Potent Anti-HIV Activities and Mechanisms of Action of a Pine Cone Extract from Pinus yunnanensis

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
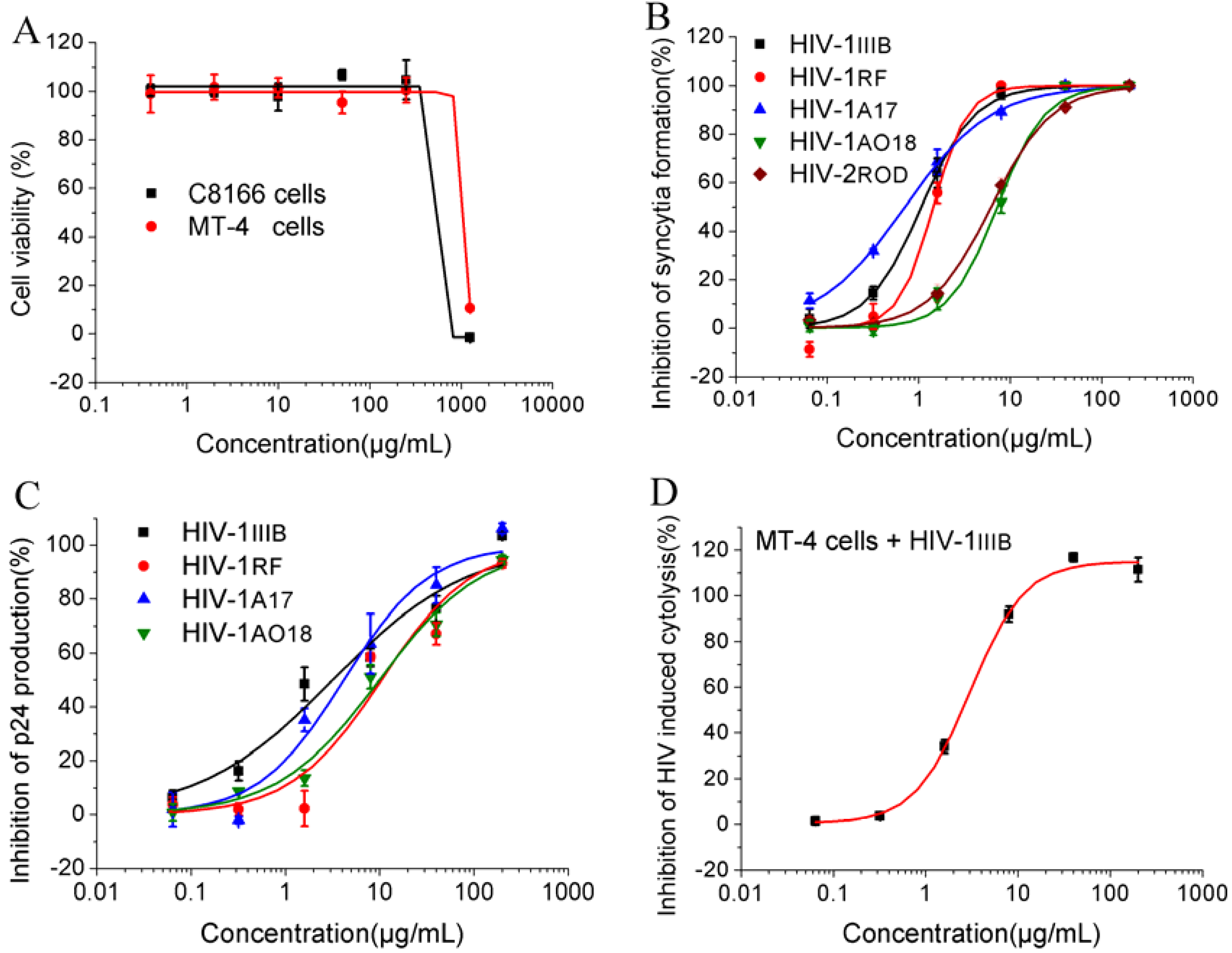
2.1. Cytotoxities and Anti-HIV Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
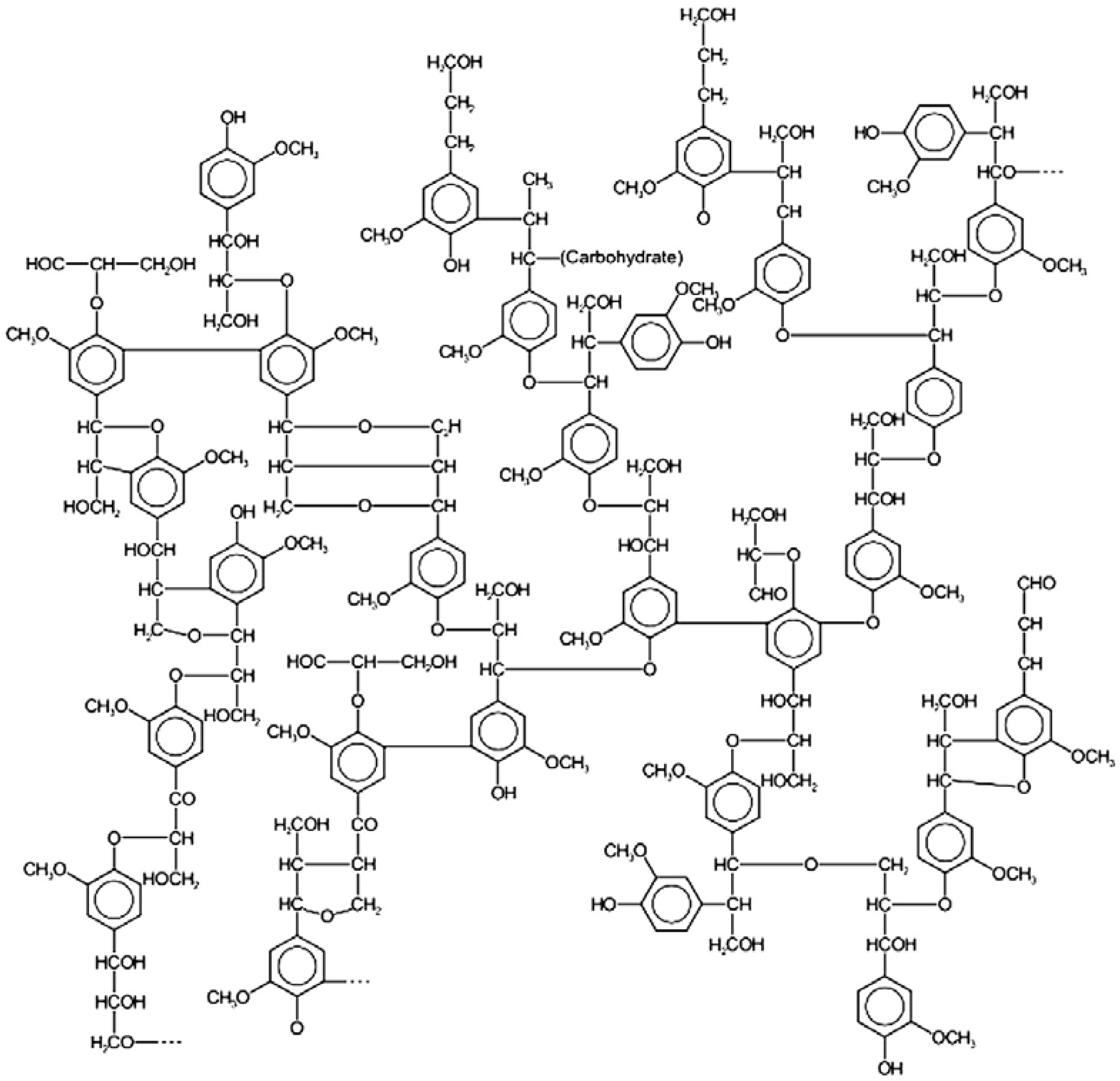{kind=link}
| Samples | Cells | HIV strains/enzyme | Assay | CC50 (μg/mL) | EC50 (μg/mL) | SI | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| YNS-PY-F | C8166 | HIV-1IIIB | CPE | 438.73 | 0.96 | 457.01 | ||||||
| p24 | 3.98 | 110.23 | ||||||||||
| HIV-1RF | CPE | 1.53 | 286.75 | |||||||||
| p24 | 9.48 | 46.28 | ||||||||||
| HIV-1A17 | CPE | 0.88 | 498.56 | |||||||||
| p24 | 4.04 | 108.60 | ||||||||||
| HIV-1AO18 | CPE | 7.20 | 60.93 | |||||||||
| p24 | 9.79 | 44.81 | ||||||||||
| HIV-2ROD | CPE | 6.17 | 71.11 | |||||||||
| H9/HIV-1IIIB | CPE | 7.60 | 57.73 | |||||||||
| MT-4 | HIV-1IIIB | MTT | 922.47 | 2.22 | 415.53 | |||||||
| — | RT | ELISA | — | 4.60 | — | |||||||
| AZT | C8166 | HIV-1IIIB | CPE | 1120.84 | 1.30 a | 862,184.62 | ||||||
| p24 | 1.94 a | 577,752.58 | ||||||||||
| HIV-1RF | CPE | 2.95 a | 379,945.76 | |||||||||
| p24 | 6.48 a | 172,969.14 | ||||||||||
| HIV-1A17 | CPE | 4.22 a | 265,601.90 | |||||||||
| p24 | 4.11 a | 272,710.46 | ||||||||||
| HIV-2ROD | CPE | 3.07 a | 365,094.46 | |||||||||
| MT-4 | HIV-1IIIB | MTT | 358.50 | 0.85 a | 421,764.71 | |||||||
| NVP | C8166 | HIV-1AO18 | CPE | — | 61.70 b | — | ||||||
| p24 | — | 42.11 b | — | |||||||||
| T20 | C8166 | H9/HIV-1IIIB | CPE | — | 14.96 a | — | ||||||
| PFA | — | RT | ELISA | — | 1.03 | — | ||||||

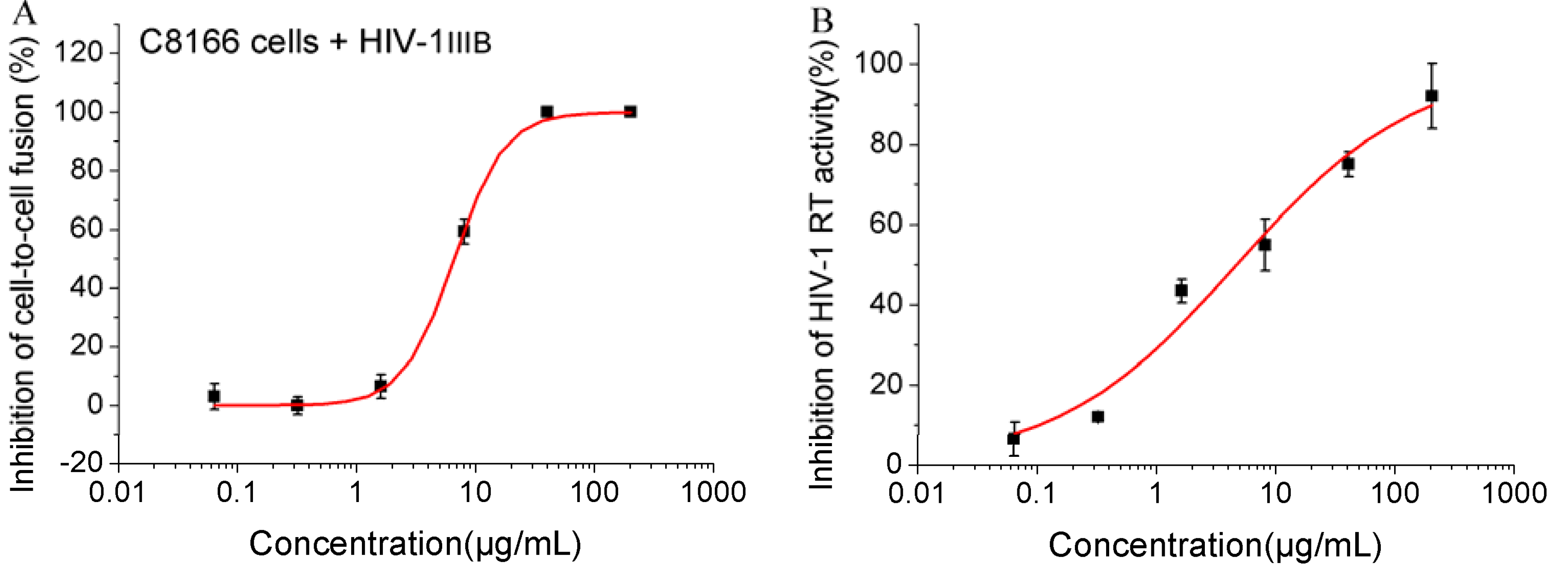
2.2. Inhibition on HIV-1 Fusion and Activities of Reverse Transcriptase

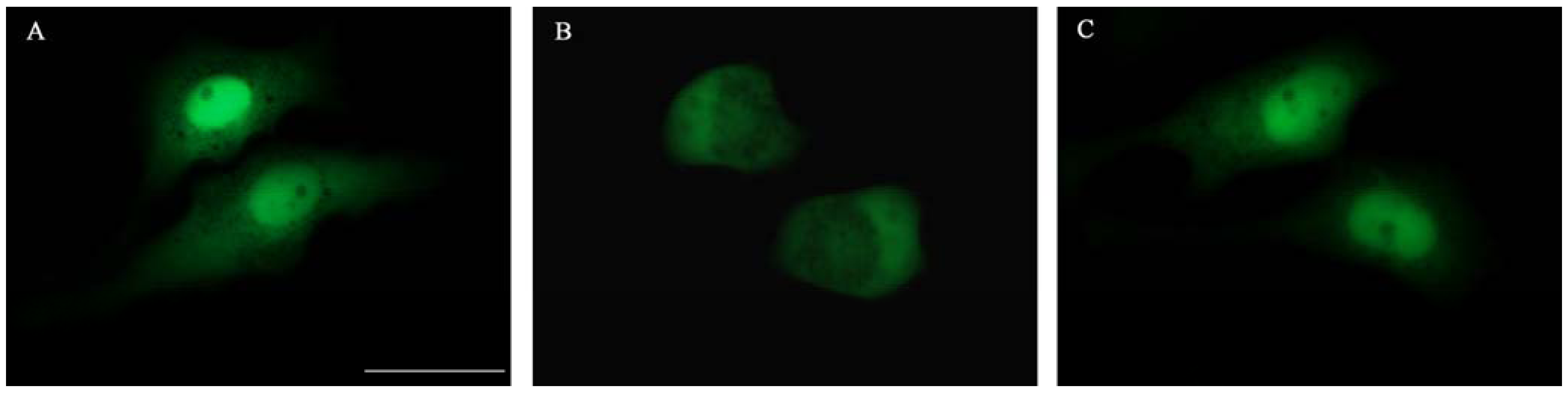
2.3. Effect on HIV-1 Integrase Nuclear Translocation

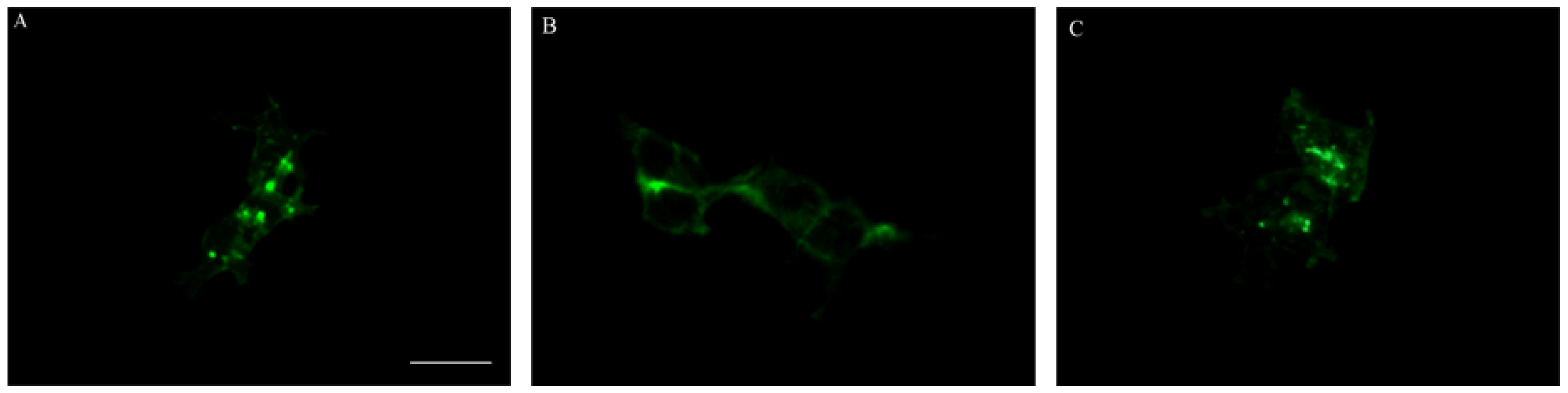
2.4. Effect on SDF-1α Induced CXCR4 Internalization

3. Experimental
3.1. Plant Materials and Extraction Procedure

3.2. Reagents and Chemicals
3.3. Plasmid, Cells and Viruses
3.4. Cytotoxicity Assay
3.5. Inhibition of YNS-PY-F on HIV Induced Cytopathic Effects (CPE)
3.6. Effect on HIV-1 Replication in Acute Infection
3.7. Effect on HIV-1IIIB Induced Cytolysis in MT-4 Cells
3.8. Effect on Cell-To-Cell Fusion
3.9. Effect on Recombinant HIV-1 Reverse Transcriptase (RT) Activity
3.10. Effect on HIV-1 Integrase (IN) Nuclear Translocation
3.11. Effect on SDF-1α Induced CXCR4 Internalization
4. Conclusions
Acknowledgments
References and Notes
- Adamson, C.S.; Freed, E.O. Novel approaches to inhibiting HIV-1 replication. Antiviral Res. 2010, 85, 119–141. [Google Scholar] [CrossRef]
- Cos, P.; Maes, L.; Vanden Berghe, D.; Hermans, N.; Pieters, L.; Vlietinck, A. Plant substances as anti-HIV agents selected according to their putative mechanism of action. J. Nat. Prod. 2004, 67, 284–293. [Google Scholar] [CrossRef]
- de Clercq, E. Current lead natural products for the chemotherapy of human immunodeficiency virus (HIV) infection. Med. Res. Rev. 2000, 20, 323–349. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, N.; Zheng, Y.T. Advances in the study of anti-HIV natural compounds derived from traditional Chinese medicines (in Chinese). Acta Pharm. Sin. 2010, 45, 141–153. [Google Scholar]
- Sakagami, H.; Kawazoe, Y.; Komatsu, N.; Simpson, A.; Nonoyama, M.; Konno, K.; Yoshida, T.; Kuroiwa, Y.; Tanuma, S. Antitumor, antiviral and immunopotentiating activities of pine cone extracts: Potential medicinal efficacy of natural and synthetic lignin-related materials (review). Anticancer Res. 1991, 11, 881–888. [Google Scholar]
- Lai, P.K.; Donovan, J.; Takayama, H.; Sakagami, H.; Tanaka, A.; Konno, K.; Nonoyama, M. Modification of human immunodeficiency viral replication by pine cone extracts. AIDS Res. Hum. Retroviruses 1990, 6, 205–217. [Google Scholar] [CrossRef]
- Tamura, Y.; Lai, P.K.; Bradley, W.G.; Konno, K.; Tanaka, A.; Nonoyama, M. A soluble factor induced by an extract from Pinus parviflora Sieb et Zucc can inhibit the replication of human-immunodeficiency-virus in vitro. Proc. Natl. Acad. Sci. USA 1991, 88, 2249–2253. [Google Scholar] [CrossRef]
- Eberhardt, T.L.; Young, R.A. Assessment of the anti-HIV activity of a pine cone isolate. Planta Med. 1996, 62, 63–65. [Google Scholar] [CrossRef]
- Satoh, K.; Kihara, T.; Ida, Y.; Sakagami, H.; Koyama, N.; Premanathan, M.; Arakaki, R.; Nakashima, H.; Komatsu, N.; Fujimaki, M.; et al. Radical modulation activity of pine cone extracts of Pinus elliottii var. Elliottii. Anticancer Res. 1999, 19, 357–364. [Google Scholar]
- Li, N.; Fu, L.K. Notes on gymnosperms. 1. Taxonomic treatments of some Chinese conifers. Novon 1997, 7, 261–264. [Google Scholar] [CrossRef]
- Young, R.C.; Friedman, M.A.; Schilsky, R.L.; Sigal, E.V. Drug safety and drug efficacy: Two sides of the same coin. Clin. Cancer Res. 2007, 13, 2533–2534. [Google Scholar] [CrossRef]
- Este, J.A. Virus entry as a target for anti-HIV intervention. Curr. Med. Chem. 2003, 10, 1617–1632. [Google Scholar] [CrossRef]
- Garcia-Sosa, A.T.; Sild, S.; Takkis, K.; Maran, U. Combined approach using ligand efficiency, cross-docking, and antitarget hits for wild-type and drug-resistant Y181C HIV-1 reverse transcriptase. J. Chem. Inf. Model. 2011, 51, 2595–2611. [Google Scholar] [CrossRef]
- Cichero, E.; Fossa, P. Docking-based 3D-QSAR analyses of pyrazole derivatives as HIV-1 non-nucleoside reverse transcriptase inhibitors. J. Mol. Model. 2012, 18, 1573–1582. [Google Scholar] [CrossRef]
- Mao, Y.; Li, Y.; Hao, M.; Zhang, S.; Ai, C. Docking, molecular dynamics and quantitative structure-activity relationship studies for HEPTs and DABOs as HIV-1 reverse transcriptase inhibitors. J. Mol. Model. 2012, 18, 2185–2198. [Google Scholar] [CrossRef]
- Sakagami, H.; Kushida, T.; Oizumi, T.; Nakashima, H.; Makino, T. Distribution of lignin-carbohydrate complex in plant kingdom and its functionality as alternative medicine. Pharmacol. Ther. 2010, 128, 91–105. [Google Scholar] [CrossRef]
- Zhou, P.; Lin, F.; Liu, J.; Lv, Y.J.; Li, H.Z.; Liu, G.M. Determination of carbohydrate content in lignin-carbohydrate complex from pine cone of Pinus yunnanensis Franch (in Chinese). Lishizhen Med. Mater Med. Res. 2011, 22, 1820–1821. [Google Scholar]
- Lesbats, P.; Botbol, Y.; Chevereau, G.; Vaillant, C.; Calmels, C.; Arneodo, A.; Andreola, M.L.; Lavigne, M.; Parissi, V. Functional coupling between HIV-1 Integrase and the SWI/SNF Chromatin remodeling complex for efficient in vitro integration into stable nucleosomes. PLoS Pathog. 2011, 7, e1001280. [Google Scholar] [CrossRef]
- Esposito, D.; Craigie, R. HIV integrase structure and function. Adv. Virus Res. 1999, 52, 319–333. [Google Scholar] [CrossRef]
- van Maele, B.; Debyser, Z. HIV-1 integrated: An interplay between HIV-1 integrase, cellular and cellular and viral. AIDS Rev. 2005, 7, 26–43. [Google Scholar]
- Maertens, G.; Cherepanov, P.; Pluymers, W.; Busschots, K.; de Clercq, E.; Debyser, Z.; Engelborghs, Y. LEDGF/p75 is essential for nuclear and chromosomal targeting of HIV-1 integrase in human cells. J. Biol. Chem. 2003, 278, 33528–33539. [Google Scholar]
- Levin, A.; Hayouka, Z.; Friedler, A.; Loyter, A. Transportin 3 and importin alpha are required for effective nuclear import of HIV-1 integrase in virus-infected cells. Nucleus 2010, 1, 422–431. [Google Scholar]
- Levin, A.; Armon-Omer, A.; Rosenbluh, J.; Melamed-Book, N.; Graessmann, A.; Waigmann, E.; Loyter, A. Inhibition of HIV-1 integrase nuclear import and replication by a peptide bearing integrase putative nuclear localization signal. Retrovirology 2009, 6, 112. [Google Scholar] [CrossRef]
- Du, L.; Chen, J.; Yang, L.M.; Zheng, Y.T.; Tang, Y.; Shen, X.; Jiang, H.L. D77, one benzoic acid derivative, functions as a novel anti-HIV-1 inhibitor targeting the interaction between integrase and cellular LEDGF/p75. Biochem. Biophys. Res. Commun. 2008, 375, 139–144. [Google Scholar] [CrossRef]
- Murakami, T.; Yamamoto, N. Role of CXCR4 in HIV infection and its potential as a therapeutic target. Future Microbiol. 2010, 5, 1025–1039. [Google Scholar] [CrossRef]
- Dong, C.Z.; Tian, S.; Madani, N.; Choi, W.T.; Kumar, S.; Liu, D.; Sodroski, J.G.; Huang, Z.; An, J. Role of CXCR4 internalization in the anti-HIV activity of stromal cell-derived factor-1alpha probed by a novel synthetically and modularly modified-chemokine analog. Exp. Biol. Med. (Maywood) 2011, 236, 1413–1419. [Google Scholar]
- Liu, G.J.; Wang, J.P.; Xiao, J.C.; Zhao, Z.W.; Zheng, Y.T. Preparation and characterization of three monoclonal antibodies against HIV-1 p24 capsid protein. Cell. Mol. Immunol. 2007, 4, 203–208. [Google Scholar]
- Wang, R.R.; Gu, Q.O.; Yang, L.M.; Chen, J.J.; Li, S.Y.; Zheng, Y.T. Anti-HIV-1 activities of extracts from the medicinal plant Rhus chinensis. J. Ethnopharmacol. 2006, 105, 269–273. [Google Scholar] [CrossRef]
- Wang, R.R.; Gu, Q.; Wang, Y.H.; Zhang, X.M.; Yang, L.M.; Zhou, J.; Chen, J.J.; Zheng, Y.T. Anti-HIV-1 activities of compounds isolated from the medicinal plant Rhus chinensis. J. Ethnopharmacol. 2008, 117, 249–256. [Google Scholar] [CrossRef]
- Zhang, X.J.; Yang, G.Y.; Wang, R.R.; Pu, J.X.; Sun, H.D.; Xiao, W.L.; Zheng, Y.T. 7,8-Secolignans from Schisandra wilsoniana and Their anti-HIV-1 activities. Chem. Biodivers. 2010, 7, 2692–2701. [Google Scholar] [CrossRef]
- Xiao, W.L.; Wang, R.R.; Zhao, W.; Tian, R.R.; Shang, S.Z.; Yang, L.M.; Yang, J.H.; Pu, J.X.; Zheng, Y.T.; Sun, H.D. Anti-HIV-1 activity of lignans from the fruits of Schisandra rubriflora. Arch. Pharm. Res. 2010, 33, 697–701. [Google Scholar] [CrossRef]
- Wang, Y.H.; Tang, J.G.; Wang, R.R.; Yang, L.M.; Dong, Z.J.; Du, L.; Shen, X.; Liu, J.K.; Zheng, Y.T. Flazinamide, a novel beta-carboline compound with anti-HIV actions. Biochem. Biophys. Res. Commun. 2007, 355, 1091–1095. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, Z.H.; Liu, J.K.; Zheng, Y.T. Xanthohumol, a novel anti-HIV-1 agent purified from Hops Humulus lupulus. Antiviral Res. 2004, 64, 189–194. [Google Scholar]
- Sample Availability: Samples of the pine cone extract (YNS-PY-F) from Pinus yunnanensis is available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.; Yang, L.-M.; Liu, G.-M.; Liu, Y.-J.; Zheng, C.-B.; Lv, Y.-J.; Li, H.-Z.; Zheng, Y.-T. Potent Anti-HIV Activities and Mechanisms of Action of a Pine Cone Extract from Pinus yunnanensis. Molecules 2012, 17, 6916-6929. https://doi.org/10.3390/molecules17066916
Zhang X, Yang L-M, Liu G-M, Liu Y-J, Zheng C-B, Lv Y-J, Li H-Z, Zheng Y-T. Potent Anti-HIV Activities and Mechanisms of Action of a Pine Cone Extract from Pinus yunnanensis. Molecules. 2012; 17(6):6916-6929. https://doi.org/10.3390/molecules17066916
Chicago/Turabian StyleZhang, Xuan, Liu-Meng Yang, Guang-Ming Liu, Ya-Juan Liu, Chang-Bo Zheng, Yong-Jun Lv, Hao-Zhi Li, and Yong-Tang Zheng. 2012. "Potent Anti-HIV Activities and Mechanisms of Action of a Pine Cone Extract from Pinus yunnanensis" Molecules 17, no. 6: 6916-6929. https://doi.org/10.3390/molecules17066916
APA StyleZhang, X., Yang, L.-M., Liu, G.-M., Liu, Y.-J., Zheng, C.-B., Lv, Y.-J., Li, H.-Z., & Zheng, Y.-T. (2012). Potent Anti-HIV Activities and Mechanisms of Action of a Pine Cone Extract from Pinus yunnanensis. Molecules, 17(6), 6916-6929. https://doi.org/10.3390/molecules17066916