Structural Diversity of Streptococcal Mutans Synthesized under Different Culture and Environmental Conditions and Its Effect on Mutanase Synthesis


Abstract
:1. Introduction
2. Results and Discussion
2.1. Structure and Some Properties of Mutans Synthesized by Cariogenic Streptococci Grown under Different Culture Conditions

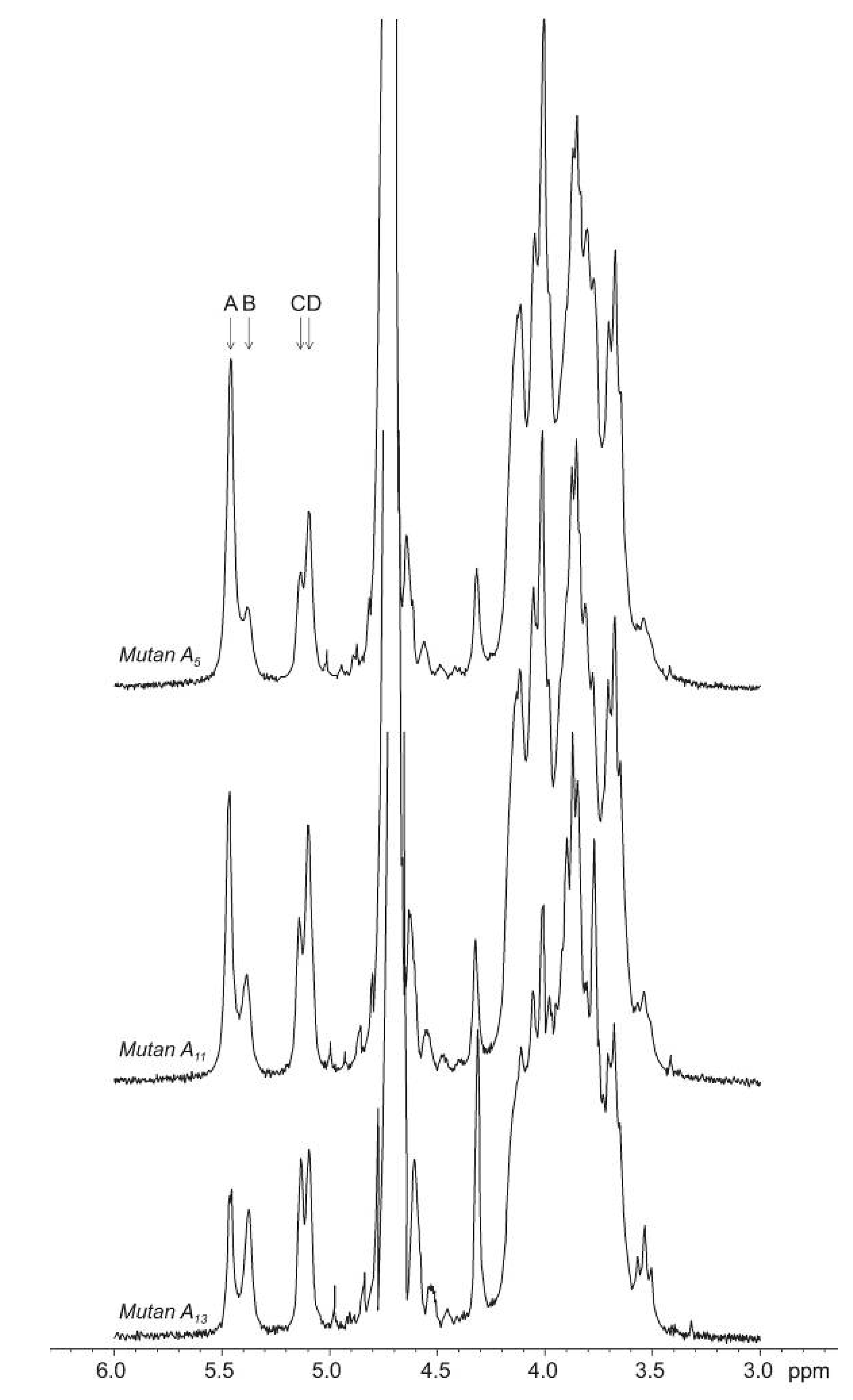{kind=link}
{kind=link}
| Strain | Medium | Mutan c | Content of glucosidic linkages (mol%) | Viscosity (mPa-s) | Optical rotation  (°) (°) | |||
|---|---|---|---|---|---|---|---|---|
| α-(1→3) chain | α-(1→6) chain | |||||||
| α-(1→3) | α-(1→3,6) d | α-(1→6) | α-(1→3,6) e | |||||
| S. sobrinus/downei 21020 | I | A1 | 38.7 | 19.7 | 22.1 | 19.5 | 8.3 | +218 |
| II | A2 | 44.0 | 11.9 | 26.4 | 17.7 | 15.8 | +208 | |
| BHI | A3 | 44.5 | 17.8 | 22.6 | 15.1 | 9.7 | +214 | |
| TTY | A4 | 48.7 | 14.0 | 24.9 | 12.4 | 17.8 | +216 | |
| THB | A5 | 49.1 | 14.0 | 24.5 | 12.4 | 6.6 | +208 | |
| TSB | A6 | 35.7 | 19.6 | 27.7 | 17.0 | 6.8 | +208 | |
| S. sobrinus 20381 | I | A7 | 33.8 | 23.0 | 24.0 | 19.2 | 8.3 | +224 |
| II | A8 | 34.0 | 17.2 | 34.0 | 14.8 | 5.3 | +206 | |
| BHI | A9 | 35.1 | 19.1 | 29.0 | 16.8 | 9.3 | +144 | |
| TTY | A10 | 36.2 | 23.7 | 20.8 | 19.3 | 10.2 | +210 | |
| THB | A11 | 37.2 | 16.9 | 30.5 | 15.4 | 7.6 | +216 | |
| TSB | A12 | 33.0 | 22.0 | 24.7 | 20.3 | 8.2 | +214 | |
| S. sobrinus 6070 | I | A13 | 23.7 | 25.5 | 27.8 | 23.0 | 4.8 | +198 |
| II | A14 | 28.7 | 25.7 | 22.7 | 22.9 | 25.4 | +224 | |
| BHI | A15 | 38.5 | 12.7 | 31.9 | 16.9 | 7.1 | +110 | |
| TTY | A16 | 32.4 | 14.5 | 38.7 | 14.4 | 19.9 | +112 | |
| THB | A17 | 26.7 | 18.4 | 33.9 | 21.0 | 6.3 | +208 | |
| TSB | - f | - | - | - | - | - | - | |
| S. mutans 6067 | I | A18 | 37.5 | 14.0 | 34.2 | 14.3 | 13.2 | +194 |
| II | - | - | - | - | - | - | - | |
| BHI | A19 | 36.9 | 13.7 | 32.7 | 16.7 | 9.6 | +196 | |
| TTY | - | - | - | - | - | - | - | |
| THB | A20 | 54.1 | 0.0 | 45.9 | 0.0 | 3.1 | +174 | |
| TSB | - | - | - | - | - | - | - | |
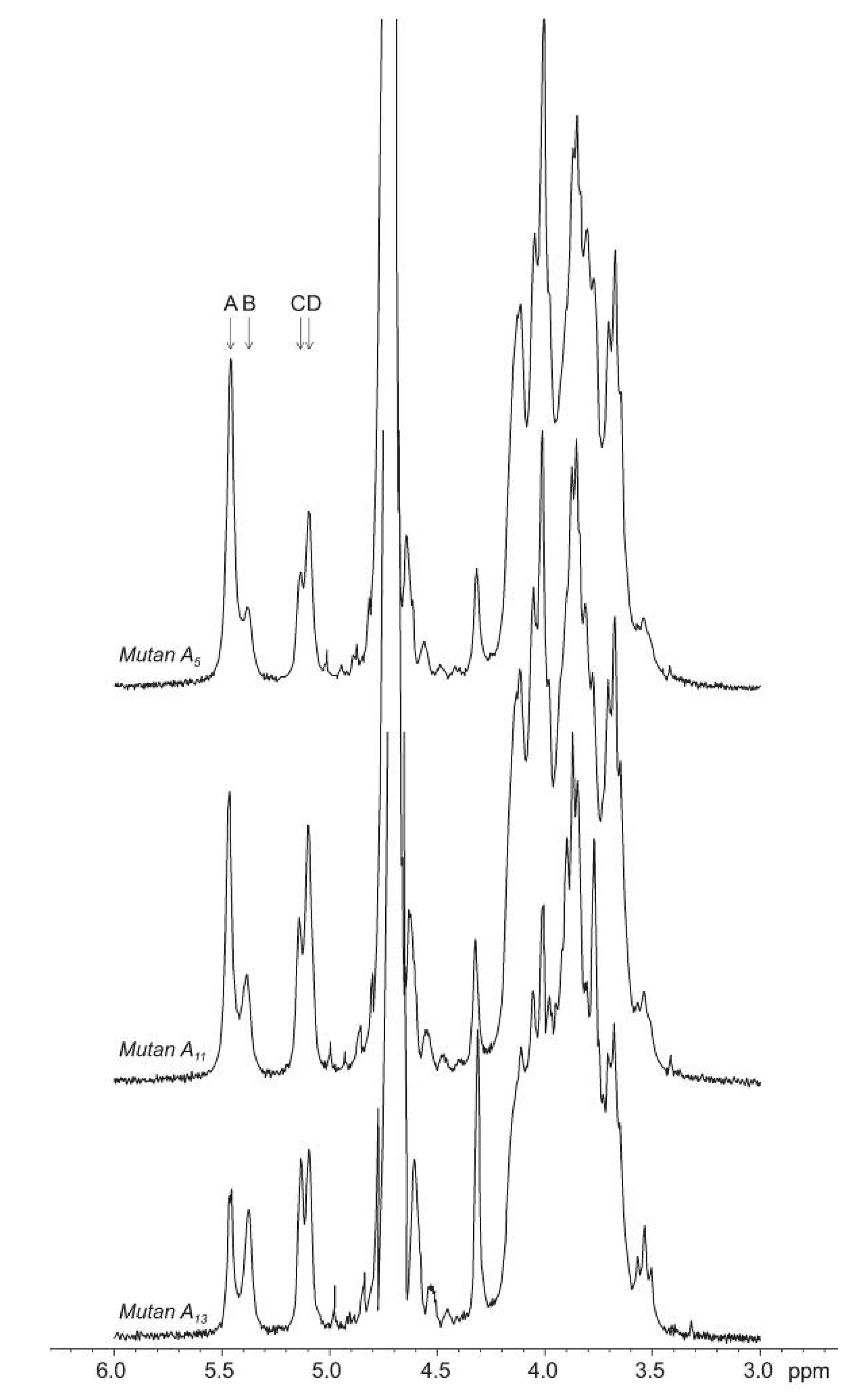 +110° and +224° (mutans A15 and A7).
+110° and +224° (mutans A15 and A7).| Factor varied a,b | Mutan c | Content of glucosidic linkages (mol%) | Viscosity (mPa-s) | Optical rotation (°) | |||
|---|---|---|---|---|---|---|---|
| α-(1→3) chain | α-(1→6) chain | ||||||
| α-(1→3) | α-(1→3,6) d | α-(1→6) | α-(1→3,6) e | ||||
| Initial pH of the medium | |||||||
| 6.0 | B1 | 56.6 | 11.7 | 20.0 | 11.7 | 11.7 | +184 |
| 6.5 | B2 | 43.6 | 15.8 | 26.0 | 14.6 | 18.5 | +242 |
| 7.0 | B3 | 43.8 | 16.1 | 26.0 | 14.1 | 16.1 | +220 |
| 7.5 | B4 | 38.5 | 17.8 | 26.8 | 16.9 | 15.5 | +222 |
| 8.0 | B5 | 41.4 | 17.9 | 24.0 | 16.7 | 13.5 | +220 |
| 8.5 | B6 | 57.9 | 9.5 | 23.2 | 9.4 | 21.0 | +240 |
| Culture temperature (°C) | |||||||
| 30 | C1 | 62.9 | 3.9 | 27.8 | 5.4 | 5.0 | +198 |
| 37 | C2 | 48.0 | 12.9 | 27.1 | 12.0 | 17.0 | +196 |
| 40 | C3 | 56.0 | 8.9 | 24.4 | 10.7 | 19.3 | +210 |
| Glucose concentration (%) | |||||||
| 0.00 | D1 | 43.0 | 15.8 | 25.6 | 15.6 | 5.4 | +148 |
| 0.05 | D2 | 49.6 | 17.9 | 17.1 | 15.4 | 8.1 | +210 |
| 0.10 | D3 | 42.7 | 17.3 | 23.5 | 16.5 | 10.5 | +148 |
| 0.25 | D4 | 48.9 | 14.8 | 24.7 | 11.6 | 3.8 | +152 |
| 0.50 | D5 | 57.5 | 14.9 | 15.6 | 12.0 | 5.9 | +146 |
| 1.00 | D6 | 59.8 | 13.6 | 14.4 | 12.4 | 5.7 | +218 |
| Cultivation time (h) | |||||||
| 6 | E1 | 57.6 | 7.2 | 27.3 | 7.9 | 22.9 | +260 |
| 12 | E2 | 41.3 | 13.2 | 33.4 | 12.1 | 11.2 | +230 |
| 18 | E3 | 33.7 | 22.2 | 23.3 | 20.8 | 6.9 | +218 |
| 24 | E4 | 34.1 | 24.4 | 18.2 | 23.3 | 3.9 | +206 |
| 36 | E5 | 43.2 | 21.1 | 16.8 | 18.9 | 2.4 | +208 |
| 48 | E6 | 42.4 | 21.5 | 16.0 | 20.1 | 2.3 | +202 |
| Kind of culture: | |||||||
| Anaerobic f | F1 | 43.5 | 16.1 | 25.7 | 14.6 | 7.7 | +222 |
| aerobic | F2 | 34.8 | 24.3 | 19.4 | 21.5 | 4.5 | +210 |
+148° to +260° (polymers D1 and E1), respectively.2.2. Structure and Some Properties of Mutans Synthesized in Post-Culture Supernates Incubated under Different Environmental Conditions
| Factor varied a | Mutan b | Content of glucosidic linkages (mol%) | Viscosity (mPa-s) | Optical rotation (°) | |||
|---|---|---|---|---|---|---|---|
| α-(1→3) chain | α-(1→6) chain | ||||||
| α-(1→3) | α-(1→3,6) c | α-(1→6) | α-(1→3,6) d | ||||
| pH of culture supernate | |||||||
| 5.0 | G1 | 26.4 | 24.2 | 26.4 | 23.0 | 4.9 | +208 |
| 5.5 | G2 | 28.2 | 22.4 | 26.5 | 22.9 | 5.3 | +210 |
| 6.0 | G3 | 31.1 | 23.9 | 22.1 | 22.9 | 5.6 | +210 |
| 6.5 | G4 | 33.6 | 23.5 | 19.2 | 23.7 | 6.0 | +216 |
| 7.0 | G5 | 39.2 | 21.1 | 18.6 | 21.1 | 23.9 | +226 |
| 7.5 | G6 | 45.0 | 19.5 | 15.3 | 20.2 | 19.6 | +220 |
| 8.0 | G7 | 60.3 | 15.0 | 11.8 | 12.9 | 30.2 | +232 |
| Reaction temperature (°C) | |||||||
| 20 | H1 | 17.3 | 25.7 | 32.0 | 25.0 | 7.1 | +184 |
| 30 | H2 | 27.7 | 23.9 | 26.2 | 22.2 | 6.6 | +206 |
| 37 | H3 | 32.5 | 22.7 | 23.8 | 21.0 | 5.7 | +218 |
| 40 | H4 | 35.5 | 21.9 | 22.4 | 20.2 | 6.0 | +214 |
| 45 | H5 | 52.9 | 14.8 | 18.7 | 13.6 | 15.2 | +234 |
| 50 | H6 | 51.0 | 12.9 | 23.9 | 12.2 | 15.5 | +240 |
| Sucrose concentration (%) e | |||||||
| 1 | I1 | 45.4 | 20.7 | 16.3 | 17.6 | 4.9 | +212 |
| 2 | I2 | 43.4 | 21.9 | 16.6 | 18.1 | 4.4 | +212 |
| 3 | I3 | 40.8 | 22.2 | 18.0 | 19.0 | 4.9 | +208 |
| 5 | I4 | 36.9 | 22.6 | 20.4 | 20.1 | 5.1 | +198 |
| 10 | I5 | 31.2 | 23.4 | 23.6 | 21.8 | 4.9 | +192 |
| 15 | I6 | 30.0 | 23.2 | 25.3 | 21.5 | 5.8 | +194 |
| 20 | I7 | 30.3 | 23.1 | 26.1 | 20.5 | 5.3 | +202 |
| Reaction time (h) | |||||||
| 6 | J1 | 31.0 | 21.6 | 26.9 | 20.5 | 11.8 | +216 |
| 12 | J2 | 33.2 | 20.7 | 26.6 | 19.5 | 11.1 | +220 |
| 18 | J3 | 34.2 | 20.8 | 25.6 | 19.4 | 9.4 | +218 |
| 24 | J4 | 33.8 | 20.5 | 26.1 | 19.6 | 9.1 | +218 |
| 36 | J5 | 37.2 | 19.7 | 24.8 | 18.3 | 7.5 | +218 |
| 48 | J6 | 38.3 | 19.4 | 24.4 | 17.9 | 6.8 | +214 |
| Sucrose source f | |||||||
| I | K1 | 30.1 | 22.9 | 21.5 | 25.5 | 5.7 | +210 |
| II | K2 | 30.0 | 22.9 | 24.9 | 22.2 | 5.1 | +200 |
| III | K3 | 30.1 | 22.7 | 25.0 | 22.2 | 5.3 | +212 |
| IV | K4 | 27.4 | 24.2 | 25.4 | 23.0 | 5.9 | +212 |
| V | K5 | 31.7 | 22.2 | 23.9 | 22.2 | 5.8 | +192 |
| VI | K6 | 32.6 | 21.7 | 22.2 | 23.5 | 5.9 | +212 |
| VII | K7 | 30.6 | 22.2 | 24.5 | 22.7 | 5.7 | +210 |
+184° to +240° (mutans H1 and H6), respectively. Data concerning the intrinsic viscosity and the optical rotation of mutans are rarely given in the literature. The values of these parameters provided by some authors [19,30,31] for water-insoluble biopolymers synthesized under optimal conditions by various strains of oral streptococci are relatively high and fluctuate within the limits of 0.46–9.6 [η]20 dL/g and from +197° to +228°, respectively. As showed by Tsumuraya and Misaki [31], the high positive values of optical rotation obtained for streptococcal mutans are indicative of α-D-glucosidic linkages. Also, Rees and Scott [32] have provided evidence that α-(1→3) linkages in the main chain of glucan confer rigidity and hence high viscosity.2.3. Effect of Structurally Diverse Mutans on Mutanase Synthesis Produced by T. harzianum
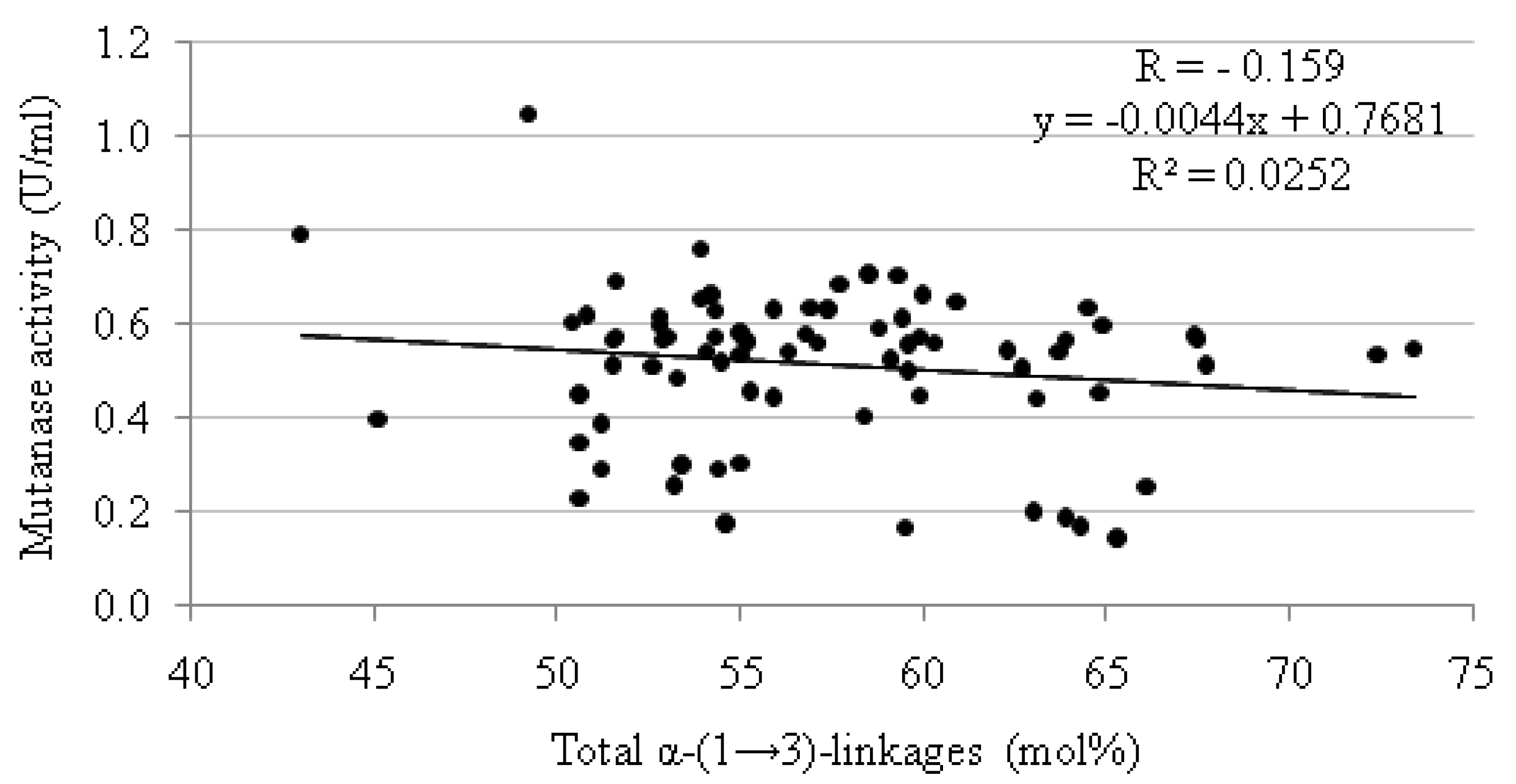
| Mutan | Mutanase activity(U/mL) | Mutan | Mutanase activity(U/mL) | Mutan | Mutanase activity(U/mL) | |||
|---|---|---|---|---|---|---|---|---|
| N° | Total content of α-(1→3)-linkages (mol%) | N° | Total content of α-(1→3)-linkages (mol%) | N° | Total content of α-(1→3)-linkages (mol%) | |||
| A1 | 58.4 | 0.406 | C1 | 66.8 | 0.665 | H3 | 55.2 | 0.564 |
| A2 | 55.9 | 0.445 | C2 | 60.9 | 0.648 | H4 | 57.4 | 0.632 |
| A3 | 62.3 | 0.545 | C3 | 64.9 | 0.600 | H5 | 67.7 | 0.514 |
| A4 | 62.7 | 0.507 | D1 | 58.8 | 0.591 | H6 | 63.9 | 0.566 |
| A5 | 63.1 | 0.441 | D2 | 67.5 | 0.570 | I1 | 66.1 | 0.253 |
| A6 | 55.3 | 0.458 | D3 | 60.0 | 0.665 | I2 | 65.3 | 0.144 |
| A7 | 56.8 | 0.579 | D4 | 63.7 | 0.542 | I3 | 63.0 | 0.201 |
| A8 | 51.2 | 0.293 | D5 | 72.4 | 0.536 | I4 | 59.5 | 0.168 |
| A9 | 54.2 | 0.665 | D6 | 73.4 | 0.547 | I5 | 54.6 | 0.175 |
| A10 | 59.9 | 0.446 | E1 | 64.8 | 0.454 | I6 | 53.2 | 0.256 |
| A11 | 54.1 | 0.542 | E2 | 54.5 | 0.519 | I7 | 53.4 | 0.300 |
| A12 | 55.0 | 0.535 | E3 | 55.9 | 0.633 | J1 | 52.6 | 0.509 |
| A13 | 49.2 | 1.051 | E4 | 58.5 | 0.707 | J2 | 53.9 | 0.656 |
| A14 | 54.4 | 0.291 | E5 | 64.3 | 0.170 | J3 | 55.0 | 0.582 |
| A15 | 51.2 | 0.389 | E6 | 63.9 | 0.188 | J4 | 54.3 | 0.572 |
| A16 | 46.9 | 0.402 | F1 | 59.6 | 0.558 | J5 | 56.9 | 0.635 |
| A17 | 45.1 | 0.396 | F2 | 59.1 | 0.527 | J6 | 57.7 | 0.686 |
| A18 | 51.5 | 0.514 | G1 | 50.6 | 0.347 | K1 | 53.0 | 0.573 |
| A19 | 50.6 | 0.452 | G2 | 50.6 | 0.230 | K2 | 52.9 | 0.566 |
| A20 | 54.1 | 0.542 | G3 | 55.0 | 0.302 | K3 | 52.8 | 0.600 |
| B1 | 68.3 | 0.585 | G4 | 57.1 | 0.560 | K4 | 51.6 | 0.691 |
| B2 | 59.4 | 0.614 | G5 | 60.3 | 0.562 | K5 | 53.9 | 0.761 |
| B3 | 59.9 | 0.572 | G6 | 64.5 | 0.634 | K6 | 54.3 | 0.631 |
| B4 | 56.3 | 0.543 | G7 | 75.3 | 0.667 | K7 | 52.8 | 0.618 |
| B5 | 59.3 | 0.703 | H1 | 43.0 | 0.792 | |||
| B6 | 67.4 | 0.576 | H2 | 51.6 | 0.573 | |||
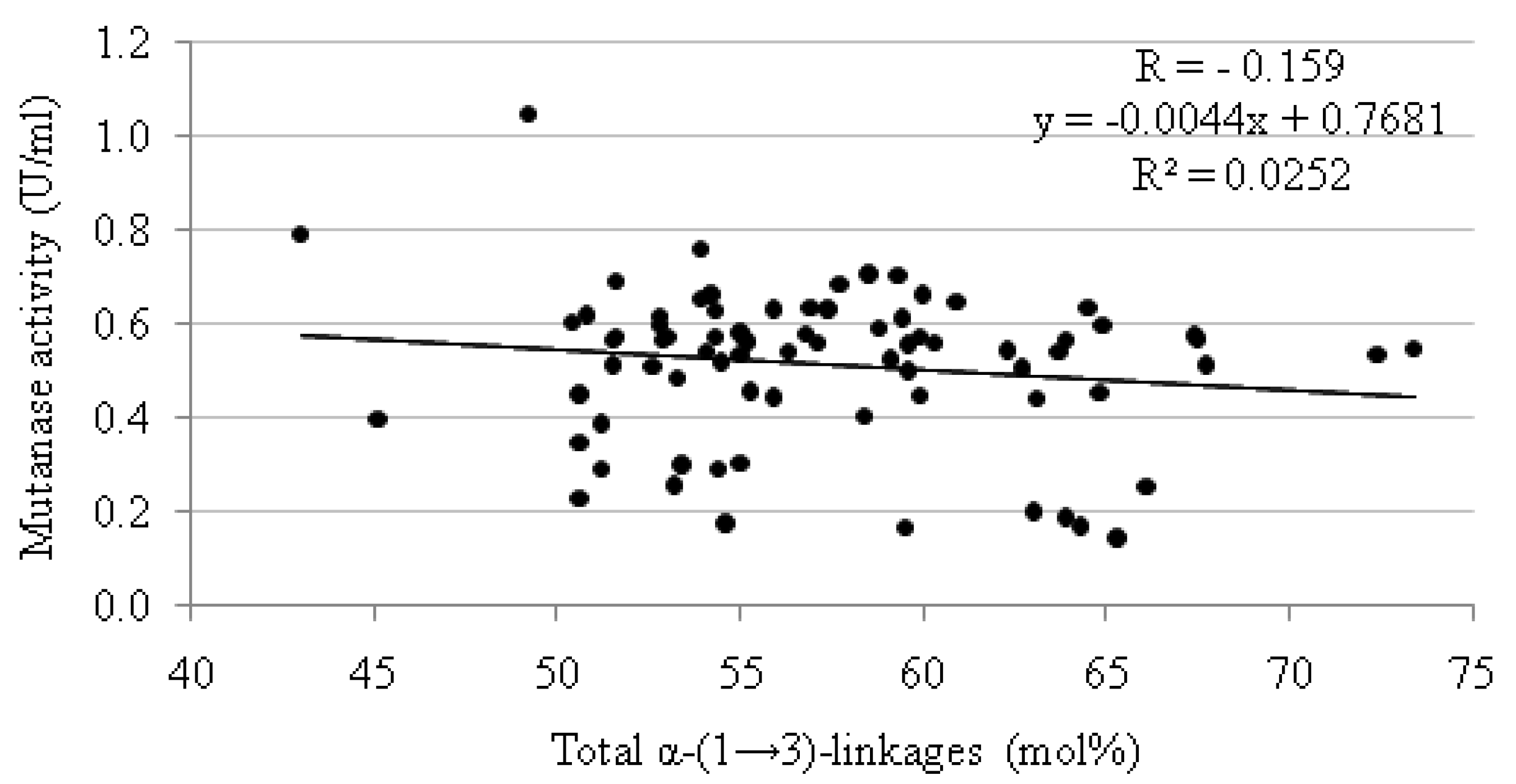
3. Experimental
3.1. Microorganisms
3.2. Bacterial Growth Conditions
3.3. Production of Mutan
3.4. Trichoderma harzianum Cultivation
3.5. Structural Studies
+ (c 1 M sodium hydroxide) was measured at 589 nm in a Perkin Elmer Automatic Polarimeter (Model 341 LC). The intrinsic viscosity of the polysaccharides (c 1 M sodium hydroxide) was measured with a Brookfield (Model DV 3) viscometer at 20 °C. 3.6. Mutanase Assay
3.7. Preparation of Dextranase-Treated Mutan (DTM) for Mutanase Activity Determination
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
- Sample Availability: Contact the authors.
References
- Russell, R.R.B. Bacterial Polysaccharides in Dental Plaque. In Bacterial Polysaccharides. Current Innovations and Future Trends; Ullrich, M., Ed.; Caister Academic Press: Norfolk, UK, 2009; pp. 143–156. [Google Scholar]
- Banas, J.A.; Vickerman, M.M. Glucan-binding proteins of the oral streptococci. Crit. Rev. Oral Biol. Med. 2003, 14, 89–99. [Google Scholar] [CrossRef]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilm. Carbohydr. Res. 2011, 45, 69–86. [Google Scholar]
- Colby, S.M.; Russell, R.R.B. Sugar metabolism by mutans streptococci. J. Appl. Microbiol. Symp. Suppl. 1997, 83, 80–88. [Google Scholar] [CrossRef]
- Pleszczyńska, M.; Marek-Kozaczuk, M.; Wiater, A.; Szczodrak, J. Paenibacillus strain MP-1: A new source of mutanase. Biotechnol. Lett. 2007, 29, 755–759. [Google Scholar] [CrossRef]
- Wiater, A.; Szczodrak, J.; Pleszczyńska, M.; Próchniak, K. Production and use of mutanase from Trichoderma harzianum for effective degradation of streptococcal mutans. Braz. J. Microbiol. 2005, 36, 137–146. [Google Scholar]
- Wiater, A.; Szczodrak, J.; Pleszczyńska, M. Optimization of conditions for the efficient production of mutan in streptococcal cultures and post-culture liquids. Acta Biol. Hung. 2005, 56, 137–150. [Google Scholar] [CrossRef]
- Kopec, L.K.; Vacca-Smith, A.M.; Bowen, W.H. Structural aspects of glucans formed in solution and on the surface of hydroxyapatite. Glycobiology 1997, 7, 929–934. [Google Scholar] [CrossRef]
- Shimamura, A. Use of 13C-NMR spectroscopy for the quantitative estimation of 3-O- and 3,6-di-O-substituted D-glucopyranosyl residues in a-D-glucans formed by the D-glucosyltransferases of Streptococcus sobrinus. Carbohydr. Res. 1989, 185, 239–248. [Google Scholar] [CrossRef]
- Yui, T.; Goto, K.; Kawano, Y.; Ogawa, K. Molecular modeling study of highly branching (1→3)-α-D-glucan, a model polysaccharide for cariogenic glucan, using the N–H mapping method. Biosci. Biotechnol. Biochem. 2000, 64, 52–60. [Google Scholar] [CrossRef]
- Wiater, A.; Szczodrak, J.; Rogalski, J. Hydrolysis of mutan and prevention of its formation in streptococcal films by fungal α-D-glucanases. Process Biochem. 2004, 39, 1481–1489. [Google Scholar] [CrossRef]
- Aires, C.P.; Koo, H.; Sassaki, G.L.; Iacomini, M.; Cury, J.A. A procedure for characterizing glucans synthesized by purified enzymes of cariogenic Streptococcus mutans. Int. J. Biol. Macromol. 2010, 46, 551–554. [Google Scholar] [CrossRef]
- Fukui, K.; Moriyama, T.; Miyake, Y.; Mizutani, K.; Tanaka, O. Purification and properties of glucosyltransferase responsible for water-insoluble glucan synthesis from Streptococcus mutans. Infect. Immun. 1982, 37, 1–9. [Google Scholar]
- Furuta, T.; Koga, T.; Nisizawa, T.; Okahashi, N.; Hamada, S. Purification and characterization of glucosyltransferases from Streptococcus mutans 6715. J. Gen. Microbiol. 1985, 131, 285–293. [Google Scholar]
- Guggenheim, B. Enzymatic hydrolysis and structure of water-insoluble glucan produced by glucosyltransferases from a strain of Streptococcus mutans. Helv. Odontol. Acta 1970, 14, 89–108. [Google Scholar]
- Wiater, A.; Choma, A.; Szczodrak, J. Insoluble glucans synthesized by cariogenic streptococci: A structural study. J. Basic Microbiol. 1999, 39, 265–273. [Google Scholar] [CrossRef]
- Inoue, M.; Koga, T.; Sato, S.; Hamada, S. Synthesis of adherent insoluble glucan by the concerted action of the two glucosyltransferase components of Streptococcus mutans. FEBS Lett. 1982, 143, 101–104. [Google Scholar] [CrossRef]
- Koga, T.; Sato, S.; Yakushiji, T.; Inoue, M. Separation of insoluble and soluble glucan-synthesizing glucosyltransferases of Streptococcus mutans OMZ 176 (serotype d. FEMS Microbiol. Lett. 1983, 16, 127–130. [Google Scholar] [CrossRef]
- Ebisu, S.; Misaki, A. The structure of water-insoluble glucans of cariogenic Streptococcus mutans, formed in the absence and presence of dextranase. Carbohydr. Res. 1974, 38, 374–381. [Google Scholar] [CrossRef]
- Aires, C.P.; Tenuta, L.M.; Carbonero, E.R.; Sassaki, G.L.; Iacomini, M.; Cury, J.A. Structural characterization of exopolysaccharides from biofilm of a cariogenic streptococci. Carbohydr. Polym. 2011, 84, 1215–1220. [Google Scholar] [CrossRef]
- Côté, G.L.; Leathers, T.D. Insoluble glucans from planktonic and biofilm cultures of mutans of Leuconostoc mesenteroides NRRL B135. Appl. Microbiol. Biotechnol. 2009, 82, 149–154. [Google Scholar] [CrossRef]
- Kopec, L.K.; Vacca-Smith, A.M.; Wunder, D.; Ng-Evans, L.; Bowen, W.H. Properties of Streptococcus sanguinis glucans formed under various conditions. Carbohydr. Res. 2001, 35, 67–74. [Google Scholar]
- Nisizawa, T.; Imai, S.; Akada, H.; Hinoide, M.; Araya, S. Extracellular glucans produced by oral streptococci. Arch. Oral Biol. 1976, 21, 207–213. [Google Scholar] [CrossRef]
- Eifuku, H.; Yoshimitsu-Narita, A.; Sato, S.; Yakushiji, T.; Inoue, M. Production and partial characterization of the extracellular polysaccharides from oral Streptococcus salivarius. Carbohydr. Res. 1989, 194, 247–260. [Google Scholar] [CrossRef]
- Quivey, R.G.; Kriger, P.S. Raffinose-induced mutanase production from Trichoderma harzianum. FEMS Microbiol. Lett. 1993, 112, 307–312. [Google Scholar] [CrossRef]
- Fuglsang, C.C.; Berka, R.M.; Wahleithner, J.A.; Kauppinen, S.; Shuster, J.R.; Rasmussen, G.; Halkier, T.; Dalbøge, H.; Henrissat, B. Biochemical analysis of recombinant fungal mutanases. J. Biol. Chem. 2000, 275, 2009–2018. [Google Scholar]
- Hamada, S.; Torii, M. Effect of sucrose in culture media on the location of glucosyltransferase of Streptococcus mutans and cell adherence to glass surfaces. Infect. Immun. 1978, 20, 592–599. [Google Scholar]
- Walker, G.J.; Brown, R.A.; Taylor, C. Activity of Streptococcus mutans α-D-glucosyltransferases released under various grown condtions. J. Dent. Res. 1984, 63, 397–400. [Google Scholar] [CrossRef]
- Walker, G.J.; Morrey-Jones, J.G.; Svensson, S.; Taylor, C. Effect of Variation in Growth Conditins on the Activity of D-Glucosyltransferases and the Synthesis of α-D-Glucans by Streptococcus mutans OMZ 176. In Glucosyltransferases, Glucans, Sucrose, and Dental Caries; Doyle, R.J., Ciardi, J.E., Eds.; DC-IRL Press: Washington, DC, USA, 1983; pp. 179–187. [Google Scholar]
- Ceska, M.; Granath, K.; Norrman, B.; Guggenheim, B. Structural and enzymatic studies on glucans synthesized with glucosyltransferases of some strains of oral streptococci. Acta Chem. Scand. 1972, 26, 2223–2230. [Google Scholar] [CrossRef]
- Tsumuraya, Y.; Misaki, A. Structure of the water-insoluble α-D-glucan of Streptococcus salivarius HHT. Carbohydr. Res. 1979, 74, 217–225. [Google Scholar] [CrossRef]
- Rees, D.A.; Scott, W.E. Polysaccharide conformation. Part VI. Computer model-building for linear and branched pyranoglycans. Correlations with biological function. Preliminary assessment of inter-residue forces in aqueous solution. Further interpretation of optical rotation in terms of chain conformation. J. Chem. Soc. B 1971, 469–479. [Google Scholar]
- Mandels, M.; Parrish, F.W.; Reese, E.T. Sophorose as an inducer of cellulase in Trichoderma viride. J. Bacteriol. 1962, 83, 400–408. [Google Scholar]
- Claridge, T.D.W. High-Resolution NMR Techniques in Organic Chemistry; Elsevier Science: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar]
- Somogyi, M. A new reagent for the determination of sugars. J. Biol. Chem. 1945, 160, 61–68. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wiater, A.; Pleszczyńska, M.; Próchniak, K.; Szczodrak, J. Structural Diversity of Streptococcal Mutans Synthesized under Different Culture and Environmental Conditions and Its Effect on Mutanase Synthesis. Molecules 2012, 17, 11800-11815. https://doi.org/10.3390/molecules171011800
Wiater A, Pleszczyńska M, Próchniak K, Szczodrak J. Structural Diversity of Streptococcal Mutans Synthesized under Different Culture and Environmental Conditions and Its Effect on Mutanase Synthesis. Molecules. 2012; 17(10):11800-11815. https://doi.org/10.3390/molecules171011800
Chicago/Turabian StyleWiater, Adrian, Małgorzata Pleszczyńska, Katarzyna Próchniak, and Janusz Szczodrak. 2012. "Structural Diversity of Streptococcal Mutans Synthesized under Different Culture and Environmental Conditions and Its Effect on Mutanase Synthesis" Molecules 17, no. 10: 11800-11815. https://doi.org/10.3390/molecules171011800