
Anti-Enterovirus 71 Agents of Natural Products


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Flavonoids
2.1. Flavonols

2.2. Flavones
2.3. Flavone Glycosides
2.4. Isoflavones
2.5. Xanthones
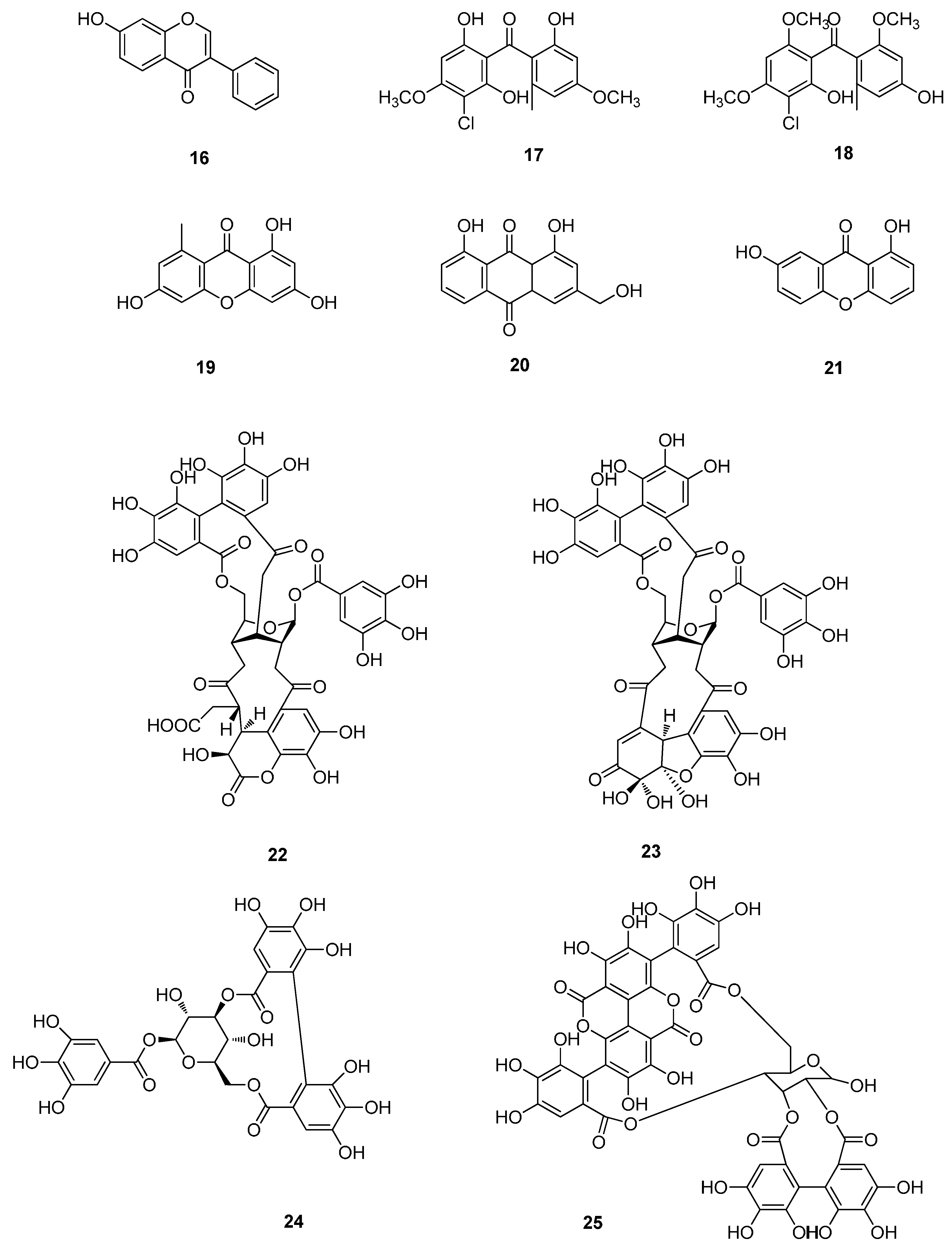
3. Polyphenols
3.1. Tannins
3.2. Polyphenols
4. Terpenoids
4.1. Monoterpenoid Glycoside

4.2. Sesquiterpenoids
4.3. Sesterterpenoids
4.4. Triterpenoids
4.4.1. Triterpenoids

4.4.2. Triterpenoids Glycosides
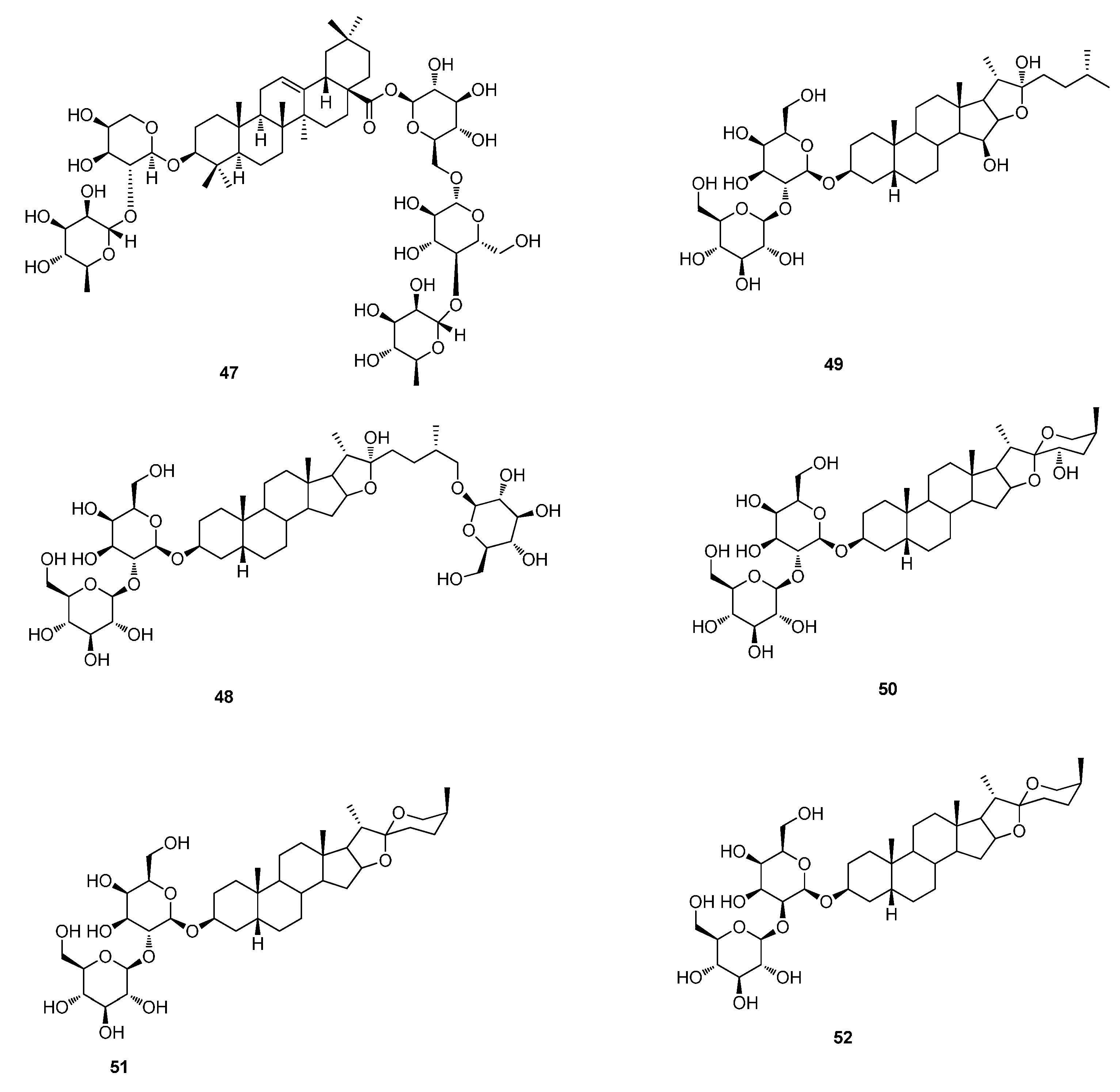
5. Steroids
5.1. Steroids Glycosides
5.2. Steroids

6. Alkaloids
7. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| Enterovirus 71 | EV71 |
| Natural product | NP |
| Hand, foot and mouth disease | HFMD |
| Cytopathic effect | CPE |
References
- Shang, L.; Xu, M.; Yin, Z. Antiviral drug discovery for the treatment of enterovirus 71 infections. Antivir. Res. 2013, 97, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.Y.; Lin, L.; Yang, Y.; Zhang, S.X.; Kong, J.Q.; Cheng, K.D.; Zhao, Y.F.; Wang, W. Advances in functional studies of nonstructural proteins and development of antiviral agents for enterovirus 71. Yaoxue Xuebao 2011, 46, 753–761. [Google Scholar]
- McMinn, P.C. Recent advances in the molecular epidemiology and control of human enterovirus 71 infection. Curr. Opin. Virol. 2012, 2, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kuo, R.L.; Shih, S.R. Strategies to develop antivirals against enterovirus 71. Virol. J. 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.X.; Ng, M.M.L.; Chu, J.J.H. Developments towards antiviral therapies against enterovirus 71. Drug Discov. Today 2010, 15, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Lai, J.K.F.; Sam, I.C.; Chan, Y.F. Recent developments in antiviral agents against enterovirus 71 infection. J. Biomed. Sci. 2014, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.C.; Weng, K.F.; Chang, S.C.; Lin, J.Y.; Huang, P.N.; Shih, S.R. Development of antiviral agents for enteroviruses. J. Antimicrob. Chemother. 2008, 62, 1169–1173. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.C.; Wang, Y.; Liu, Y.P.; Zhang, R.Q.; Li, X.; Su, W.H.; Long, F.; Luo, X.D.; Peng, T. Inhibition of enterovirus 71 replication by chrysosplenetin and penduletin. Eur. J. Pharm. Sci. 2011, 44, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, T.; Du, J.; Cui, S.; Yang, F.; Jin, Q. Anti-Enterovirus 71 Effects of Chrysin and Its Phosphate Ester. PLoS ONE 2014, 9, e89668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Qiao, H.; Lv, Y.; Wang, J.; Chen, X.; Hou, Y.; Tan, R.; Li, E. Apigenin Inhibits Enterovirus-71 Infection by Disrupting Viral RNA Association with trans-Acting Factors. PLoS ONE 2014, 9, e110429. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.J.; Lin, C.W.; Lai, C.C.; Lan, Y.C.; Lai, C.H.; Hung, C.H.; Hsueh, K.C.; Lin, T.H.; Chang, H.C.; Wan, L.; et al. Kaempferol inhibits enterovirus 71 replication and internal ribosome entry site (IRES) activity through FUBP and HNRP proteins. Food Chem. 2011, 128, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.Y.; Cheng, M.L.; Weng, S.F.; Leu, Y.L.; Chiu, D.T.Y. Antiviral Effect of Epigallocatechin Gallate on Enterovirus 71. J. Agric. Food Chem. 2009, 57, 6140–6147. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Su, W.; Jin, J.; Chen, J.; Li, X.; Zhang, X.; Sun, M.; Sun, S.; Fan, P.; An, D.; et al. Identification of Luteolin as Enterovirus 71 and Coxsackievirus A16 Inhibitors through Reporter Viruses and Cell Viability-Based Screening. Viruses 2014, 6, 2778–2795. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, D.; Ge, M.; Li, Z.; Jiang, J.; Li, Y. Formononetin inhibits enterovirus 71 replication by regulating COX-2/PGE(2) expression. Virol. J. 2015, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Meng, S.; Li, Z.R.; Peng, Z.G.; Han, Y.X.; Guo, S.S.; Cui, X.L.; Li, Y.H.; Jiang, J.D. The antiviral effect of 7-hydroxyisoflavone against Enterovirus 71 in vitro. J. Asian Nat. Prod. Res. 2012, 15, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Lin, X.; Lu, X.; Wan, J.; Zhou, X.; Liao, S.; Tu, Z.; Xu, S.; Liu, Y. Sesquiterpenoids and xanthones derivatives produced by sponge-derived fungus Stachybotry sp. HH1 ZSDS1F1-2. J. Antibiot. 2015, 68, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Wu, C.F.; Hsiao, N.W.; Chang, C.Y.; Li, S.W.; Wan, L.; Lin, Y.J.; Lin, W.Y. Aloe-emodin is an interferon-inducing agent with antiviral activity against Japanese encephalitis virus and enterovirus 71. Int. J. Antimicrob. Agents 2008, 32, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tao, L.; Fu, W.W.; Liang, S.; Yang, Y.F.; Yuan, Q.H.; Yang, D.J.; Lu, A.P.; Xu, H.X. Prenylated Benzoylphloroglucinols and Xanthones from the Leaves of Garcinia oblongifolia with Antienteroviral Activity. J. Nat. Prod. 2014, 77, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiu, J.; Liu, J.; Zhang, L.; Li, X.; Xu, Y.; Qin, C.; Zhang, L. Chebulagic Acid, a Hydrolyzable Tannin, Exhibited Antiviral Activity in Vitro and in Vivo against Human Enterovirus 71. Int. J. Mol. Sci. 2013, 14, 9618–9627. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.J.; Zhang, L.F.; Fan, X.X.; Qin, C.; Liu, J.N. Antiviral effect of geraniin on human enterovirus 71 in vitro and in vivo. Bioorg. Med. Chem. Lett. 2012, 22, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.G.; Song, J.H.; Hong, E.H.; Lee, B.R.; Kwon, Y.S.; Chang, S.Y.; Kim, S.H.; Lee, S.W.; Park, J.H.; Ko, H.J. Antiviral effects of Phyllanthus urinaria containing corilagin against human enterovirus 71 and Coxsackievirus A16 in vitro. Arch. Pharm. Res. 2015, 38, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiu, J.; Zhang, L.; Qin, C.; Liu, J. Antiviral activity of punicalagin toward human enterovirus 71 in vitro and in vivo. Phytomedicine 2012, 20, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Song, J.H.; Park, K.S.; Baek, S.H. In vitro anti-enterovirus 71 activity of gallic acid from Woodfordia fruticosa flowers. Lett. Appl. Microb. 2010, 50, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Y.; Gu, Z.; Wang, Y.; Shi, M.; Ji, Y.; Sun, J.; Xu, X.; Zhang, L.; Jiang, J.; et al. Resveratrol Inhibits Enterovirus 71 Replication and Pro-Inflammatory Cytokine Secretion in Rhabdosarcoma Cells through Blocking IKKs/NF-kappa B Signaling Pathway. PLoS ONE 2015, 10, e0116879. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.L.; Chen, Q.; Li, J.D.; Zhu, W.C.; Guo, J.; Yu, S.Y. Inhibitory effect of resveratrol against enterovirus type 71 in vitro. J. South. Med. Univ. 2011, 31, 665–668. [Google Scholar]
- Hung, H.C.; Chen, T.C.; Fang, M.Y.; Yen, K.J.; Shih, S.R.; Hsu, J.T.A.; Tseng, C.P. Inhibition of enterovirus 71 replication and the viral 3D polymerase by aurintricarboxylic acid. J. Antimicrob. Chemother. 2010, 65, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Zhao, C.H.; Yang, Z. Methyl 3,4-dihydroxyphenylacetate prevents enterovirus 71 proliferation in rhabdomyosarcoma cells. Yaoxue Xuebao 2012, 47, 1257–1260. [Google Scholar]
- Wang, J.F.; Wei, X.Y.; Lu, X.; Xu, F.Q.; Wan, J.T.; Lin, X.P.; Zhou, X.F.; Liao, S.R.; Yang, B.; Tu, Z.C.; et al. Eight new polyketide metabolites from the fungus Pestalotiopsis vaccinii endogenous with the mangrove plant Kandelia candel (L.) Druce. Tetrahedron 2014, 70, 9695–9701. [Google Scholar] [CrossRef]
- Lin, Y.J.; Lai, C.C.; Lai, C.H.; Sue, S.C.; Lin, C.W.; Hung, C.H.; Lin, T.H.; Hsu, W.Y.; Huang, S.M.; Hung, Y.L.; et al. Inhibition of enterovirus 71 infections and viral IRES activity by Fructus gardeniae and geniposide. Eur. J. Med. Chem. 2013, 62, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Lin, J.; Zhang, T.; Tu, Z.; et al. Cytotoxic and antiviral nitrobenzoyl sesquiterpenoids from the marine-derived fungus Aspergillus ochraceus Jcma1F17. Med. Chem. Commun. 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Choi, H.J.; Lim, C.H.; Song, J.H.; Baek, S.H.; Kwon, D.H. Antiviral activity of raoulic acid from Raoulia australis against Picornaviruses. Phytomedicine 2009, 16, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Chiang, L.C.; Ng, L.T.; Cheng, P.W.; Chiang, W.; Lin, C.C. Antiviral activities of extracts and selected pure constituents of Ocimum basilicum. Clin. Exp. Pharmacol. Physiol. 2005, 32, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.H.; Xu, J.; Zhang, Y.Q.; Zhao, L.X.; Feng, B. Inhibition of Human Enterovirus 71 Replication by Pentacyclic Triterpenes and Their Novel Synthetic Derivatives. Chem. Pharm. Bull. 2014, 62, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tao, J.; Yang, X.; Yang, Z.; Zhang, L.; Liu, H.; Wu, K.; Wu, J. Antiviral effects of two Ganoderma lucidum triterpenoids against enterovirus 71 infection. Biochem. Biophys. Res. Commun. 2014, 449, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, X.; Wang, W.; Zhang, Y.; Yang, Z.; Jin, Y.; Ge, H.M.; Li, E.; Yang, G. Glycyrrhizic acid as the antiviral component of Glycyrrhiza uralensis Fisch against coxsackievirus A16 and enterovirus 71 of hand foot and mouth disease. J. Ethnopharm. 2013, 147, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Choi, H.J.; Song, H.H.; Hong, E.H.; Lee, B.R.; Oh, S.R.; Choi, K.; Yeo, S.G.; Lee, Y.P.; Cho, S.; et al. Antiviral activity of ginsenosides against coxsackievirus B3, enterovirus 71, and human rhinovirus 3. J. Ginseng Res. 2014, 38, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Yeo, S.G.; Hong, E.H.; Lee, B.R.; Kim, J.W.; Kim, J.; Jeong, H.; Kwon, Y.; Kim, H.; Lee, S.; et al. Antiviral Activity of Hederasaponin B from Hedera helix against Enterovirus 71 Subgenotypes C3 and C4a. Biomol. Ther. 2014, 22, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Tao, L.; Chau, S.L.; Wu, R.; Zhang, H.; Yang, Y.; Yang, D.; Bian, Z.; Lu, A.; Han, Q.; et al. Folding fan mode counter-current chromatography offers fast blind screening for drug discovery. Case study: Finding anti-enterovirus 71 agents from Anemarrhena asphodeloides. J. Chromatogr. A 2014, 1368, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, L.; Sun, S.; Zhang, H.; Ma, T.; Su, W.; Jiang, C. Identification of Cinobufagin and Resibufogenin as Inhibitors of Enterovirus 71 Infection. Chem. Res. Chin. Univ. 2014, 30, 953–958. [Google Scholar] [CrossRef]
- Yang, Y.; Xiu, J.; Zhang, X.; Zhang, L.; Yan, K.; Qin, C.; Liu, J. Antiviral Effect of Matrine against Human Enterovirus 71. Molecules 2012, 17, 10370–10376. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, Y.; Xu, Y.; Ma, C.; Qin, C.; Zhang, L. Lycorine reduces mortality of human enterovirus 71-infected mice by inhibiting virus replication. Virol. J. 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ma, J.; Xiu, J.; Bai, L.; Guan, F.; Zhang, L.; Liu, J.; Zhang, L. Deferoxamine Compensates for Decreases in B Cell Counts and Reduces Mortality in Enterovirus 71-Infected Mice. Mar. Drugs 2014, 12, 4086–4095. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Wang, J.; Wang, L.; Ma, S.; Liu, Y. Anti-Enterovirus 71 Agents of Natural Products. Molecules 2015, 20, 16320-16333. https://doi.org/10.3390/molecules200916320
Wang L, Wang J, Wang L, Ma S, Liu Y. Anti-Enterovirus 71 Agents of Natural Products. Molecules. 2015; 20(9):16320-16333. https://doi.org/10.3390/molecules200916320
Chicago/Turabian StyleWang, Liyan, Junfeng Wang, Lishu Wang, Shurong Ma, and Yonghong Liu. 2015. "Anti-Enterovirus 71 Agents of Natural Products" Molecules 20, no. 9: 16320-16333. https://doi.org/10.3390/molecules200916320