Diterpenoid Tanshinones and Phenolic Acids from Cultured Hairy Roots of Salvia miltiorrhiza Bunge and Their Antimicrobial Activities


Abstract
:1. Introduction

2. Results and Discussion
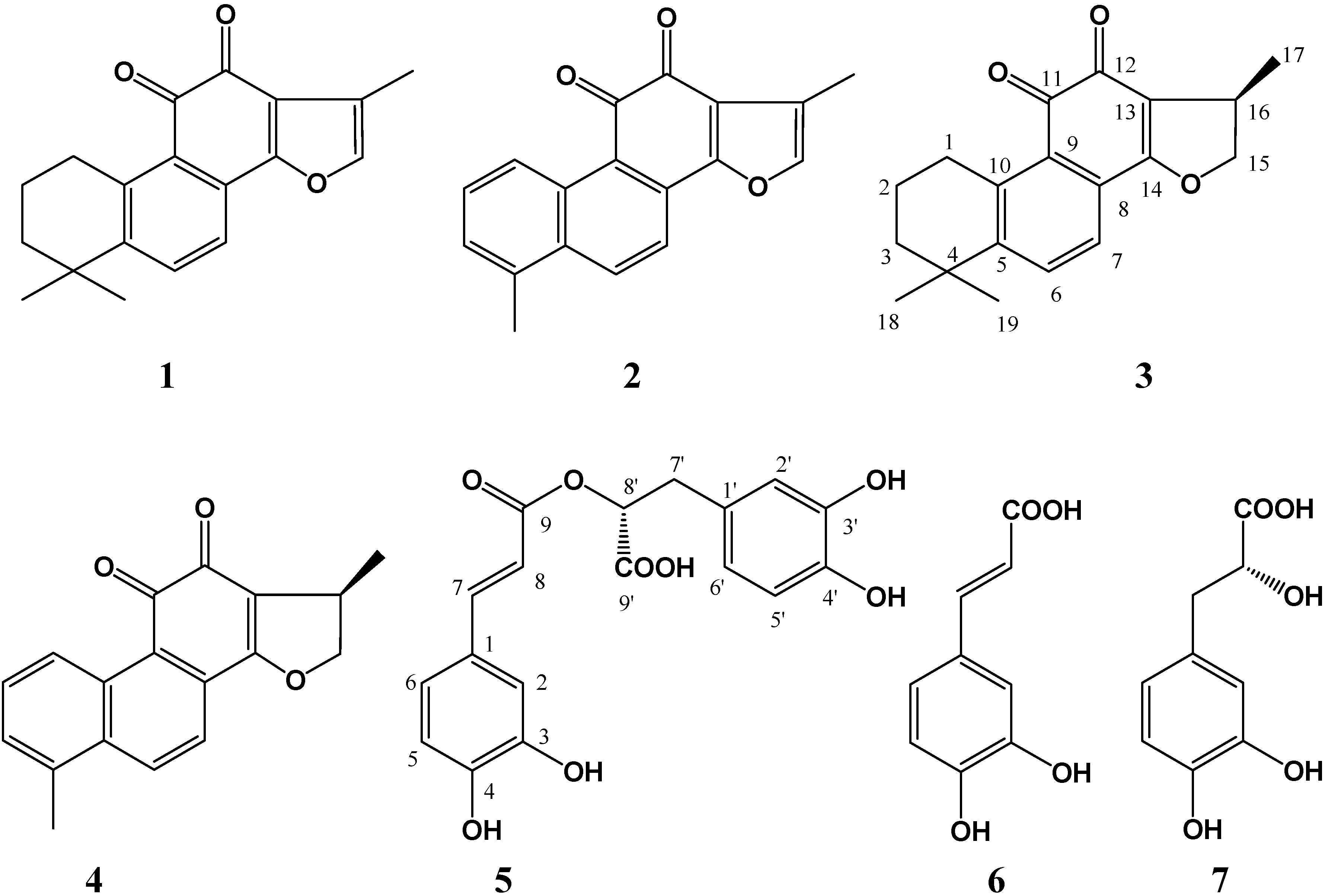
2.1. Isolation and identification
2.2. Antimicrobial activity

{kind=link}
| Micro-organism | MIC (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | CK+ | |
| A. t | 50 | 25 | 12.5 | 6.25 | 100 | 100 | 150 | 6.25 |
| E. c | 50 | 50 | 12.5 | 25 | 100 | 100 | 150 | 25 |
| P. l | 25 | 25 | 12.5 | 12.5 | 50 | 50 | 100 | 12.5 |
| R. s | 50 | 50 | 12.5 | 12.5 | 50 | 100 | 100 | 12.5 |
| X. v | 25 | 25 | 6.25 | 12.5 | 50 | 100 | 100 | 12.5 |
| B. s | 50 | 50 | 25 | 25 | 100 | 150 | 150 | 50 |
| S. a | 200 | 150 | 100 | 100 | 200 | 200 | >200 | 100 |
| S. h | 100 | 100 | 50 | 50 | 150 | 150 | 200 | 50 |
| M. o | 50 | 12.5 | 6.25 | 3.13 | 150 | 200 | >200 | 6.25 |
| Micro-organism | IC50 (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | CK+ | |
| A. t | 31.25 ± 0.56d | 13.56 ± 0.26e | 7.28 ± 0.35f | 4.03 ± 0.09g | 62.78 ± 1.21c | 65.32 ± 0.76b | 87.36 ± 1.02a | 3.83 ± 0.05g |
| E. c | 27.54 ± 0.38d | 20.29 ± 0.43e | 7.83 ± 0.51g | 11.17 ± 0.13f | 52.15 ± 0.82c | 57.28 ± 0.63b | 96.75 ± 1.36a | 7.55 ± 0.18g |
| P. l | 16.32 ± 0.25d | 11.18 ± 0.37e | 7.32 ± 0.62g | 9.10 ± 0.23f | 27.03 ± 0.51c | 32.65 ± 0.35b | 64.02 ± 0.73a | 6.82 ± 0.10g |
| R.s | 35.36 ± 0.52c | 31.58 ± 0.85e | 7.12 ± 0.23f | 7.78 ± 0.26f | 33.15 ± 0.63d | 63.27 ± 0.79b | 68.25 ± 0.64a | 6.75 ± 0.06f |
| X. v | 14.43 ± 0.31d | 11.09 ± 0.29e | 3.66 ± 0.16h | 7.57 ± 0.51f | 38.56 ± 0.49c | 57.23 ± 0.61b | 72.38 ± 0.89a | 5.72 ± 0.12g |
| B. s | 32.59 ± 0.67d | 27.54 ± 0.45e | 15.48 ± 0.34g | 17.61 ± 0.38f | 73.86 ± 0.92c | 91.85 ± 1.04b | 94.32 ± 1.16a | 27.35 ± 1.06e |
| S. a | 108.87 ± 1.83c | 83.06 ± 1.32d | 50.25 ± 1.42g | 57.38 ± 0.92f | 131.82 ± 1.58b | 142.36 ± 2.13a | nd | 65.98 ± 1.32e |
| S. h | 73.34 ± 1.21d | 61.81 ± 1.05e | 28.87 ± 0.88h | 35.56 ± 1.23f | 83.61 ± 1.32c | 102.58 ± 1.83b | 164.26 ± 2.35a | 31.94 ± 1.18g |
| M. o | 33.02 ± 0.87c | 6.10 ± 0.64d | 3.40 ± 0.12e | 0.91 ± 0.07f | 92.53 ± 1.36b | 128.35 ± 1.25a | nd | 2.86 ± 0.16e |
3. Experimental
3.1. General
3.2. Plant material
3.3. Extraction, fractionation and identification
3.4. Antibacterial activity assay
3.5. Antifungal activity assay
4. Conclusions
Acknowledgements
References
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Zhou, L.; Wu, J. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 2006, 23, 789–810. [Google Scholar] [CrossRef]
- Zhong, J.-J. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. Adv. Biochem. Engin. Biotechnol. 2001, 72, 1–26. [Google Scholar]
- Zhou, L.; Yang, C.; Li, J.; Wang, S.; Wu, J. Heptasaccharide and octasaccharide isolated from Paris polyphylla var. yunnanensis and their plant growth-regulatory activity. Plant Sci. 2003, 165, 571–575. [Google Scholar] [CrossRef]
- Zhou, L.; Cao, X.; Zhang, R.; Peng, Y.; Zhao, S.; Wu, J. Stimulation of saponin production in Panax ginseng hairy roots by two oligosaccharides from Paris polyphylla var. yunnanensis. Biotechnol. Lett. 2007, 29, 631–634. [Google Scholar] [CrossRef]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Adv. Biochem. Eng. Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Zhou, L.; Zuo, Z.; Chow, M.S.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef]
- Jiang, R.W.; Lau, K.M.; Hon, P.M.; Mak, T.C.W.; Woo, K.S.; Fung, K.P. Chemistry and biological activities of caffeic acid derivatives from Salvia miltiorrhiza. Curr. Med. Chem. 2005, 12, 237–246. [Google Scholar] [CrossRef]
- Wang, X.B.; Morris-Natschke, S.L.; Lee, K.H. New developments in the chemistry and biology of the bioactive constituents of Tanshen. Med. Res. Rev. 2007, 27, 133–148. [Google Scholar] [CrossRef]
- Hu, Z.B.; Alfermann, A.W. Diterpenoid production in hairy root cultures of Salvia miltiorrhiza. Phytochemistry 1993, 32, 699–703. [Google Scholar] [CrossRef]
- Chen, H.; Chen, F.; Zhang, Y.L.; Song, J.Y. Production of lithospermic acid B and rosmarinic acid in hairy root cultures of Salvia miltiorrhiza. J. Ind. Microbiol. Biotechnol. 1999, 12, 133–138. [Google Scholar]
- Zhao, J.; Zhou, L.; Wu, J. Promotion of Salvia miltiorrhiza hairy root growth and tanshinone production by polysaccharide-protein fractions of plant growth-promoting rhizobacterium Bacillus cereus. Process Biochem. 2010, 45, 1517–1522. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Choi, D.-Y.; Woo, E.-R. Inhibition of osteoclast differentiation by tanshinones from the root of Salvia miltiorrhiza Bunge. Arch. Pharm. Res. 2005, 28, 909–913. [Google Scholar] [CrossRef]
- Huang, W.G.; Jiang, Y.Y.; Li, Q.; Li, J.; Li, J.; Lu, W.; Cai, J.C. Synthesis and biological evaluation of (±)-cryptotanshinone and its simplified analogues as potent CDC25 inhibitors. Tetrahedron 2005, 61, 1863–1870. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Rosmarinic acid derivatives from Salvia officinalis. Phytochemistry 1999, 51, 91–94. [Google Scholar]
- Zhou, L.; Li, D.; Wang, J.; Liu, Y.; Wu, J. Antibacterial phenolic acids from the spines of Gleditsia sinensis Lam. Nat. Prod. Res. 2007, 21, 283–291. [Google Scholar] [CrossRef]
- Kelley, C.J.; Harruff, R.C.; Carmack, M. Polyphenolic acids of Lithospermum ruderale. II. Carbon-13 nuclear magnetic resonance of lithospermic and rosmarinic acids. J. Org. Chem. 1976, 41, 449–455. [Google Scholar] [CrossRef]
- Wang, J.W.; Wu, J.Y. Tanshinone biosynthesis in Salvia miltiorrhiza and production in plant tissue cultures. Appl. Microbiol. Biotechnol. 2010, 88, 437–449. [Google Scholar] [CrossRef]
- Gao, W.; Hillwig, M.L.; Huang, L.Q.; Cui, G.H.; Wang, X.Y.; Kong, J.Q.; Yang, B.; Peters, R.J. Functional genomics approach to tanshinone biosynthesis provides stereochemical insights. Org. Lett. 2009, 11, 5170–5173. [Google Scholar] [CrossRef]
- Don, M.-J.; Shen, C.-C.; Syu, W.-J.; Ding, Y.-H.; Sun, C.-M. Cytotoxic and aromatic constituents from Salvia miltiorrhiza. Phytochemistry 2006, 67, 497–503. [Google Scholar]
- Gao, Y.; Song, Y.; Yang, Y.; Liu, W.; Tang, J. Pharmacology of tanshinone. Acta Pharm. Sin. 1979, 14, 75–81. [Google Scholar]
- Zhu, J.; Luo, H. Antibacterial study of tanshinone IIA. J. China Pharm. Univ. 2004, 35, 368–370. [Google Scholar]
- Mothana, R.A.A.; Jansen, R.; Gruenert, R.; Bednarski, P.J.; Lindequist, U. Antimicrobial and cytotoxic abietane diterpenoids from the roots of Meriandera benghalensis (Roxb.) Benth. Pharmazie 2009, 64, 613–615. [Google Scholar]
- Jiang, R.-W.; Lau, K.-M.; Hon, P.-M.; Mak, T.C.W.; Woo, K.-S.; Fung, K.-P. Chemistry and biological activities of caffeic acid derivatives from Salvia miltiorrhiza. Curr. Med. Chem. 2005, 12, 237–246. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F.A. A revised medium for rapid growth and biosynthesis with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wang, J.; Liu, H.; Zhao, J.; Gao, H.; Zhou, L.; Liu, Z.; Chen, Y.; Sui, P. Antimicrobial and antioxidant activities of the root bark essential oil of Periploca sepium and its main component 2-hydroxy-4-methoxybenzaldehyde. Molecules 2010, 15, 5807–5817. [Google Scholar] [CrossRef]
- Abe, K.; Matsuki, N. Measurement of cellular 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction activity and lactate dehydrogenase release using MTT. Neurosci. Res. 2000, 38, 325–329. [Google Scholar] [CrossRef]
- Sakuma, M. Probit analysis of preference data. Appl. Entomol. Zool. 1998, 33, 339–347. [Google Scholar]
- Liu, H.; Wang, J.; Zhao, J.; Lu, S.; Wang, J.; Jiang, W.; Ma, Z.; Zhou, L. Isoquinoline alkaloids from Macleaya cordata active against plant microbial pathogens. Nat. Prod. Commun. 2009, 4, 1557–1560. [Google Scholar]
- Wang, J.; Zhao, J.; Liu, H.; Zhou, L.; Liu, Z.; Wang, J.; Han, J.; Yu, Z.; Yang, F. Chemical analysis and biological activity of the essential oils of two valerianaceous species from China: Nardostachys chinensis and Valeriana officinalis. Molecules 2010, 15, 6411–6422. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, J.; Lou, J.; Mou, Y.; Li, P.; Wu, J.; Zhou, L. Diterpenoid Tanshinones and Phenolic Acids from Cultured Hairy Roots of Salvia miltiorrhiza Bunge and Their Antimicrobial Activities. Molecules 2011, 16, 2259-2267. https://doi.org/10.3390/molecules16032259
Zhao J, Lou J, Mou Y, Li P, Wu J, Zhou L. Diterpenoid Tanshinones and Phenolic Acids from Cultured Hairy Roots of Salvia miltiorrhiza Bunge and Their Antimicrobial Activities. Molecules. 2011; 16(3):2259-2267. https://doi.org/10.3390/molecules16032259
Chicago/Turabian StyleZhao, Jianglin, Jingfeng Lou, Yan Mou, Peiqin Li, Jianyong Wu, and Ligang Zhou. 2011. "Diterpenoid Tanshinones and Phenolic Acids from Cultured Hairy Roots of Salvia miltiorrhiza Bunge and Their Antimicrobial Activities" Molecules 16, no. 3: 2259-2267. https://doi.org/10.3390/molecules16032259