Effects of Mogrosides on High-Fat-Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
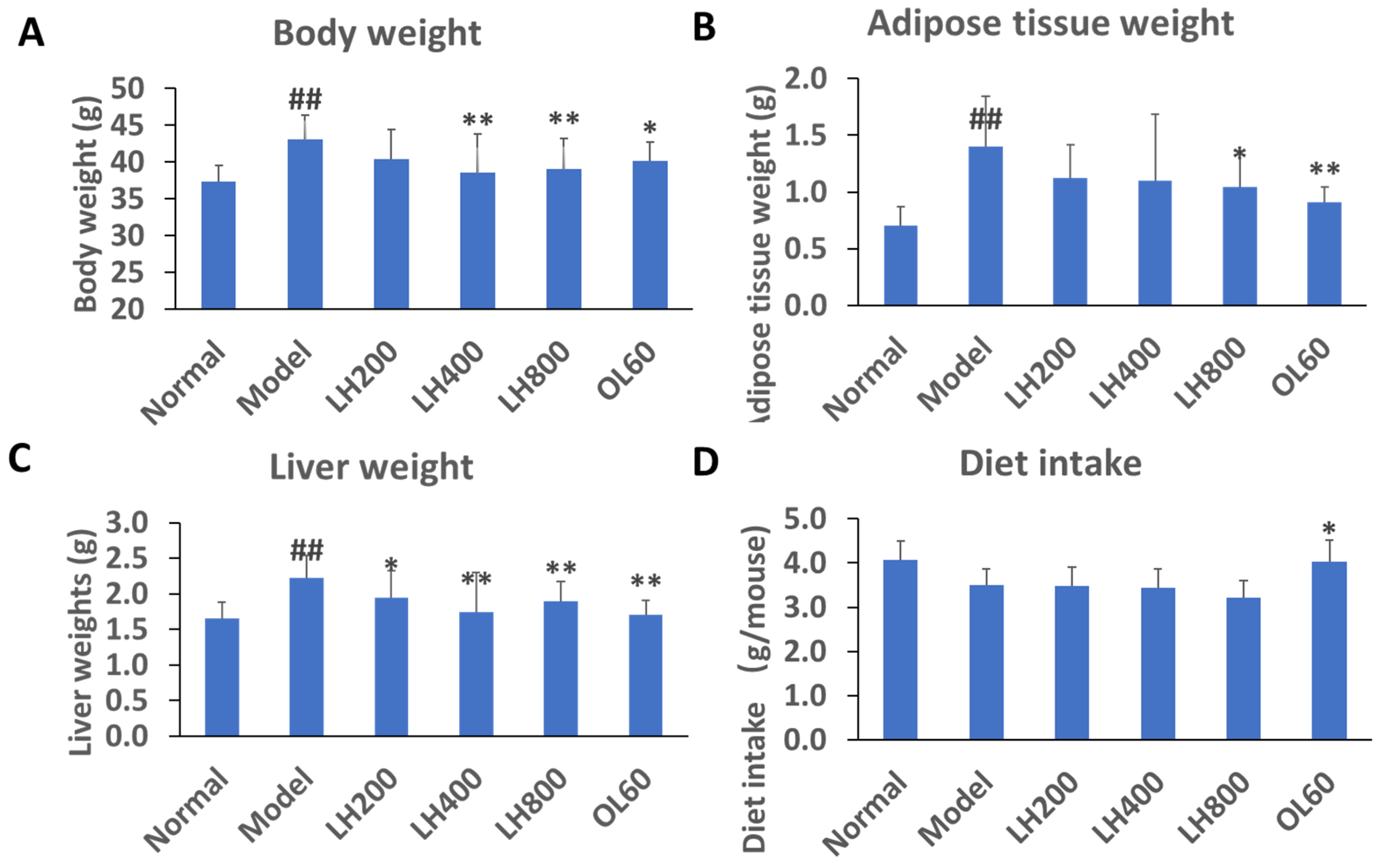
2.1. Effects of LH on Biochemical Parameters
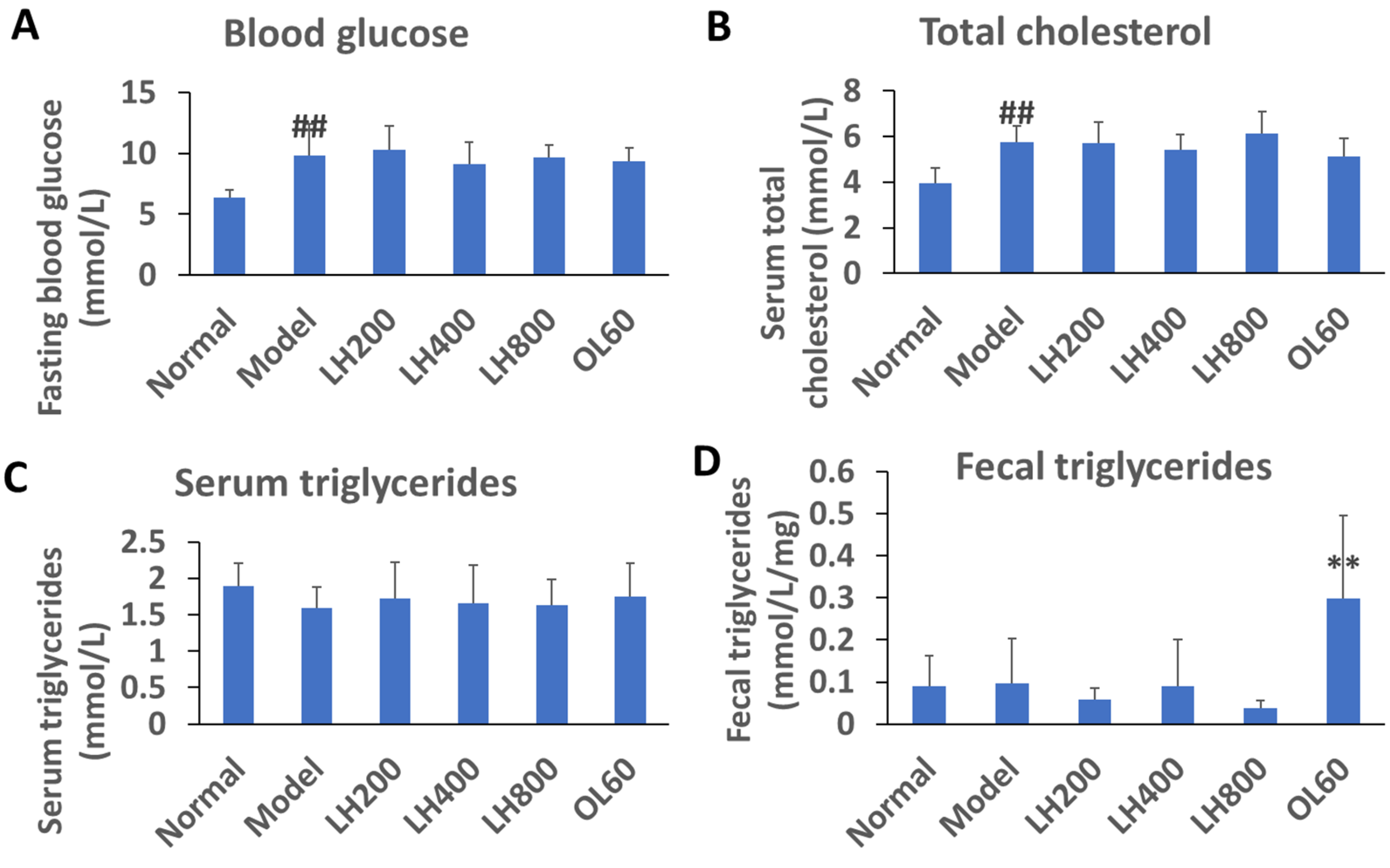
2.2. Effects of LH on Blood Glucose, Lipid, and Fecal TG Levels
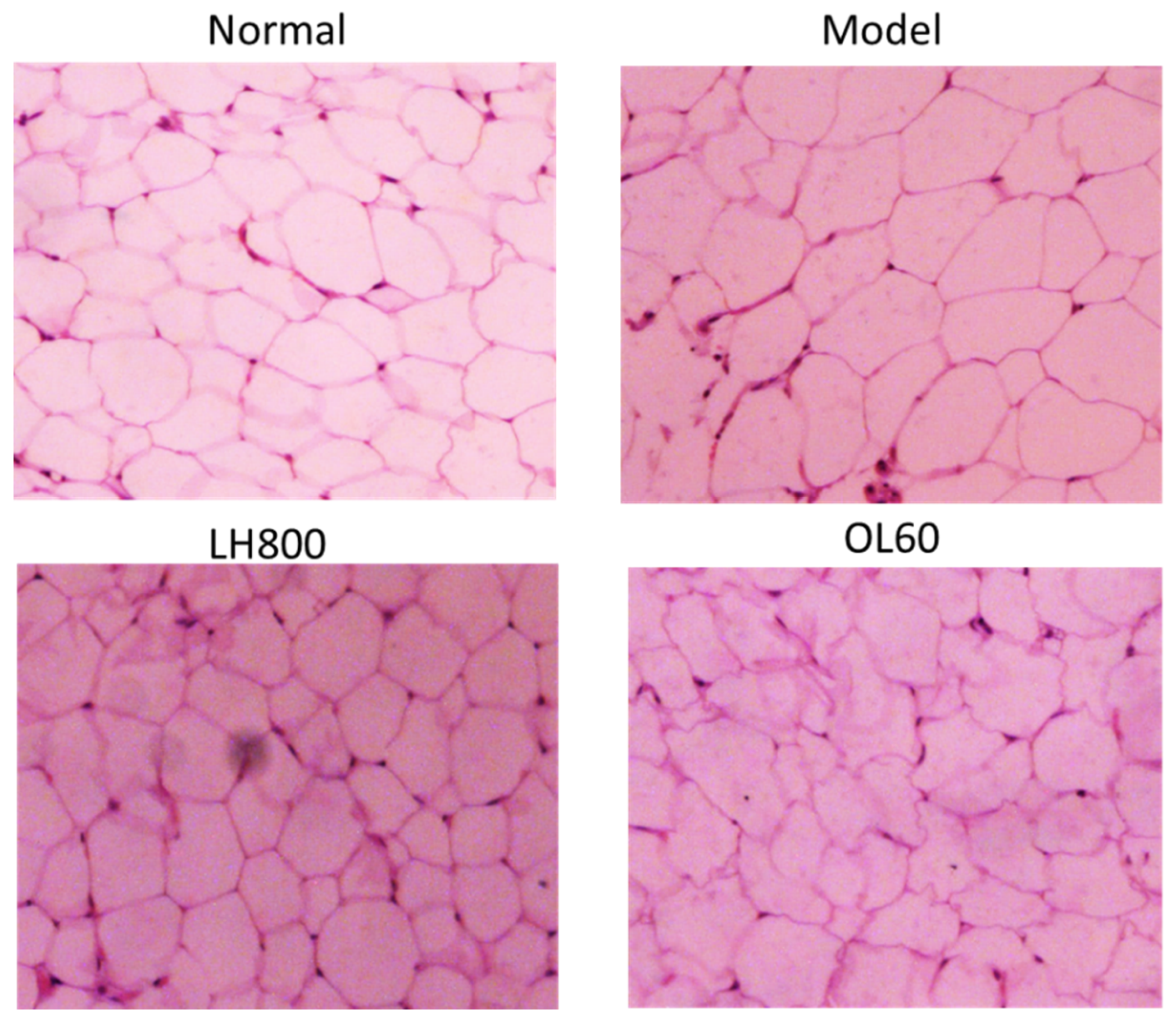
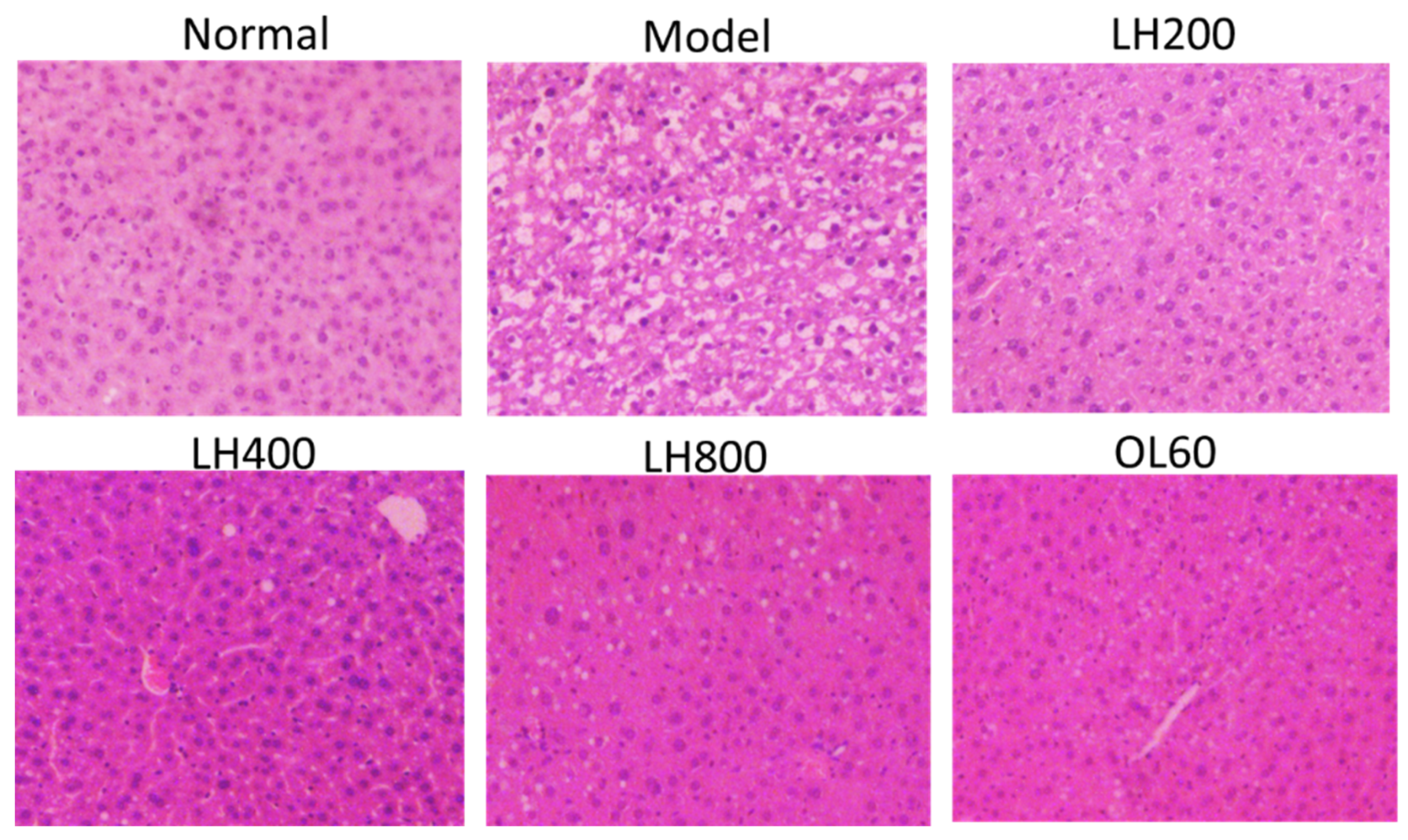
2.3. Effects of LH on Abdominal Adipose Tissue Size, Liver Morphology, and Pathology
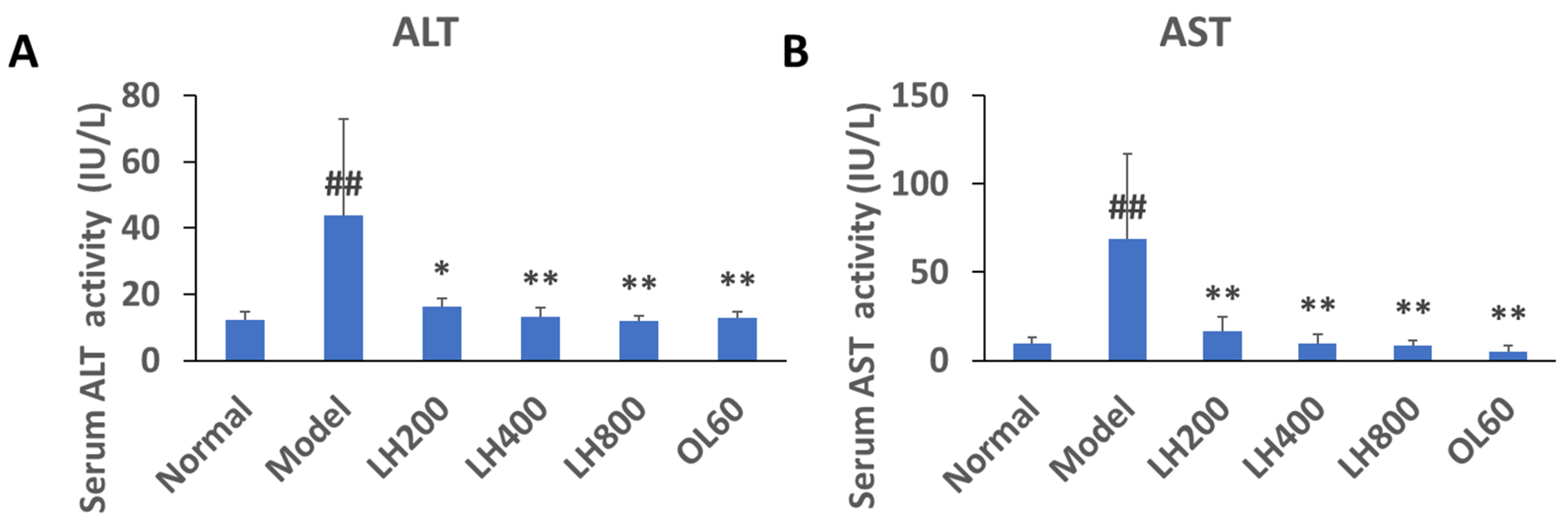
2.4. Effects of LH on Serum ALT and AST Activity
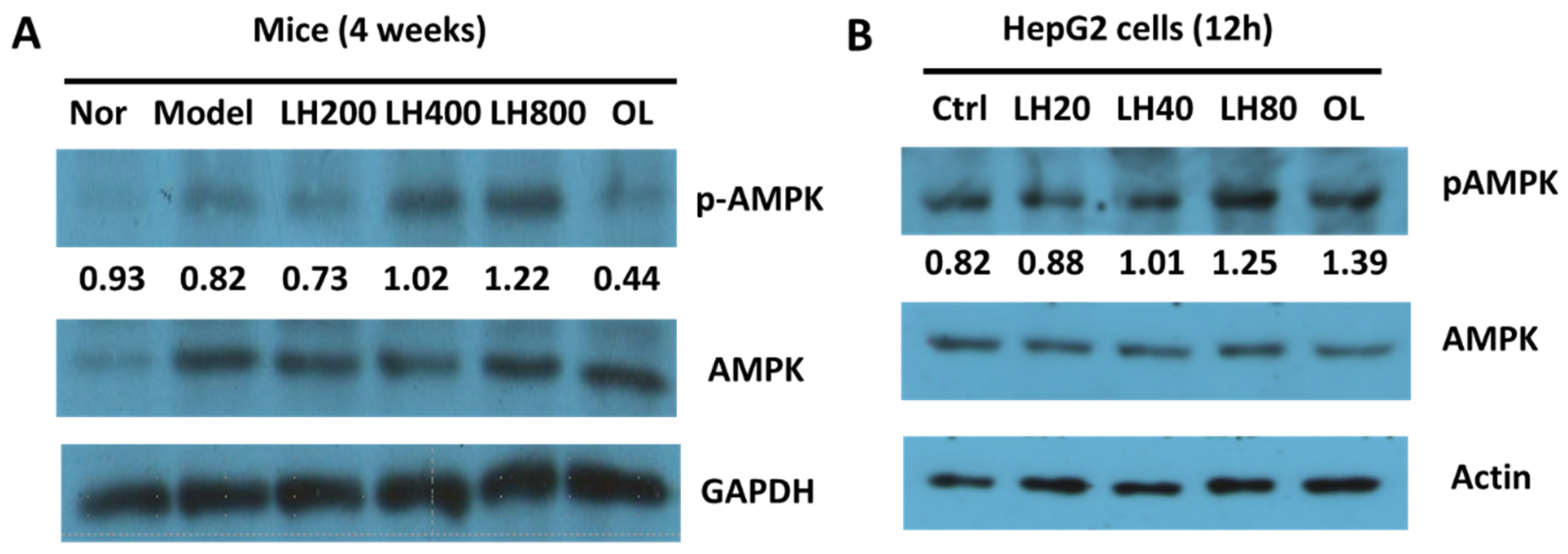
2.5. Effects of LH on Liver AMPK Phosphorylation
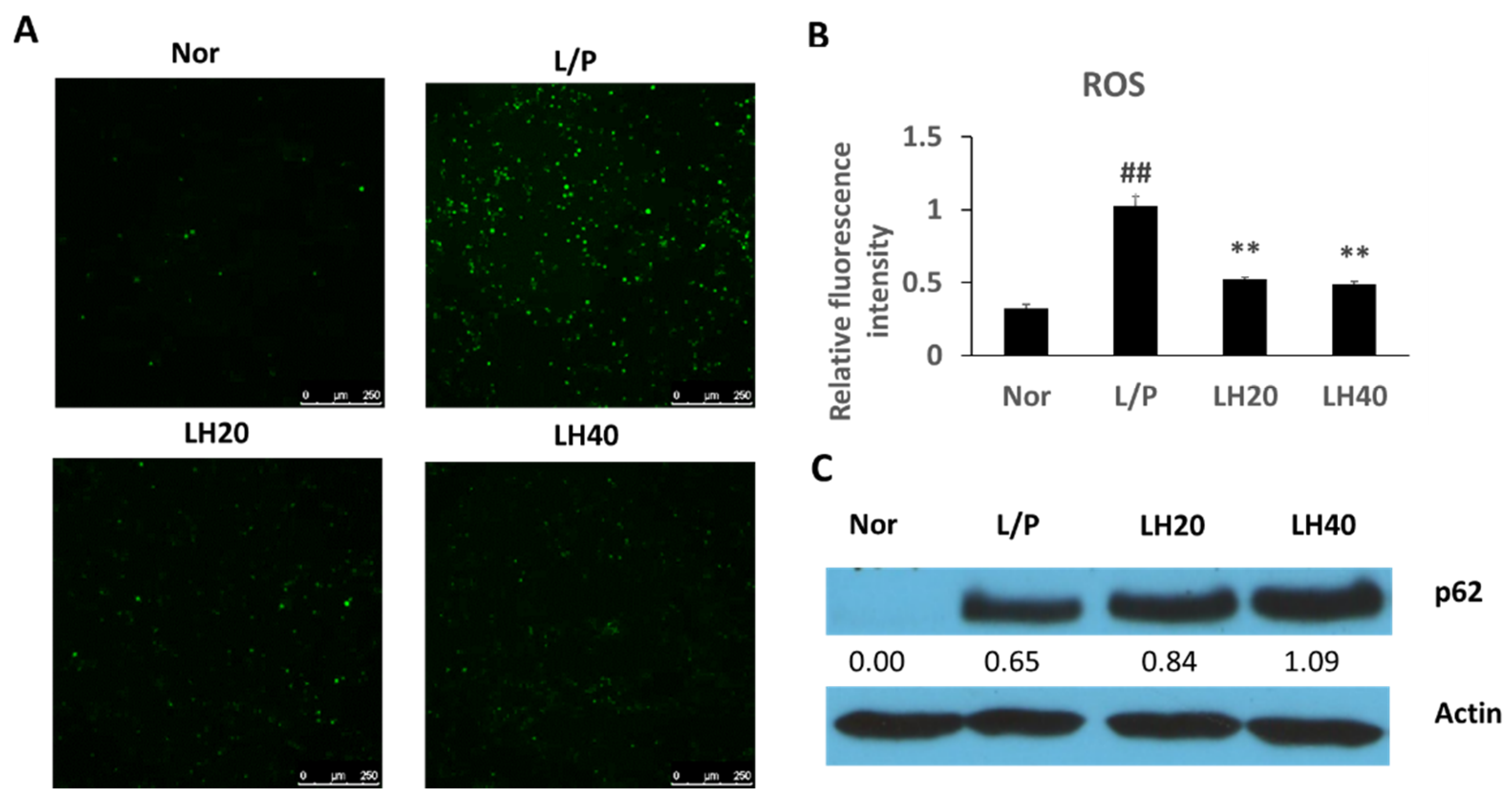
2.6. Effects of LH on Antioxidative Defenses in THP-1 Cells
3. Discussion
4. Materials and Methods
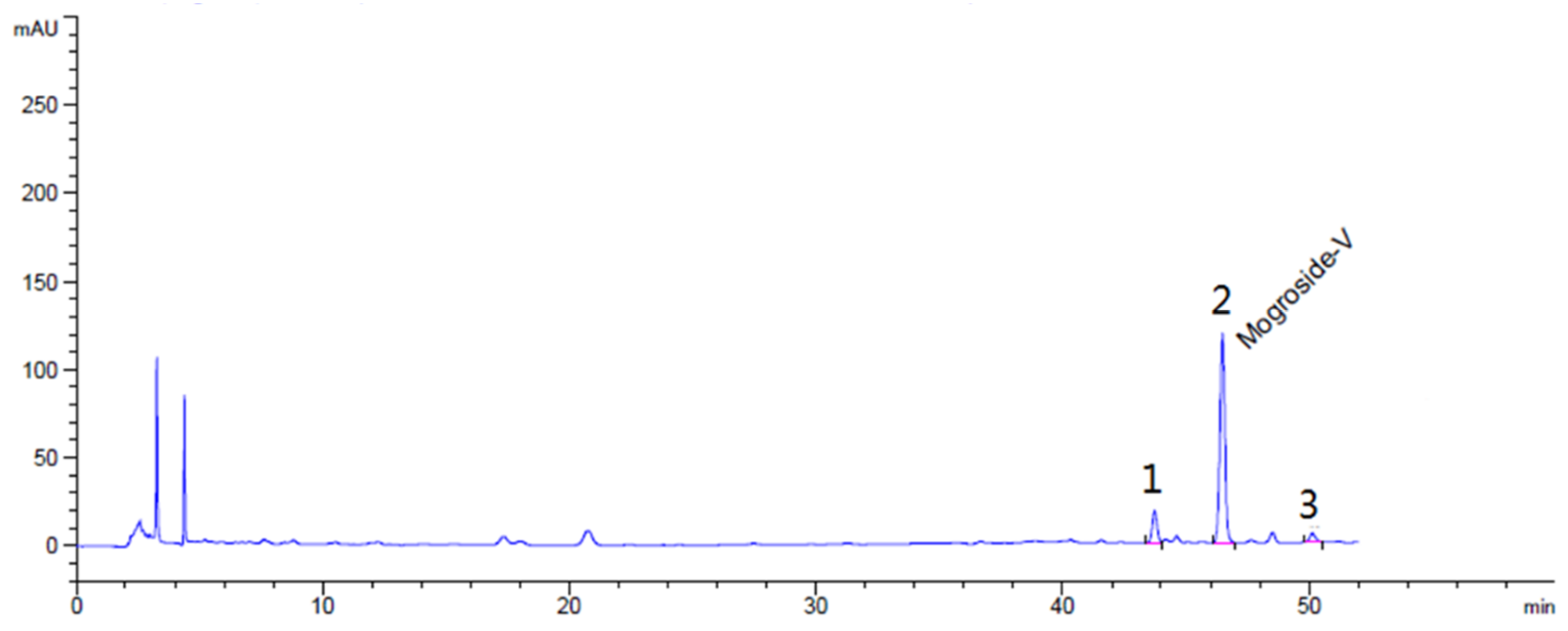
4.1. Chemicals and Reagents
4.2. High-Fat-Diet-Induced Mice
4.3. Biochemical Analysis
4.4. Cell Culture
4.5. ROS Assay
4.6. Western Blot Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Secombe, P.; Harley, S.; Chapman, M.; Aromataris, E. Feeding the critically ill obese patient: A systematic review protocol. JBI Database Syst. Rev. Implement. Rep. 2015, 13, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Seidell, J.C.; Halberstadt, J. The global burden of obesity and the challenges of prevention. Ann. Nutr. Metab. 2015, 66 (Suppl. 2), 7–12. [Google Scholar] [CrossRef] [PubMed]
- Pati, G.K.; Singh, S.P. Nonalcoholic fatty liver disease in south asia. Euroasian J. Hepato-Gastroenterol. 2016, 6, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.F.; Al-Mahtab, M.; Rahman, S.; Debnath, C.R. Non-alcoholic fatty liver disease (NAFLD)—A Review. Mymensingh Med. J. MMJ 2015, 24, 873–880. [Google Scholar] [PubMed]
- Vranesic Bender, D.; Nutrizio, M.; Josic, M.; Ljubas Kelecic, D.; Karas, I.; Premuzic, M.; Domislovic, V.; Rotim, C.; Krznaric, Z. Nutritional status and nutrition quality in patients with non-alcoholic fatty liver disease. Acta Clin. Croatica 2017, 56, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Bemis, T.; Brychta, R.; Chen, K.Y.; Courville, A.; Crayner, E.J.; Goodwin, S.; Guo, J.; Howard, L.; Knuth, N.D.; et al. Calorie for calorie, dietary fat restriction results in more body fat loss than carbohydrate restriction in people with obesity. Cell Metab. 2015, 22, 427–436. [Google Scholar] [CrossRef] [PubMed]
- White, A.H.; James, S.A.; Paulson, S.W.; Beebe, L.A. Sugar sweetened beverage consumption among adults with children in the home. Front. Nutr. 2018, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Bray, G.A.; Popkin, B.M. Dietary sugar and body weight: Have we reached a crisis in the epidemic of obesity and diabetes?: health be damned! Pour on the sugar. Diabetes Care 2014, 37, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B. Resolved: There is sufficient scientific evidence that decreasing sugar-sweetened beverage consumption will reduce the prevalence of obesity and obesity-related diseases. Obes. Rev. 2013, 14, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Dai, L.; Liu, Y.; Dou, D.; Sun, Y.; Ma, L. Pharmacological activities of mogrosides. Future Med. Chem. 2018, 10, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Krentz, A.J.; Fujioka, K.; Hompesch, M. Evolution of pharmacological obesity treatments: focus on adverse side-effect profiles. Diabetes Obes. Metab. 2016, 18, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Arch, J.R. Horizons in the pharmacotherapy of obesity. Curr. Obes. Rep. 2015, 4, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Menke, A.L.; Driessen, A.; Koek, G.H.; Lindeman, J.H.; Stoop, R.; Havekes, L.M.; Kleemann, R.; van den Hoek, A.M. Establishment of a general NAFLD scoring system for rodent models and comparison to human liver pathology. PLoS ONE 2014, 9, e115922. [Google Scholar] [CrossRef] [PubMed]
- Rej, R. Aminotransferases in disease. Clin. Lab. Med. 1989, 9, 667–687. [Google Scholar] [PubMed]
- Engin, A. Non-alcoholic fatty liver disease. Adv. Exp. Med. Biol. 2017, 960, 443–467. [Google Scholar] [PubMed]
- Tarantino, G.; Savastano, S.; Colao, A. Hepatic steatosis, low-grade chronic inflammation and hormone/growth factor/adipokine imbalance. World J. Gastroenterol. 2010, 16, 4773–4783. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, H.M.; Holloway, G.P.; Steinberg, G.R. AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: implications for obesity. Mol. Cell. Endocrinol. 2013, 366, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.K.; Marcinko, K.; Desjardins, E.M.; Lally, J.S.; Ford, R.J.; Steinberg, G.R. Treatment of nonalcoholic fatty liver disease: role of AMPK. Am. J. Physiol.-Endocrinol. Metab. 2016, 311, E730–E740. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Zheng, M.; Wang, Y.; Liu, C.; Chen, S. Protective effects and mechanisms of mogroside V on LPS-induced acute lung injury in mice. Pharm. Biol. 2014, 52, 729–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Xu, C.; Lei, F.; Liao, M.; Wang, W.; Xu, N.; Zhang, Y.; Xie, W. MiR-30a targets IL-α and regulates islet functions as an inflammation buffer and response factor. Sci. Rep. 2017, 7, 5270. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, W.; Zhong, J.; Lei, F.; Xu, N.; Zhang, Y.; Xie, W. Canagliflozin exerts anti-inflammatory effects by inhibiting intracellular glucose metabolism and promoting autophagy in immune cells. Biochem. Pharmacol. 2018, 152, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Alisi, A.; Carpino, G.; Oliveira, F.L.; Panera, N.; Nobili, V.; Gaudio, E. The role of tissue macrophage-mediated inflammation on NAFLD pathogenesis and its clinical implications. Mediat. Inflamm. 2017, 2017, 8162421. [Google Scholar] [CrossRef] [PubMed]
- Klisic, A.; Isakovic, A.; Kocic, G.; Kavaric, N.; Jovanovic, M.; Zvrko, E.; Skerovic, V.; Ninic, A. Relationship between oxidative stress, inflammation and dyslipidemia with fatty liver index in patients with type 2 diabetes mellitus. Exp. Clin. Endocrinol. Diabetes 2018, 126, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cano, M.; Handa, J.T. p62 provides dual cytoprotection against oxidative stress in the retinal pigment epithelium. Biochim. Biophys. Acta 2014, 1843, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.D.; Wang, H.; Zhang, J.F.; Kung, H.F.; Zhao, Y.N.; Zhang, Y. Proteomic profile of visceral adipose tissues between low-fat diet-fed obesity-resistant and obesity-prone C57BL/6 mice. Mol. Med. Rep. 2010, 3, 1047–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds (Mogrosides or mogroside V) are available from the authors. |









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Song, Y.; Ding, Y.; Wang, W.; Liao, L.; Zhong, J.; Sun, P.; Lei, F.; Zhang, Y.; Xie, W. Effects of Mogrosides on High-Fat-Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice. Molecules 2018, 23, 1894. https://doi.org/10.3390/molecules23081894
Zhang X, Song Y, Ding Y, Wang W, Liao L, Zhong J, Sun P, Lei F, Zhang Y, Xie W. Effects of Mogrosides on High-Fat-Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice. Molecules. 2018; 23(8):1894. https://doi.org/10.3390/molecules23081894
Chicago/Turabian StyleZhang, Xiaobing, Yunfei Song, Yipei Ding, Wei Wang, Ling Liao, Jin Zhong, Pengbo Sun, Fan Lei, Yaou Zhang, and Weidong Xie. 2018. "Effects of Mogrosides on High-Fat-Diet-Induced Obesity and Nonalcoholic Fatty Liver Disease in Mice" Molecules 23, no. 8: 1894. https://doi.org/10.3390/molecules23081894