Three Constituents of Moringa oleifera Seeds Regulate Expression of Th17-Relevant Cytokines and Ameliorate TPA-Induced Psoriasis-Like Skin Lesions in Mice

 and
and 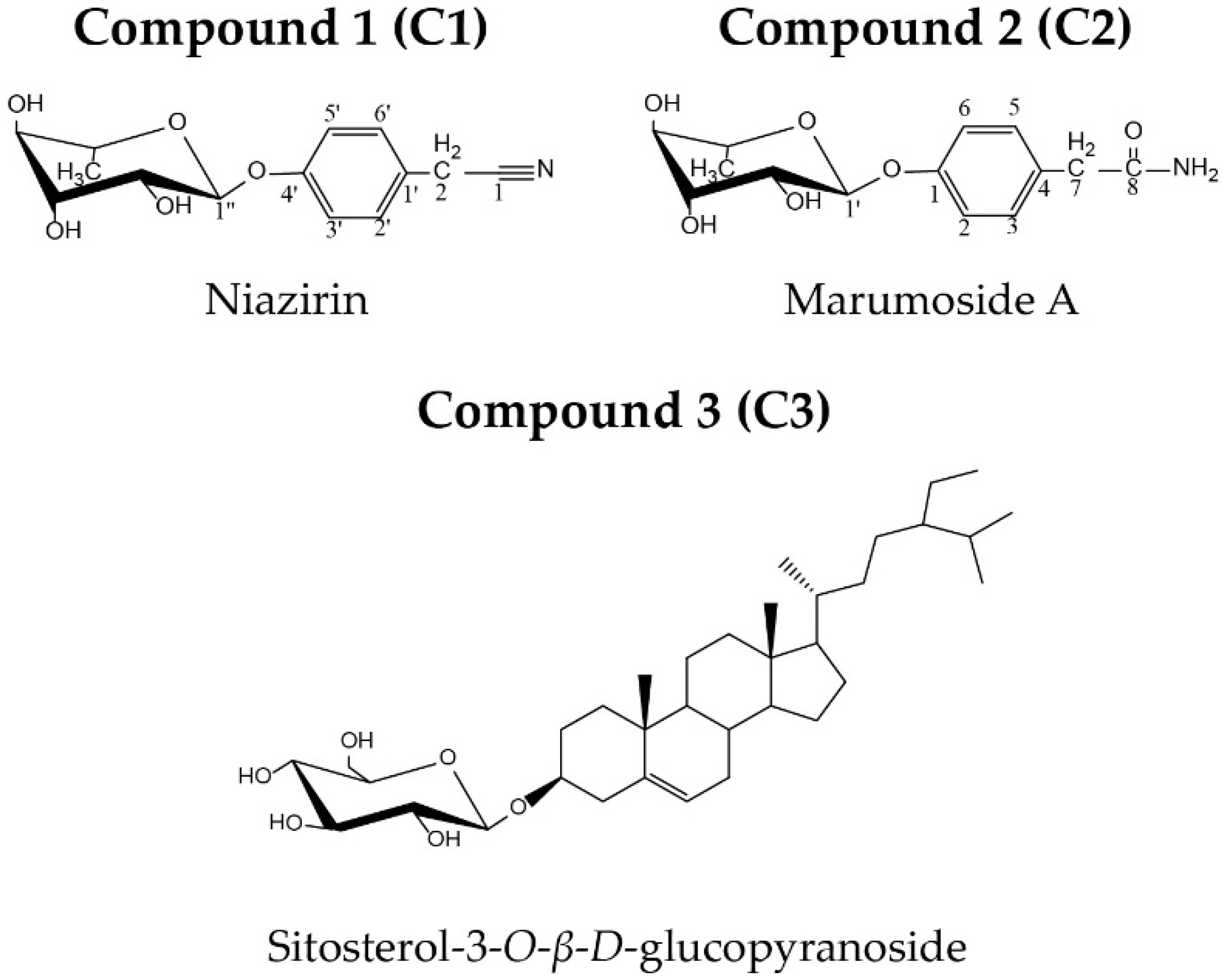{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. C.haracterization of Isolated Compounds
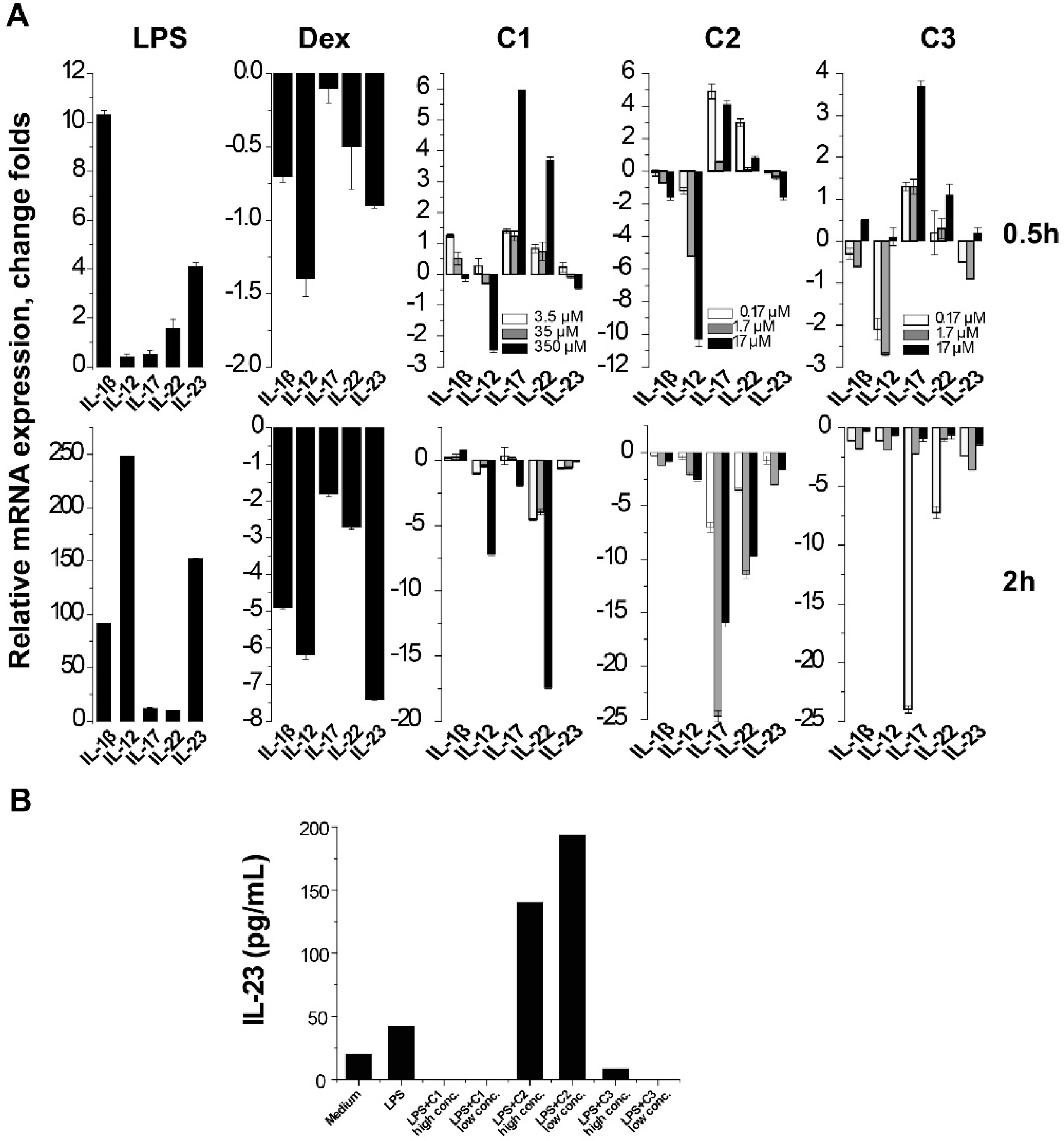
2.2. Suppression of Th17-Relevant Cytokines in LPS-Stimulated THP-1 Cells
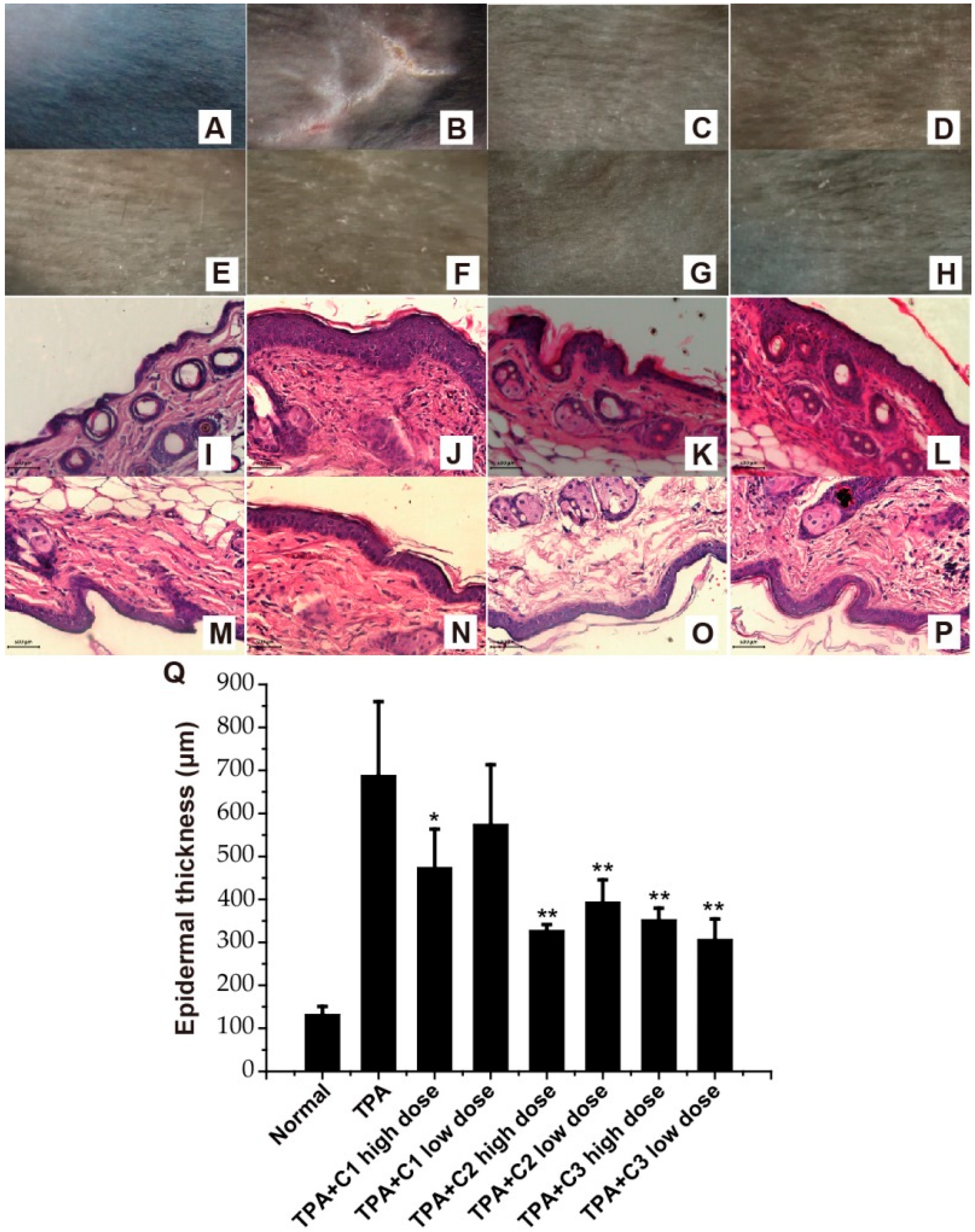
2.3. Amelioration of TPA-Induced Psoriasis—Like Skin Lesions
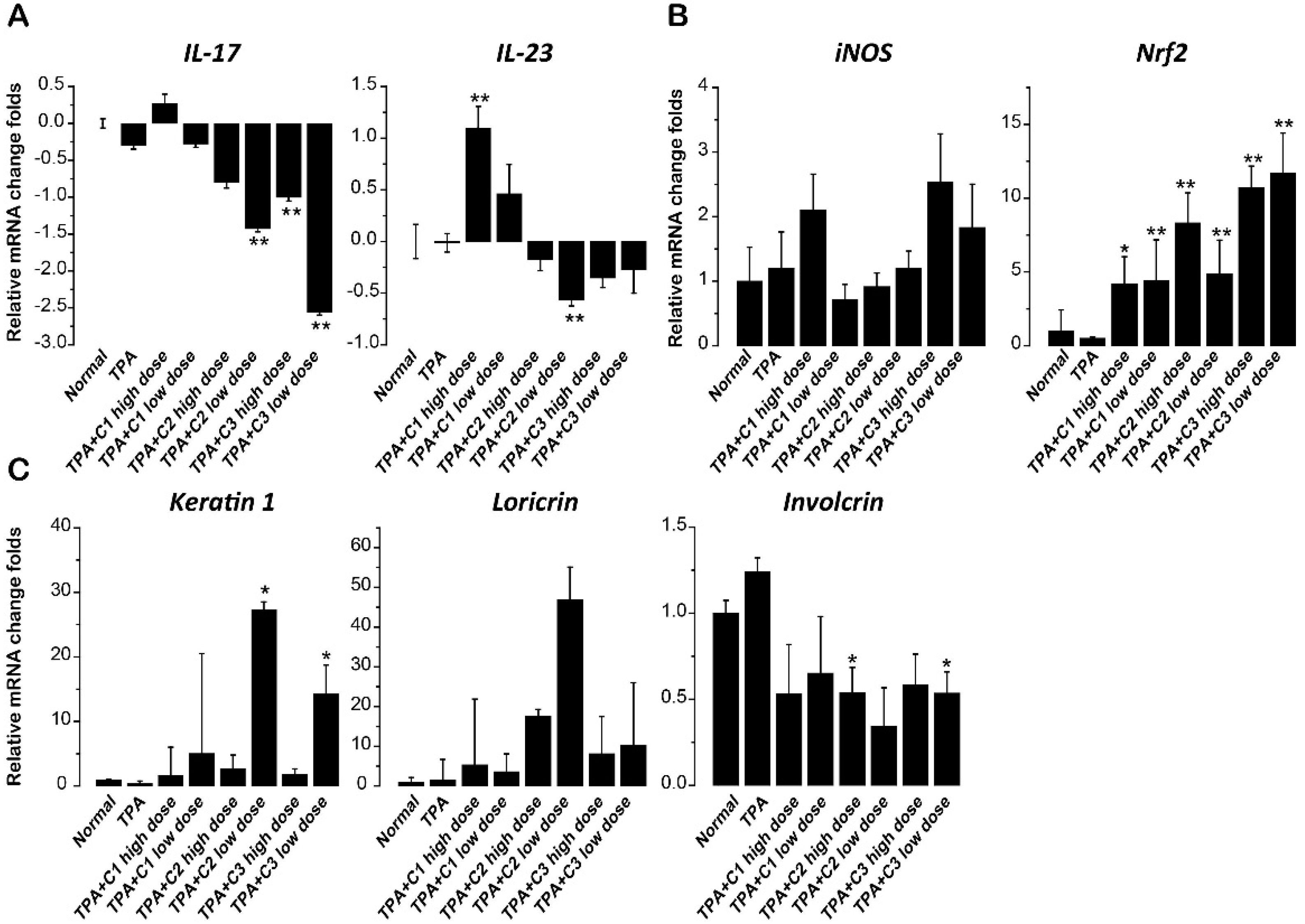
2.4. Effect of Compounds on Th17-Relevant Cytokines in Skin
2.5. Effect of Compounds on Inducible NO Synthase (iNOS) and Nuclear Factor Erythroid-2-Related Factor 2 (Nrf2)
2.6. Effect of Compounds on Differentiation Markers of Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Isolation
4.3. General Experimental Procedures for Structural Determination
4.4. THP-1 Cell Culture and Compound Treatment
4.5. RNA Isolation, Reverse Transcription, and Quantitative PCR Analysis
4.6. Enzyme-Linked Immunosorbent Assay (ELISA) Detection of IL-23 in Culture Supernatant
4.7. Preparation of Ointment
4.8. Treatment of TPA-Induced Psoriasis-Like Skin Inflammation in C57BL/6J Mice
4.9. Histology
4.10. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fahey, J.W. Moringa oleifera: A review of the medical evidence for its nutritional, therapeutic, and prophylactic properties. Part 1. Trees Life J. 2005, 1, 1–15. [Google Scholar]
- Faizi, S.; Siddiqui, B.S.; Saleem, R.; Siddiqui, S.; Aftab, K.; Gilani, A.H. Fully acetylated carbamate and hypotensive thiocarbamate glycosides from Moringa oleifera. Phytochemistry 1995, 38, 957–963. [Google Scholar] [CrossRef]
- Villarruel-Lopez, A.; Lopez-de la Mora, D.A.; Vazquez-Paulino, O.D.; Puebla-Mora, A.G.; Torres-Vitela, M.R.; Guerrero-Quiroz, L.A.; Nuno, K. Effect of Moringa oleifera consumption on diabetic rats. BMC Complement. Altern. Med. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Annongu, A.; Karim, O.R.; Toye, A.A.; Sola-Ojo, F.E.; Kayode, R.M.O.; Badmos, A.H.A.; Alli, O.I.; Adeyemi, K.D. Geo-assessment of chemical composition and nutritional evaluation of Moringa oleifera seeds in nutrition of broilers. J. Agric. Sci. 2014, 6, 119. [Google Scholar] [CrossRef]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Cultivation, genetic, ethnopharmacology, phytochemistry and pharmacology of Moringa oleifera leaves: An overview. Int. J. Mol. Sci. 2015, 16, 12791–12835. [Google Scholar] [CrossRef]
- Vergara Jimenez, M.; Almatrafi, M.M.; Fernandez, M.L. Bioactive components in Moringa Oleifera leaves protect against chronic disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef]
- Toma, A.; Deyno, S. Phytochemistry and pharmacological activities of Moringa oleifera. Int. J. Pharmacogn. 2014, 1, 222–231. [Google Scholar]
- Cacares, C.A.; Saravia, A.; Rizzo, S.; Zabala, L.; Leon, E.D.; Nave, F. Pharmacologic properties of Moringa oleifera. 2: Screening for antispasmodic, antiinflammatory and diuretic activity. J. Ethnopharmacol. 1992, 36, 233–237. [Google Scholar] [CrossRef]
- Gupta, R.; Mathur, M.; Bajaj, V.K.; Katariya, P.; Yadav, S.; Kamal, R.; Gupta, R.S. Evaluation of antidiabetic and antioxidant activity of Moringa oleifera in experimental diabetes. J. Diabetes 2012, 4, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Arulselvan, P.; Tan, W.S.; Gothai, S.; Muniandy, K.; Fakurazi, S.; Esa, N.M.; Alarfaj, A.A.; Kumar, S.S. Anti-inflammatory potential of ethyl acetate fraction of Moringa oleifera in downregulating the NF-κB signaling pathway in lipopolysaccharide-stimulated macrophages. Molecules 2016, 21, 1452. [Google Scholar] [CrossRef]
- Kooltheat, N.; Sranujit, R.P.; Chumark, P.; Potup, P.; Laytragoon-Lewin, N.; Usuwanthim, K. An ethyl acetate fraction of Moringa oleifera Lam. inhibits human macrophage cytokine production induced by cigarette smoke. Nutrients 2014, 6, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Savelkoul, H.F.J.; Wichers, H.J. Transcription profiles of LPS-stimulated THP-1 monocytes and macrophages: A tool to study inflammation modulating effects of food-derived compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Di Meglio, P.; Nestle, F.O. The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J. Investig. Dermatol. 2009, 129, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Faizi, S.; Siddiqui, B.S.; Saleem, R.; Siddiqui, S.; Aftab, K.; Gilani, A.U.H. Isolation and structure elucidation of new nitrile and mustard oil glycosides from Moringa-Oleifera and their effect on blood-pressure. J. Nat. Prod. 1994, 57, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Vudhgirl, S.; Prasad, R.B.N.; Kota, A. Synthesis and biological evaluation of marumoside A isolated from Moringa Oleifera and its lipid derivatives. Int. J. Pharm. Sci. Res. 2016, 2, 607–617. [Google Scholar]
- Khatun, M.; Billah, M.; Quader, A. Sterols and sterol glucoside from Phyllanthus species. Dhaka Uni. J. Sci. 2012, 60, 5–10. [Google Scholar] [CrossRef]
- Amigo, M.; Paya, M.; De Rosa, S.; Terencio, M.C. Antipsoriatic effects of avarol-3′-thiosalicylate are mediated by inhibition of TNF-alpha generation and NF-κB activation in mouse skin. Br. J. Pharmacol. 2007, 152, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Lowes, M.A.; Bowcock, A.M.; Krueger, J.G. Pathogenesis and therapy of psoriasis. Nature 2007, 445, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, H.; Takahashi, H.; Honma, M.; Ishida-Yamamoto, A. Unique keratinization process in psoriasis: Late differentiation markers are abolished because of the premature cell death. J. Dermatol. 2004, 31, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, R.A.; Fischer, S.M.; Locniskar, M.F. Effect of 12-O-tetradecanoylphorbol-13-acetate on inhibition of expression of keratin 1 mRNA in mouse keratinocytes mimicked by 12(S)-hydroxyeicosatetraenoic acid. Mol. Carcinog. 1997, 19, 157–164. [Google Scholar] [CrossRef]
- Nair, R.P.; Duffin, K.C.; Helms, C.; Ding, J.; Stuart, P.E.; Goldgar, D.; Gudjonsson, J.E.; Li, Y.; Tejasvi, T.; Feng, B.J.; et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-κB pathways. Nat. Genet. 2009, 41, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fotiadou, C.; Lazaridou, E.; Sotiriou, E.; Gerou, S.; Kyrgidis, A.; Vakirlis, E.; Ioannides, D. IL-17A, IL-22, and IL-23 as markers of psoriasis activity: A cross-sectional, hospital-based study. J. Cutan. Med. Surg. 2015, 19, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Conrad, C. The IL-12 family member p40 chain as a master switch and novel therapeutic target in psoriasis. J. Investig. Dermatol. 2004, 123, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.; Chu, A.; Ludviksson, B.R.; Berg, E.L.; Ehrhardt, R.O. IL-12, independently of IFN-gamma, plays a crucial role in the pathogenesis of a murine psoriasis-like skin disorder. J. Immunol. 1999, 162, 7480–7491. [Google Scholar] [PubMed]
- Takahashi, T.; Koga, Y.; Kainoh, M. Anti-IL-12/IL-23p40 antibody ameliorates dermatitis and skin barrier dysfunction in mice with imiquimod-induced psoriasis-like dermatitis. Eur. J. Pharmacol. 2018, 828, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Van Belle, A.B.; Heusch, M.; Lemaire, M.M.; Hendrickx, E.; Warnier, G.; Dunussi-Joannopoulos, K.; Fouser, L.A.; Renauld, J.C.; Dumoutier, L. IL-22 is required for imiquimod-induced psoriasiform skin inflammation in mice. J. Immunol. 2012, 188, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.L.; Liang, S.; Li, J.; Napierata, L.; Brown, T.; Benoit, S.; Senices, M.; Gill, D.; Dunussi-Joannopoulos, K.; Collins, M.; et al. IL-22 is required for Th17 cell-mediated pathology in a mouse model of psoriasis-like skin inflammation. J. Clin. Investig. 2008, 118, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, R.; Figueiredo, C.P.; Passos, G.F.; Calixto, J.B. Reduced skin inflammatory response in mice lacking inducible nitric oxide synthase. Biochem. Pharmacol. 2009, 78, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Bae, J.H.; Kang, S.G.; Cho, S.W.; Chun, D.I.; Nam, S.M.; Kim, C.H.; Nam, H.S.; Lee, S.H.; Cho, M.K. Pro-oxidant status and Nrf2 levels in psoriasis vulgaris skin tissues and dimethyl fumarate-treated HaCaT cells. Arch. Pharm. Res. 2017, 40, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Bruck, J.; Holstein, J.; Glocova, I.; Seidel, U.; Geisel, J.; Kanno, T.; Kumagai, J.; Mato, N.; Sudowe, S.; Widmaier, K.; et al. Nutritional control of IL-23/Th17-mediated autoimmune disease through HO-1/STAT3 activation. Sci. Rep. 2017, 7, 44482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arul, S.; Dayalan, H.; Jegadeesan, M.; Damodharan, P. Induction of differentiation in psoriatic keratinocytes by propylthiouracil and fructose. BBA Clin. 2016, 6, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, N.M.; Muley, M.M.; Jaji, M.S.; Vijaykanth, G.; Raghul, J.; Reddy, N.K.; Vishwakarma, S.L.; Rajesh, N.B.; Mookkan, J.; Krishnan, U.M.; et al. Topical atorvastatin ameliorates 12-O-tetradecanoylphorbol-13-acetate induced skin inflammation by reducing cutaneous cytokine levels and NF-κB activation. Arch. Pharm. Res. 2015, 38, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
Sample Availability: Samples of the compounds are available from Xin-an Huang ([email protected]). |




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, N.; Tang, Q.; Wu, W.-T.; Huang, X.-A.; Xu, Q.; Rong, G.-L.; Chen, S.; Song, J.-P. Three Constituents of Moringa oleifera Seeds Regulate Expression of Th17-Relevant Cytokines and Ameliorate TPA-Induced Psoriasis-Like Skin Lesions in Mice. Molecules 2018, 23, 3256. https://doi.org/10.3390/molecules23123256
Ma N, Tang Q, Wu W-T, Huang X-A, Xu Q, Rong G-L, Chen S, Song J-P. Three Constituents of Moringa oleifera Seeds Regulate Expression of Th17-Relevant Cytokines and Ameliorate TPA-Induced Psoriasis-Like Skin Lesions in Mice. Molecules. 2018; 23(12):3256. https://doi.org/10.3390/molecules23123256
Chicago/Turabian StyleMa, Nuan, Qin Tang, Wan-Ting Wu, Xin-An Huang, Qin Xu, Guang-Li Rong, Song Chen, and Jian-Ping Song. 2018. "Three Constituents of Moringa oleifera Seeds Regulate Expression of Th17-Relevant Cytokines and Ameliorate TPA-Induced Psoriasis-Like Skin Lesions in Mice" Molecules 23, no. 12: 3256. https://doi.org/10.3390/molecules23123256