Folinic Acid Increases Protein Arginine Methylation in Human Endothelial Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Determination of Total Hcy (tHcy) in Medium and of Intracellular AdoMet and AdoHcy
2.4. Evaluation of Protein Arginine Methylation Status in Cells
2.5. Statistical Analysis
3. Results
3.1. High Methionine Availability Offsets the Effect of Folinic Acid on Homocysteine Export from Endothelial Cells
3.2. Folinic Acid Decreases Cellular Methylation Capacity in Endothelial Cells under Low Methionine Availability
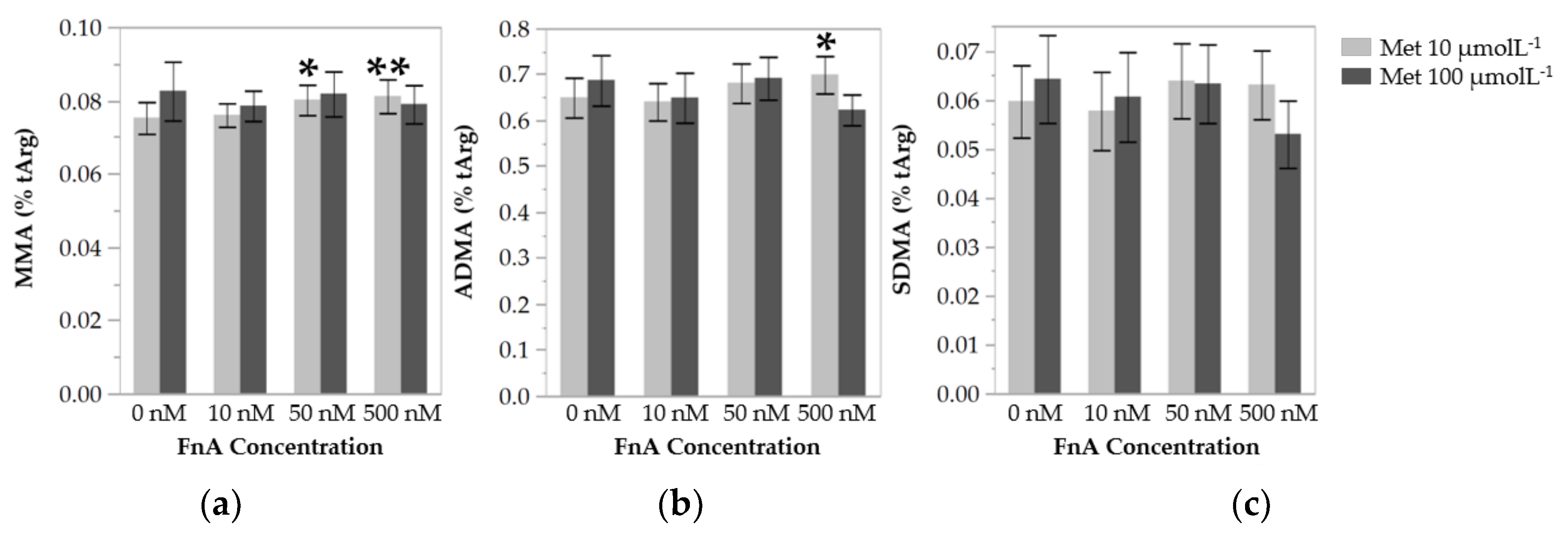
3.3. Folinic Acid Lowers Protein Arginine Methylation in Endothelial Cells under Low Methionine Availability
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Blom, H.J.; Shaw, G.M.; den Heijer, M.; Finnell, R.H. Neural tube defects and folate: Case far from closed. Nat. Rev. Neurosci. 2006, 7, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Gonda, T.A.; Kim, Y.-I.; Salas, M.C.; Gamble, M.V.; Shibata, W.; Muthupalani, S.; Sohn, K.-J.; Abrams, J.A.; Fox, J.G.; Wang, T.C.; et al. Folic acid increases global DNA methylation and reduces inflammation to prevent Helicobacter-associated gastric cancer in mice. Gastroenterology 2012, 142, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.-Y.; Chen, H.-M.; Chen, Y.-X.; Wang, Y.-C.; Wang, Z.-H.; Tang, J.-T.; Ge, Z.-Z.; Chen, X.-Y.; Sheng, J.-Q.; Fang, D.-C.; et al. Folic acid prevents the initial occurrence of sporadic colorectal adenoma in Chinese older than 50 years of age: A randomized clinical trial. Cancer Prev. Res. (Phila). 2013, 6, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Rivera, I.; Blom, H.J.; Jakobs, C.; Tavares de Almeida, I. Homocysteine metabolism, hyperhomocysteinaemia and vascular disease: An overview. J. Inherit. Metab. Dis. 2006, 29, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-H.; Tang, J.-Y.; Wu, M.-J.; Lu, J.; Wei, X.; Qin, Y.-Y.; Wang, C.; Xu, J.-F.; He, J. Effect of folic acid supplementation on cardiovascular outcomes: A systematic review and meta-analysis. PLoS ONE 2011, 6, e25142. [Google Scholar] [CrossRef] [PubMed]
- Jardine, M.J.; Kang, A.; Zoungas, S.; Navaneethan, S.D.; Ninomiya, T.; Nigwekar, S.U.; Gallagher, M.P.; Cass, A.; Strippoli, G.; Perkovic, V. The effect of folic acid based homocysteine lowering on cardiovascular events in people with kidney disease: Systematic review and meta-analysis. BMJ 2012, 344, e3533. [Google Scholar] [CrossRef] [PubMed]
- Barroso, M.; Handy, D.E.; Castro, R. The Link between Hyperhomocysteinemia and Hypomethylation. J. Inborn Errors Metab. Screen. 2017, 5. [Google Scholar] [CrossRef]
- Finkelstein, J.D.; Martin, J.J. Methionine metabolism in mammals. Adaptation to methionine excess. J. Biol. Chem. 1986, 261, 1582–1587. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Rivera, I.; Struys, E.A.; Jansen, E.E.; Ravasco, P.; Camilo, M.E.; Blom, H.J.; Jakobs, C.; Tavares de Almeida, I. Increased homocysteine and S-adenosylhomocysteine concentrations and DNA hypomethylation in vascular disease. Clin. Chem. 2003, 49, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Esse, R.; Florindo, C.; Imbard, A.; Rocha, M.S.; de Vriese, A.S.; Smulders, Y.M.; Teerlink, T.; Tavares de Almeida, I.; Castro, R.; Blom, H.J. Global protein and histone arginine methylation are affected in a tissue-specific manner in a rat model of diet-induced hyperhomocysteinemia. Biochim. Biophys. Acta 2013, 1832, 1708–1714. [Google Scholar] [CrossRef] [PubMed]
- Friso, S.; Choi, S.-W.; Girelli, D.; Mason, J.B.; Dolnikowski, G.G.; Bagley, P.J.; Olivieri, O.; Jacques, P.F.; Rosenberg, I.H.; Corrocher, R.; et al. A common mutation in the 5,10-methylenetetrahydrofolate reductase gene affects genomic DNA methylation through an interaction with folate status. Proc. Natl. Acad. Sci. USA 2002, 99, 5606–5611. [Google Scholar] [CrossRef] [PubMed]
- Ingrosso, D.; Cimmino, A.; Perna, A.F.; Masella, L.; De Santo, N.G.; De Bonis, M.L.; Vacca, M.; D’Esposito, M.; D’Urso, M.; Galletti, P.; et al. Folate treatment and unbalanced methylation and changes of allelic expression induced by hyperhomocysteinaemia in patients with uraemia. Lancet 2003, 361, 1693–1699. [Google Scholar] [CrossRef]
- James, S.J.; Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Caudill, M.A. Elevation in S-adenosylhomocysteine and DNA hypomethylation: Potential epigenetic mechanism for homocysteine-related pathology. J. Nutr. 2002, 132, 2361S–2366S. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Hine, R.J.; James, S.J. Increase in plasma homocysteine associated with parallel increases in plasma S-adenosylhomocysteine and lymphocyte DNA hypomethylation. J. Biol. Chem. 2000, 275, 29318–29323. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Karaplis, A.C.; Ackerman, S.L.; Pogribny, I.P.; Melnyk, S.; Lussier-Cacan, S.; Chen, M.F.; Pai, A.; John, S.W.; Smith, R.S.; et al. Mice deficient in methylenetetrahydrofolate reductase exhibit hyperhomocysteinemia and decreased methylation capacity, with neuropathology and aortic lipid deposition. Hum. Mol. Genet. 2001, 10, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Choumenkovitch, S.F.; Selhub, J.; Bagley, P.J.; Maeda, N.; Nadeau, M.R.; Smith, D.E.; Choi, S.-W. In the cystathionine beta-synthase knockout mouse, elevations in total plasma homocysteine increase tissue S-adenosylhomocysteine, but responses of S-adenosylmethionine and DNA methylation are tissue specific. J. Nutr. 2002, 132, 2157–2160. [Google Scholar] [CrossRef] [PubMed]
- Esse, R.; Leandro, P.; Tavares de Almeida, I.; Blom, H.J.; Castro, R. Deciphering Protein Arginine Methylation in Mammals. In Methylation—From DNA, RNA and Histones to Diseases and Treatment; Dricu, A., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Bedford, M.T.; Clarke, S.G. Protein arginine methylation in mammals: Who, what, and why. Mol. Cell 2009, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Esse, R.; Rocha, M.S.; Barroso, M.; Florindo, C.; Teerlink, T.; Kok, R.M.; Smulders, Y.M.; Rivera, I.; Leandro, P.; Koolwijk, P.; et al. Protein arginine methylation is more prone to inhibition by S-adenosylhomocysteine than DNA methylation in vascular endothelial cells. PLoS ONE 2013, 8, e55483. [Google Scholar] [CrossRef]
- Esse, R.; Imbard, A.; Florindo, C.; Gupta, S.; Quinlivan, E.P.; Davids, M.; Teerlink, T.; Tavares de Almeida, I.; Kruger, W.D.; Blom, H.J.; et al. Protein arginine hypomethylation in a mouse model of cystathionine β-synthase deficiency. FASEB J. 2014, 2686–2695. [Google Scholar] [CrossRef] [PubMed]
- Lonn, E.; Yusuf, S.; Arnold, M.J.; Sheridan, P.; Pogue, J.; Micks, M.; McQueen, M.J.; Probstfield, J.; Fodor, G.; Held, C.; et al. Homocysteine lowering with folic acid and B vitamins in vascular disease. N. Engl. J. Med. 2006, 354, 1567–1577. [Google Scholar] [CrossRef] [PubMed]
- Smulders, Y.M.; Blom, H.J. The homocysteine controversy. J. Inherit. Metab. Dis. 2011, 34, 93–99. [Google Scholar] [CrossRef] [PubMed]
- McDowell, I.F.; Lang, D. Homocysteine and endothelial dysfunction: A link with cardiovascular disease. J. Nutr. 2000, 130, 369S–372S. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Rivera, I.; Martins, C.; Struys, E.A.; Jansen, E.E.; Clode, N.; Graça, L.M.; Blom, H.J.; Jakobs, C.; de Almeida, I.T. Intracellular S-adenosylhomocysteine increased levels are associated with DNA hypomethylation in HUVEC. J. Mol. Med. (Berl.) 2005, 83, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Barroso, M.; Kao, D.; Blom, H.J.; Tavares de Almeida, I.; Castro, R.; Loscalzo, J.; Handy, D.E. S-adenosylhomocysteine induces inflammation through NFkB: A possible role for EZH2 in endothelial cell activation. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Barroso, M.; Florindo, C.; Kalwa, H.; Silva, Z.; Turanov, A.A.; Carlson, B.A.; De Almeida, I.T.; Blom, H.J.; Gladyshev, V.N.; Hatfield, D.L.; et al. Inhibition of cellular methyltransferases promotes endothelial cell activation by suppressing glutathione peroxidase 1 protein expression. J. Biol. Chem. 2014, 289, 15350–15362. [Google Scholar] [CrossRef] [PubMed]
- Doshi, S.N.; McDowell, I.F.; Moat, S.J.; Lang, D.; Newcombe, R.G.; Kredan, M.B.; Lewis, M.J.; Goodfellow, J. Folate improves endothelial function in coronary artery disease: An effect mediated by reduction of intracellular superoxide? Arterioscler. Thromb. Vasc Biol. 2001, 21, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Van der Molen, E.F.; van den Heuvel, L.P.; te Poele Pothoff, M.T.; Monnens, I.A.; Eskes, T.K.; Blom, H.J. The effect of folic acid on the homocysteine metabolism in human umbilical vein endothelial cells (HUVECs). Eur. J. Clin. Investig. 1996, 26, 304–309. [Google Scholar] [CrossRef]
- Litynski, P.; Loehrer, F.; Linder, L.; Todesco, L.; Fowler, B. Effect of low doses of 5-methyltetrahydrofolate and folic acid on plasma homocysteine in healthy subjects with or without the 677CT polymorphism of methylenetetrahydrofolate reductase. Eur. J. Clin. Investig. 2002, 32, 662–668. [Google Scholar] [CrossRef]
- Holmes, W.B.; Appling, D.R. Cloning and characterization of methenyltetrahydrofolate synthetase from Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 20205–20213. [Google Scholar] [CrossRef] [PubMed]
- Selhub, J.; Miller, J.W. The pathogenesis of homocysteinemia: Interruption of the coordinate regulation by S-adenosylmethionine of the remethylation and transsulfuration of homocysteine. Am. J. Clin. Nutr. 1992, 55, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Nijhout, H.F.; Reed, M.C.; Anderson, D.F.; Mattingly, J.C.; James, S.J.; Ulrich, C.M. Long-range allosteric interactions between the folate and methionine cycles stabilize DNA methylation reaction rate. Epigenetics 2006, 1, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Smulders, Y.M.; Teerlink, T.; Struys, E.A.; De Meer, K.; Kostense, P.J.; Jakobs, C.; Dekker, J.M.; Nijpels, G.; Heine, R.J.; et al. S-adenosylhomocysteine and the ratio of S-adenosylmethionine to S-adenosylhomocysteine are not related to folate, cobalamin and vitamin B6 concentrations. Eur. J. Clin. Investig. 2003, 33, 17–25. [Google Scholar] [CrossRef]
- Green, T.J.; Skeaff, C.M.; McMahon, J.A.; Venn, B.J.; Williams, S.M.; Devlin, A.M.; Innis, S.M. Homocysteine-lowering vitamins do not lower plasma S-adenosylhomocysteine in older people with elevated homocysteine concentrations. Br. J. Nutr. 2010, 103, 1629–1634. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esse, R.; Teerlink, T.; Koolwijk, P.; Tavares de Almeida, I.; Blom, H.J.; Castro, R. Folinic Acid Increases Protein Arginine Methylation in Human Endothelial Cells. Nutrients 2018, 10, 404. https://doi.org/10.3390/nu10040404
Esse R, Teerlink T, Koolwijk P, Tavares de Almeida I, Blom HJ, Castro R. Folinic Acid Increases Protein Arginine Methylation in Human Endothelial Cells. Nutrients. 2018; 10(4):404. https://doi.org/10.3390/nu10040404
Chicago/Turabian StyleEsse, Ruben, Tom Teerlink, Pieter Koolwijk, Isabel Tavares de Almeida, Henk J. Blom, and Rita Castro. 2018. "Folinic Acid Increases Protein Arginine Methylation in Human Endothelial Cells" Nutrients 10, no. 4: 404. https://doi.org/10.3390/nu10040404