Lectin from Canavalia brasiliensis Seeds (ConBr) Is a Valuable Biotechnological Tool to Stimulate the Growth of Rhizobium tropici in Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
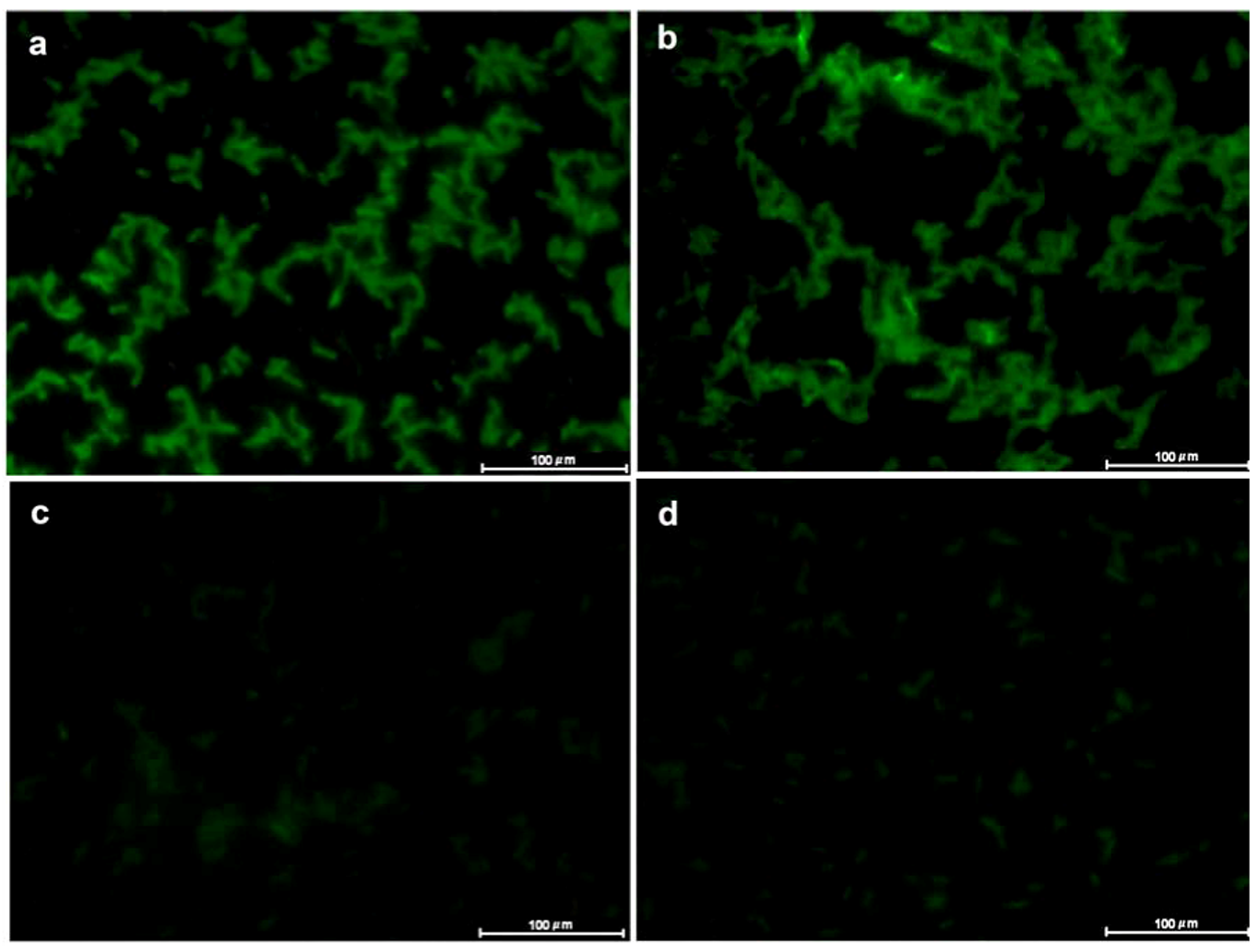
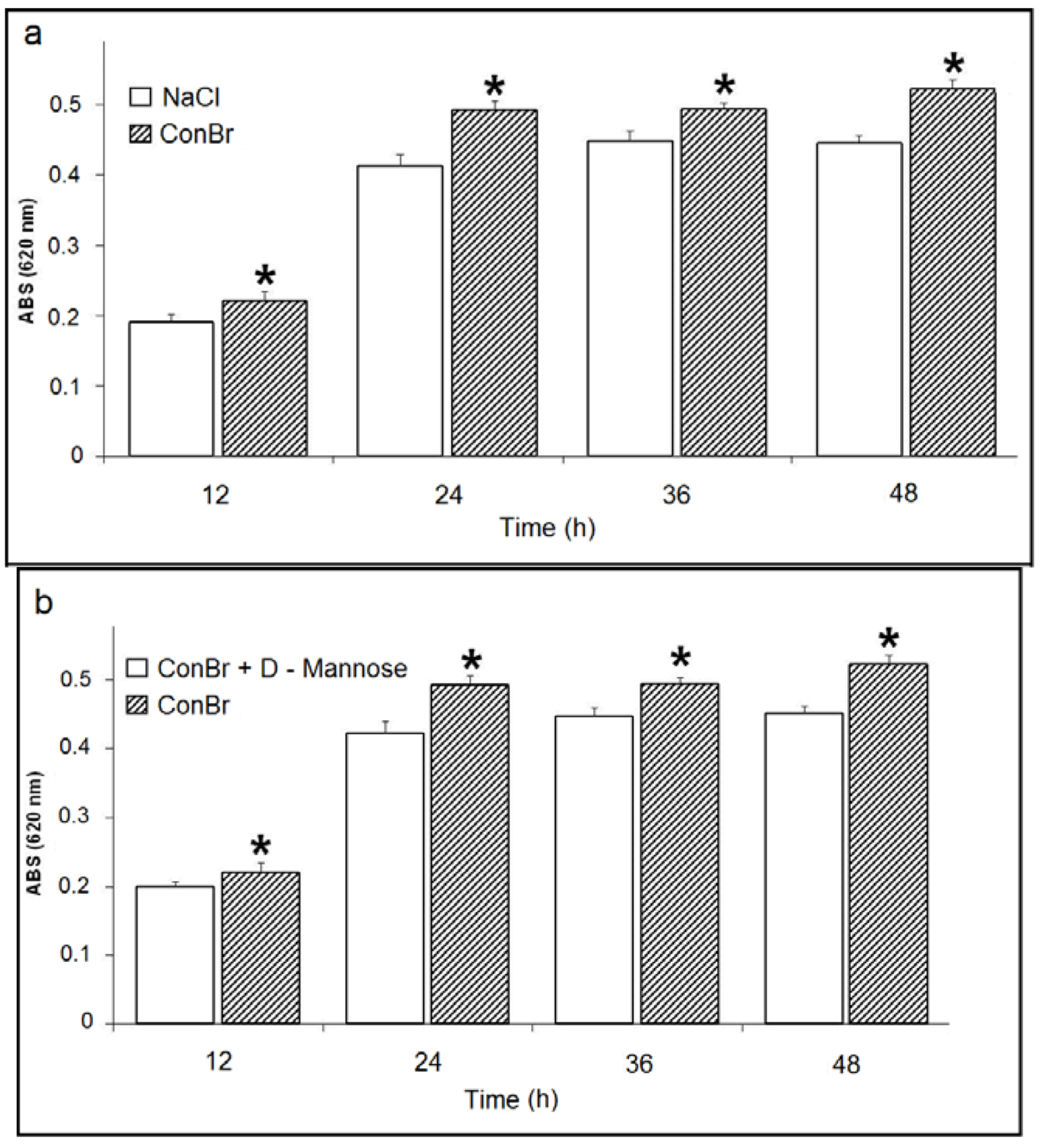
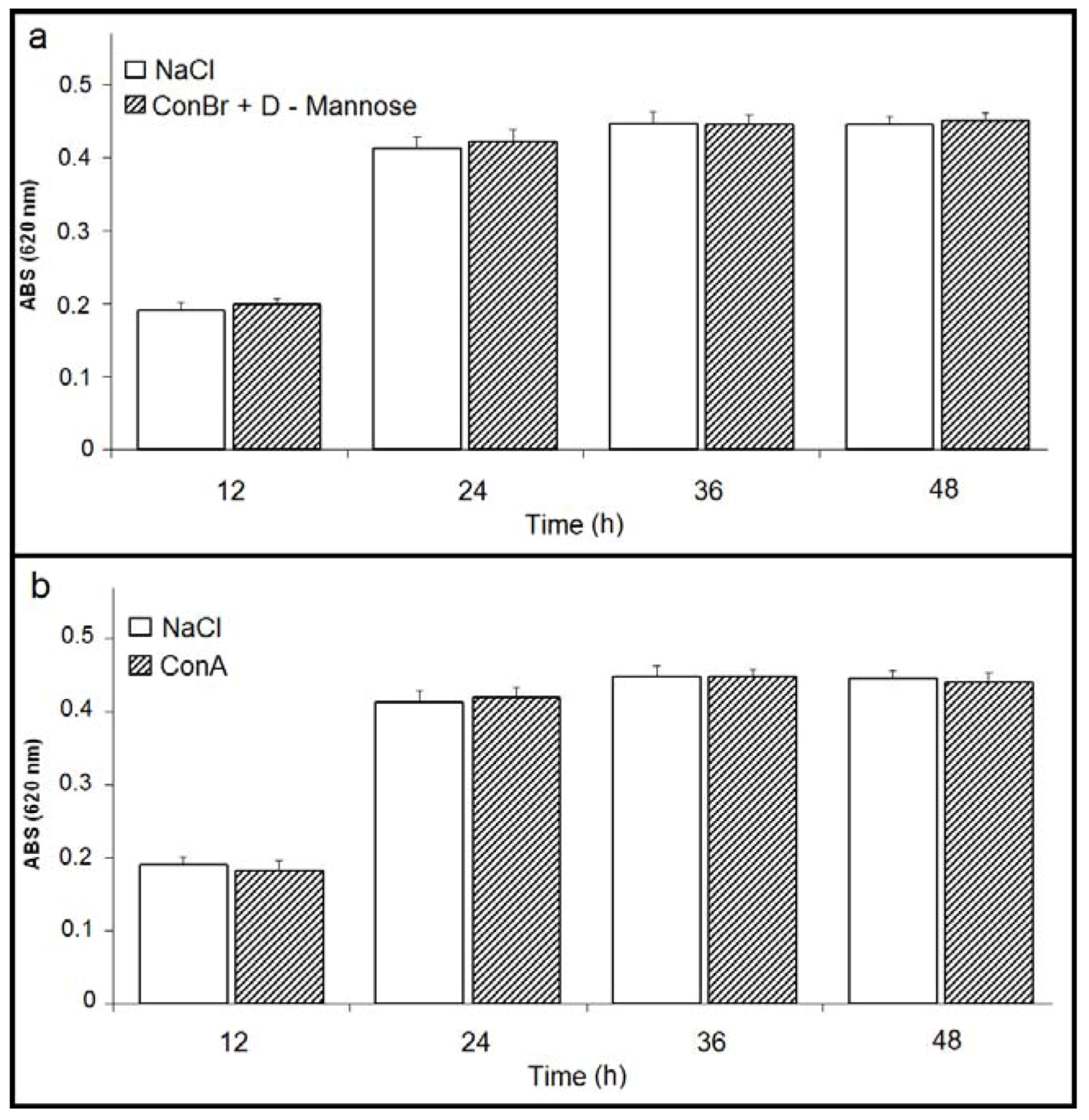
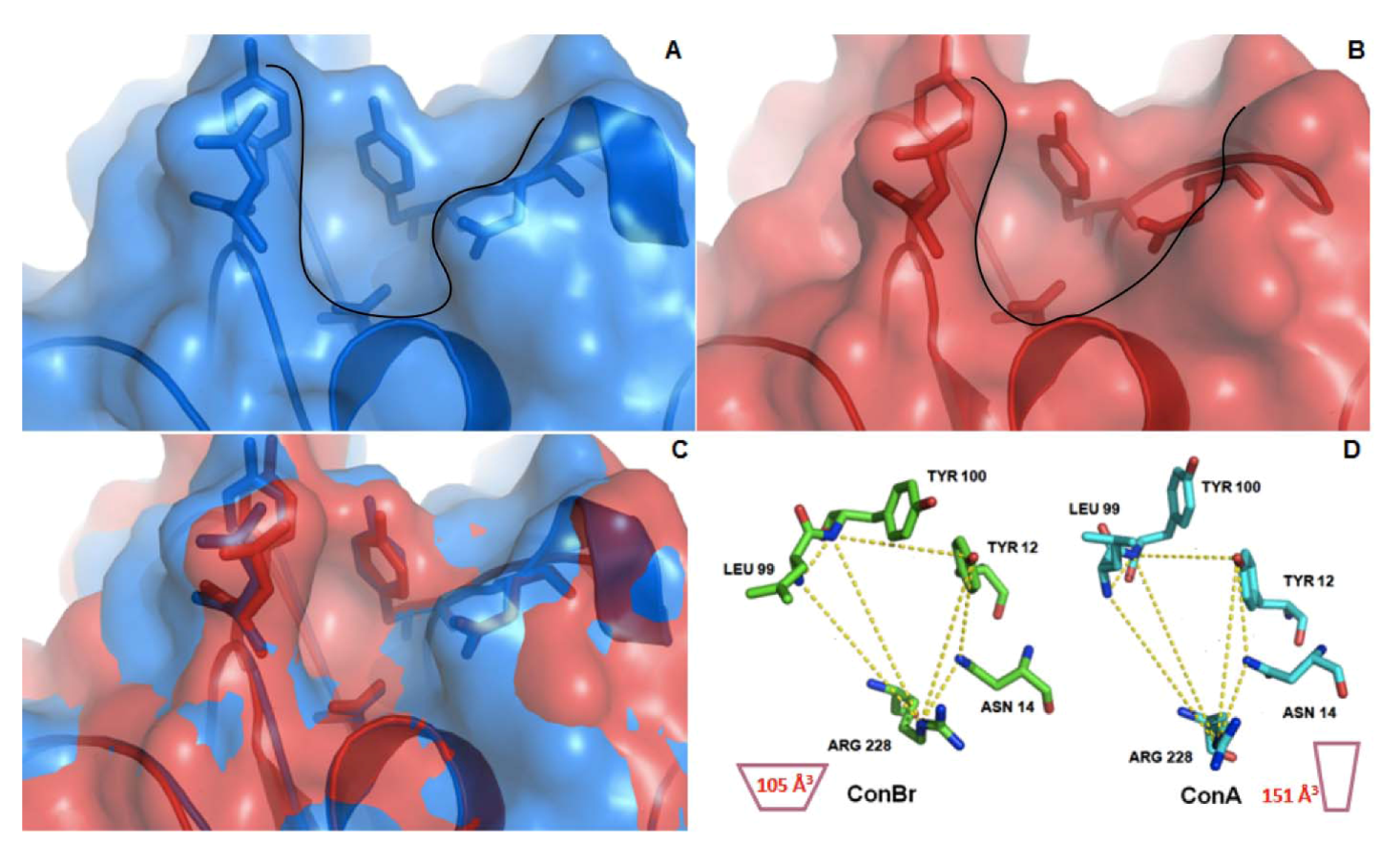
2. Results and Discussion
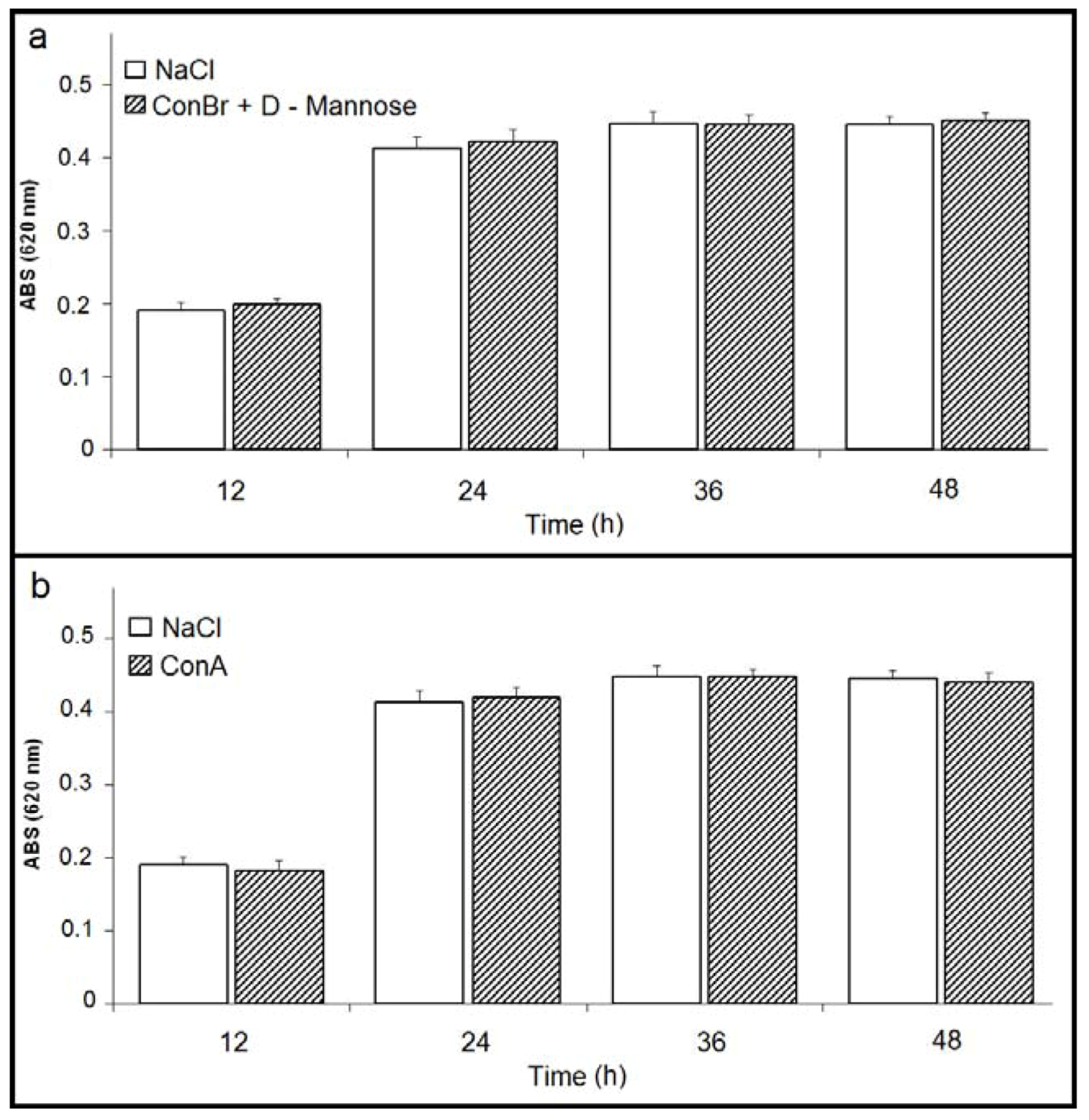
 ) p < 0.01 related to 0.15 M NaCl; (b) Comparison between the effects of ConBr and ConBr complexed with D-mannose on the growth of R. tropici. ( ) p < 0.01 related to ConBr plus D-mannose.
) p < 0.01 related to 0.15 M NaCl; (b) Comparison between the effects of ConBr and ConBr complexed with D-mannose on the growth of R. tropici. ( ) p < 0.01 related to ConBr plus D-mannose.
) p < 0.01 related to 0.15 M NaCl; (b) Comparison between the effects of ConBr and ConBr complexed with D-mannose on the growth of R. tropici. ( ) p < 0.01 related to ConBr plus D-mannose.
) p < 0.01 related to 0.15 M NaCl; (b) Comparison between the effects of ConBr and ConBr complexed with D-mannose on the growth of R. tropici. ( ) p < 0.01 related to ConBr plus D-mannose.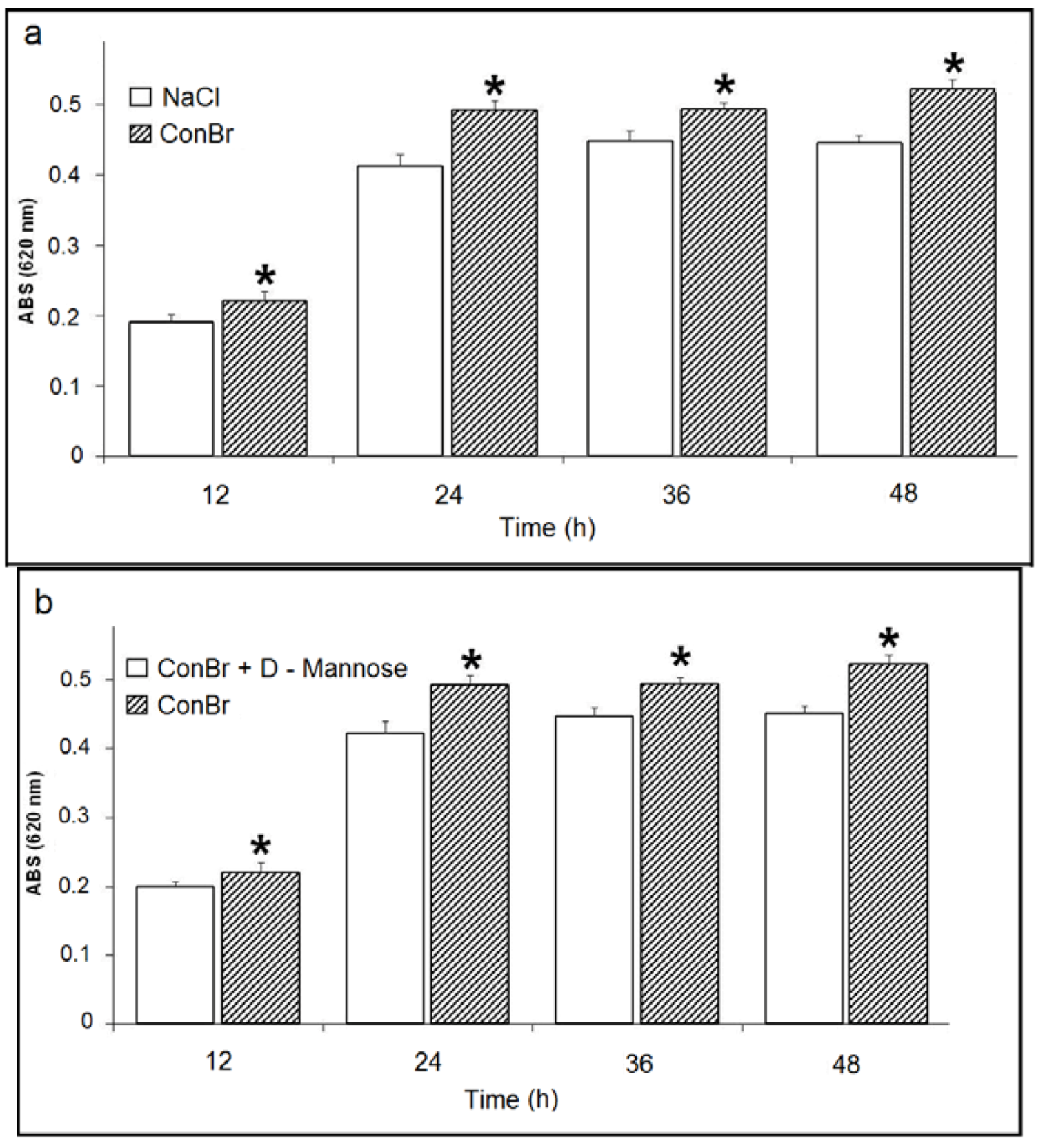
3. Experimental
3.1. Microorganism
3.2. Lectin Purification
3.3. Lectin Complexation with Mannose
3.4. FITC-Labeled Proteins
3.5. Cultivation of Microorganisms
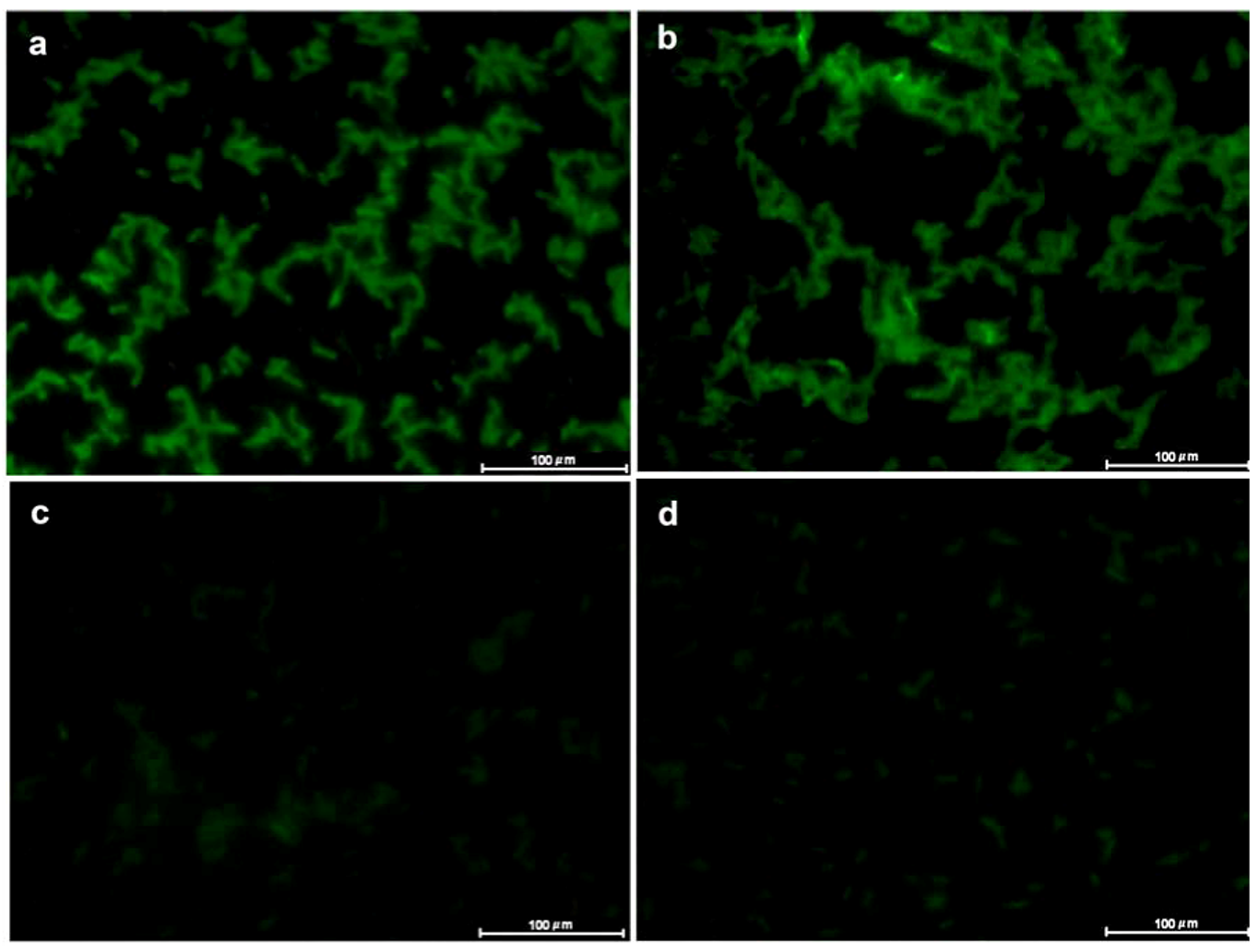
3.6. Lectin Fluorescence Microscopy
3.7. Bacterial Growth Assay
3.8. Statistical Analysis
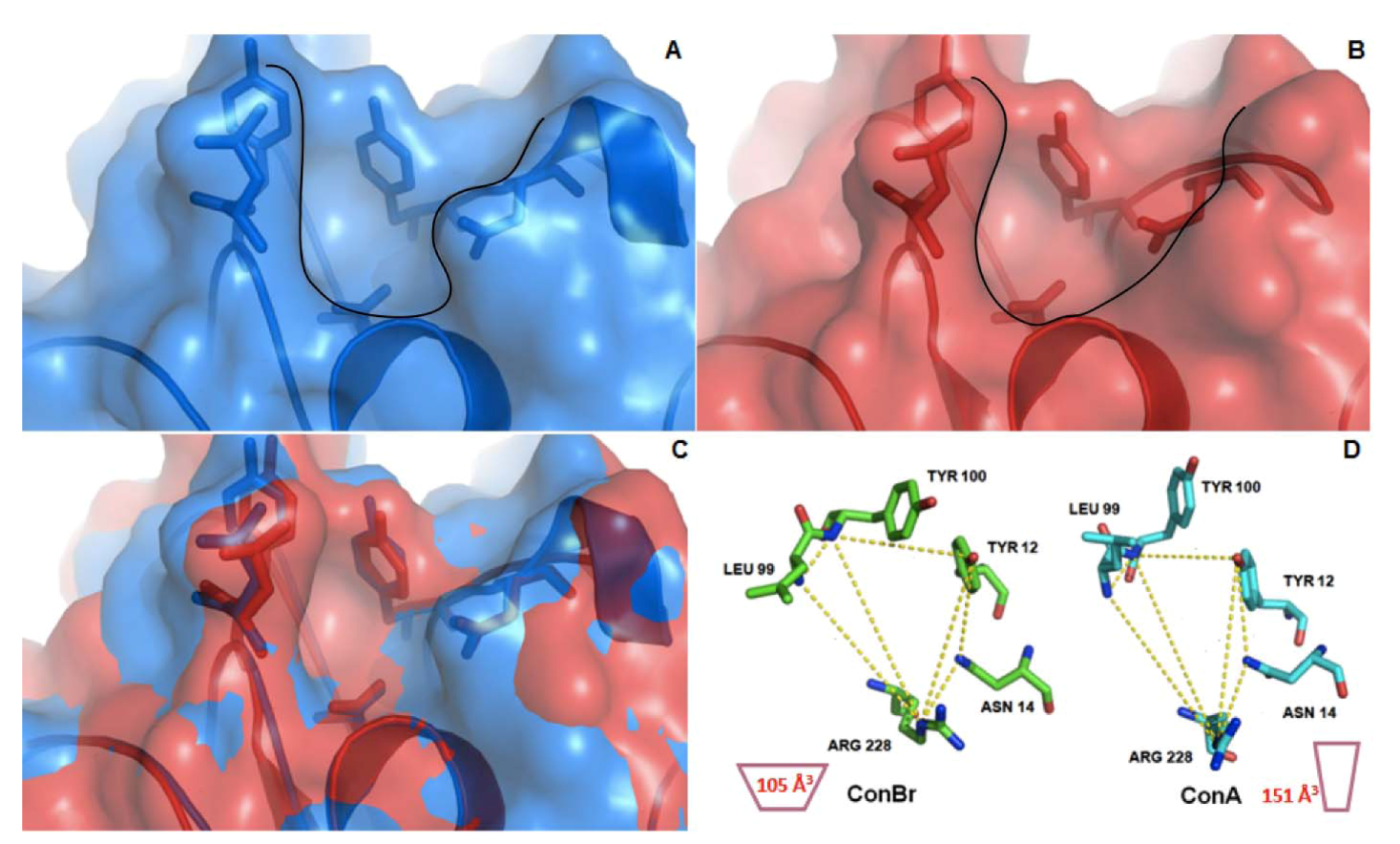
3.9. Structure/Function Analysis
4. Conclusions
Acknowledgments
- Sample Availability: Contact the authors.
References and Notes
- Ribeiro, R.A.; Barcellos, F.G.; Thompson, F.L.; Hungria, M. Multilocus sequence analysis of Brazilian Rhizobium microsymbionts of common bean (Phaseolus vulgaris L.) reveals unexpected taxonomic diversity. Res. Microbiol. 2009, 160, 297–306. [Google Scholar]
- Masson-Boivin, C.; Giraud, E.; Perret, X.; Batut, J. Establishing nitrogen-fixing symbiosis with legumes: How many rhizobium recipes? Trends Microbiol. 2009, 17, 458–466. [Google Scholar] [CrossRef]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef]
- Pinto, F.G.S.; Hungria, M.; Mercante, F.M. Polyphasic characterization of Brazilian Rhizobium tropici strains effective in fixing N2 with common bean (Phaseolus vulgaris L.). Soil Biol. Biochem. 2007, 39, 1851–1864. [Google Scholar]
- Hungria, M.; Campo, R.J.; Mendes, I.C. Benefits of inoculation of the common bean (Phaseolus vulgaris) crop with efficient and competitive Rhizobium tropici strains. Biol. Fert. Soils 2003, 39, 88–93. [Google Scholar] [CrossRef]
- Pinto, F.G.S.; Chueire, L.M.O.; Vasconcelos, A.T.R.; Nicolás, M.F.; Almeida, L.G.P.; Souza, R.C.; Menna, P.; Barcellos, F.G.; Megías, M.; Hungria, M. Novel genes related to nodulation, secretion systems, and surface structures revealed by a genome draft of Rhizobium tropici strain PRF 81. Funct. Integr. Genomics 2009, 9, 263–270. [Google Scholar] [CrossRef]
- Agrawal, P.; Kumar, S.; Jaiswal, Y.K.; Das, H.R.; Das, R.H. As Mesorhizobium lipopolysaccharide (LPS) specific lectin (CRL) from the roots of nodulating host plant, Cicer arietinum. Biochimie 2001, 3, 440–449. [Google Scholar]
- de Hoff, P.L.; Brill, L.M.; Hirsch, A.M. Plant lectins: The ties that bind in root symbiosis and plant defense. Mol. Genet. Genomics 2009, 282, 1–15. [Google Scholar] [CrossRef]
- Bajaj, M.; Soni, G.; Singh, C.K. Interaction of pea (Pisum sativum L.) lectins with rhizobial strains. Microbiol. Res. 2001, 156, 71–74. [Google Scholar]
- Martinez, C.R.; Albertini, A.V.P.; Figueiredo, M.V.B.; Silva, V.L.; Sampaio, A.H.; Cavada, B.S.; Lima-Filho, J.L. Respiratory stimulus in Rhizobium sp. by legume lectins. World J. Microbiol. Biotechnol. 2004, 20, 77–83. [Google Scholar] [CrossRef]
- Martínez, C.R.; Chanway, C.P.; Albertini, A.V.; Figueiredo, M.V.; Sampaio, A.H.; Castellon, R.R.; Cavada, B.S.; Lima-Filho, J.L. The interaction of Vatairea macrocarpa and Rhizobium tropici: Net H+ efflux stimulus and alteration of extracellular Na+ concentration. FEMS Microbiol. Lett. 2004, 1238, 17–22. [Google Scholar]
- Sytnikov, D.M.; Kots, S.Y.; Datsenko, V.K.A. Efficacy of biological preparations of soybean root nodule bacteria modified with a homologous lectin. Prikl. Biokhim. Mikrobiol. 2007, 43, 304–310. [Google Scholar]
- Cavada, B.S.; Barbosa, T.; Arruda, S.; Grangeiro, T.B.; Barral-Netto, M. Revisiting proteus: Do minor changes in lectin structure matter in biological activity? Lessons from and potential biotechnological uses of the Diocleinae subtribe lectins. Curr. Protein Pept. Sci. 2001, 2, 123–135. [Google Scholar] [CrossRef]
- Andrade, J.L.; Arruda, S.; Barbosa, T.; Paim, L.; Ramos, M.V.; Cavada, B.S. Lectin-induced nitric oxide production. Cell. Immunol. 2009, 194, 98–102. [Google Scholar]
- Barauna, A.C.; Kaster, M.P.; Heckert, B.T.; Nascimento, K.S.; Rossi, F.M.; Teixeira, E.H.; Cavada, B.S.; Rodrigues, A.L.S.; Leal, R.B. Antidepressant-like effect of lectin from Canavalia brasiliensis (ConBr) administered centrally in mice. Pharmacol. Biochem. Behav. 2006, 85, 160–169. [Google Scholar] [CrossRef]
- Barbosa, T.; Arruda, S.; Cavada, B.S.; Grangeiro, T.B.; Freitas, L.A.R.; Barral Netto, M. In vivo lymphocyte activation and apoptosis by lectins of the Diocleinae subtribe. Mem. Inst. Oswaldo Cruz. 2001, 96, 673–678. [Google Scholar] [CrossRef]
- Pinto, F.G.S.; Chueire, L.M.O.; Vasconcelos, A.T.R.; Nicolás, M.F.; Almeida, L.G.P.; Souza, R.C.; Menna, P.; Barcellos, F.G.; Megías, M.; Hungria, M. Novel genes related to nodulation, secretion systems, and surface structures revealed by a genome draft of Rhizobium tropici strain PRF 81. Funct. Integr. Genomics 2009, 9, 263–270. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. Lectins—Proteins with a sweet tooth: Functions in cell recognition. Essays Biochem. 1995, 30, 59–75. [Google Scholar]
- Varki, A. Biological roles of oligosaccharides: All of the theories are correct. Glycobiology 1993, 3, 97–130. [Google Scholar] [CrossRef]
- Strathmann, M.; Wingender, J.; Flemming, H.C. Application of fluorescently labelled lectins for the visualization and biochemical characterization of polysaccharides in biofilms of Pseudomonas aeruginosa. J. Microbiol. Methods 2002, 20, 237–248. [Google Scholar]
- Biswas, S.; Saroha, A.; Das, H.R. A lectin from Sesbania aculeata (Dhaincha) roots and its possible function. Biochemistry Mosc. 2009, 74, 329–335. [Google Scholar] [CrossRef]
- Antonyuk, L.P.; Fomina, O.R.; Ignatov, V.V. Effect of wheat lectin on the metabolism of Azospirillum brasiliense: Induction of protein synthesis. Microbiology (Engl. Transl. Mikrobiologiya) 2001, 66, 140–145. [Google Scholar]
- Araujo-Filho, J.H.; Vasconcelos, I.M.; Martins-Miranda, A.S.; Gondim, D.M.F.; Oliveira, J.T.A. A ConA-like lectin from Dioclea guianensis Benth. has antifungal activity against Colletotrichum gloeosporioides, unlike Its Homologues, ConM and ConA. J. Agric. Food Chem. 2010, 58, 4090–4096. [Google Scholar]
- Barral-Netto, M.; Santos, S.B.; Barral, A.; Moreira, L.I.; Santos, C.F.; Moreira, R.A.; Oliveira, J.T.; Cavada, B.S. Human lymphocyte stimulation by legume lectins from the Diocleae tribe. Immunol. Invest. 1992, 21, 297–303. [Google Scholar] [CrossRef]
- Gadelha, C.A.; Moreno, F.B.; Santi-Gadelha, T.; Cajazeiras, J.B.; Rocha, B.A.; Assreuy, A.M.; Lima Mota, M.R.; Pinto, N.V.; Meireles, A.V.P.; Borges, J.C.; et al. Native crystal structure of a nitric oxide-releasing lectin from the seeds of Canavalia maritima. J. Struct. Biol. 2005, 152, 185–194. [Google Scholar] [CrossRef]
- Brinda, K.V.; Mitra, N.; Surolia, A.; Vishveshwara, S. Determinants of quaternary association in legume lectins. Protein Sci. 2004, 13, 1735–1749. [Google Scholar] [CrossRef]
- Loris, R.; Hamelryck, T.; Bouckaert, J.; Wyns, L. Legume lectin structure. Biochim. Biophys. Acta 1998, 1383, 9–36. [Google Scholar] [CrossRef]
- Sanz-Aparício, J.; Hermoso, J.; Granjeiro, T.B.; Calvete, J.J.; Cavada, B.S. The crystal structure of Canavalia brasiliensis lectin suggests a correlation between its quaternary conformation and its distinct lectin biological properties from Concanavalin A. FEBS Lett. 1997, 405, 114–118. [Google Scholar] [CrossRef]
- Bezerra, E.H.; Rocha, B.A.; Bagano, C.S.; Moura, T.R.; Bezerra, G.A.; Bezerra, M.J.; Benevides, R.G.; Sampaio, A.H.; Assreuy, A.M.; Delatorre, P.; et al. Structural analysis of ConBr reveals molecular correlation between the carbohydrate recognition domain and endothelial NO synthase activation. Biochem. Biophys. Res. Commun. 2011, 8, 566–570. [Google Scholar]
- Ramos, M.V.; Cavada, B.S.; Mazard, A.M.; Roug, P. Interaction of Diocleinae lectins with glycoproteins based in surface plasmon resonance. Mem. Inst. Oswaldo Cruz 2002, 97, 275–279. [Google Scholar] [CrossRef]
- Bezerra, G.A.; Oliveira, T.M.; Moreno, F.B.; Souza, E.P.; Rocha, B.A.; Benevides, R.G.; Delatorre, P.; Azevedo, W.F., Jr.; Cavada, B.S. Structural analysis of Canavalia maritima and Canavalia gladiata lectins complexed with different dimannosides: New insights into the understanding of the structure-biological activity relationship in legume lectins. J. Struct. Biol. 2007, 160, 168–176. [Google Scholar] [CrossRef]
- Moreira, R.A.; Cavada, B.S. Lectin from Canavalia brasiliensis (Mart.). Isolation, characterisation and behaviour during germination. Biol. Plant 1984, 26, 113–120. [Google Scholar]
- Vincent, J.M. A Manual for the Practical Study of Root-Nodule Bacteria; Blackwell: Oxford, UK, 1970; p. 164. [Google Scholar]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Laurie, A.T.; Jackson, R.M. Q-SiteFinder: An energy-based method for the prediction of protein-ligand binding sites. Bioinformatics 2005, 21, 1908–1916. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Vasconcelos, M.A.; Cunha, C.O.; Arruda, F.V.S.; Carneiro, V.A.; Mercante, F.M.; Do Nascimento Neto, L.G.; De Sousa, G.S.; Rocha, B.A.M.; Teixeira, E.H.; Cavada, B.S.; et al. Lectin from Canavalia brasiliensis Seeds (ConBr) Is a Valuable Biotechnological Tool to Stimulate the Growth of Rhizobium tropici in Vitro. Molecules 2012, 17, 5244-5254. https://doi.org/10.3390/molecules17055244
De Vasconcelos MA, Cunha CO, Arruda FVS, Carneiro VA, Mercante FM, Do Nascimento Neto LG, De Sousa GS, Rocha BAM, Teixeira EH, Cavada BS, et al. Lectin from Canavalia brasiliensis Seeds (ConBr) Is a Valuable Biotechnological Tool to Stimulate the Growth of Rhizobium tropici in Vitro. Molecules. 2012; 17(5):5244-5254. https://doi.org/10.3390/molecules17055244
Chicago/Turabian StyleDe Vasconcelos, Mayron Alves, Claudio Oliveira Cunha, Francisco Vassiliepe Sousa Arruda, Victor Alves Carneiro, Fabio Martins Mercante, Luiz Gonzaga Do Nascimento Neto, Giselly Soares De Sousa, Bruno Anderson Matias Rocha, Edson Holanda Teixeira, Benildo Sousa Cavada, and et al. 2012. "Lectin from Canavalia brasiliensis Seeds (ConBr) Is a Valuable Biotechnological Tool to Stimulate the Growth of Rhizobium tropici in Vitro" Molecules 17, no. 5: 5244-5254. https://doi.org/10.3390/molecules17055244