Metabolic Profiling of Lactococcus lactis Under Different Culture Conditions

Abstract
:1. Introduction
2. Results and Discussion
2.1. Homolactic and Mixed-Acid Fermentation

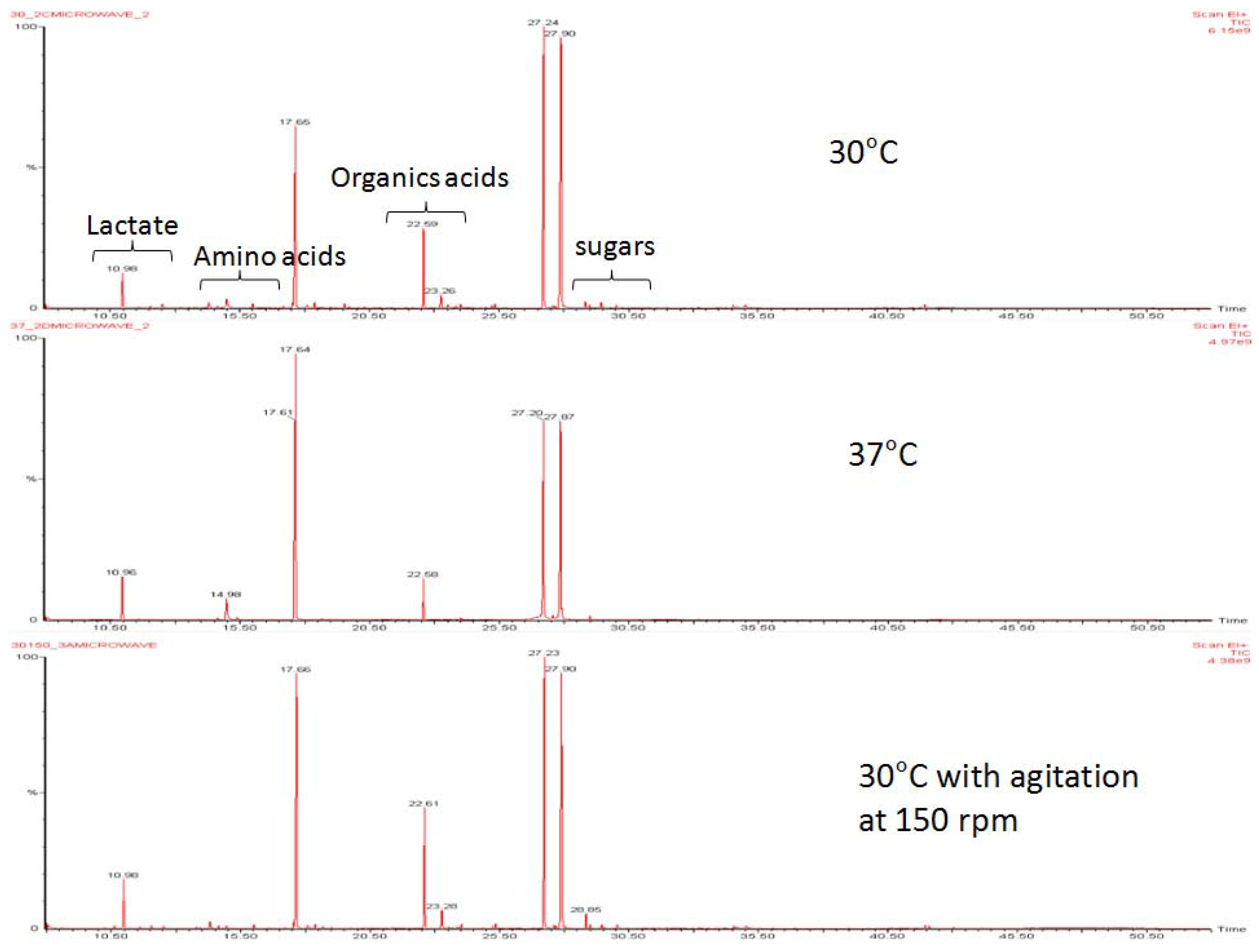{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 30 °C | Groups | 37 °C | Groups | 30 °C agitated | Groups |
|---|---|---|---|---|---|
| Alanine | A.A | Proline | A.A | Phenylalanine | A.A |
| Leucine | A.A | Leucine | A.A | Leucine | A.A |
| Norleucine | A.A | Phenylalanine | A.A | Serine | A.A |
| Phenylalanine | A.A | 1,3-Propanediol | Alcohol | Acetyl-L-Lysine | A.A |
| Threonine | A.A | 1,2-Butanediol | Alcohol | Aspartic acid | A.A |
| Glycine | A.A | Propanol | Alcohol | Threonine | A.A |
| Ethanol | Alcohol | Decanoic acid | F.A | Glutamine | A.A |
| Propanol | Alcohol | Picolinic acid | F.A | Propanol | Alcohol |
| Acetone | Ketone | Steric acid | F.A | 2-Propanol | Alcohol |
| 2-butanone | Ketone | Hexanoic acid | F.A | 1,2-Benzenediol | Alcohol |
| 2-heptanone | Ketone | Hexadecanoic acid | F.A | Isoleucine | Alcohol |
| Citric acid | TCA | Acetone | Ketone | 1,2-Octanediol | Alcohol |
| Fumaric acid | TCA | 2-Pentanone | Ketone | Cadaverine | Biogenic |
| Acetic acid | O.A | Oxalacetic acid | TCA | Palmitic acid | F.A |
| Aminobutyric acid | O.A | Lactate | O.A | Hexadecanoic acid | F.A |
| Hexanoic acid | O.A | 3-Hydroxybutyric acid | O.A | 2,4-Dihydroxyacetophenone | Ketone |
| Lactate | O.A | Mercaptoacetic acid | O.A | Heptadecane | Ketone |
| Mercaptoacetic acid | O.A | Acetic acid | O.A | 2-Pentanone | Ketone |
| Pentadecanoic acid | O.A | Butanoic acid | O.A | Ethanone | Ketone |
| Propanoic acid | O.A | Octadecanoic acid | O.A | Succinic acid | TCA |
| Glycerin | Polyol | Pentadecanoic acid | O.A | Malic acid | TCA |
| Glycerol | Polyol | Hexanoic acid | O.A | Lactate | O.A |
| Uracil | Pyrimidine | 3-Hydroxypyruvic acid | O.A | 2-Propenoic acid | O.A |
| Mannose | Sugar | Ascorbic acid | O.A | Mercaptoacetic acid | O.A |
| Ribose | Sugar | Acetic acid | O.A | 3-Hydroxypyruvic acid | O.A |
| Xylose | Sugar | Glycerin | Polyol | Ascorbic acid | O.A |
| Ethanethiol | Sulfur | Glycerol | Polyol | Acetic acid | O.A |
| Glyceraldehyde | Ribitol | Polyol | Butanoic acid | O.A | |
| Methanethiol | Sulfurs | Phthalic acid | O.A | ||
| 2-Hydroxyisophthalic acid | O.A | ||||
| Phosphoric acid | O.A | ||||
| Phenylacetic acid | O.A | ||||
| Benzoic acid | O.A | ||||
| Mandelic acid | O.A | ||||
| Oxalic acid | O.A | ||||
| Malonic acid | O.A | ||||
| Hexanoic acid | O.A | ||||
| 2-Butenedioic acid | O.A | ||||
| Aminomalonic acid | O.A | ||||
| Pentanoic acid | O.A |
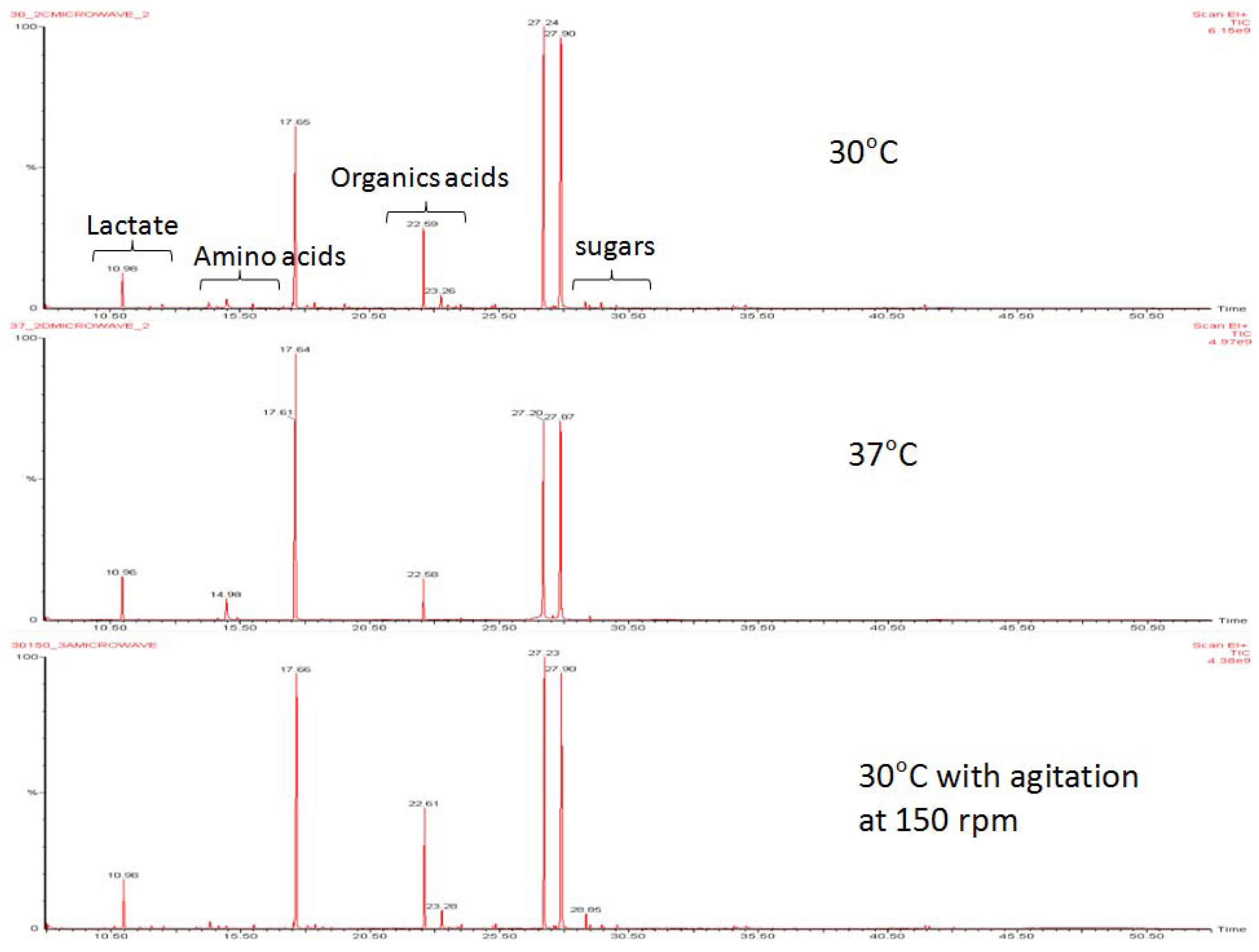
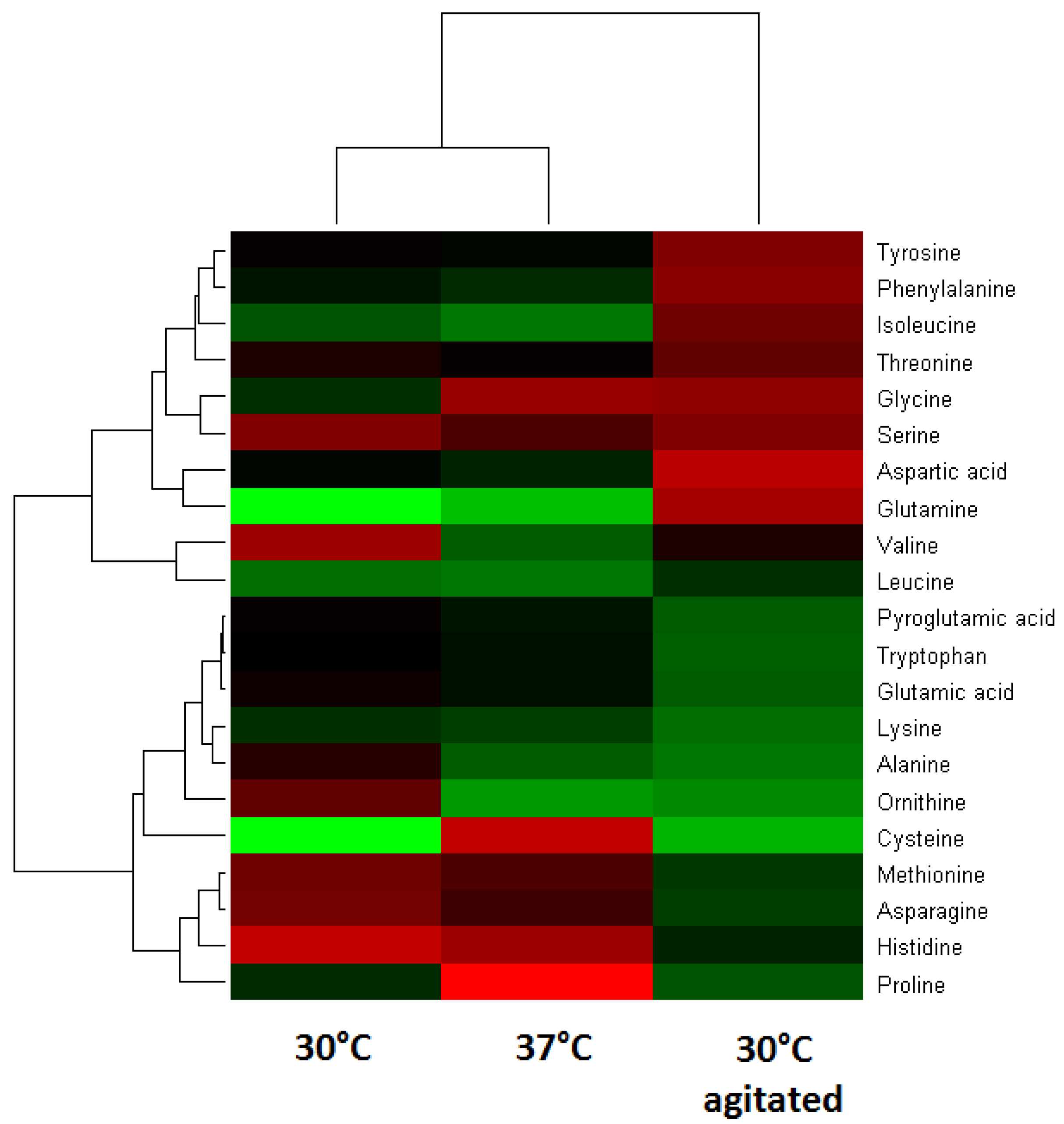
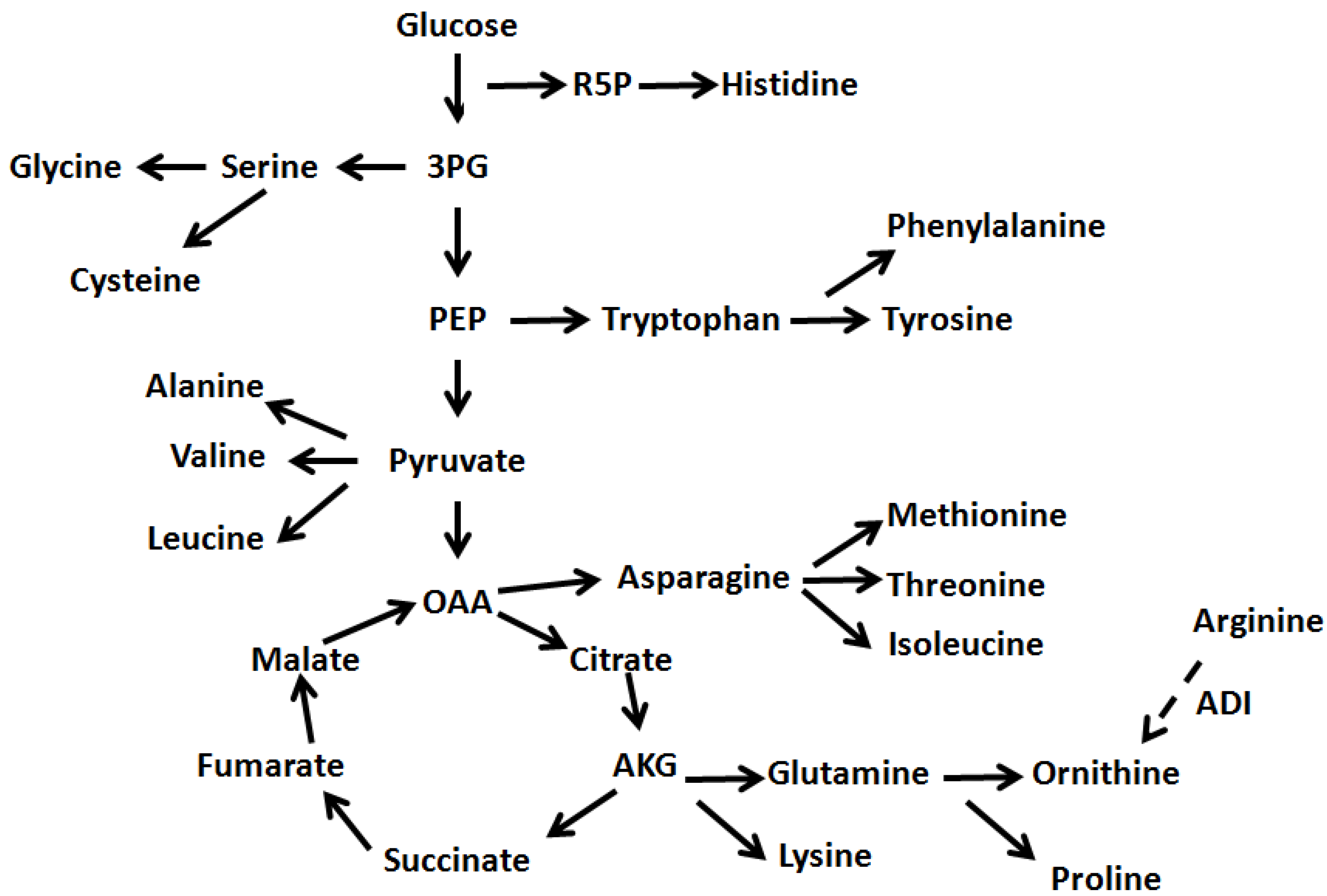
2.2. Changes of Amino Acids and Fermentation End Products

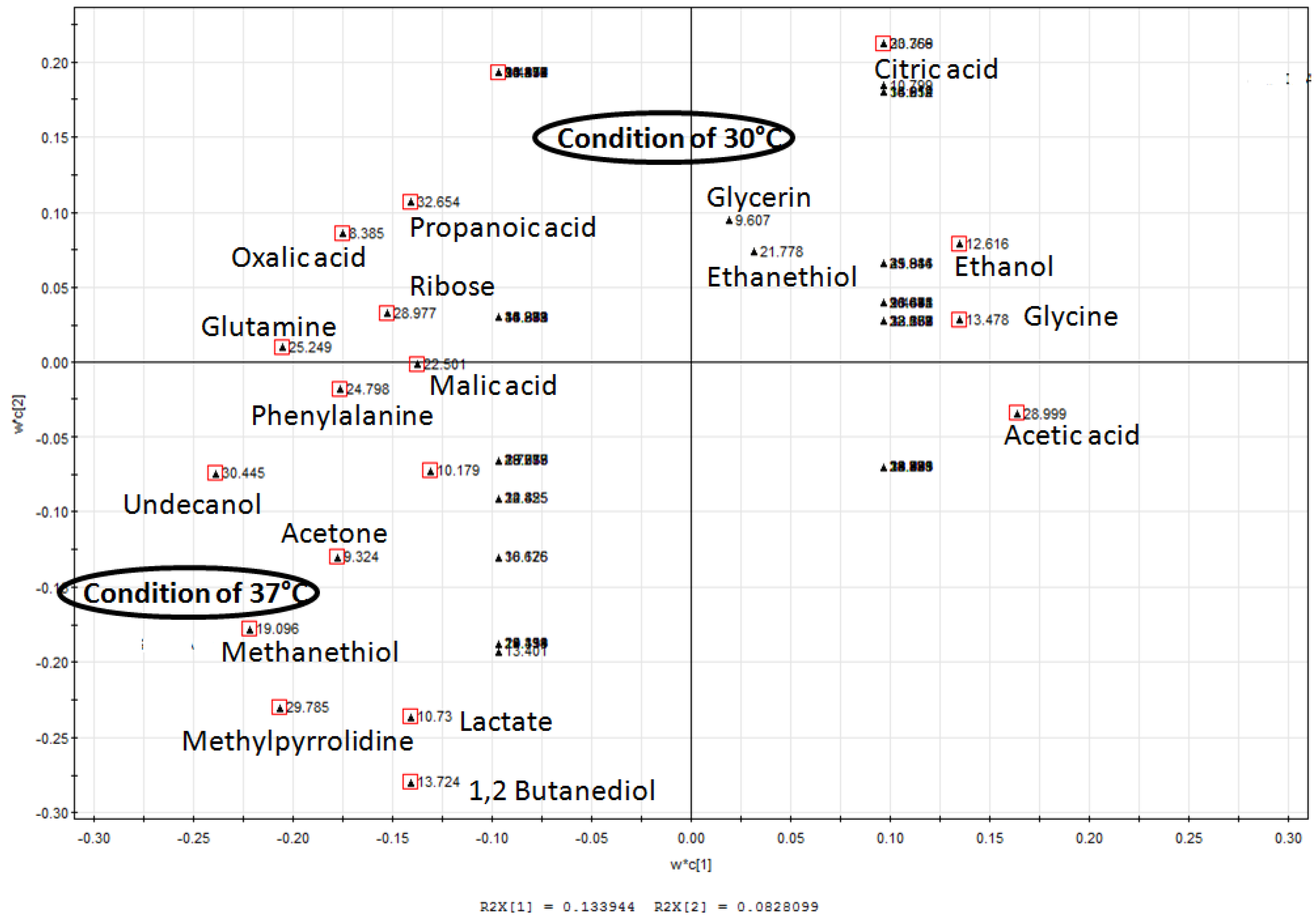
2.3. Changes of Amino Acids in Response to Temperature and Agitation

2.4. Comparison of Metabolites Detected Using TMS, MCF and HS Analysis
| Organic acids | Amino acids | Fatty acid |
|---|---|---|
| 2-aminobutyric acid | Alanine | Caprylate |
| 4-aminobutyric acid | Asparagine | 10,12-Otacedecadienoate |
| Caproic acid | Aspartic acid | |
| Citraconate | Cysteine | |
| Fumaric acids | Glutamic acid | |
| Glutaric acid | Glycine | |
| Glyceric acid | Histidine | |
| Hydroxybenzoate | Isoleucine | |
| Itaconic acid | Leucine | |
| Lactate | Lysine | |
| Malic acid | Methionine | |
| Malonic acid | Ornithine | |
| Nicotinic acid | Proline | |
| Oxalacetic acid | Threonine | |
| Oxalic acid | Tryptophan | |
| Succinic acid | Valine |
| Groups of metabolites | TMS | MCF |
|---|---|---|
| Alcohols | Yes | No |
| Aldehydes | Yes | No |
| Amines | Yes | No |
| Amino acids | Yes | Yes |
| Fatty acids | Yes | Yes |
| Ketones | Yes | No |
| Organic acids | Yes | Yes |
| Sugars | Yes | No |
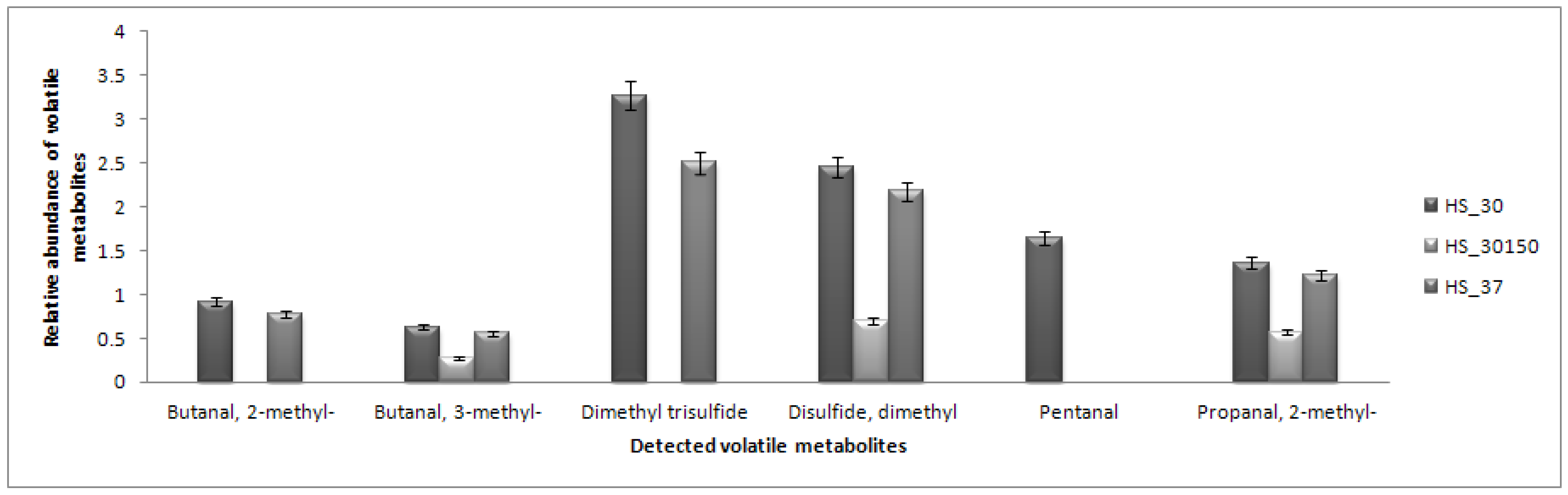
2.5. Headspace (HS) Analysis
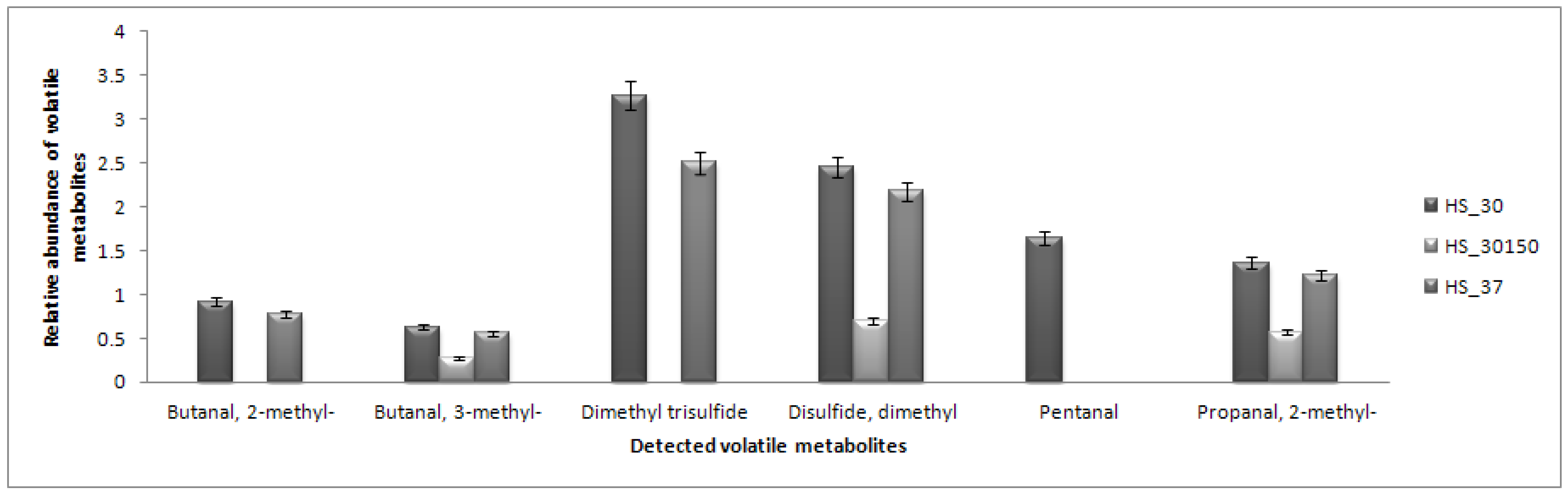
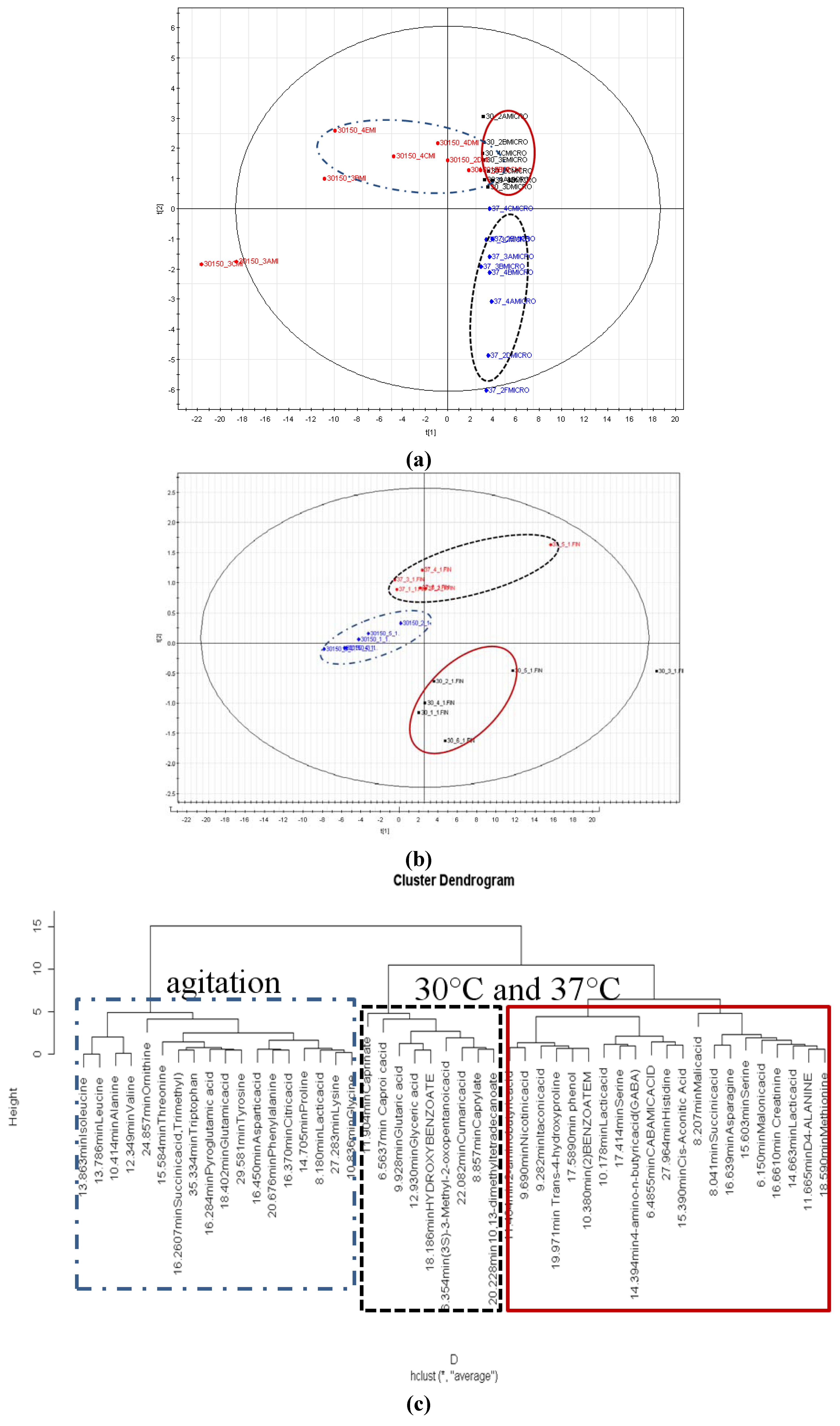
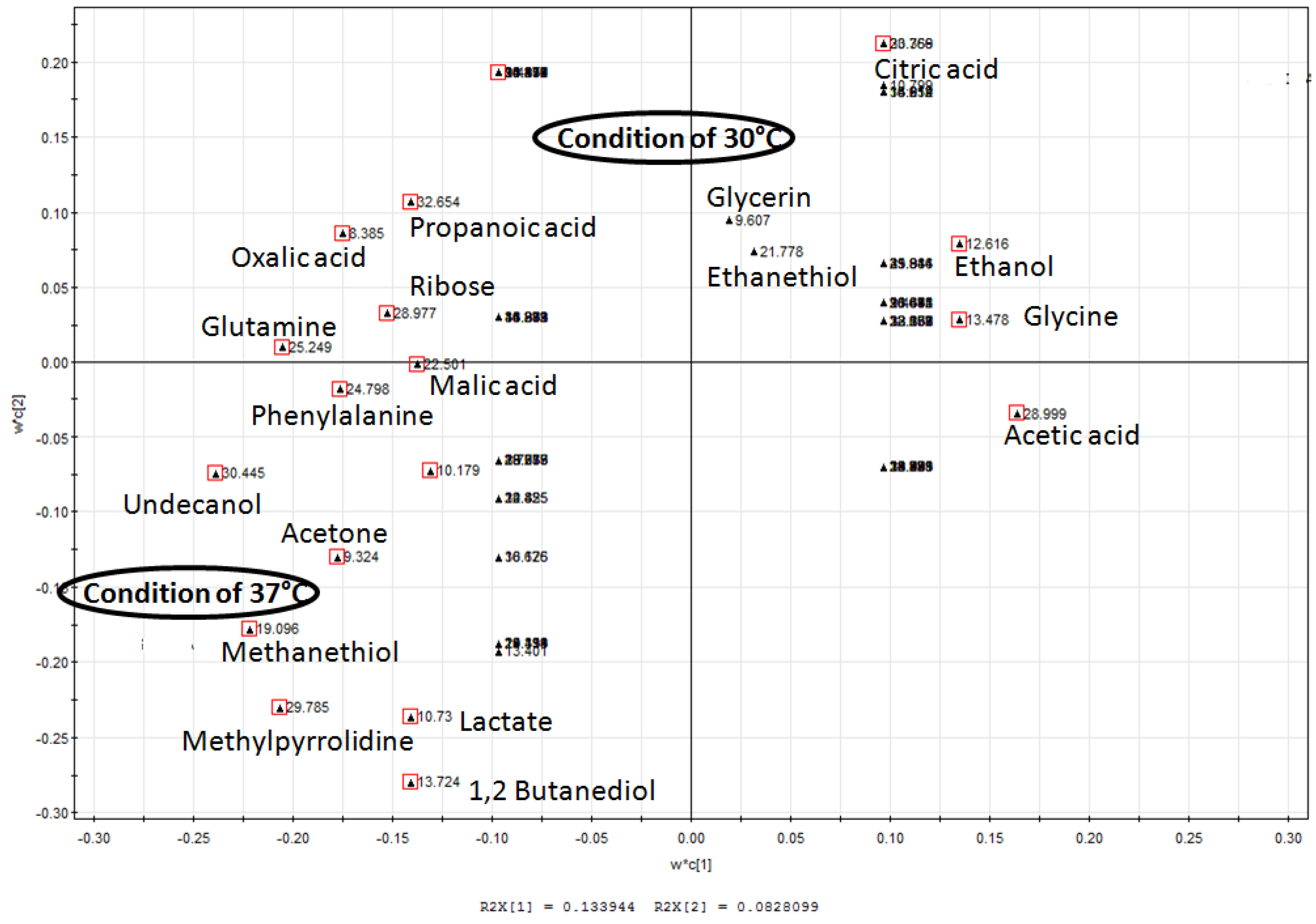
2.6. Partial Least Square Discriminant Analysis
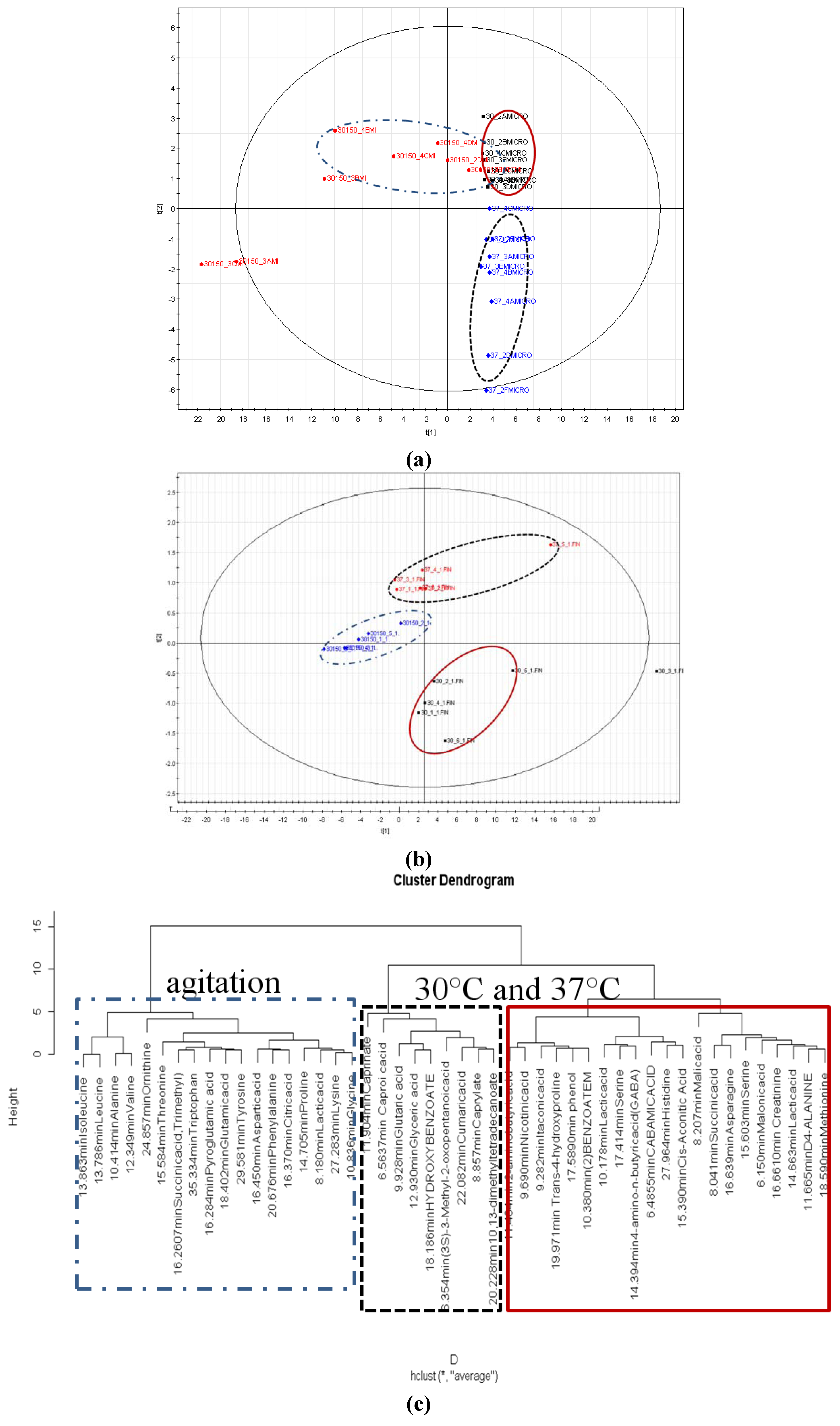
3. Experimental
3.1. Chemicals
3.2. Microbial Cultivation
3.3. Growth Estimation
3.4. Extraction of Extracellular Samples
3.5. Sample Derivatization Using TMS
3.6. Sample Derivatization Using MCF
3.7. Sample Preparation for Headspace Analysis (HS)
3.8. GC-MS Parameter for Samples Prepared by TMS
3.9. GC-MS Parameters for Samples Prepared by MCF
3.10. GC-MS Parameters for Headspace Analysis (HS)
3.11. Data Analysis and Validation
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Losiane, E.G.; Sylvain, M. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb. Cell Fact. 2011, 10, S20. [Google Scholar] [CrossRef]
- Bermúdez-Humarán, L.G.; Kharrat, P.; Chatel, J.-M.; Langella, P. Lactococci and lactobacilli as muscosal delivery vectors for therapeutic proteins and DNA vaccines. Microb. Cell Fact. 2011, 10, S4. [Google Scholar] [CrossRef]
- Mireille, Y.; Chistophe, G.; Emilie, C.; Gaelle, B.; Veronique, M. The initial efficiency of the proteolytic systems of Lactococcus lactis strains determines their responses to a cheese environment. Int. Dairy J. 2011, 21, 335–345. [Google Scholar]
- Guimont, C. Change of free amino acids in M17 medium after growth of Streptococcus thermophilus and identification of a glutamine transport ATP-binding protein. Int. Dairy J. 2002, 12, 729–736. [Google Scholar] [CrossRef]
- Amel, T.; Nassra, D.; Maryse, L.; Denis, R.; Gisele, L. Comparative transcriptome analysis of Lactococcus lactis subsp. cremoris strains under conditions simulating Cheddar cheese manufacture. 2011, 146, 263–275. [Google Scholar]
- Ziadi, M.; Bergot, G.; Courtin, P.; Chambellon, E.; Hamdi, M.; Yvon, M. Amino acid catabolism by Lactococcus lactis during milk fermentation. Int. Dairy J. 2010, 20, 5–31. [Google Scholar]
- Ayad, E.H.E.; Verheul, A.; de Jong, C.; Wouters, J.T.M.; Smit, G. Flavour forming abilities and amino acid requirements of Lactococcus lactis strains isolated from artisanal and non-dairy origin. Int. Dairy J. 1999, 19, 725–735. [Google Scholar]
- Clementine, D.; Emma, R.; Christophe, G.; Pascal, L.; Veronique, M.; Muriel, C.B. Investigation of the adaptation of Lactococcus lactis to isoleucine starvation integrating dynamic transcriptome and proteome information. Microb. Cell Fact. 2011, 10, S18. [Google Scholar] [CrossRef]
- Grade, S.; Carbonell, M.; Fernandez-grarcia, E.; Medina, M.; Nunez, M. Volatile compounds in Hispanico Cheese manufactured using a mesophilic starter a thermophilic starter and bacteriocin-producing Lactococcus lactis subps. cremoris INIA415. J. Agric. Food Chem. 2002, 50, 6752–6757. [Google Scholar]
- Zhu, D.; Zhou, X.; Jin, Y.Y. Metabolome profiling reveals adaptive evolution of S. Cerevisiae during repeated vacuum fermentations. Metabolomics 2010, 870, 222–232. [Google Scholar]
- Villas-Boas, S.G.; Delicado, S.G.; Akesson, M.; Nielsen, J. Simultaneous analysis of amino and nonamino organic acids as methyl chloroformate derivatives using gas chromatography-mass spectrometry. Anal. Biochem. 2003, 322, 134–138. [Google Scholar]
- Cheng, W.P.; Yang, X.Y.; Hegeman, A.D.; Gray, W.M.; Cohen, J.D. Microscale analysis of amino acids using gas chromatography-mass spectrometry after methyl chloroformate derivatization. J. Chromatogr. B 2010, 878, 2199–2208. [Google Scholar] [CrossRef]
- Liebeke, M.; Wunder, A.; Lalk, M. A rapid microwave-assisted derivatization of bacterial metabolome samples for gas chromatography/mass spectrometry analysis. Anal. Biochem. 2009, 401, 312–314. [Google Scholar]
- Urancken, G.; Rimaux, T.; Wouters, D.; Leny, F.; De Vuyst, L. The arginine deiminase pathway of L. fermentum IMDO 13010 response to growth under stress conditions of both temperature and salt. Food Microbiol. 2009, 7, 720–727. [Google Scholar]
- Dong, X.; Quinn, P.J.; Wang, X. Metabolic engineering of Escherichia coli glutamicum for the production of L- threonine. Biotechnol. Adv. 2011, 29, 11–23. [Google Scholar] [CrossRef]
- Brice, S.; Patrice, P.; Dusko, S.E.; Pierre, R.; Eric, G. Sulfur amino acid metabolism and its control in L. lactis IL1403. J. Bacteriol. 2005, 11, 3762–3778. [Google Scholar]
- Beale, D.J.; Dunn, M.S.; Morrison, P.D.; Porter, N.A.; Marlow, D.R. Characterisation of bulk water samples from copper pipes undergoing microbially influenced corrosion by diagnostic metabolomic profiling. Corros. Sci. 2012, 55, 272–279. [Google Scholar] [CrossRef]
- Kouremenos, K.A.; Harynuk, J.J.; Winniford, W.L.; Morrison, P.D.; Marriott, P.J. One-pot microwave derivatization of target compounds relevent to metabolomics with comprehensive two-dimensional gas chromatography. J. Chromatogr. B 2010, 878, 1761–1770. [Google Scholar]
- Chu, T.Y.; Chang, C.H.; Liao, Y.C.; Chen, Y.C. Microwave-accelerated derivatization processes for the determination of phenolic acids by gas chromatography-mass spectrometry. Talanta 2001, 54, 1163–1171. [Google Scholar]
- Ranz, A.; Maier, E.; Motter, H.; Lankmayr, E. Extraction and derivatization of polar herbicides for GC-MS analyses. J. Sep. Sci. 2008, 31, 3021–3029. [Google Scholar] [CrossRef]
- Bowden, J.A.; Dominic, M.C.; Whitney, L.S.; Diana, C.M.M.; Timothy, J.G.; Richard, A.Y. Enhanced Analysis of Steroids by Gas Chromatography/Mass Spectrometry using Microwave-Accelerated Derivatization. Anal. Chem. 2009, 16, 6725–6734. [Google Scholar]
- Beale, D.J.; Michael, S.D.; Donavan, M. Application of GC-MS metabolic profiling to “blue-green water” from microbial influenced corrosion in copper pipes. Corros. Sci. 2010, 52, 3140–3145. [Google Scholar]
- Sandra, L.S.; Markus, D.; Oliver Kappe, C. Microwave-assisted derivatization procedures for gas chromatography/mass spectrometry analysis. Mol. Divers. 2010, 14, 869–888. [Google Scholar]
- Ruiz-Matute, A.I.; Hernandez-Hernandez, O.; Rodriguez-Sanchez, S.; Sanz, M.L.; Martinez-Castro, I. Derivatization of carbohydrates for GC and GC-MS analyses. J. Chromatogr. B 2011, 879, 1226–1240. [Google Scholar]
- Villas-Boas, S.G.; Noel, S.; Lane, G.A.; Attwood, G.; Cookson, A. Extracellular metabolomics: A metabolic footprinting approach to assess fiber degradation in complex media. Anal. Biochem. 2006, 349, 297–305. [Google Scholar]
- Husek, P.; Matucha, P.; Vrankova, A.; Simek, P. Simple plasma work-up for a fast chromatographic analysis of homocysteine, cysteine, methionine and aromatic amino acids. J. Chromatogr. B 2003, 789, 311–322. [Google Scholar] [CrossRef]
- Smart, K.F.; Aggio, R.M.B.; Houtte, J.R.V.; Villas-Boas, S.G. Analytical platform for metabolome analysis of microbial cells using methyl chloroformate derivatization followed by gas chromatography-mass spectrometry. Nat. Protocol 2010, 5, 1709–1729. [Google Scholar]
- Yvon, M.; Rijnen, L. Cheese flavour formation by amino acid catabolism. Int. Dairy J. 2001, 11, 185–201. [Google Scholar] [CrossRef]
- Marilley, L.; Casey, M.G. Flavours of cheese products: Metabolic pathways analytical tool and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef]
- Terzaghi, B.E.; Sandine, W.E. Improved medium for lactic streptococci and their bacteriophages. App. Microbiol. 1975, 29, 807–813. [Google Scholar]
- Roessner, U.; Wagner, C.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Simultaneous analysis of metabolites in potato tuber by gas chromatography-mass spectrometry. Plant J. 2010, 23, 131–142. [Google Scholar]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, 652–660. [Google Scholar]
- Sample Availability: Samples of the metabolites lists are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azizan, K.A.; Baharum, S.N.; Mohd Noor, N. Metabolic Profiling of Lactococcus lactis Under Different Culture Conditions. Molecules 2012, 17, 8022-8036. https://doi.org/10.3390/molecules17078022
Azizan KA, Baharum SN, Mohd Noor N. Metabolic Profiling of Lactococcus lactis Under Different Culture Conditions. Molecules. 2012; 17(7):8022-8036. https://doi.org/10.3390/molecules17078022
Chicago/Turabian StyleAzizan, Kamalrul Azlan, Syarul Nataqain Baharum, and Normah Mohd Noor. 2012. "Metabolic Profiling of Lactococcus lactis Under Different Culture Conditions" Molecules 17, no. 7: 8022-8036. https://doi.org/10.3390/molecules17078022
APA StyleAzizan, K. A., Baharum, S. N., & Mohd Noor, N. (2012). Metabolic Profiling of Lactococcus lactis Under Different Culture Conditions. Molecules, 17(7), 8022-8036. https://doi.org/10.3390/molecules17078022