite profiling of four major flavonoids of Herba Epimedii in zebrafish. Molecules 2012, 17, 420–432. [Google Scholar]
- Wei, Y.J.; Ning, Q.; Jia, X.B.; Gong, Z.N. Thoughts and methods for metabolic study of Chinese materia medica based on zebrafish model. Chin. Tradit. Herb. Drugs 2009, 40, 1009–1011. [Google Scholar]
- Du, G.H.; Zhang, J.T. The general situation and progress of the modern research of red sage root (Radix Salviae miltiorrhizae). Her. Med. 2004a, 23, 355–360. [Google Scholar]
- Du, G.H.; Zhang, J.T. The general situation and progress of the modern research of red sage root (Radix Salviae miltiorrhizae). Her. Med. 2004b, 23, 435–440. [Google Scholar]
- Li, P.; Wang, G.J.; Li, J.; Hao, H.P.; Zheng, C.N. Identification of tanshinone IIA metabolites in rat liver microsomes by liquid chromatography-tandem mass spectrometry. J. Chromatogr. 2006, 1104, 366–369. [Google Scholar]
- Li, J.; Wang, G.J.; Li, P.; Hao, H.P. Simultaneous determination of tanshinone IIA and cryptotanshinone in rat plasma by liquid chromatography-electrospray ionisation-mass spectrometry. J. Chromatogr. Analyt. Technol. Biomed. Life Sci. 2005, 826, 26–30. [Google Scholar] [CrossRef]
- Li, P.; Wang, G.J.; Li, J.; Hao, H.P.; Zheng, C.N. Simultaneous determination of tanshinone IIA and its three hydroxylated metabolites by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 815–822. [Google Scholar]
- Li, P.; Wang, G.J.; Li, J.; Hao, H.P.; Zheng, C.N. Characterization of metabolites of tanshinone IIA in rats by liquid chromatography/tandem mass spectrometry. J. Mass Spectrom. 2006, 41, 670–684. [Google Scholar] [CrossRef]
- Mao, S.J.; Jin, H.; Bi, Y.Q.; Liang, Z.; Li, H.; Hou, S.X. Ion-pair reversed-phase HPLC method for determination of sodium tanshinone IIA sulfonate in biological samples and its pharmacokinetics and biodistribution in mice. Chem. Pharm. Bull. 2007, 55, 753–756. [Google Scholar] [CrossRef]
- Sun, J.H.; Yang, M.; Han, J.; Wang, B.; Ma, X.; Xu, M.; Liu, P.; Guo, D.A. Profiling the metabolic difference of seven tanshinones using high-performance liquid chromatography/multi-stage mass spectrometry with data-dependent acquisition. Rapid Commun. Mass Spectrom. 2007, 21, 2211–2226. [Google Scholar]
- Hao, H.P.; Wang, G.J.; Li, P.; Li, J.; Ding, Z. Simultaneous quantification of cryptotanshinone and its active metabolite tanshinone IIA in plasma by liquid chromatography/tandem mass spectrometry (LC-MS/MS). J. Pharm. Biomed. Anal. 2006, 40, 382–388. [Google Scholar] [CrossRef]
- Wang, M.; Dai, H.; Li, X.; Li, Y.; Wang, L.; Xue, M. Structural elucidation of metabolites of tanshinone I and its analogue dihydrotanshinone I in rats by HPLC-ESI-MSn. J. Chromatogr. Analyt. Technol. Biomed. Life Sci. 2010, 878, 915–924. [Google Scholar] [CrossRef]
- Xue, M.; Cui, Y.; Wang, H.Q.; Luo, Y.J.; Zhang, B.; Zhou, Z.T. Pharmacokinetics of cryptotanshinone and its metabolite in pigs. Acta Pharm. Sin. 1999, 34, 81–84. [Google Scholar]
- Sun, J.H.; Yang, M.; Wang, X.M.; Xua, M.; Liu, A.H.; Guo, D.A. Identification of tanshinones and their metabolites in rat bile after oral administration of TTE-50, a standardized extract of Salvia miltiorrhiza by HPLC-ESI-DAD-MSn. J. Pharm. Biomed. Anal. 2007, 44, 564–574. [Google Scholar] [CrossRef]
- Li, Z.H.; Alex, D.; Siu, S.O.; Chu, I.K.; Renn, J.; Winkler, C.; Lou, S.; Tsui, S.K.; Zhao, H.Y.; Yan, W.R.; et al. Combined in vivo imaging and omics approaches reveal metabolism of icaritin and its glycosides in zebrafish larvae. Mol. BioSyst. 2011, 7, 2128–2138. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds tanshinone IIA, cryptotanshinone and tanshinone I are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wei, Y.; Li, P.; Wang, C.; Peng, Y.; Shu, L.; Jia, X.; Ma, W.; Wang, B. Metabolism of Tanshinone IIA, Cryptotanshinone and Tanshinone I from Radix Salvia Miltiorrhiza in Zebrafish. Molecules 2012, 17, 8617-8632. https://doi.org/10.3390/molecules17078617
Wei Y, Li P, Wang C, Peng Y, Shu L, Jia X, Ma W, Wang B. Metabolism of Tanshinone IIA, Cryptotanshinone and Tanshinone I from Radix Salvia Miltiorrhiza in Zebrafish. Molecules. 2012; 17(7):8617-8632. https://doi.org/10.3390/molecules17078617
Chicago/Turabian StyleWei, Yingjie, Ping Li, Changmei Wang, Yunru Peng, Luan Shu, Xiaobin Jia, Wenquan Ma, and Bing Wang. 2012. "Metabolism of Tanshinone IIA, Cryptotanshinone and Tanshinone I from Radix Salvia Miltiorrhiza in Zebrafish" Molecules 17, no. 7: 8617-8632. https://doi.org/10.3390/molecules17078617