WRINKLED1, a “Master Regulator” in Transcriptional Control of Plant Oil Biosynthesis


1
School of Biological Sciences, Nanyang Technological University, Singapore 637551, Singapore
2
Department of Plant and Soil Sciences, Kentucky Tobacco Research and Development Center, University of Kentucky, Lexington, KY 40546, USA
*
Author to whom correspondence should be addressed.
Plants 2019, 8(7), 238; https://doi.org/10.3390/plants8070238
Submission received: 20 June 2019
/
Revised: 18 July 2019
/
Accepted: 18 July 2019
/
Published: 22 July 2019
(This article belongs to the Special Issue Lipid Metabolism in Plants)
{kind=link}
Abstract
:A majority of plant species generate and accumulate triacylglycerol (TAG) in their seeds, which is the main resource of carbon and energy supporting the process of seedling development. Plant seed oils have broad ranges of uses, being not only important for human diets but also renewable feedstock of industrial applications. The WRINKLED1 (WRI1) transcription factor is vital for the transcriptional control of plant oil biosynthetic pathways. Since the identification of the Arabidopsis WRI1 gene (AtWRI1) fifteen years ago, tremendous progress has been made in understanding the functions of WRI1 at multiple levels, ranging from the identification of AtWRI1 target genes to location of the AtWRI1 binding motif, and from discovery of intrinsic structural disorder in WRI1 to fine-tuning of WRI1 modulation by post-translational modifications and protein-protein interactions. The expanding knowledge on the functional understanding of the WRI1 regulatory mechanism not only provides a clearer picture of transcriptional regulation of plant oil biosynthetic pathway, but also helps generate new strategies to better utilize WRI1 for developing novel oil crops.
1. Introduction
Plants mainly biosynthesize and accumulate plant oils (mostly as triacylglycerol, or TAG) in their seeds which serve as the carbon and energy resource for supporting seedling development. TAG, which is derived from seed oil and known to most people as vegetable oils, represents a highly energy-rich resource in nature (e.g., TAG has higher energy density compared to carbohydrates and proteins). In addition to the importance for the human diet, plant seed oils are also important as a renewable feedstock of industrial uses. Plant oils have broad applications for non-food industrial uses such as the manufacturing of soaps, detergents, and lubricants. Plant oils are also converted to biodiesel by chemical reactions. The global demand for vegetable oils is rapidly increasing and estimated to double by 2030 [1], escalating the necessity of increasing plant oil production to meet growing demand.
TAG biosynthesis involves cooperation between plastids and the endoplasmic reticulum (ER) [2]. Fatty acids biosynthesized in the plastids are exported for TAG assembly in the ER. The initial step of fatty acid biosynthesis starts with acetyl-CoA carboxylase, which converts acetyl-CoA and bicarbonate into malonyl-CoA. Fatty acids are then biosynthesized with two carbon augmentation via the fatty acid synthase (FAS) complex. Synthesized fatty acids are transported from plastids to ER in forms of acyl-CoA esters and TAG assembly in ER occurs mainly through the eukaryotic phospholipid biosynthetic pathway [3]. The final step of TAG biosynthesis, the conversion of diacylglycerol (DAG) to TAG, either using acyl-CoA or phospholipids, is mediated by diacylglycerol acyltransferase (DGAT) or phosphatidylcholine:diacylglycerol acyltransferase (PDAT) [4,5,6]. The regulatory mechanism of TAG biosynthesis is sophisticated and not understood thoroughly. Elucidation of the molecular mechanism of TAG biosynthesis is thus vital for both fundamental research of plant lipid biochemistry and design of new oil crops with the goal of increasing oil content.
2. WRI1 Acts as a Key Transcriptional Regulator in Governing Plant Oil Biosynthesis
A few transcriptional regulators, including WRINKLED1 (WRI1), are essential for plant oil biosynthesis. Pioneer work identifying WRI1 was performed by the laboratory of Christoph Benning through Arabidopsis mutant screening. In 1998, Benning’s lab discovered wri1-1, the Arabidopsis loss-of-function mutant of AtWRI1, which displays a phenotype of 80% reduced seed oil content compared to the wild-type (WT) [7]. WRI1 is further characterized as a member of the APETALA2 (AP2) transcription factor family [8,9] (also see Figure 1A). In 2002, microarray analysis of developing seeds of WT and wri1-1, conducted in the laboratories of John Ohlrogge and Christoph Benning, indicated that the majority of the genes with decreased expression in the wri1-1 mutant encode enzymes in the glycolytic and fatty acid biosynthetic pathways [10]. Subsequent work has validated numerous genes involved in glycolysis and fatty acid biosynthesis as direct targets of AtWRI1 [11,12] and characterized the AtWRI1 binding motif, the AW-box [12]. Given its importance, WRI1 is thus considered a “master regulator” for transcriptional regulation of TAG biosynthesis [1].
3. Regulators Involved in Mediating the Expression of WRI1
Central seed developmental regulators, LEAFY COTYLEDON1 (LEC1) and LEAFY COTYLEDON2 (LEC2), have been proposed to be key upstream transcription factors, which control the expression of AtWRI1. AtWRI1 expression is elevated in tnp (a gain-of-function LEC1 mutant) [13] and LEC1-overexpression transgenic plants [14], implicating a potential function of LEC1 in activating AtWRI1 expression [15,16]. The molecular and genetic evidence also suggests that LEC2 plays an important role in controlling the expression of AtWRI1. Reduced expression of AtWRI1 was found in a lec2 loss-of-function mutant [11]. Using inducible transgenic LEC2 plants, Baud et al. have revealed that induction of LEC2 activates AtWRI1 expression [11]. The mechanism of LEC1/LEC2 controlling WRI1 expression seems to be conserved in other plant species as well. For instance, the overexpression of soybean LEC2 (GmLEC2) leads to the upregulated expression of soybean WRI1 (GmWRI1) [17]. Expression of maize WRI1 (ZmWRI1) is also elevated in transgenic maize embryos overexpressing maize LEC1 (ZmLEC1). This mechanism is substantiated via co-expression of ZmLEC1 with a promoterZmWRI1:GUS reporter in maize cell culture. ZmLEC1 is able to activate GUS expression and elevate the GUS activity of a promoterZmWRI1:GUS reporter in maize cell culture [18]. Nevertheless, the molecular basis of LEC1/LEC2 binding to the promoter of AtWRI1 has not been elucidated [15,16]. Pelletier et al. recently conducted chromatin immunoprecipitation (ChIP) followed by DNA microarray (ChIP-chip) assays and confirmed that AtWRI1 is a direct target of LEC1 [19].
Molecular and genetic evidence also suggests that the Arabidopsis transcription factor FUSCA3 (FUS3) activates the expression of AtWRI1. Previous comparative microarray results indicated that the expression of AtWRI1 is decreased in a fus3 loss-of-function mutant compared to WT [20]. It has been hypothesized that FUS3 controls AtWRI1 expression in a similar manner as LEC2, based on the functional redundancy between LEC2 and FUS3 [20]. Recently, Perry’s team conducted ChIP-chip assays and provided convincing evidence validating AtWRI1 as a direct target gene of FUS3 [21]. The addition of sucrose activates the expression of AtWRI1 in Arabidopsis seedlings [9]. AtWRI1 is also shown to trigger the expression of some sugar-responsive genes, suggesting a potential role of AtWRI1 in mediating the process of carbon flow to TAG [9]. LEC1 and LEC2 do not affect the expression of FUS3, hence the activation of AtWRI1 (triggered by sucrose treatment) is concluded to be mediated by FUS3 directly [22].
Possible upstream WRI1 regulators, other than LEC1, LEC2, and FUS3, might exist in other plant species. For example, the expression of oil palm LEC1 (EgLEC1), EgLEC2, and EgFUS3 is low in the oil palm mesocarp where EgWRI1 is highly expressed during fruit ripening, raising the speculation that expression of EgWRI1 is regulated by novel regulators [23]. Recent work showed that EgNF-YA3, EgNF-YC2, and EgABI5 are able to bind to the EgWRI1 promoter and activate EgWRI1 expression [24].
The Arabidopsis transcription factor MYB89 has recently been identified as a new regulator, which represses AtWRI1 expression. The overexpression of MYB89 causes reduced expression of AtWRI1, and increased AtWRI1 expression is detected in the myb89 mutant. ChIP experiments further validated that MYB89 binds to the AtWRI1 promoter, suggesting that AtWRI1 is a direct target of MYB89 [25].
4. Molecular Regulatory Mechanism of AtWRI1 Activity
Since the identification of AtWRI1 in 2004 [8], advancements in understanding AtWRI1 function include identification of the AtWRI1 target genes in oil biosynthesis and characterization of the AtWRI1 binding motif. The last five years have seen significantly more publications on WRI1. However, little is known regarding other regulatory mechanisms mediating the activity of WRI1, such as post-translational modifications and interaction with other protein regulators. In addition, functional domains and motifs in WRI1 protein also remain to be characterized.
The first WRI1-interacting partner has been identified through yeast two-hybrid (Y2H) screening. Using CULLIN3-based E3 ligase adaptor BTB/POZMATH 1 (BPM1) as prey led to the identification of AtWRI1 as a BPM1-interacting protein [26]. AtWRI1 interacts with other BPM proteins as well. The AtWRI1 protein is unstable and its assembly with E3 ligase adaptor BPMs mediates its degradation by the 26S proteasome [26]. This study bridges the connection between E3 ligase and plant oil biosynthesis through controlling WRI1 protein stability (Figure 1B).
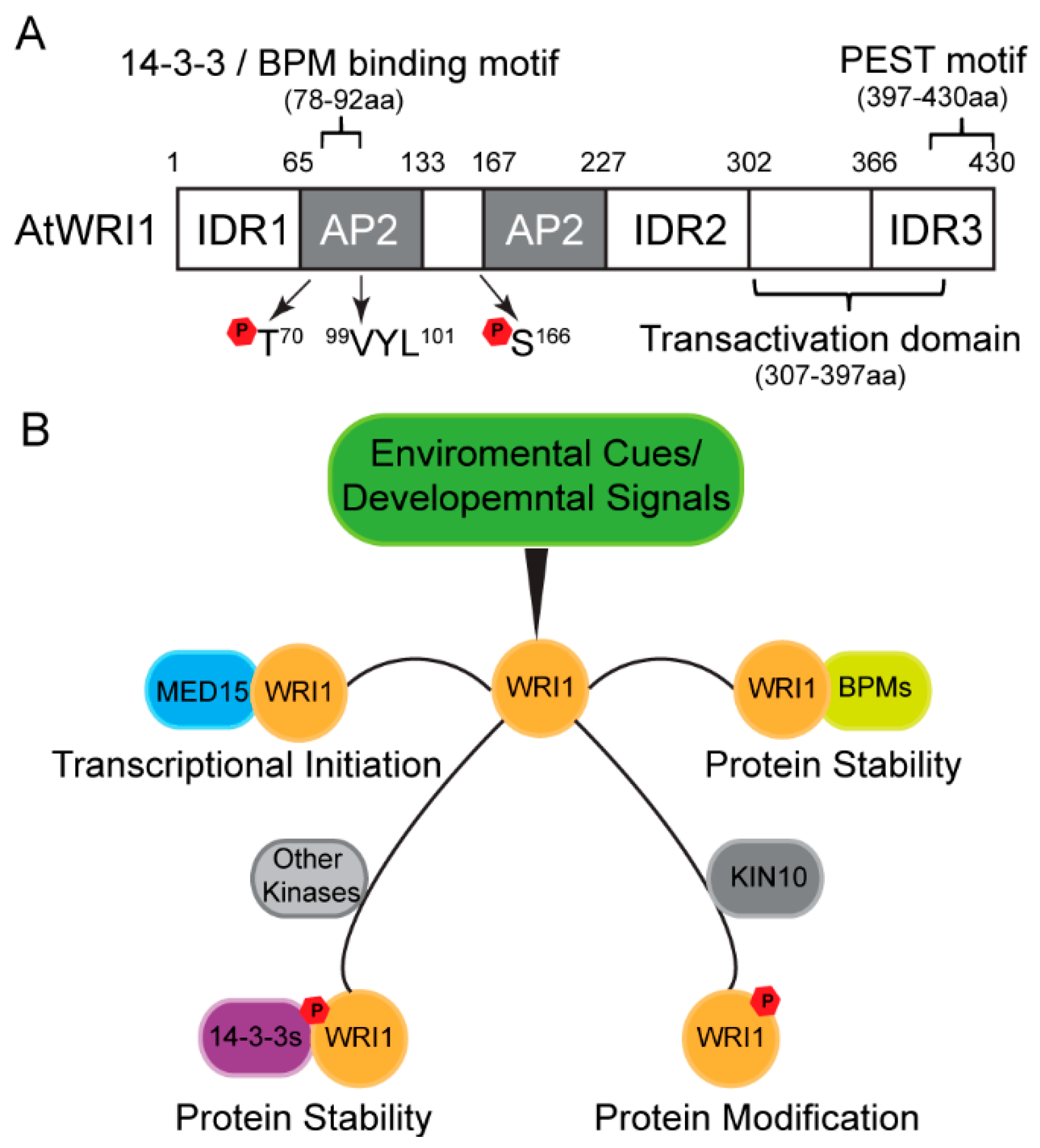
Many proteins do not have a rigid three-dimensional structure, and intrinsic disordered regions (IDRs) have been broadly identified in eukaryotic proteins [27,28,29]. Recent work showed that AtWRI1 possesses three IDRs, as predicted by in silico analysis [30]. Functional characterization led to the identification of a PEST motif (a peptide signal for proteolysis), located in IDR3 of AtWRI1, and a transactivation domain (TAD) [30]. Both the IDR3-PEST motif and TAD are located at the C-terminus without overlapping (Figure 1A). Engineered AtWRI1s with either deletion of IDR3-PEST or mutations in possible phosphorylation sites in IDR3-PEST, results in increased protein stability and enhanced oil production compared to the native form of AtWRI1. Therefore, phosphorylation at the IDR3-PEST motif is proposed as a possible regulatory mechanism for AtWRI1 [30].
Multiple novel AtWRI1-interacting proteins that modulate AtWRI1 activity have recently been identified by several research groups. Ma et al. found that AtWRI1 physically interacts with 14-3-3 proteins in a Y2H and bimolecular fluorescence complementation (BiFC) assay in planta [31]. Overexpression of 14-3-3 increased AtWRI1-regulated oil biosynthesis and enhanced the protein stability and transcriptional activity of AtWRI1 (Figure 1B). Functional characterization revealed that the binding motifs of 14-3-3 and BPM overlap in AtWRI1 protein (Figure 1A). Hence, it is speculated that AtWRI1-14-3-3 interaction blocks BPM interaction with AtWRI1, or detachment of AtWRI1 from BPMs [31,32].
Zhai et al. found that KIN10 kinase physically interacts with AtWRI1 and triggers phosphorylation of AtWRI1 [33] (Figure 1B). AtWRI1 phosphorylation triggered by KIN10 is critical for the degradation of AtWRI1. The AtWRI1 mutant with mutations of two KIN10 phosphorylation residues (T70 and S166) abolishes KIN10-triggered phosphorylation and leads to enhanced protein stability of AtWRI1 [33] (Figure 1A). The proximity of the 14-3-3 binding motif and one KIN10 phosphorylation site in AtWRI1 suggests that these two modifications possibly overlap in the proteasome pathway [33]. Zhai et al. further found that trehalose 6-phosphate (T6P) plays a role in stabilizing the WRI1 protein and enhancing fatty acid biosynthesis by repressing KIN10 activity [34]. AtWRI1 protein level is higher in adg1suc2 double mutant compared to WT, which is speculated via the repression KIN10-mediated phosphorylation on AtWRI1 [35].
The Arabidopsis mediator complex MED15 subunit is another novel candidate regulator, which physically interacts with AtWRI1 in multiple protein-protein interaction assays [36] (Figure 1B). Arabidopsis plants overexpressing MED15 showed upregulation of AtWRI1 target genes in oil biosynthetic pathways. ChIP experiments subsequently demonstrated that MED15 binds to the promoter regions of these AtWRI1 target genes [36]. Nevertheless, transgenic wri1 plants overexpressing MED15 showed the unregulated expression of AtWRI1 target genes, suggesting that MED15 possibly interacts with other transcription factors to control the expression of AtWRI1 target genes [36].
5. WRI1 Orthologs Identified in Various Plant Species
WRI1 orthologs have been identified in a variety of plant species, both monocot and dicot, such as Avena sativa [37], Brassica napus [38], Brachypodium distachyon [39], Camelina sativa [40], Cocos nucifera [41], Cyperus esculentus [37], Elaeis guineensis [23,42], Glycine max [17,43], Gossypium spp [44], Jatropha curcas [45], Persea americana [46], Ricinus communis [47], Solanum tuberosum [37], and Zea mays [18,48]. Expression of AtWRI1 and WRI1 orthologs rescue the reduced-oil phenotypes of wri1 loss-of-function mutants [8,40,42,45,48]. Many WRI1 orthologs are highly expressed in developing seeds, similar to the expression pattern of AtWRI1. However, some WRI1s were found to be highly expressed in non-seed tissues. Expression of EgWRI1 is enhanced in mesocarp (oil-producing tissues) and significantly increased during the fruit ripening process [23]. In addition, PaWRI1 [46] and CeWRI1 [37] display high expression in avocado mesocarp and nutsedge stem tuber, respectively. Protein structural features and functional motifs/domains, that have been characterized in AtWRI1 (e.g., “VYL” [42], IDR [30] and the PEST motif [30]), were found to be conserved in WRI1s identified from other plant species [30,39,40,42,45,49,50].
6. Newly Identified Targets of AtWRI1 That Are Not in Oil Biosynthetic Pathway
AtWRI1 is able to bind to the promoters of PINs (PIN-FORMEDs) at the AW-box, but binds the promoter of GH3.3 (which encodes an enzyme participating in the auxin degradation) via a non-AW-box element, suggesting a role in modulating auxin homeostasis [51]. AtWRI1 is a homolog of CitAP2.10 (a Citrus sinensis AP2 transcription factor involved in (+)-valencene production) and able to activate the expression of C. sinensis Terpene Synthase 1 (CsTPS1) in a dual-luciferase assay [52]. At present, it is unknown how AtWRI1 regulation of these alternative target genes is associated with mediating plant development and growth. Nevertheless, the potential effects of alternative target genes of WRI1 on other plant physiological processes (e.g., auxin homeostasis [51]) needs to be taken into consideration in the application of WRI1 for bioengineering oil production.
7. Applications of WRI1 in Bioengineering of Plant Oil Production
Transgenic plants overexpressing AtWRI1 or WRI1 orthologs have been shown to elevate seed oil content [8,18,38,39,41,45,53]. Overexpression of AtWRI1 and BdWRI1 elevate oil content in leaves of transgenic Arabidopsis and Brachypodium seedlings [39,54]. Using the embryo-preferred OLEOSIN (OLE) promoter to drive ZmWRI1 expression led to significantly elevated seed oil content in transgenic maize, while no oil increase was detected when used the starch endosperm-specific 19 KD ZEIN promoter to drive ZmWRI1 expression [18]. In addition, selection of the FUS3 promoter to drive AtWRI1 expression in transgenic Arabidopsis, aiming to extend oil production during the mid-phase of seed development, is an effective way to enhance seed oil content [55].
Transient overproduction of AtWRI1 or WRI1 orthologs have also been successfully used for producing TAG in tobacco leaves [30,37,56]. The transient co-expression of AtWRI1 and DGAT1 in tobacco leaves has resulted in significantly increased oil content compared to the sole expression of WRI1, suggesting a synergistic effect between WRI1 and DGAT1 [56]. Transient ectopic expression of WRI1 variants, including those with phosphorylation deficient mutations in the IDR3-PEST motif or removal of IDR3-PEST, lead to stabilized WRI1s and increased oil biosynthesis in tobacco leaves compared to overexpression of native WRI1 [30]. Transient co-expression of AtWRI1 and 14-3-3 in tobacco leaves increases AtWRI1 stability and oil production [31]. The AtWRI1K2RK3R mutant (with mutations in ubiquitination target sites) also displays increased protein stability and enhanced oil production compared to native AtWRI1 in a tobacco transient expression assay [33].
In addition, ectopic expression of some transcription factors from other plant species, which are capable of activating WRI1 expression, have been shown to effectively increase oil content in seeds. For example, transgenic Arabidopsis plants overexpressing GmZF351 or GmDREBL display enhanced seed oil contents. GmZF351 or GmDREBL have been found to be able to bind to the AtWRI1 promoter [57,58]. Ectopic expression of ZmLEC1 significantly enhances seed oil contents in Arabidopsis, Camelina, and maize [18,59].
Unusual fatty acids, e.g., hydroxy fatty acids (HFAs), have high value for industrial uses due to their special physical and chemical properties. However, transgenic plants overexpressing a hydroxylase gene accumulate a low amount of HFAs, while the seed oil content is decreased [60,61]. The reduction in seed oil was thought to be due to feedback inhibition of fatty acid synthesis [61]. To overcome this bottleneck, Adhikari et al. generated transgenic Arabidopsis plants co-expressing WRI1 and a gene encoding a castor fatty acid hydroxylase, RcFAH12. The proportion of HFA and whole seed oil content in the transgenic plants significantly increased, suggesting WRI1 effectively rescues the feedback inhibition of fatty acid biosynthesis due to ectopic expression of a hydroxylase gene [62].
8. Future Perspectives
Since the identification of the wri1-1 mutant in 1998 [7], the work in the past two decades confirms the important role of WRI1 in transcriptional control of plant fatty acid biosynthesis. However, there are still important questions to be addressed. For instance, the molecular mechanism regulating WRI1 expression is still unclear. How seed development regulators, such as LEC1, LEC2, and FUS3, control WRI1 expression also requires further elucidation. Alternative regulators may be involved in modulating WRI1 expression in response to developmental or environmental signals. Transcription factors mediating the expression of EgWRI1 were recently identified in oil palm [24]. Whether regulation of WRI1 expression by these regulators is conserved in other plant species requires further investigation. Deeper investigations of the WRI1 promoters from different plant species and the multi-layer gene regulatory network will address this question, as well as other questions related to the broader biological functions of WRI1.
Efforts have been initiated to connect the protein structural features with the molecular function of WRI1. Characterizations of the IDR, TAD, PEST motif, 14-3-3 and BPM binding motif, and KIN10 phosphorylation sites in AtWRI1 [30,31,33] are advancing our understanding of the WRI1 regulatory mechanism at the molecular level. In addition to more detailed characterization of the functional domains and motifs in the WRI1 protein, future work should focus on investigating WRI1 dynamics fine-tuned by protein-protein interactions and in response to cellular signals and environmental cues. Potential phosphorylation residues have been identified in AtWRI1 [30] and some have been experimentally shown to be vital for the function of AtWRI1 [30,33]. Continuing discovery and characterization of novel kinases involved in modifying AtWRI1 will be necessary. In addition, it has been speculated that phosphorylation might play dual roles in modulating the stability of AtWRI1 in response to different developmental signals during plant embryo development [31]. Thus, it will be interesting to see how modifications mediated by different upstream kinases and altered signals might be involved in the regulation of AtWRI1.
AtWRI1 has been recently found to bind the AW-box or new binding elements in promoters of targets not involved in glycolysis and fatty acid biosynthesis [51]. Recent work by Liu et al. found that AtWRI1 directly upregulates the expression of genes encoding BIOTIN ATTACHMENT DOMAIN-CONTAINING (BADC) proteins, which are responsible for inhibiting fatty acid biosynthesis [63]. This work might suggest a novel mechanism in WRI1 regulation of fatty acid synthesis. A deeper understanding of WRI1 on mediating the gene expression of the targets, which are not linked to oil production, might contribute to overcoming the undesirable effects associated with overexpressing WRI1s in transgenic plants [8,15,39]. Alternative approaches, such as protein engineering and genome editing, may be used to enhance WRI1 binding specificities to the target genes in oil biosynthetic pathways.
Author Contributions
Q.K. and W.M. conceived the ideas, prepared the figures and wrote the first draft. L.Y. and W.M. reviewed and edited the figures and manuscript. All authors read and approved the final version of the manuscript.
Funding
This research was funded by a Nanyang Technological University Startup grant and a Ministry of Education (MOE) of Singapore Tier 1 to W.M. (grant number: 2018-T1-002-019).
Acknowledgments
We apologize to all authors whose important work is not cited because of space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.B.; Chapman, K.D. The seeds of green energy: Expanding the contribution of plant oils as biofuels. Biochemist 2011, 33, 34–38. [Google Scholar] [Green Version]
- Bates, P.D.; Durrett, T.P.; Ohlrogge, J.B.; Pollard, M. Analysis of acyl fluxes through multiple pathways of triacylglycerol synthesis in developing soybean embryos. Plant Physiol. 2009, 150, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, A.; Stahl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Wei, Y.; Jako, C.; Kumar, A.; Selvaraj, G.; Taylor, D.C. The Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol acyltransferase gene. Plant J. 1999, 19, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Focks, N.; Benning, C. wrinkled1: A novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism. Plant Physiol. 1998, 118, 91–101. [Google Scholar] [CrossRef]
- Cernac, A.; Benning, C. WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 2004, 40, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Masaki, T.; Mitsui, N.; Tsukagoshi, H.; Nishii, T.; Morikami, A.; Nakamura, K. ACTIVATOR of Spomin::LUC1/WRINKLED1 of Arabidopsis thaliana transactivates sugar-inducible promoters. Plant Cell Physiol. 2005, 46, 547–556. [Google Scholar] [CrossRef]
- Ruuska, S.A.; Girke, T.; Benning, C.; Ohlrogge, J.B. Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 2002, 14, 1191–1206. [Google Scholar] [CrossRef]
- Baud, S.; Mendoza, M.S.; To, A.; Harscoet, E.; Lepiniec, L.; Dubreucq, B. WRINKLED1 specifies the regulatory action of LEAFY COTYLEDON2 towards fatty acid metabolism during seed maturation in Arabidopsis. Plant J. 2007, 50, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Maeo, K.; Tokuda, T.; Ayame, A.; Mitsui, N.; Kawai, T.; Tsukagoshi, H.; Ishiguro, S.; Nakamura, K. An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis. Plant J. 2009, 60, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Casson, S.A.; Lindsey, K. The turnip mutant of Arabidopsis reveals that LEAFY COTYLEDON1 expression mediates the effects of auxin and sugars to promote embryonic cell identity. Plant Physiol. 2006, 142, 526–541. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Tan, H.; Zheng, Q.; Fu, F.; Liang, Y.; Zhang, J.; Yang, X.; Wang, T.; Chong, K.; Wang, X.J.; et al. LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Marchive, C.; Nikovics, K.; To, A.; Lepiniec, L.; Baud, S. Transcriptional regulation of fatty acid production in higher plants: Molecular bases and biotechnological outcomes. Eur. J. Lipid Sci. Technol. 2014, 116, 1332–1343. [Google Scholar]
- Santos-Mendoza, M.; Dubreucq, B.; Baud, S.; Parcy, F.; Caboche, M.; Lepiniec, L. Deciphering gene regulatory networks that control seed development and maturation in Arabidopsis. Plant J. 2008, 54, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Manan, S.; Ahmad, M.Z.; Zhang, G.; Chen, B.; Haq, B.U.; Yang, J.; Zhao, J. Soybean LEC2 Regulates Subsets of Genes Involved in Controlling the Biosynthesis and Catabolism of Seed Storage Substances and Seed Development. Front. Plant Sci. 2017, 8, 1604. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Allen, W.B.; Zheng, P.; Li, C.; Glassman, K.; Ranch, J.; Nubel, D.; Tarczynski, M.C. Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol. 2010, 153, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.M.; Kwong, R.W.; Park, S.; Le, B.H.; Baden, R.; Cagliari, A.; Hashimoto, M.; Munoz, M.D.; Fischer, R.L.; Goldberg, R.B.; et al. LEC1 sequentially regulates the transcription of genes involved in diverse developmental processes during seed development. Proc. Natl. Acad. Sci. USA 2017, 114, E6710–E6719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, A.; Kagaya, Y.; Usui, H.; Hobo, T.; Takeda, S.; Hattori, T. Diverse roles and mechanisms of gene regulation by the Arabidopsis seed maturation master regulator FUS3 revealed by microarray analysis. Plant Cell Physiol. 2010, 51, 2031–2046. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Perry, S.E. Identification of direct targets of FUSCA3, a key regulator of Arabidopsis seed development. Plant Physiol. 2013, 161, 1251–1264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cao, X.; Jia, Q.; Ohlrogge, J. FUSCA3 activates triacylglycerol accumulation in Arabidopsis seedlings and tobacco BY2 cells. Plant J. 2016, 88, 95–107. [Google Scholar] [CrossRef]
- Bourgis, F.; Kilaru, A.; Cao, X.; Ngando-Ebongue, G.F.; Drira, N.; Ohlrogge, J.B.; Arondel, V. Comparative transcriptome and metabolite analysis of oil palm and date palm mesocarp that differ dramatically in carbon partitioning. Proc. Natl. Acad. Sci. USA 2011, 108, 12527–12532. [Google Scholar] [CrossRef] [Green Version]
- Yeap, W.C.; Lee, F.C.; Shabari Shan, D.K.; Musa, H.; Appleton, D.R.; Kulaveerasingam, H. WRI1-1, ABI5, NF-YA3 and NF-YC2 increase oil biosynthesis in coordination with hormonal signaling during fruit development in oil palm. Plant J. 2017, 91, 97–113. [Google Scholar] [CrossRef]
- Li, D.; Jin, C.; Duan, S.; Zhu, Y.; Qi, S.; Liu, K.; Gao, C.; Ma, H.; Zhang, M.; Liao, Y.; et al. MYB89 Transcription Factor Represses Seed Oil Accumulation. Plant Physiol. 2017, 173, 1211–1225. [Google Scholar] [CrossRef]
- Chen, L.; Lee, J.H.; Weber, H.; Tohge, T.; Witt, S.; Roje, S.; Fernie, A.R.; Hellmann, H. Arabidopsis BPM Proteins Function as Substrate Adaptors to a CULLIN3-Based E3 Ligase to Affect Fatty Acid Metabolism in Plants. Plant Cell 2013, 25, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, I.; Guittard-Crilat, E.; Maldiney, R.; Habricot, Y.; Lignon, S.; Lebrun, R.; Miginiac, E.; Ruelland, E.; Jeannette, E.; Lebreton, S. The intrinsically disordered C-terminal region of Arabidopsis thaliana TCP8 transcription factor acts both as a transactivation and self-assembly domain. Mol. Biosyst. 2013, 9, 2282–2295. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef]
- Kragelund, B.B.; Jensen, M.K.; Skriver, K. Order by disorder in plant signaling. Trends Plant Sci. 2012, 17, 625–632. [Google Scholar] [CrossRef]
- Ma, W.; Kong, Q.; Grix, M.; Mantyla, J.J.; Yang, Y.; Benning, C.; Ohlrogge, J.B. Deletion of a C-terminal intrinsically disordered region of WRINKLED1 affects its stability and enhances oil accumulation in Arabidopsis. Plant J. 2015, 83, 864–874. [Google Scholar] [CrossRef]
- Ma, W.; Kong, Q.; Mantyla, J.J.; Yang, Y.; Ohlrogge, J.B.; Benning, C. 14-3-3 protein mediates plant seed oil biosynthesis through interaction with AtWRI1. Plant J. 2016, 88, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Ma, W. WRINKLED1 as a novel 14-3-3 client: Function of 14-3-3 proteins in plant lipid metabolism. Plant Signal. Behav. 2018, 13, e1482176. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Liu, H.; Shanklin, J. Phosphorylation of WRINKLED1 by KIN10 Results in Its Proteasomal Degradation, Providing a Link between Energy Homeostasis and Lipid Biosynthesis. Plant Cell 2017, 29, 871–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Z.; Keereetaweep, J.; Liu, H.; Feil, R.; Lunn, J.E.; Shanklin, J. Trehalose 6-Phosphate Positively Regulates Fatty Acid Synthesis by Stabilizing WRINKLED1. Plant Cell 2018, 30, 2616–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Z.; Liu, H.; Xu, C.; Shanklin, J. Sugar Potentiation of Fatty Acid and Triacylglycerol Accumulation. Plant Physiol. 2017, 175, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jang, I.C.; Chua, N.H. The Mediator Complex MED15 Subunit Mediates Activation of Downstream Lipid-Related Genes by the WRINKLED1 Transcription Factor. Plant Physiol. 2016, 171, 1951–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimberg, A.; Carlsson, A.S.; Marttila, S.; Bhalerao, R.; Hofvander, P. Transcriptional transitions in Nicotiana benthamiana leaves upon induction of oil synthesis by WRINKLED1 homologs from diverse species and tissues. BMC Plant Biol. 2015, 15, 192. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hua, W.; Zhan, G.; Wei, F.; Wang, X.; Liu, G.; Wang, H. Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of wri1-like gene from Brassica napus. Plant Physiol. Biochem. 2010, 48, 9–15. [Google Scholar] [CrossRef]
- Yang, Y.; Munz, J.; Cass, C.; Zienkiewicz, A.; Kong, Q.; Ma, W.; Sedbrook, J.; Benning, C. Ectopic Expression of WRINKLED1 Affects Fatty Acid Homeostasis in Brachypodium distachyon Vegetative Tissues. Plant Physiol. 2015, 169, 1836–1847. [Google Scholar] [CrossRef]
- An, D.; Kim, H.; Ju, S.; Go, Y.S.; Kim, H.U.; Suh, M.C. Expression of Camelina WRINKLED1 Isoforms Rescue the Seed Phenotype of the Arabidopsis wri1 Mutant and Increase the Triacylglycerol Content in Tobacco Leaves. Front. Plant Sci. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Ye, R.; Gao, L.; Zhang, L.; Wang, R.; Mao, T.; Zheng, Y.; Li, D.; Lin, Y. Characterization and Ectopic Expression of CoWRI1, an AP2/EREBP Domain-Containing Transcription Factor from Coconut (Cocos nucifera L.) Endosperm, Changes the Seeds Oil Content in Transgenic Arabidopsis thaliana and Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 63. [Google Scholar] [CrossRef]
- Ma, W.; Kong, Q.; Arondel, V.; Kilaru, A.; Bates, P.D.; Thrower, N.A.; Benning, C.; Ohlrogge, J.B. Wrinkled1, a ubiquitous regulator in oil accumulating tissues from Arabidopsis embryos to oil palm mesocarp. PLoS ONE 2013, 8, e68887. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, M.; Li, S.; Sun, L.; Wang, W.; Cai, C.; Dierking, E.C.; Ma, J. Plasticity and innovation of regulatory mechanisms underlying seed oil content mediated by duplicated genes in the palaeopolyploid soybean. Plant J. 2017, 90, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Ye, J.; Geng, Y.F.; Sun, Y.W.; Gao, S.Q.; Zhang, B.P.; Chen, W.; Chua, N.H. Dissecting functions of KATANIN and WRINKLED1 in cotton fiber development by virus-induced gene silencing. Plant Physiol. 2012, 160, 738–748. [Google Scholar] [CrossRef]
- Ye, J.; Wang, C.; Sun, Y.; Qu, J.; Mao, H.; Chua, N.H. Overexpression of a Transcription Factor Increases Lipid Content in a Woody Perennial Jatropha curcas. Front. Plant Sci. 2018, 9, 1479. [Google Scholar] [CrossRef] [Green Version]
- Kilaru, A.; Cao, X.; Dabbs, P.B.; Sung, H.J.; Rahman, M.M.; Thrower, N.; Zynda, G.; Podicheti, R.; Ibarra-Laclette, E.; Herrera-Estrella, L.; et al. Oil biosynthesis in a basal angiosperm: Transcriptome analysis of Persea Americana mesocarp. BMC Plant Biol. 2015, 15, 203. [Google Scholar] [CrossRef]
- Tajima, D.; Kaneko, A.; Sakamoto, M.; Ito, Y.; Hue, N.T.; Miyazaki, M.; Ishibashi, Y.; Yuasa, T.; Iwaya-Inoue, M. Wrinkled 1 (WRI1) Homologs, AP2-Type Transcription Factors Involving Master Regulation of Seed Storage Oil Synthesis in Castor Bean (Ricinus communis L.). Am. J. Plant Sci. 2013, 4, 333–339. [Google Scholar] [CrossRef]
- Pouvreau, B.; Baud, S.; Vernoud, V.; Morin, V.; Py, C.; Gendrot, G.; Pichon, J.P.; Rouster, J.; Paul, W.; Rogowsky, P.M. Duplicate maize Wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis. Plant Physiol. 2011, 156, 674–686. [Google Scholar] [CrossRef]
- Kong, Q.; Ma, W. WRINKLED1 transcription factor: How much do we know about its regulatory mechanism? Plant Sci. 2018, 272, 153–156. [Google Scholar] [CrossRef]
- Tang, T.; Du, C.; Song, H.; Aziz, U.; Wang, L.; Zhao, C.; Zhang, M. Genome-wide analysis reveals the evolution and structural features of WRINKLED1 in plants. Mol. Genet. Genom. 2019, 294, 329–341. [Google Scholar] [CrossRef]
- Kong, Q.; Ma, W.; Yang, H.; Ma, G.; Mantyla, J.J.; Benning, C. The Arabidopsis WRINKLED1 transcription factor affects auxin homeostasis in roots. J. Exp. Bot. 2017, 68, 4627–4634. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.L.; Yin, X.R.; Zhang, B.; Xie, X.L.; Jiang, Q.; Grierson, D.; Chen, K.S. CitAP2.10 activation of the terpene synthase CsTPS1 is associated with the synthesis of (+)-valencene in ‘Newhall’ orange. J. Exp. Bot. 2016, 67, 4105–4115. [Google Scholar] [CrossRef]
- An, D.; Suh, M.C. Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content in Camelina sativa. Plant Biotechnol. Rep. 2015, 9, 137–148. [Google Scholar] [CrossRef]
- Durrett, T.P.; Weise, S.E.; Benning, C. Increasing the energy density of vegetative tissues by diverting carbon from starch to oil biosynthesis in transgenic Arabidopsis. Plant Biotechnol. J. 2011, 9, 874–883. [Google Scholar] [CrossRef]
- Kanai, M.; Mano, S.; Kondo, M.; Hayashi, M.; Nishimura, M. Extension of oil biosynthesis during the mid-phase of seed development enhances oil content in Arabidopsis seeds. Plant Biotechnol. J. 2016, 14, 1241–1250. [Google Scholar] [CrossRef]
- Vanhercke, T.; El Tahchy, A.; Shrestha, P.; Zhou, X.R.; Singh, S.P.; Petrie, J.R. Synergistic effect of WRI1 and DGAT1 coexpression on triacylglycerol biosynthesis in plants. FEBS Lett. 2013, 587, 364–369. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.T.; Lu, X.; Song, Q.X.; Chen, H.W.; Wei, W.; Tao, J.J.; Bian, X.H.; Shen, M.; Ma, B.; Zhang, W.K.; et al. Selection for a Zinc-Finger Protein Contributes to Seed Oil Increase during Soybean Domestication. Plant Physiol. 2017, 173, 2208–2224. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Lu, X.; Zhao, F.Y.; Li, Q.T.; Niu, S.L.; Wei, W.; Zhang, W.K.; Ma, B.; Chen, S.Y.; Zhang, J.S. Soybean GmDREBL Increases Lipid Content in Seeds of Transgenic Arabidopsis. Sci. Rep. 2016, 6, 34307. [Google Scholar] [CrossRef]
- Zhu, Y.; Xie, L.; Chen, G.Q.; Lee, M.Y.; Loque, D.; Scheller, H.V. A transgene design for enhancing oil content in Arabidopsis and Camelina seeds. Biotechnol. Biofuels 2018, 11, 46. [Google Scholar] [CrossRef]
- Bates, P.D.; Browse, J. The pathway of triacylglycerol synthesis through phosphatidylcholine in Arabidopsis produces a bottleneck for the accumulation of unusual fatty acids in transgenic seeds. Plant J. 2011, 68, 387–399. [Google Scholar] [CrossRef]
- Bates, P.D.; Johnson, S.R.; Cao, X.; Li, J.; Nam, J.W.; Jaworski, J.G.; Ohlrogge, J.B.; Browse, J. Fatty acid synthesis is inhibited by inefficient utilization of unusual fatty acids for glycerolipid assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, N.D.; Bates, P.D.; Browse, J. WRINKLED1 Rescues Feedback Inhibition of Fatty Acid Synthesis in Hydroxylase-Expressing Seeds. Plant Physiol. 2016, 171, 179–191. [Google Scholar] [CrossRef]
- Liu, H.; Zhai, Z.; Kuczynski, K.; Keereetaweep, J.; Schwender, J.; Shanklin, J. WRINKLED1 regulates BIOTIN ATTACHMENT DOMAIN-CONTAINING proteins that inhibit fatty acid synthesis. Plant Physiol. 2019. [Google Scholar] [CrossRef]
Figure 1.
Arabidopsis WRINKLED1 (AtWRI1) structural features and regulatory mechanism of AtWRI1-mediated oil biosynthesis. (A) Schematic diagram of domains and motifs of AtWRI1, including two APETALA2 (AP2) domains, three intrinsically disordered regions (IDRs), a functional motif of “VYL”, the transactivation domain (TAD), the binding motifs of 14-3-3 and E3 ligase adaptor (BPM), PEST degradation signal sequence, and the KIN10 phosphorylation sites. (B) Post-translational regulatory mechanisms for AtWRI1 mediated by phosphorylation and newly identified interacting regulators, including Mediator Subunit 15 (MED15), BPMs, and kinase KIN10. The regulators modulate the protein stability and transcriptional activity of AtWRI1 via interactions and modifications, which in turn alters the gene expression of AtWRI1 targets.
Figure 1.
Arabidopsis WRINKLED1 (AtWRI1) structural features and regulatory mechanism of AtWRI1-mediated oil biosynthesis. (A) Schematic diagram of domains and motifs of AtWRI1, including two APETALA2 (AP2) domains, three intrinsically disordered regions (IDRs), a functional motif of “VYL”, the transactivation domain (TAD), the binding motifs of 14-3-3 and E3 ligase adaptor (BPM), PEST degradation signal sequence, and the KIN10 phosphorylation sites. (B) Post-translational regulatory mechanisms for AtWRI1 mediated by phosphorylation and newly identified interacting regulators, including Mediator Subunit 15 (MED15), BPMs, and kinase KIN10. The regulators modulate the protein stability and transcriptional activity of AtWRI1 via interactions and modifications, which in turn alters the gene expression of AtWRI1 targets.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kong, Q.; Yuan, L.; Ma, W. WRINKLED1, a “Master Regulator” in Transcriptional Control of Plant Oil Biosynthesis. Plants 2019, 8, 238. https://doi.org/10.3390/plants8070238
AMA Style
Kong Q, Yuan L, Ma W. WRINKLED1, a “Master Regulator” in Transcriptional Control of Plant Oil Biosynthesis. Plants. 2019; 8(7):238. https://doi.org/10.3390/plants8070238
Chicago/Turabian StyleKong, Que, Ling Yuan, and Wei Ma. 2019. "WRINKLED1, a “Master Regulator” in Transcriptional Control of Plant Oil Biosynthesis" Plants 8, no. 7: 238. https://doi.org/10.3390/plants8070238
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.