
Construction and Quality Analysis of Transgenic Rehmannia glutinosa Containing TMV and CMV Coat Protein

Abstract
:1. Introduction
2. Results
2.1. Establishment and Optimization of Transformation System
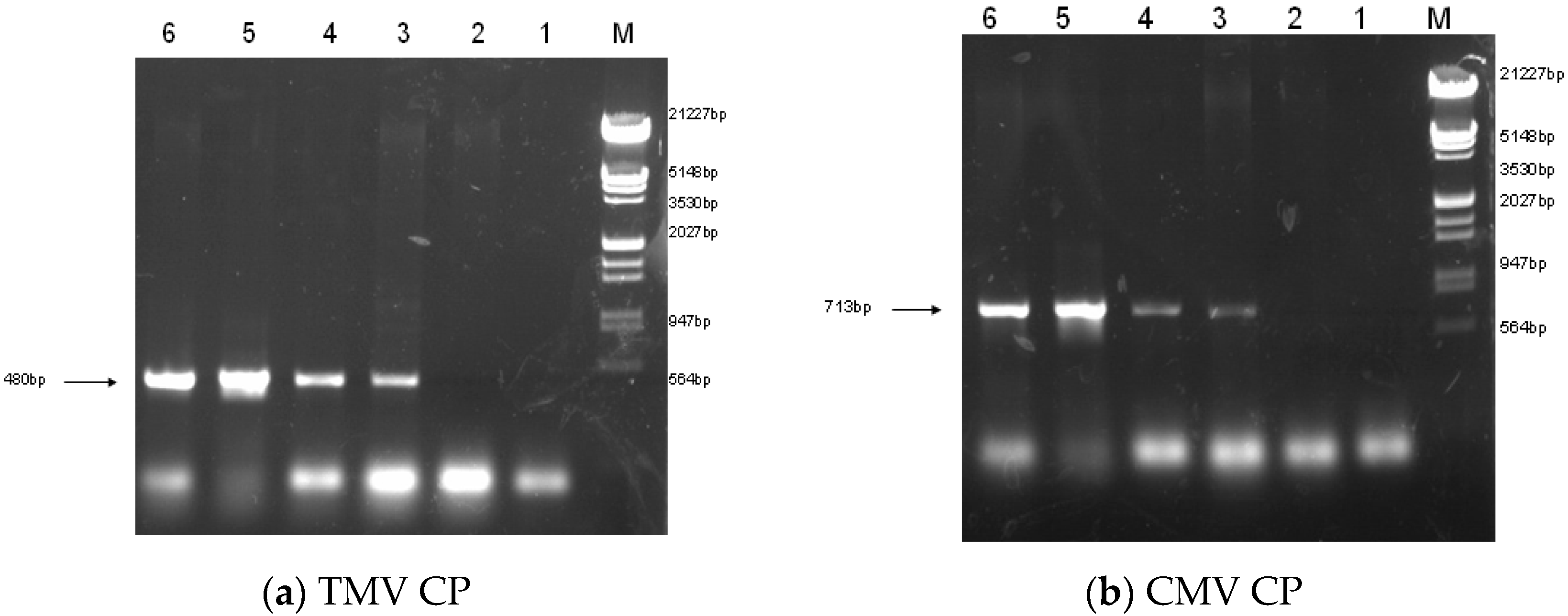
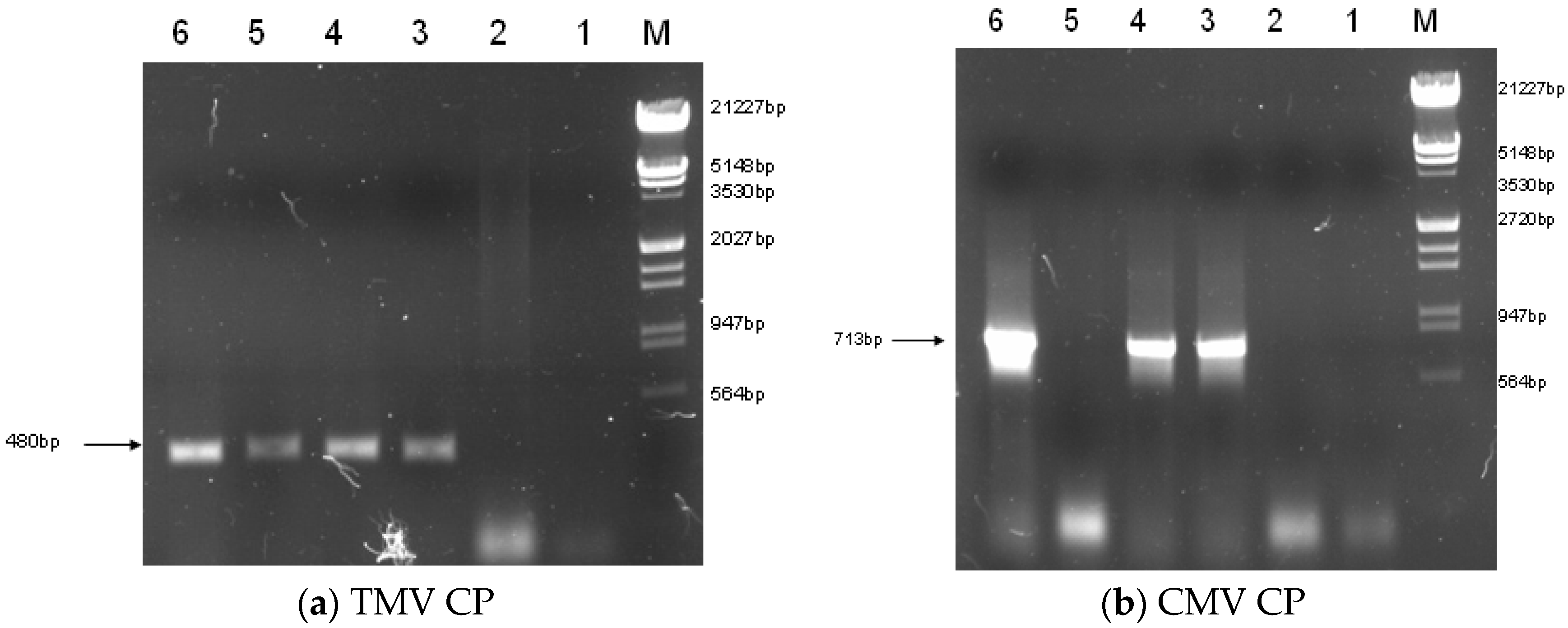
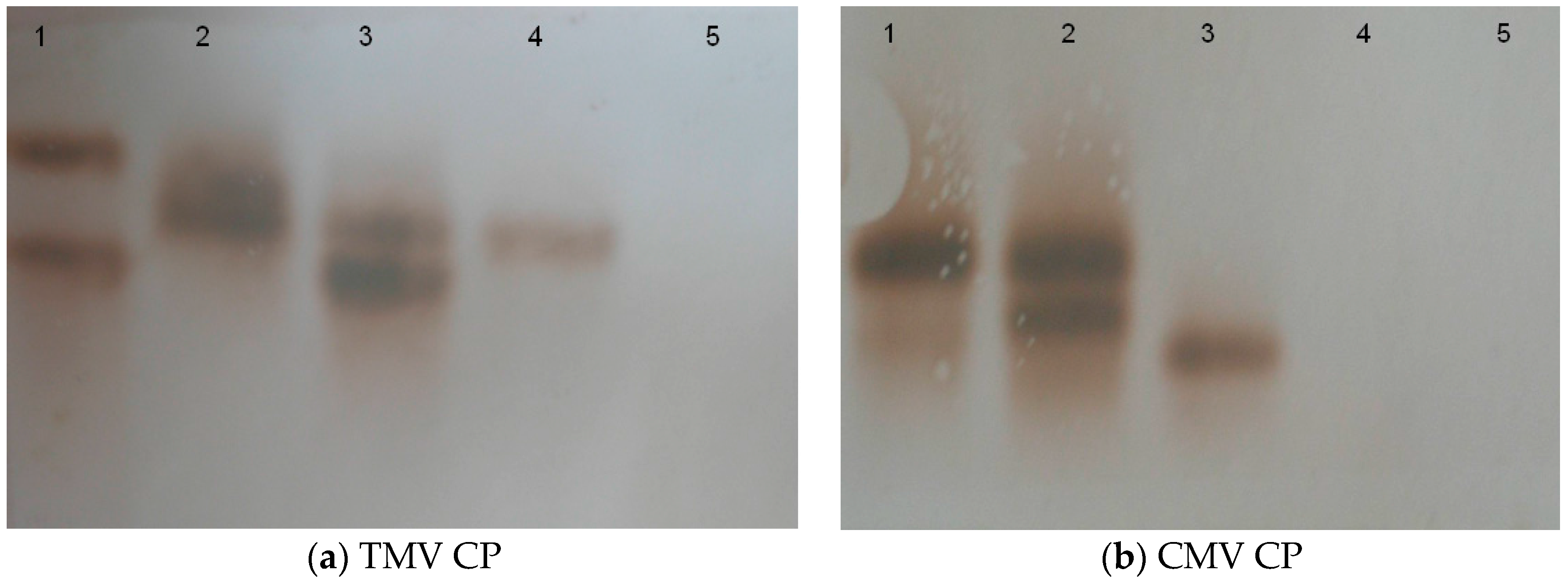
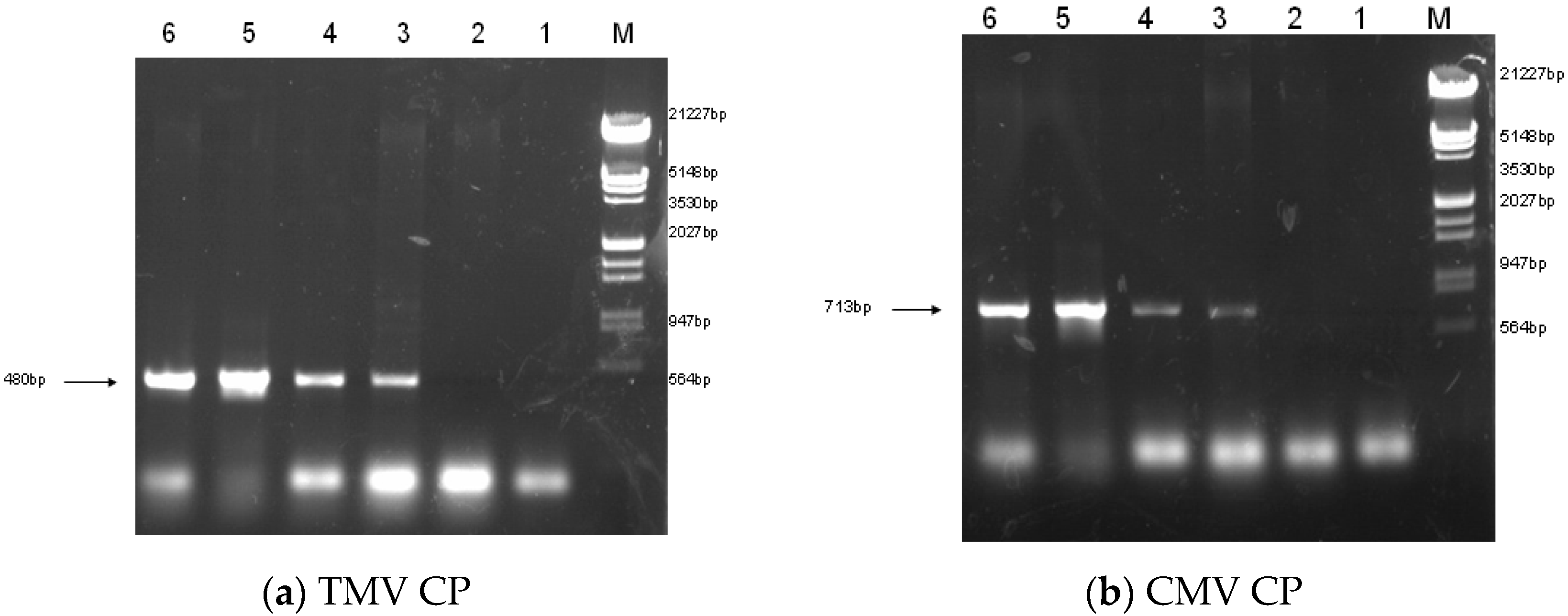
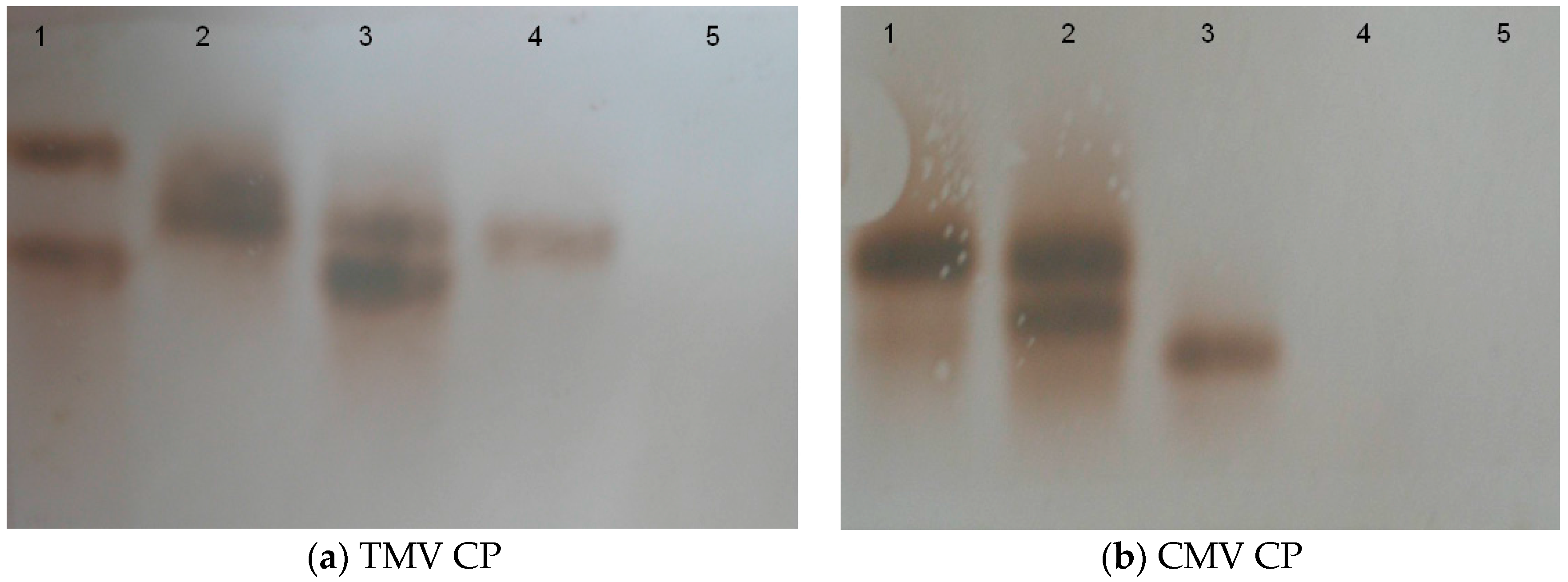
2.2. Construction of Transgenic Plants
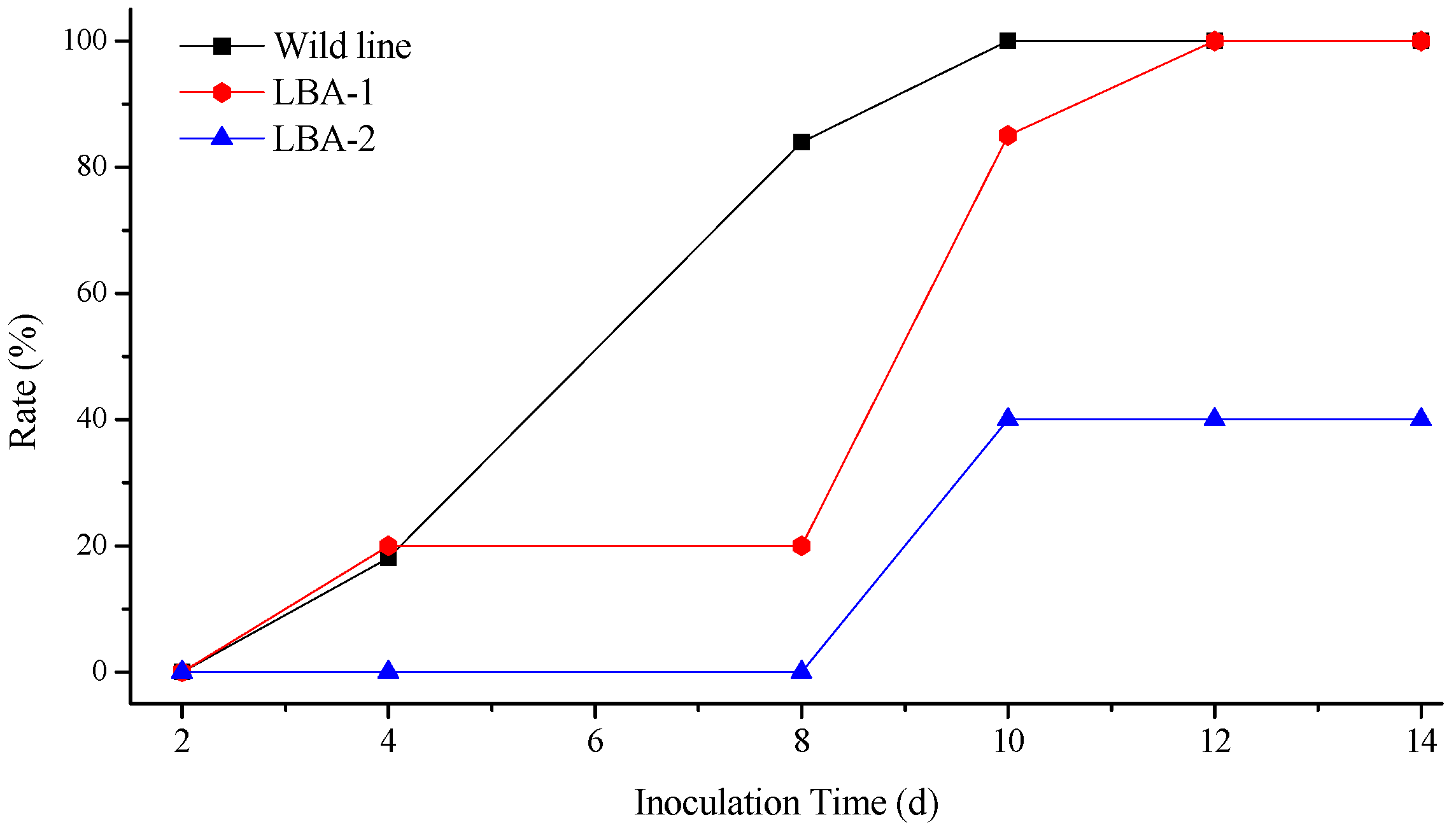
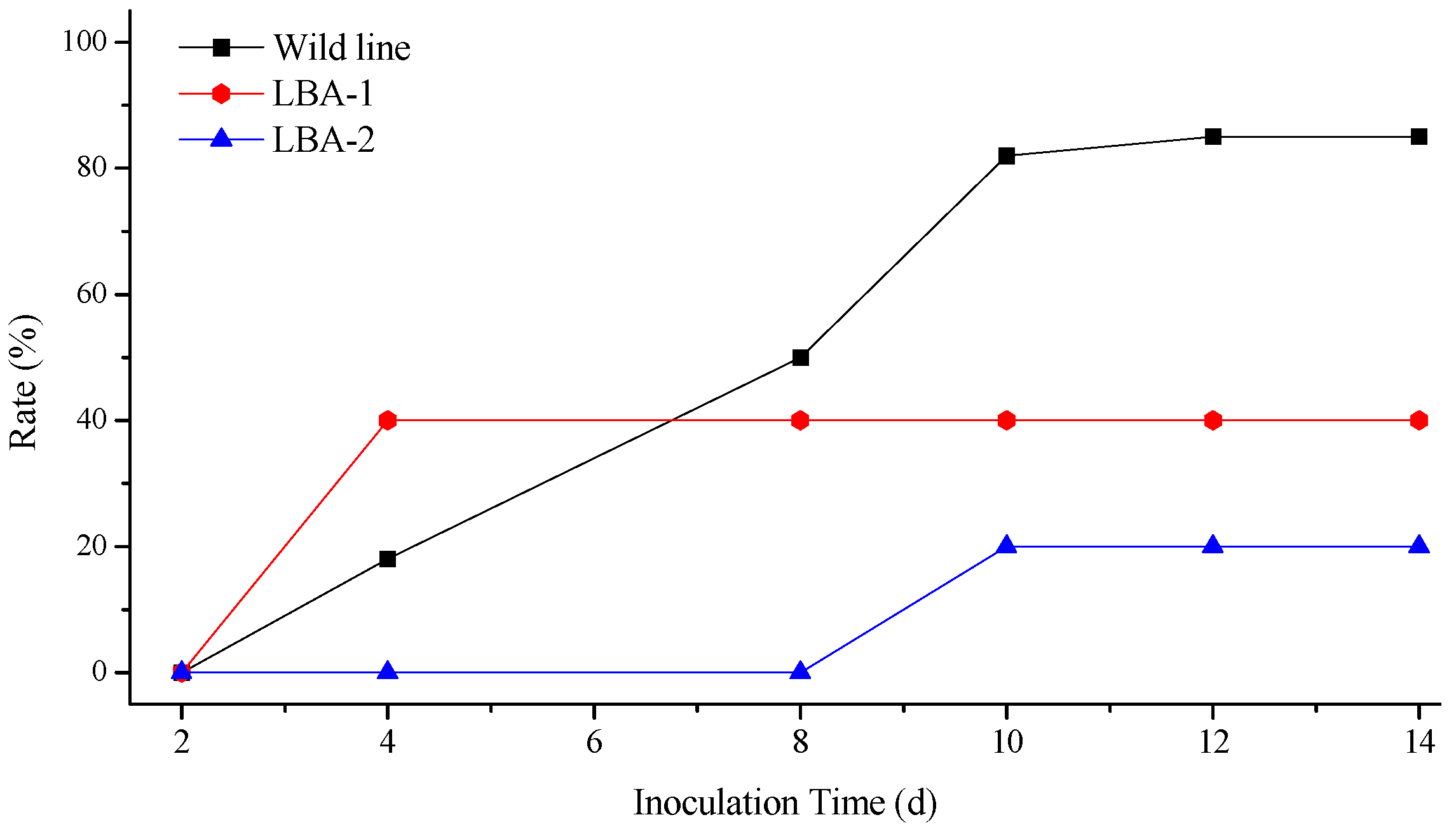
2.3. Virus Resistance of Transgenic Lines
2.4. Quality Evaluation of Transgenic Rehmannia glutinosa
3. Discussion
4. Materials and Methods
4.1. Reagents and Plant Material
4.2. Construction of the Plant Expression Plasmid
4.3. Leaf Disc Transformation of Rehmannia glutinosa
4.4. PCR of Transgenic Plants
4.5. RT-PCR of Transgenic Plants
4.6. Southern Blotting
4.7. TMV and CMV Infections
4.8. Chromatographic Fingerprint Analysis of Rehmanniae Radix
4.9. Determination of Leonurine, Echinacoside, Acteoside, Catalpol, Aucubin and Polysaccharides
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, R.-X.; Li, M.-X.; Jia, Z.-P. Rehmannia glutinosa: Review of botany, chemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 199–214. [Google Scholar] [CrossRef]
- Wang, G.; Dong, C.; Shang, Y.; Sun, Y.-A.; Fu, D.; Zhao, J. Characterization of radix rehmanniae processing procedure using FT-IR spectroscopy through nonnegative independent component analysis. Anal. Bioanal. Chem. 2009, 394, 827–833. [Google Scholar] [CrossRef]
- Feng, W.; Lv, Y.; Zheng, X.; Zhang, Y.; Cao, Y.; Pei, Y. A new megastigmane from fresh roots of Rehmannia glutinosa. Acta Pharm. Sin. B 2013, 3, 333–336. [Google Scholar] [CrossRef]
- Waisundara, V.Y.; Huang, M.; Hsu, A.; Huang, D.; Tan, B.K.-H. Characterization of the anti-diabetic and antioxidant effects of Rehmannia glutinosa in streptozotocin-induced diabetic Wistar rats. Am. J. Chin. Med. 2008, 36, 1083–1104. [Google Scholar] [CrossRef]
- Paek, K.; Yu, K.; Park, S.; Sung, N.; Park, C. Internat. Symposium on Medicinal and Aromatic Plants 390. In Micropropagation of Rehmannia glutinosa as Medicinal Plant by Shoot Tip and Root Segment Culture; International Society for Horticultural Science: Kyoto, Japan, 1994; pp. 113–120. [Google Scholar]
- Liu, Z.; Lou, Z.; Ding, X.; Li, X.; Qi, Y.; Zhu, Z.; Chai, Y. Global characterization of neutral saccharides in crude and processed Radix Rehmanniae by hydrophilic interaction liquid chromatography tandem electrospray ionization time-of-flight mass spectrometry. Food Chem. 2013, 141, 2833–2840. [Google Scholar] [CrossRef]
- Wang, M.; Li, M.; Huang, L.; Chen, Y.; Wu, Z.; Li, G.; Wei, M.; Fang, R. TMV and CMV widely infect cultivated Rehmannia glutinosa Libosch. Zhi Wu Bing Li Xue Bao 2006, 36, 189–192. [Google Scholar]
- Liao, J.; Hu, C.; Kao, J.; Deng, T. Identification of Tobacco mosaic virus infecting Rehmannia glutinosa. Plant Pathol. Bull. 2007, 16, 61–69. [Google Scholar]
- Beachy, R.N.; Loesch-Fries, S.; Tumer, N.E. Coat protein-mediated resistance against virus infection. Annu. Rev. Phytopathol. 1990, 28, 451–472. [Google Scholar] [CrossRef]
- Abel, P.P.; Nelson, R.S.; De, B.; Hoffmann, N.; Rogers, S.G.; Fraley, R.T.; Beachy, R.N. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science 1986, 232, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.-Q.; Fang, R.-X.; Shang, H.-S.; Wang, X.; Zhang, F.-L.; Li, Y.-R.; Zhang, J.-C.; Cheng, X.-Y.; Wang, G.-L.; Mang, K.-Q. Development of CMV-and TMV-resistant chili pepper: Field perfermance and biosafety assessment. Mol. Breed. 2003, 11, 25–35. [Google Scholar] [CrossRef]
- Lee, Y.H.; Jung, M.; Shin, S.H.; Lee, J.H.; Choi, S.H.; Her, N.H.; Lee, J.H.; Ryu, K.H.; Paek, K.Y.; Harn, C.H. Transgenic peppers that are highly tolerant to a new CMV pathotype. Plant Cell Rep. 2009, 28, 223–232. [Google Scholar] [CrossRef]
- Tougou, M.; Furutani, N.; Yamagishi, N.; Shizukawa, Y.; Takahata, Y.; Hidaka, S. Development of resistant transgenic soybeans with inverted repeat-coat protein genes of soybean dwarf virus. Plant Cell Rep. 2006, 25, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Tumer, N.E.; O’Connell, K.M.; Nelson, R.S.; Sanders, P.R.; Beachy, R.N.; Fraley, R.T.; Shah, D.M. Expression of alfalfa mosaic virus coat protein gene confers cross-protection in transgenic tobacco and tomato plants. EMBO J. 1987, 6, 1181–1188. [Google Scholar] [PubMed]
- Fehér, A.; Skryabin, K.G.; Balázs, E.; Preiszner, J.; Shulga, O.A.; Zakharyev, V.M.; Dudits, D. Expression of PVX coat protein gene under the control of extensin-gene promoter confers virus resistance on transgenic potato plants. Plant Cell Rep. 1992, 11, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering resistance to mixed virus infection in a commercial potato cultivar: Resistance to potato virus X and potato virus Y in transgenic Russet Burbank. Nat. Biotechnol. 1990, 8, 127–134. [Google Scholar] [CrossRef]
- Ting, H.; Meifu, K.; Wei, C.; Yunfeng, W.; Julong, C. Identification of tobacco variety resistance to Tobacco mosaic virus and Cucumber mosaic virus. Plant Prot. 2012. [Google Scholar] [CrossRef]
- Peocházková, Z. Interaction of cucumber mosaic virus and potato virus Y with tobacco mosaic virus. Biol. Plantarum 1970, 12, 297–304. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, H.; Stupak, M.; Fütterer, J.; Gruissem, W.; Zhang, P. Engineering resistance to geminiviruses—Review and perspectives. Plant Biotechnol. J. 2007, 5, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Halpin, C. Gene stacking in transgenic plants—The challenge for 21st century plant biotechnology. Plant Biotechnol. J. 2005, 3, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Van Vu, T.; Choudhury, N.R.; Mukherjee, S.K. Transgenic tomato plants expressing artificial microRNAs for silencing the pre-coat and coat proteins of a begomovirus, Tomato leaf curl New Delhi virus, show tolerance to virus infection. Virus Res. 2013, 172, 35–45. [Google Scholar]
- Hackland, A.; Rybicki, E.; Thomson, J. Coat protein-mediated resistance in transgenic plants. Arch. Virol. 1994, 139, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Schillberg, S.; Zimmermann, S.; Zhang, M.-Y.; Fischer, R. Antibody-based resistance to plant pathogens. Transgenic Res. 2001, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Doreste, V.; Ramos, P.; Enriquez, G.; Rodriguez, R.; Peral, R.; Pujol, M. Transgenic potato plants expressing the potato virus X (PVX) coat protein gene developed resistance to the viral infection. Phytoparasitica 2002, 30, 177–185. [Google Scholar] [CrossRef]
- Woerdenbag, H.J.; Lugt, C.B.; Pras, N. Artemisia annua L.: A source of novel antimalarial drugs. Pharm. Weekbl. 1990, 12, 169–181. [Google Scholar] [CrossRef]
- Xie, P.; Chen, S.; Liang, Y.-Z.; Wang, X.; Tian, R.; Upton, R. Chromatographic fingerprint analysis—A rational approach for quality assessment of traditional Chinese herbal medicine. J. Chrom. A 2006, 1112, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bauer, R.; Melchart, D.; Xiao, P.-G.; Staudinger, A. Chromatographic Fingerprint Analysis of Herbal Medicines; Springer: Berlin, Gemany, 2011. [Google Scholar]
- Novak, W.K.; Haslberger, A.G. Substantial equivalence of antinutrients and inherent plant toxins in genetically modified novel foods. Food Chem. Toxicol. 2000, 38, 473–483. [Google Scholar] [CrossRef]
- Kuiper, H.A.; Kleter, G.A.; Noteborn, H.P.; Kok, E.J. Substantial equivalence—An appropriate paradigm for the safety assessment of genetically modified foods? Toxicology 2002, 181, 427–431. [Google Scholar] [CrossRef]
- Parani, M.; Rudrabhatla, S.; Myers, R.; Weirich, H.; Smith, B.; Leaman, D.W.; Goldman, S.L. Microarray analysis of nitric oxide responsive transcripts in Arabidopsis. Plant Biotechnol. J. 2004, 2, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Candelier-Harvey, P.; Hull, R. Cucumber mosaic virus genome is encapsidated in alfalfa mosaic virus coat protein expressed in transgenic tobacco plants. Transgenic Res. 1993, 2, 277–285. [Google Scholar] [CrossRef]
- Xu, J.; Wu, J.; Zhu, L.Y.; Shen, H.; Xu, J.D.; Jensen, S.R.; Jia, X.B.; Zhang, Q.W.; Li, S.L. Simultaneous determination of iridoid glycosides, phenethylalcohol glycosides and furfural derivatives in Rehmanniae Radix by high performance liquid chromatography coupled with triple-quadrupole mass spectrometry. Food Chem. 2012, 135, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | Mean |
|---|---|---|---|---|---|---|---|---|---|---|---|
| LBA-1 | 0.899 | 0.941 | 0.893 | 0.916 | 0.918 | 0.989 | 0.99 | 0.987 | 0.991 | 0.982 | 0.951 |
| LBA-2 | 0.940 | 0.953 | 0.899 | 0.910 | 0.902 | 0.937 | 0.926 | 0.928 | 0.991 | 0.987 | 0.937 |
| Lines | Polysaccharides (mg/g) | Main Compounds (mg/g) | ||||
|---|---|---|---|---|---|---|
| Aucubin | Catalpol | Ajugol | Acteoside | Echinacoside | ||
| Wild lines | 33.66 ± 1.82 | 17.80 ± 2.88 | 683.57 ± 120.12 | 189.21 ± 22.49 | 1.25 ± 0.45 | 5.71 ± 1.34 |
| LBA-1 | 33.47 ± 1.61 | 17.84 ± 3.55 | 687.20 ± 112.35 | 186.93 ± 18.11 | 1.25 ± 0.46 | 5.74 ± 2.34 |
| LBA-2 | 33.22 ± 1.69 | 17.76 ± 3.75 | 682.67 ± 132.77 | 187.59 ± 22.12 | 1.26 ± 0.28 | 5.73 ± 1.60 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, Z.; Shen, Y.; Li, J.; Lin, Z.; Chen, M.; Wang, M.; Li, M.; Dong, H.; Huang, L. Construction and Quality Analysis of Transgenic Rehmannia glutinosa Containing TMV and CMV Coat Protein. Molecules 2016, 21, 1134. https://doi.org/10.3390/molecules21091134
Teng Z, Shen Y, Li J, Lin Z, Chen M, Wang M, Li M, Dong H, Huang L. Construction and Quality Analysis of Transgenic Rehmannia glutinosa Containing TMV and CMV Coat Protein. Molecules. 2016; 21(9):1134. https://doi.org/10.3390/molecules21091134
Chicago/Turabian StyleTeng, Zhongqiu, Ye Shen, Jing Li, Zhongping Lin, Min Chen, Min Wang, Man Li, Hongran Dong, and Luqi Huang. 2016. "Construction and Quality Analysis of Transgenic Rehmannia glutinosa Containing TMV and CMV Coat Protein" Molecules 21, no. 9: 1134. https://doi.org/10.3390/molecules21091134