Betaine Improves Intestinal Functions by Enhancing Digestive Enzymes, Ameliorating Intestinal Morphology, and Enriching Intestinal Microbiota in High-salt stressed Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sampling
2.3. Osmolarity and Electrolytes Assays
2.4. Plasma Endocrine Hormone Assays
2.5. Digestive Enzymes Assays
2.6. DNA Isolation and 16S rRNA Gene Sequencing
2.7. Data Processing
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Osmolarity and Electrolytes
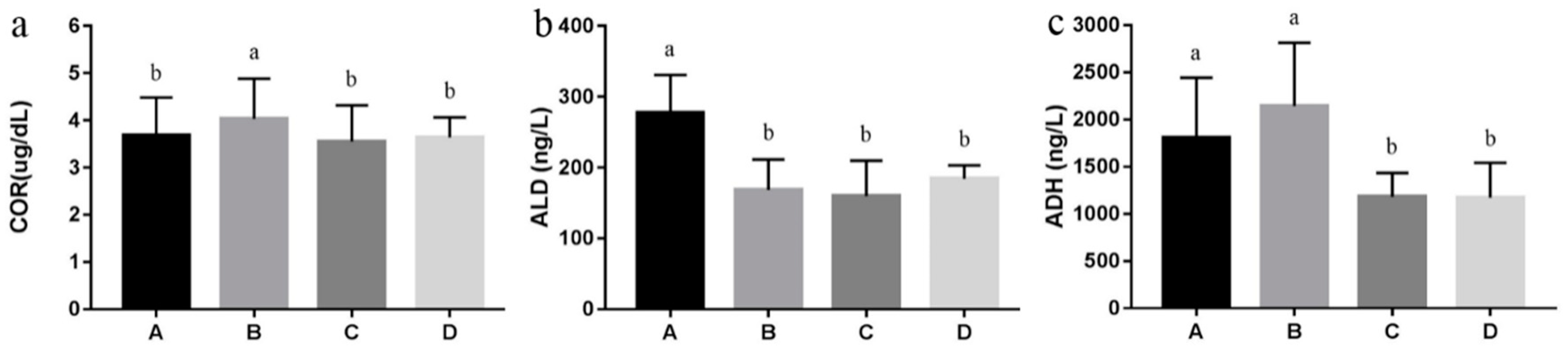
3.3. Endocrine Hormones
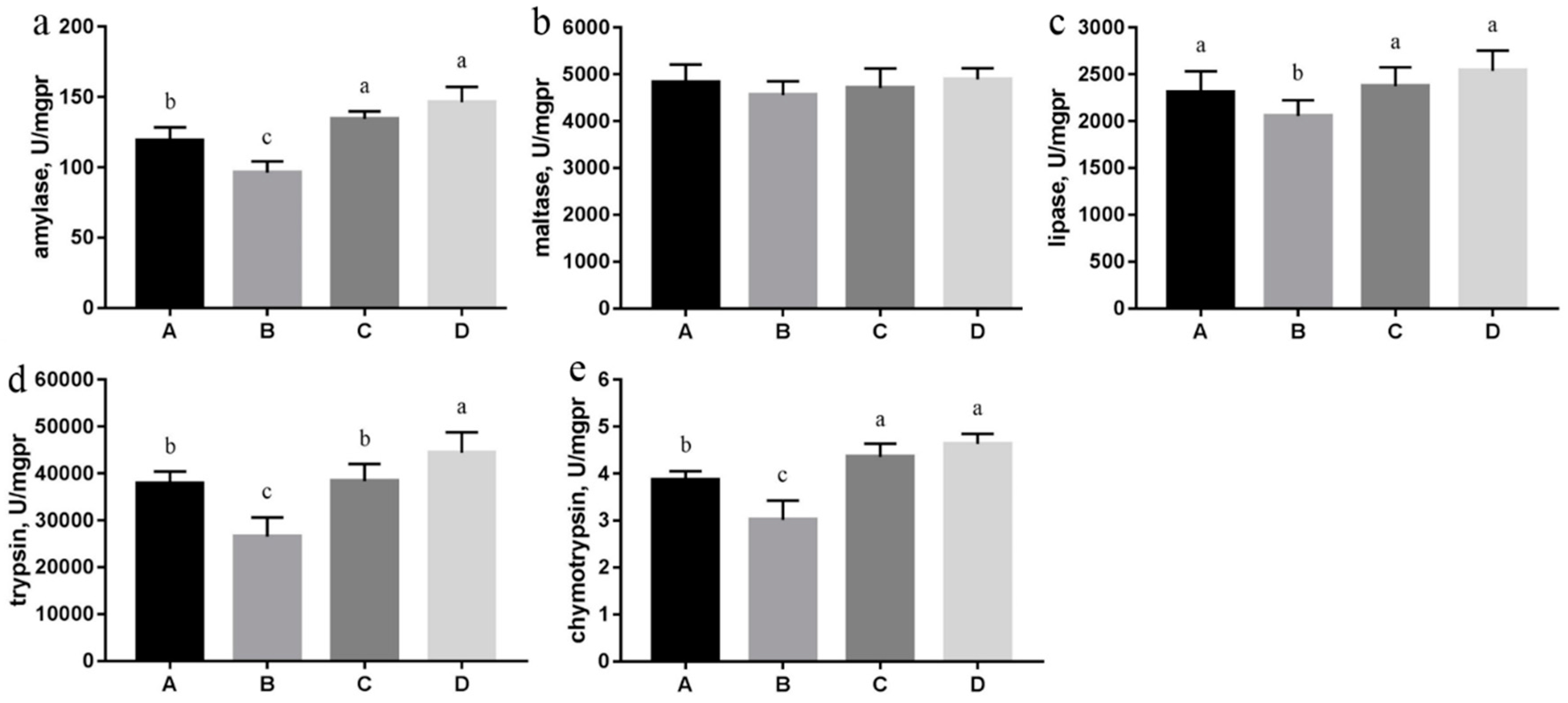
3.4. Digestive Enzyme Activities
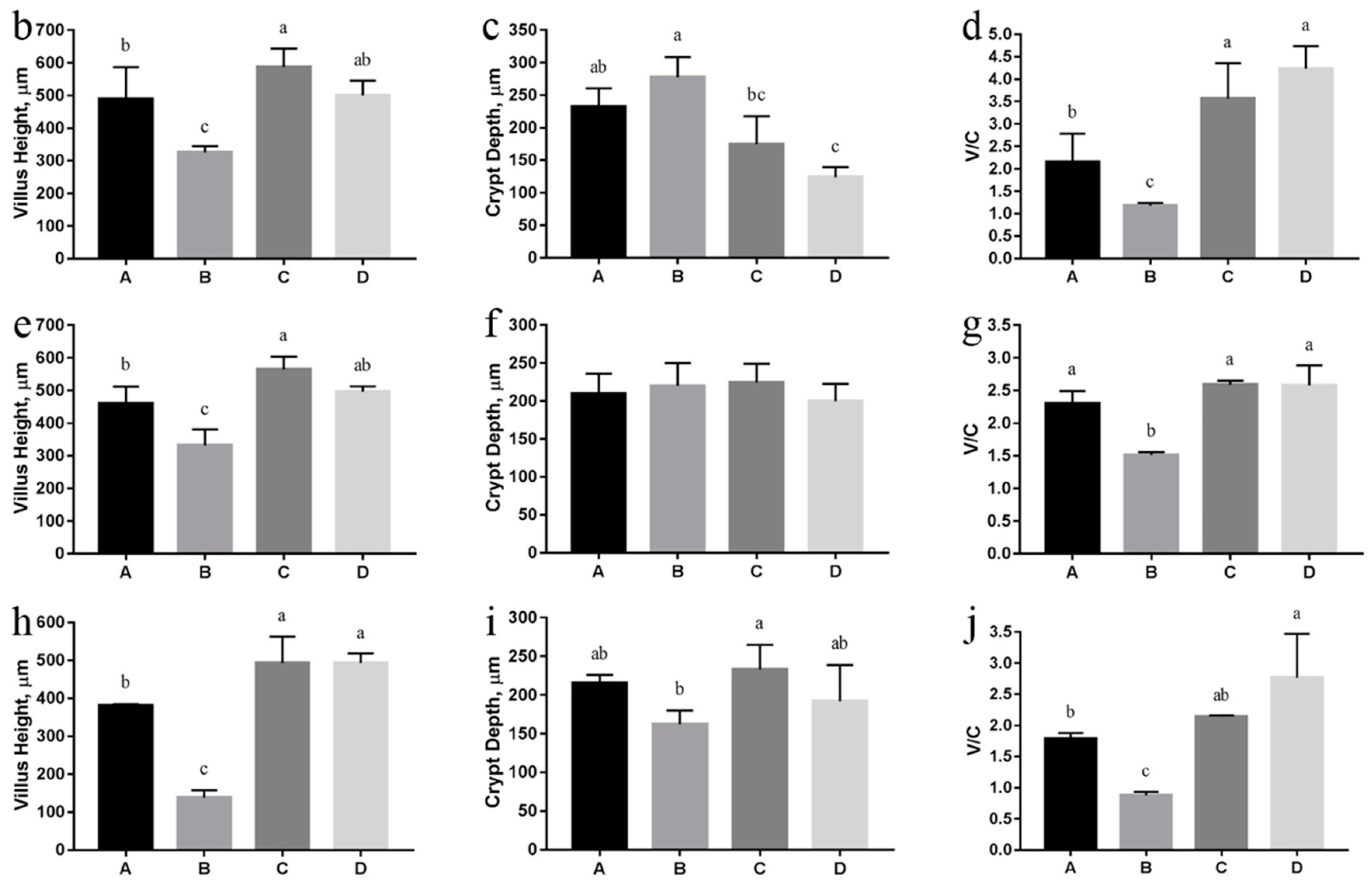
3.5. Intestinal Morphology
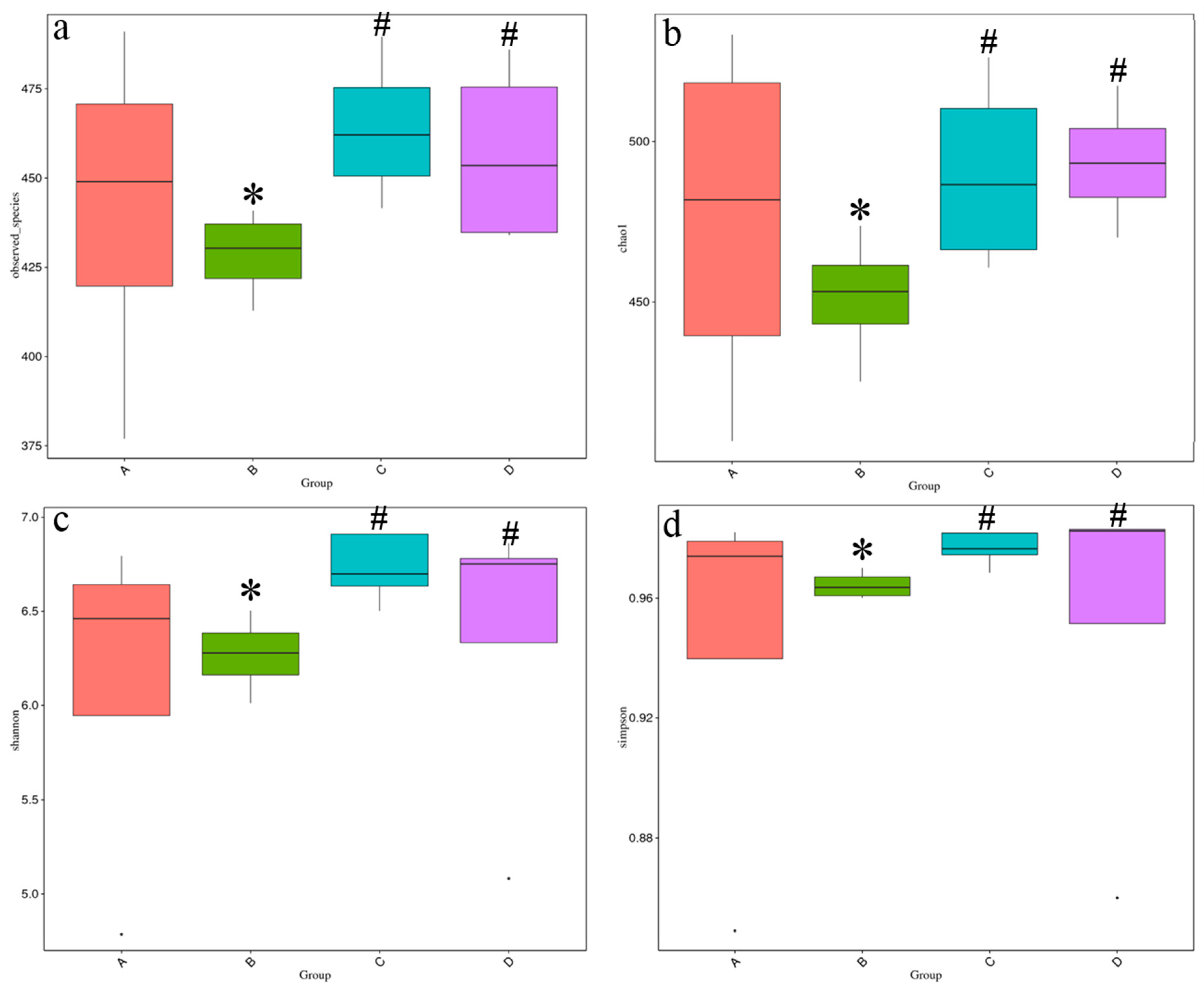
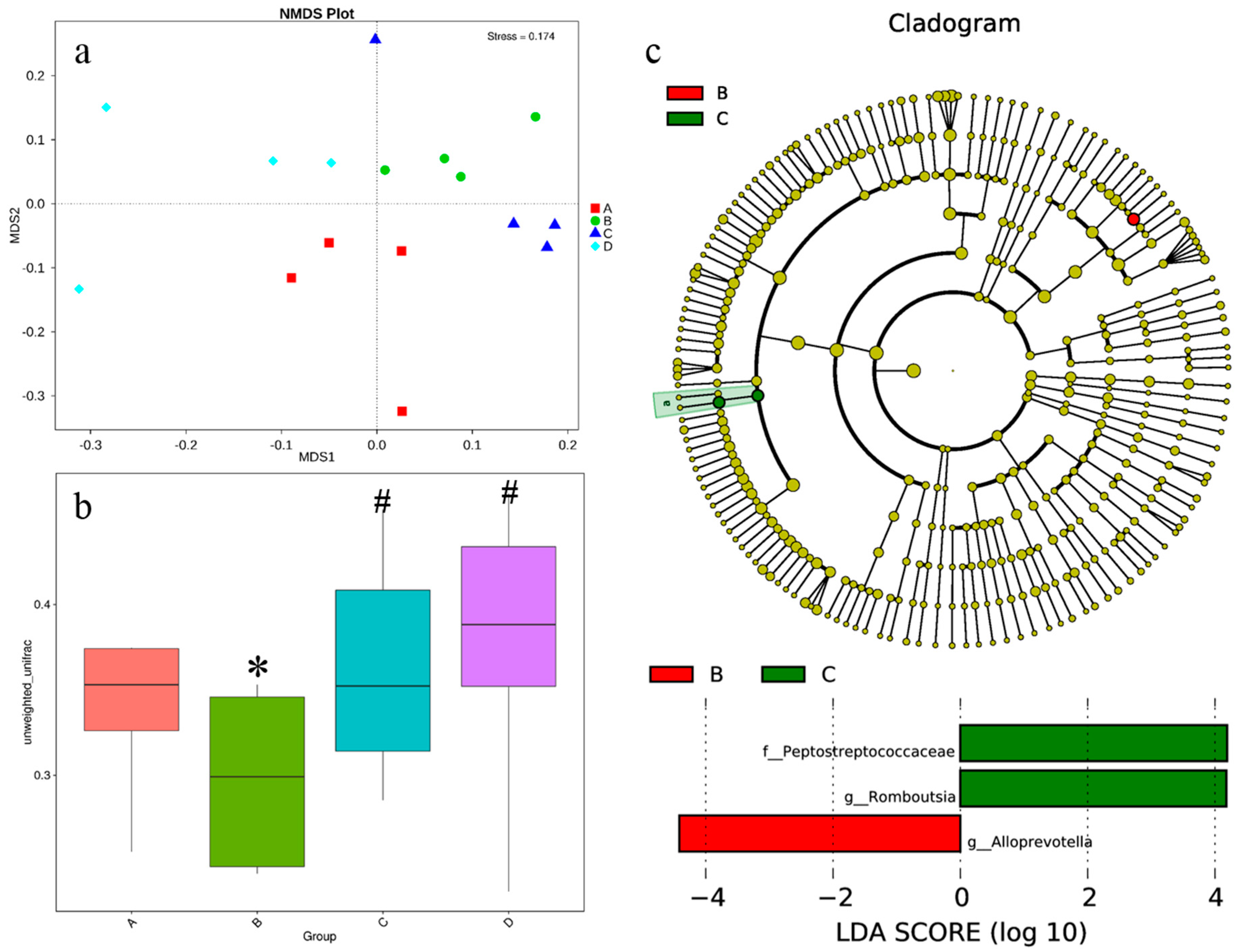
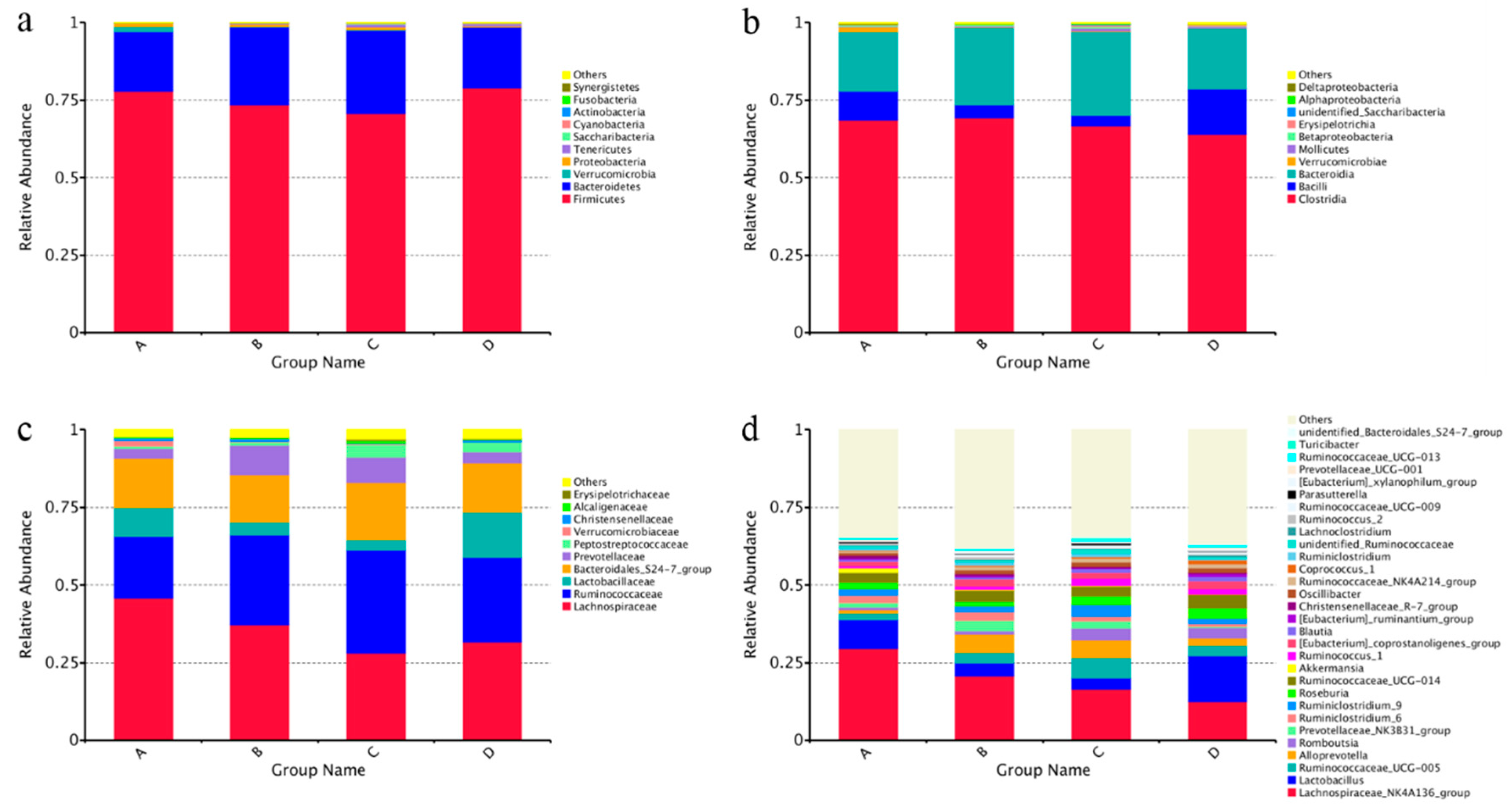
3.6. Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Zwart, F.J.; Slow, S.; Payne, R.J.; Lever, M.; George, P.M.; Gerrard, J.A.; Chambers, S.T. Glycine betaine and glycine betaine analogues in common foods. Food Chem. 2003, 83, 197–204. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Mar, M.; Howe, J.C.; Holden, J.M. Concentrations of choline-containing compounds and betaine in common foods. J. Nutr. 2003, 133, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Craig, S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Huang, D.; Hu, Q.; Wu, J.; Wang, Y.; Feng, J. Betaine alleviates hepatic lipid accumulation via enhancing hepatic lipid export and fatty acid oxidation in rats fed with a high-fat diet. Br. J. Nutr. 2015, 113, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A. Betaine: An old therapy for a new scourge. Am. J. Gastroenterol. 2001, 96, 2534–2536. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L. Nonalcoholic fatty liver disease: Relationship to insulin sensitivity and oxidative stress. Treatment approaches using vitamin E, magnesium, and betaine. Altern. Med. Rev. J. Clin. Ther. 2002, 7, 276–291. [Google Scholar]
- Tokar, J.L.; Berg, C.L. Therapeutic Options in Nonalcoholic Fatty Liver Disease. Curr. Treat. Opt. Gastroenterol. 2002, 5, 425–436. [Google Scholar] [CrossRef]
- Day, C.R.; Kempson, S.A. Betaine chemistry, roles, and potential use in liver disease. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1860, 1098–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappes, R.M.; Kempf, B.; Bremer, E. Three transport systems for the osmoprotectant glycine betaine operate in Bacilluss subtilis: Characterization of OpuD. J. Bacteriol. 1996, 178, 5071–5079. [Google Scholar] [CrossRef] [PubMed]
- Csonka, L.N. Physiological and genetic responses of bacteria to osmotic-stress. Microbiol. Rev. 1989, 53, 121–147. [Google Scholar] [PubMed]
- Yancey, P.H. Living with Water Stress: Evolution of Osmolyte Systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.H.; Wunz, T.M.; Silva, A.L. Betaine transport in the gill of a marine mussel, mytilus-californianus. Am. J. Physiol. 1992, 263, r226–r232. [Google Scholar] [CrossRef] [PubMed]
- Dawson, K.M.; Baltz, J.M. Organic osmolytes and embryos: Substrates of the Gly and beta transport systems protect mouse zygotes against the effects of raised osmolarity. Biol. Reprod. 1997, 56, 1550–1558. [Google Scholar] [CrossRef] [PubMed]
- Oyaas, K.; Ellingsen, T.E.; Dyrset, N.; Levine, D.W. Transport of osmoprotective compounds in hybridoma cells exposed to hyperosmotic stress. Cytotechnology 1995, 17, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Weik, C.; Warskulat, U.; Bode, J.; Peters-Regehr, T.; Haussinger, D. Compatible organic osmolytes in rat liver sinusoidal endothelial cells. Hepatology 1998, 27, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Wettstein, M.; Weik, C.; Holneicher, C.; Haussinger, D. Betaine as an osmolyte in rat liver: Metabolism and cell-to-cell interactions. Hepatology 1998, 27, 787–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters-Regehr, T.; Bode, J.G.; Kubitz, R.; Haussinger, D. Organic osmolyte transport in quiescent and activated rat hepatic stellate cells (Ito cells). Hepatology 1999, 29, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Petronini, P.G.; De Angelis, E.M.; Borghetti, P.; Borghetti, A.F.; Wheeler, K.P. Modulation by betaine of cellular responses to osmotic stress. Biochem. J. 1992, 282, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales, J. A meta-analysis of the effects of dietary betaine supplementation on finishing performance and carcass characteristics of pigs. Anim. Feed Sci. Technol. 2011, 165, 68–78. [Google Scholar] [CrossRef]
- Kidd, M.T.; Garlich, P. Nutritional and osmoregulatory functions of betaine. World Poult. Sci. J. 1997, 53, 125–139. [Google Scholar] [CrossRef]
- Ratriyanto, A.; Mosenthin, R.; Bauer, E.; Eklund, M. Metabolic, osmoregulatory and nutritional functions of betaine in monogastric animals. Asian Australas. J. Anim. Sci. 2009, 22, 1461–1476. [Google Scholar] [CrossRef]
- Fetterer, R.H.; Augustine, P.C.; Allen, P.C.; Barfield, R.C. The effect of dietary betaine on intestinal and plasma levels of betaine in uninfected and coccidia-infected broiler chicks. Parasitol. Res. 2003, 90, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Augustine, P.C.; Danforth, H.D. Influence of betaine and salinomycin on intestinal absorption of methionine and glucose and on the ultrastructure of intestinal cells and parasite developmental stages in chicks infected with Eimeria acervulina. Avian Dis. 1999, 43, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Ferket, P.R. Flushing syndrome in commercial turkeys during the grow-out stage. In Proceedings of the Smithkline Beecham Pacesetter Conference, National Turkey Federation Annual Meeting, Orlando, FL, USA, 10 January 1995. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents—Final report of the American institute of nutrition ad hoc writing committee on the reformulation of the AIN-76a rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Klasing, K.C.; Adler, K.L.; Remus, J.C.; Calvert, C.C. Dietary betaine increases intraepithelial lymphocytes in the duodenum of coccidia-infected chicks and increases functional properties of phagocytes. J. Nutr. 2002, 132, 2274–2282. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 11–57. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.C.; Clayton, M.K. A similarity measure based on species proportions. Commun. Stat.-Theory Methods 2005, 34, 2123–2131. [Google Scholar] [CrossRef]
- Zeidel, M.L. Salt and water: Not so simple. J. Clin. Investig. 2017, 127, 1625–1626. [Google Scholar] [CrossRef] [PubMed]
- Deloof, S.; De Seze, C.; Montel, V.; Chatelain, A. Atrial natriuretic peptide and aldosterone secretions, and atrial natriuretic peptide-binding sites in kidneys and adrenal glands of pregnant and fetal rats in late gestation in response to a high-salt diet. Eur. J. Endocrinol. 2000, 142, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mähler, A.; Balogh, A.; Markó, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Leandro, S.M.; Furukawa, L.N.S.; Shimizu, M.H.M.; Casarini, D.E.; Seguro, A.C.; Patriarca, G.; Coelho, M.S.; Dolnikoff, M.S.; Heimann, J.C. Low birth weight in response to salt restriction during pregnancy is not due to alterations in uterine-placental blood flow or the placental and peripheral renin–angiotensin system. Physiol. Behav. 2008, 95, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Kurtz, A. Influence of salt intake on atrial natriuretic peptide gene expression in rats. Pflugers Arch. Eur. J. Physiol. 1997, 433, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Bub, A.; Rechkemmer, G.; Hock, D.; Forssmann, W.G. Urodilatin secretion in salt-loaded Wistar rats. Eur. J. Med. Res. 1996, 1, 417–424. [Google Scholar] [PubMed]
- Mcknight, J.A.; Roberts, G.; Sheridan, B.; Atkinson, A.B. The effect of low and high sodium diets on plasma atrial-natriuretic-factor, the renin-aldosterone system and blood-pressure in subjects with essential-hypertension. Clin. Endocrinol. 1994, 40, 73–77. [Google Scholar] [CrossRef]
- Seymour, A.A.; Asaad, M.M.; Abboaoffei, B.; Smith, P.L.; Rogers, W.L. Sodium loads enhance the natriuretic responses to atrial-natriuretic-peptide and neutral endopeptidase inhibitors in conscious cynomolgus monkeys. Clin. Exp. Pharmacol. Physiol. 1994, 21, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Kitada, K.; Daub, S.; Zhang, Y.; Klein, J.D.; Nakano, D.; Pedchenko, T.; Lantier, L.; LaRocque, L.M.; Marton, A.; Neubert, P.; et al. High salt intake reprioritizes osmolyte and energy metabolism for body fluid conservation. J. Clin. Investig. 2017, 127, 1944–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knepper, M.A.; Kwon, T.H.; Nielsen, S. Molecular Physiology of Water Balance. N. Engl. J. Med. 2015, 372, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Kettunen, H.; Peuranen, S.; Tiihonen, K. Betaine aids in the osmoregulation of duodenal epithelium of broiler chicks, and affects the movement of water across the small intestinal epithelium in vitro. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 129, 595–603. [Google Scholar] [CrossRef]
- Pollard, A.; Wyn Jones, R. Enzyme activities in concentrated solutions of glycinebetaine and other solutes. Planta 1979, 144, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xia, X.; Zhang, J.; Guo, Y.; Zhang, W. An Overlooked Effect of Glycine Betaine on Fermentation: Prevents Caramelization and Increases the L-Lysine Production. J. Microbiol. Biotechnol. 2014, 24, 1368–1376. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wu, Z.; Xian, M.; Liu, H.; Cheng, T.; Cao, Y. Not only osmoprotectant: Betaine increased lactate dehydrogenase activity and L-lactate production in lactobacilli. Bioresour. Technol. 2013, 148, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Chen, J.; Wu, J. Optimization of pullulanase production in Escherichia coli by regulation of process conditions and supplement with natural osmolytes. Bioresour. Technol. 2013, 146, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Kultz, D.; Chakravarty, D. Hyperosmolality in the form of elevated NaCl but not urea causes DNA damage in murine kidney cells. Proc. Natl. Acad. Sci. USA 2001, 98, 1999–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auton, M.; Rösgen, J.; Sinev, M.; Holthauzen, L.M.F.; Bolen, D.W. Osmolyte effects on protein stability and solubility: A balancing act between backbone and side-chains. Biophys. Chem. 2011, 159, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolen, D.W.; Baskakov, I.V. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol. 2001, 310, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Bruzdziak, P.; Panuszko, A.; Stangret, J. Influence of Osmolytes on Protein and Water Structure: A Step to Understanding the Mechanism of Protein Stabilization. J. Phys. Chem. B 2013, 117, 11502–11508. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, H.; Tiihonen, K.; Peuranen, S.; Saarinen, M.T.; Remus, J.C. Dietary betaine accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in healthy and coccidia-infected broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 759–769. [Google Scholar] [CrossRef]
- Hooper, L.V. Bacterial contributions to mammalian gut development. Trends Microbiol. 2004, 12, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Ferrer, M. Functional Redundancy-Induced Stability of Gut Microbiota Subjected to Disturbance. Trends Microbiol. 2016, 24, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Downes, J.; Dewhirst, F.E.; Tanner, A.C.R.; Wade, W.G. Description of Alloprevotella rava gen. nov., sp. nov., isolated from the human oral cavity, and reclassification of Prevotella tannerae Moore et al. 1994 as Alloprevotella tannerae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, J.; Fuentes, S.; Grievink, W.; van Niftrik, L.; Tindall, B.J.; Timmerman, H.M.; Rijkers, G.T.; Smidt, H. Characterization of Romboutsia ilealis gen. nov., sp. nov., isolated from the gastro-intestinal tract of a rat, and proposal for the reclassification of five closely related members of the genus Clostridium into the genera Romboutsia gen. nov., Intestinibacter gen. nov., Terrisporobacter gen. nov. and Asaccharospora gen. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1600–1616. [Google Scholar] [PubMed]
- Gerritsen, J.; Hornung, B.; Renckens, B.; van Hijum, S.A.F.T.; Martins Dos Santos, V.A.P.; Rijkers, G.T.; Schaap, P.J.; de Vos, W.M.; Smidt, H. Genomic and functional analysis of Romboutsia ilealis CRIBT reveals adaptation to the small intestine. PeerJ 2017, 5, e3698. [Google Scholar] [CrossRef] [PubMed]
- Bielinska, K.; Radkowski, M.; Grochowska, M.; Perlejewski, K.; Huc, T.; Jaworska, K.; Motooka, D.; Nakamura, S.; Ufnal, M. High salt intake increases plasma trimethylamine N-oxide (TMAO) concentration and produces gut dysbiosis in rats. Nutrition 2018, 54, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.J.; Ohye, D.F.; Christian, J.H. Tolerance of bacteria to high concentrations of NaCl and glycerol in the growth medium. Appl. Microbiol. 1971, 21, 363–364. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | A | B | C | D | SEM | p |
|---|---|---|---|---|---|---|
| Initial weight (g) | 128.4 | 128.1 | 127.9 | 127.8 | 0.90 | 0.9956 |
| Final weight (g) | 320.6 | 309.0 | 315.8 | 317.9 | 2.86 | 0.5424 |
| ADG (g/day) | 6.87 | 6.46 | 6.71 | 6.79 | 0.10 | 0.5562 |
| ADFI (g/day) | 24.77 | 26.32 | 26.74 | 26.07 | 0.46 | 0.5675 |
| Water intake (mL/day) | 32.74 b | 57.37 a | 58.99 a | 58.00 a | 4.16 | 0.0002 |
| Liver index (%) | 4.07 | 4.28 | 4.42 | 4.24 | 0.06 | 0.2475 |
| Kidney index (%) | 0.79 b | 0.91 a | 0.90 a | 0.88 a | 0.01 | 0.0160 |
| Items | A | B | C | D | SEM | p |
|---|---|---|---|---|---|---|
| Plasma | 336 | 329 | 338 | 333 | 1.2 | 0.8451 |
| Liver | 570 | 579 | 563 | 555 | 6.3 | 0.6042 |
| Kidney | 498 | 500 | 466 | 489 | 6.2 | 0.2071 |
| Digesta | ||||||
| Duodenum | 612 | 650 | 570 | 605 | 16.6 | 0.3940 |
| Jejunum | 490 | 514 | 490 | 476 | 13.0 | 0.7976 |
| Ileum | 413 | 361 | 380 | 374 | 10.6 | 0.3752 |
| Mucosa | ||||||
| Duodenum | 620 | 585 | 680 | 633 | 18.8 | 0.3823 |
| Jejunum | 520 | 548 | 560 | 505 | 15.4 | 0.6194 |
| Ileum | 475 | 493 | 518 | 528 | 24.4 | 0.8943 |
| Items | A | B | C | D | SEM | p |
|---|---|---|---|---|---|---|
| Plasma | ||||||
| Na | 142.2 | 143.1 | 141.8 | 141.3 | 0.32 | 0.2730 |
| K | 5.75 | 5.82 | 5.84 | 5.95 | 0.09 | 0.8801 |
| Cl | 100.3 a | 99.8 ab | 98.5 bc | 97.9 c | 0.33 | 0.0174 |
| Liver | ||||||
| Na | 98.6 | 99.0 | 101.9 | 101.4 | 1.15 | 0.6940 |
| K | 83.9 | 83.6 | 84.2 | 83.8 | 0.62 | 0.9901 |
| Cl | 118.6 b | 130.6 a | 134.3 a | 134.4 a | 1.74 | 0.0008 |
| Kidney | ||||||
| Na | 119.6 | 124.6 | 126.3 | 121.3 | 1.38 | 0.3140 |
| K | 67.1 | 67.1 | 67.6 | 68.7 | 1.21 | 0.9668 |
| Cl | 97.9 b | 102.5 ab | 105.8 a | 104.4 a | 0.96 | 0.0150 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Li, S.; Fang, S.; Yang, X.; Feng, J. Betaine Improves Intestinal Functions by Enhancing Digestive Enzymes, Ameliorating Intestinal Morphology, and Enriching Intestinal Microbiota in High-salt stressed Rats. Nutrients 2018, 10, 907. https://doi.org/10.3390/nu10070907
Wang H, Li S, Fang S, Yang X, Feng J. Betaine Improves Intestinal Functions by Enhancing Digestive Enzymes, Ameliorating Intestinal Morphology, and Enriching Intestinal Microbiota in High-salt stressed Rats. Nutrients. 2018; 10(7):907. https://doi.org/10.3390/nu10070907
Chicago/Turabian StyleWang, Haichao, Sisi Li, Shenglin Fang, Xiaojing Yang, and Jie Feng. 2018. "Betaine Improves Intestinal Functions by Enhancing Digestive Enzymes, Ameliorating Intestinal Morphology, and Enriching Intestinal Microbiota in High-salt stressed Rats" Nutrients 10, no. 7: 907. https://doi.org/10.3390/nu10070907