Sulfoxaflor and Natural Pyrethrin with Piperonyl Butoxide Are Effective Alternatives to Neonicotinoids against Juveniles of Philaenus spumarius, the European Vector of Xylella fastidiosa

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Pesticide Screening in Laboratory Conditions
2.3. Greenhouse Experiments under Semi-Field Conditions
2.4. Statistics
3. Results
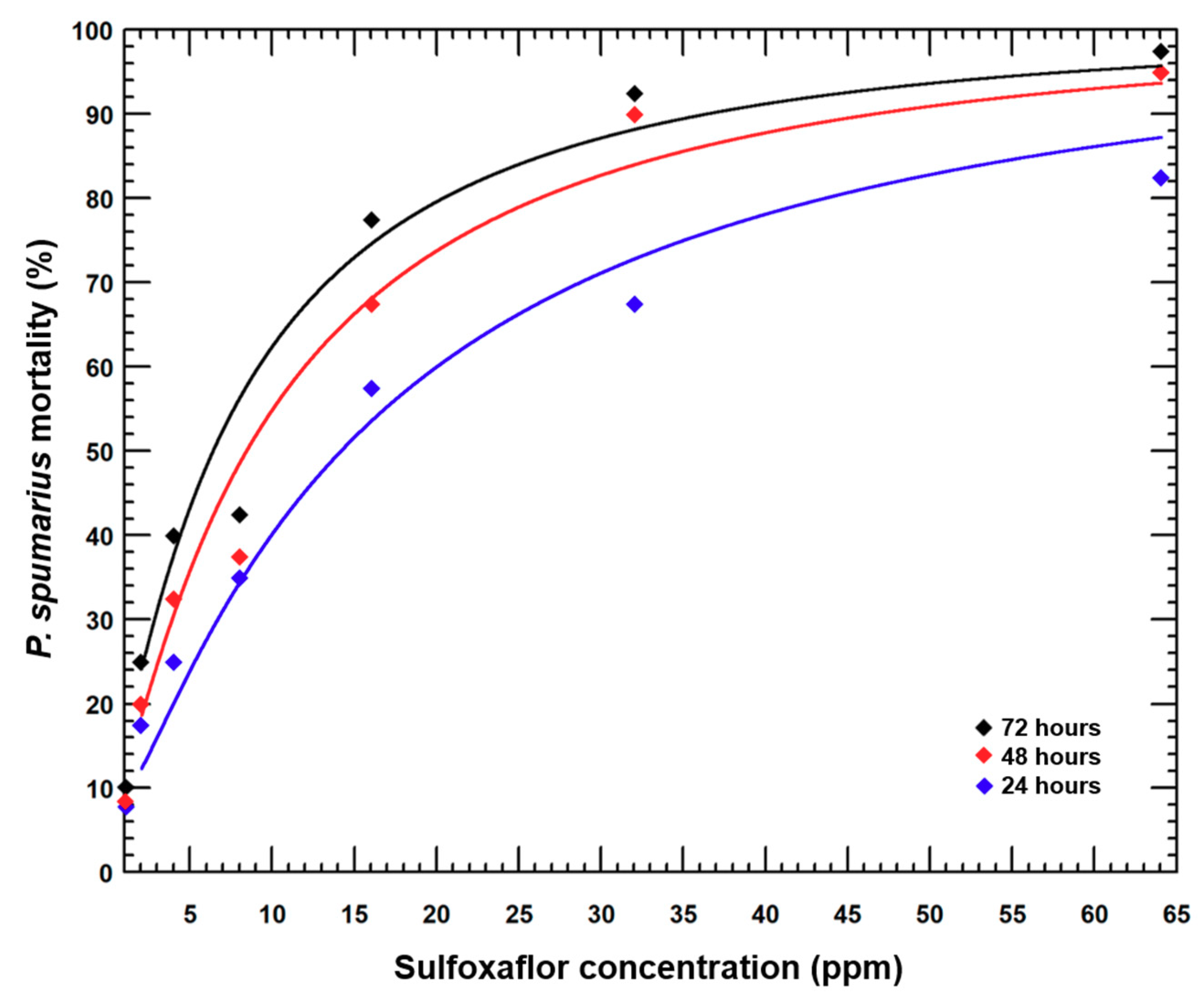
3.1. Sulfoxaflor Efficacy under Laboratory Conditions
3.2. Sulfoxaflor Efficacy under Glasshouse Conditions
3.3. Pesticide Screening under Laboratory Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wells, J.M.; Raju, B.C.; Hung, H.Y.; Weisburg, W.G.; Mandelco-Paul, L.; Brenner, D.J. Xylella-fastidiosa gen. nov., sp. nov: Gram-negative, xylem-limited, fastidious plant bacteria related to Xanthomonas spp. Int. J. Bacteriol. 1987, 37, 136–143. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory, with the identification and evaluation of risk reduction options. EFSA J. 2015, 13, 1–262. [Google Scholar]
- Tsagkarakis, A.E.; Afentoulis, D.G.; Matared, M.; Thanou, Z.N.; Stamatakou, G.D.; Kalaitzaki, A.P.; Tzobanoglou, D.K.; Goumas, D.; Trantas, E.; Zarboutis, I.; et al. Identification and seasonal abundance of Auchenorrhyncha with a focus on potential insect vectors of Xylella fastidiosa in olive orchards in three regions of Greece. J. Econ. Entomol. 2018, 111, 2536–2545. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.P.P.; Nunney, L. How do plant diseases caused by Xylella fastidiosa emerge? Plant Dis. 2015, 99, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Cornara, D.; Cavalieri, V.; Dongiovanni, C.; Altamura, G.; Palmisamno, F.; Bosco, D.; Porcelli, F.; Almeida, R.P.P.; Saponari, M. Transmission of Xylella fastidosa by natural infected Philaenus spumarius (Hemioptera, aphrophoridae) to different host plants. J. Appl. Entomol. 2017, 141, 80–87. [Google Scholar] [CrossRef]
- Overall, L.M.; Rebek, E.J. Insect vectors and current management strategies for diseases caused by Xylella fastidiosa in the Southern United States. J. Integr. Pest Manag. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Purcell, A.H. Xylella fastidiosa: Cause of Pierce’s disease of grapevine and other emergent diseases. Plant Dis. 2002, 86, 1056–1066. [Google Scholar] [CrossRef]
- Chang, C.J.; Garnier, M.; Zreik, L.; Rossetti, V.; Bové, J.M. Culture and serological detection of the xylem-limited bacterium causing citrus variegated chlorosis and its identification as a strain of Xylella fastidiosa. Curr. Microbiol. 1993, 27, 137–142. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Protection Organization). EPPO Global Database, Xylella Fastidiosa. Available online: https://gd.eppo.int/taxon/XYLEFA/distribution (accessed on 1 February 2019).
- Almeida, R.P.P. Can Apulia’s olive trees be saved? Science 2016, 353, 346–348. [Google Scholar] [CrossRef]
- Martelli, G.P.; Boscia, D.; Porcelli, F.; Saponari, M. The olive quick decline syndrome in south-east Italy: A threatening phytosanitary emergency. Eur. J. Plant Pathol. 2015, 144, 235–243. [Google Scholar] [CrossRef]
- Olmo, D.; Nieto, A.; Adrover, F.; Urbano, A.; Beidas, O.; Juan, A.; Marco-Noales, E.; López, M.M.; Navarro, I.; Monterde, A.; et al. First detection of Xylella fastidiosa infecting cherry (Prunus avium) and Polygala myrtifolia plants in Mallorca Island, Spain. Plant Dis. 2017, 101, 1820. [Google Scholar] [CrossRef]
- Morente, M.; Cornara, D.; Plaza, M.; Durán, J.M.; Capiscol, C.; Trillo, R.; Ruiz, M.; Ruz, C.; Sanjuan, S.; Pereira, J.A.; et al. Distribution and relative abundance of insect vectors of Xylella fastidiosa in olive groves of the Iberian peninsula. Insects 2018, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Redak, R.A.; Purcell, A.H.; Lopes, J.R.; Blua, M.J.; Mizell, R.F., III; Andersen, P.C. The biology of xylem fluid-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Tubajika, K.M.; Civerolo, E.L.; Purteka, G.; Hashim, J.M.; Luvisi, D. The effects of kaolin, harpin, and imidacloprid on development of Pierce’s disease in grape. Crop Prot. 2007, 26, 92–99. [Google Scholar] [CrossRef]
- Spotti Lopes, J.R.S.; Krugner, R. Transmission ecology and epidemiology of the citrus variegated chlorosis strain of Xylella fastidiosa. In Vector-Mediated Transmission of Plant Pathogens; Lopes, J.R.S., Krugner, R., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2016; Chapter 14; pp. 195–208. [Google Scholar]
- Cornara, D.; Bosco, D.; Fereres, A. Philaenus spumarius: When an old acquaintance becomes a new threat to Europe agriculture. J. Pest Sci. 2018, 91, 957–972. [Google Scholar] [CrossRef]
- Severin, H.H.P. Spittle-insect vectors of Pierce’s disease virus: II. Life history and virus transmission. Hilgardia 1950, 19, 357–381. [Google Scholar] [CrossRef]
- Almeida, R.P.P.; Blua, M.J.; Lopes, J.R.S.; Purcell, A.H. Vector transmission of Xylella fastidiosa: Applying fundamental knowledge to generate disease management strategies. Ann. Entomol. Soc. Am. 2005, 98, 775–786. [Google Scholar] [CrossRef]
- Elbeaino, T.; Yaseen, T.; Valentini, F.; Ben Moussa, I.E.; Mazzoni, V.; D’Onghia, A.M. Identification of three potential insect vectors of Xylella fastidiosa in southern Italy. Phytopathol. Mediterr. 2014, 53, 328–332. [Google Scholar]
- Cornara, D.; Saponari, M.; Zeilinger, A.R.; de Stradis, A.; Boscia, D.; Loconsole, G.; Bosco, D.; Martelli, G.P.; Almeida, R.P.P.; Porcelli, F. Spittlebugs as vectors of Xylella fastidiosa in olive orchards in Italy. J. Pest Sci. 2017, 90, 521–530. [Google Scholar] [CrossRef]
- Janse, J.D.; Obradovic, A. Xylella fastidiosa: Its biology, diagnosis, control and risks. J. Plant Pathol. 2010, 92 (Suppl. 1), S35–S48. [Google Scholar]
- Purcell, A.H.; Finlay, A. Evidence for noncirculative transmission of Pierce’s disease bacterium by sharpshooter leafhoppers. Phytopathology 1979, 69, 393–395. [Google Scholar] [CrossRef]
- Cornara, D.; Sicard, A.; Zeilinger, A.R.; Porcelli, F. Transmission of Xylella fastidiosa to grapevine by the meadow spittlebug. Phytoparasitica 2016, 106, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Latini, A.; Foxi, C.; Borfecchia, F.; Lentini, A.; De Cecco, L.; Iantosca, D.; Serafini, M.; Laneri, U.; Citterio, M.; Campiotti, A.; et al. Tacking the vector of Xylella fastidiosa: Geo-statistical analysis of long-term field observations on host plants influencing the distribution of Philaenus spumarius nymphs. Environ. Sci. Pollut. Res. 2019, 26, 6503–6516. [Google Scholar] [CrossRef] [PubMed]
- Grandgirard, J.; Hoddle, M.S.; Petit, J.N.; Roderick, G.K.; Davies, N. Classical biological control of the glassy-winged sharpshooter, Homalodisca vitripennis, by the egg parasitoid Gonatocerus ashmeadi in the Society, Marquesas and Austral archipelagos of French Polynesia. Biol. Control 2009, 48, 155–163. [Google Scholar] [CrossRef]
- Irvin, N.A.; Pinckard, T.R.; Perring, T.M.; Hoddle, M.S. Evaluating the potential of buckwheat and cahaba vetch as nectar producing cover crops for enhancing biological control of Homalodisca vitripennis in California vineyards. Biol. Control 2014, 76, 10–18. [Google Scholar] [CrossRef]
- Morgan, D.J.W.; Triapitsyn, S.V.; Redak, R.A.; Bezark, L.G.; Hoddle, M.S. Biological control of the glassy-winged sharp shooter: Current status and future potential. In Proceedings of the California Conference on Biological Control, Riverside, CA, USA, 11–12 July 2000; Hoddle, M.S., Ed.; Center for Biological Control, College of Natural Resources, University of California: Berkeley, CA, USA, 2000; pp. 167–171. [Google Scholar]
- Harper, G.; Whittaker, J.B. The role of natural enemies in the colour polymorphism of Philaenus spumarius L. J. Anim. Ecol. 1976, 45, 91–104. [Google Scholar] [CrossRef]
- Carles-Tolrá Hjorth-Andersen, M. Catalogue of Spanish and Andorran Diptera; SEA monography 8: Zarafgoza, Spain, 2002. [Google Scholar]
- Ben-Ze´Ev, I.; Kenneth, R.G. Zoophthora radicans and Zoophthora petchi sp. nov. (Zygomycetes: Entomophthorales), two species of the ‘Sphaerosperma group’ attacking leaf-hoppers and frog-hoppers (Hom.). Entomophaga 1981, 26, 131–142. [Google Scholar] [CrossRef]
- Prabhaker, N.; Castle, S.J.; Toscano, N.C. Susceptibility of immature stages of Homalodisca coagulate (Hemiptera: Cicadellidae) to selected insecticides. J. Econ. Entomol. 2006, 99, 1805–1812. [Google Scholar] [CrossRef]
- Dongiovanni, C.; Di Carolo, M.; Fumarola, G.; Tauro, D.; Altamura, G.; Cavalieri, V. Evaluation of insecticides for the control of juveniles of Philaenus spumarius L., 2015–2017. Arthropod Manag. Tests 2018, 43, 1–2. [Google Scholar] [CrossRef]
- Dongiovanni, C.; Fumarola, G.; Di Carolo, M.; Tauro, D.; Ciniero, A.; Altamura, G.; Palmisano, F.; Silleti, M.R.; Pollastro, P.; Cavalieri, V. Sputacchina dell’olivo insetticidi a confront. L’Inf. Agrar. 2018, 24–25, 51–56. [Google Scholar]
- OJEU (Official Journal of the European Union). Commission implementing regulations (EU) 2018/783, 784 and 785 of 29 May 2018 amending Implementing regulation /EU) No 540/211 as regards the conditions of approval of the active substances imidacloprid, clothianidin and thiamethoxam. OJEU 2018, 132, 31–44. [Google Scholar]
- IRAC (Insecticide Resistance Action Committee). Insecticide Resistance Action Committee. 2019. Available online: https://www.irac-online.org (accessed on 1 February 2019).
- Morente, M.; Cornara, D.; Moreno, A.; Fereres, A. Continuous indoor rearing of Philaenus spumarius, the main European vector of Xylella fastidiosa. J. Appl. Entomol. 2018, 142, 901–904. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- MAPA (Ministry of Agriculture, Fishery and Food). CLOSER. 2019. Available online: https://www.mapa.gob.es/agricultura/pags/fitos/registro/productos/pdf/ES-00461.pdf (accessed on 1 February 2019).
- Kakko, I.; Toimela, T.; Tähti, H. Piperonyl butoxide potentiates the synaptosome ATPase inhibiting effect of pyrethrin. Chemosphere 2000, 40, 301–305. [Google Scholar] [CrossRef]
- MAPA (Ministry of Agriculture, Fishery and Food). Insecticida Vac®. 2019. Available online: https://www.mapa.gob.es/agricultura/pags/fitos/registro/productos/pdf/19884.pdf (accessed on 25 June 2019).
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; WH Freeman and Co.: New York, NY, USA, 1995; p. 887. [Google Scholar]
- MAPA (Ministry of Agriculture, Fishery and Food). List of Active Substances Included, Excluded and under Evaluation. 2019. Available online: https://www.mapa.gob.es/agricultura/pags/fitos/registro/fichas/pdf/Lista_Sustancias_activas_aceptadas_excluidas.pdf (accessed on 1 February 2019).
- Dongiovanni, C.; Cavalieri, V.; Altamura, G.; Di Carolo, M.; Fumarola, G.; Saponari, M.; Porcelli, F. Preliminary results of comparative efficacy evalutation trials against Philaenus spumarius L., vector of Xylella fastidiosa. Options Méditerr. Sér. A. Sémin. Méditerr. 2016, 121, 79–80. [Google Scholar]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorgan. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- OJEU (Official Journal of the European Union). Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009, establishing a framework for Community action to achieve the sustainable use of pesticides. OJEU 2009, 309, 71–86. [Google Scholar]
- Bürgel, K.; Daniel, C.; Wyss, E. Effects of autumn kaolin treatments on the rosy apple aphid Dysaphis plantaginea and possible mode of action. J. Appl. Entomol. 2005, 129, 311–314. [Google Scholar] [CrossRef]
- Tubajika, K.M.; Civerolo, E.L.; Purteka, G.; Wendel, L.; Ciomperlik, M.; Bartels, D.; Luvisi, D.; Hashim, J.M. Messenger and particle film Surround reduces Pierce’s disease development in grape. Phytopathology S 2003, 93, 84. [Google Scholar]
- Whittaker, J.B. Cercopid spittle as a microhabitat. Oikos 1970, 21, 59–64. [Google Scholar] [CrossRef]
- MAPA (Ministry of Agriculture, Fishery and Food). Commercials against Philaenus Spumarius in Spain. 2019. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/productos-fitosanitarios/registro/productos/forexi.asp?e=0&plagEfecto=908 (accessed on 1 February 2019).


{kind=link}
| Active Ingredient | Commercial Product | Company | IRAC | Authorized | MFRC # Commercial Product | Replicates * |
|---|---|---|---|---|---|---|
| Water-mock | - | - | - | - | - | 29 |
| 1% PBO † | Piperonyl butoxide 90% | Alfa Aesar (Lancashire, United Kingdom) | - | No | 10 mL/L | 12 |
| 3% PBO | Piperonyl butoxide 90% | Alfa Aesar (Lancashire, United Kingdom) | - | No | 30 mL/L | 12 |
| Kaolin | Surround® 95% [WP] P/P | Tessenderlo (Overpelt, Belgium) | - | Yes | 50 g/L | 12 |
| Azadirachtin | Align® 3.2% [EC] P/V | Sipcam Inagra (Valencia, Spain) | UN ‡ | Yes | 1.5 mL/L | 12 |
| Delthamethrin | Decis Evo® 2.5% [EW] P/V | Bayer (Madrid, Spain) | 3A | Yes | 0.5 mL/L | 12 |
| λ-cyhalothrin | Karate Zeon® 10% [CS] P/V | Syngenta (Madrid, Spain) | 3A | Yes | 0.2 mL/L | 12 |
| Pyrethrin | Pirecris® α 2% [EC] P/V | Seipasa (Valencia, Spain) | 3A | Yes | 1.5 mL/L | 13 |
| Pyrethrin + 1% PBO | Pirecris® 2% [EC] P/V | Seipasa (Valencia, Spain) | 3A | No | 1.5 + 10 mL/L | 12 |
| Pyrethrin + 3% PBO | Pirecris® 2% [EC] P/V | Seipasa (Valencia, Spain) | 3A | No | 1.5 + 30 mL/L | 12 |
| Sulfoxaflor | Closer® [SC] P/V | Dow Agrosciences (Madrid, Spain) | 4C | Yes | 0.24 mL/L | 12 |
| Pymetrozine | Plenum® 50% [WG] P/P | Syngenta (Madrid, Spain) | 9 | Yes | 0.5 g/L | 12 |
| Spirotetramat | Movento® 150 O-TEQ 15% [OD] P/V | Bayer (Madrid, Spain) | 23 | Yes | 0.5 mL/L | 12 |
| Treatment | Spittle † | Insects Settled on S. oleraceus (%) | Insects Walking inside the Cage (%) | Total Insects Alive inside the Cage (%) | Number of Adults # | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | ||
| Water-Mock Control | Yes | 78.8 ± 4.9 a | 73.3 ± 5.5 a | 58.8 ± 7.8 a | 8.8 ± 3.3 a | 9.6 ± 2.5 a | 18.8 ± 9.0 a | 87.5 ± 4.2 a | 82.9 ± 4.6 a | 77.5 ± 5.4 a | 6.7 ± 2.2 a | 12.3 ± 3.7 a | 25.2 ± 2.0 a |
| Sulfoxaflor 32 ppm | No | 12.5 ± 2.2 b | 8.8 ± 2.1 b | 7.5 ± 2.0 b | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 12.5 ± 2.2 b | 8.8 ± 2.1 b | 7.5 ± 2.0 b | 0.3 ± 0.6 b | 0.3 ± 0.2 b | 1.3 ± 0.5 b |
| Sulfoxaflor 64 ppm | No | 3.8 ± 1.5 c | 2.5 ± 1.6 bc | 1.7 ± 1.2 bc | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 3.8 ± 1.5 c | 2.5 ± 1.6 bc | 1.7 ± 1.2 bc | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b |
| Delthamethrin 12.5 ppm | No | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b |
| Statistics | F = 19.579 df = 3(20) p < 0.001 | F = 9.775 df = 3(20) p < 0.001 | F = 6.019 df = 3(20) p = 0.004 | F = 8.733 df = 3(20) p = 0.001 | F = 12.864 df = 3(20) p < 0.001 | F = 11.307 df = 3(20) p < 0.001 | H = 20.713 df = 3 p < 0.001 | F = 13.800 df = 3(20) p < 0.001 | H = 18.803 df = 3 p < 0.001 | H = 15.555 df = 3 p < 0.001 | H = 14.589 df = 3 p < 0.001 | H = 19.768 df = 3 p < 0.001 | |
| Active Ingredient | MFRC † Commercial Product | Plants with Spittle/Plants Tested at 72 h | Corrected Mortality (%) | ||
|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | |||
| Pyrethrin + 3% PBO | 1.50 + 30 mL/L | 0/12 = 0% | 96.0 ± 2.6 a | 96.0 ± 2.6 a | 96.0 ± 2.6 a |
| Pyrethrin + 1% PBO | 1.50 + 10 mL/L | 0/12 = 0% | 97.5 ± 1.3 a | 96.7 ± 1.4 a | 95.9 ± 1.9 a |
| λ-cyhalothrin | 0.20 mL/L | 0/12 = 0% | 96.7 ± 1.4 a | 95.0 ± 1.5 a | 94.2 ± 1.9 a |
| Delthamethrin | 0.50 mL/L | 0/12 = 0% | 86.8 ± 2.8 b | 86.8 ± 2.5 b | 83.5 ± 3.1 b |
| Sulfoxaflor | 0.24 mL/L | 7/12 = 58% | 61.7 ± 5.8 c | 76.7 ± 5.4 c | 82.5 ± 3.9 b |
| Pyrethrin | 1.50 mL/L | 10/13 = 77% | 23.9 ± 6.7 d | 24.6 ± 6.6 d | 32.3 ± 7.6 c |
| Pymetrozine | 0.50 g/L | 8/12 = 67% | 15.0 ± 3.6 d | 20.9 ± 6.0 d | 28.4 ± 5.9 c |
| 1% PBO | 10 mL/L | 10/12 = 83% | 13.3 ± 3.1 d | 20.2 ± 4.3 d | 26.7 ± 5.0 c |
| Azadirachtin | 1.5 mL/L | 6/12 = 50% | 7.5 ± 3.0 d | 17.5 ± 4.6 d | 25.0 ± 6.3 c |
| 3% PBO | 30 mL/L | 11/12 = 92% | 17.5 ± 2.5 d | 20.2 ± 0.0 d | 22.5 ± 2.5 c |
| Spirotetramat | 0.5 mL/L | 8/12 = 67% | 15.8 ± 3.6 d | 19.2 ± 3.6 d | 21.7 ± 4.9 c |
| Kaolin | 50 g/L | 8/12 = 67% | 12.5 ± 5.7 d | 16.7 ± 6.3 d | 18.3 ± 6.6 c |
| Water-mock # | - | 16/21 = 76% | 4.0 ± 1.1 | 4.5 ± 1.1 | 5.5 ± 1.2 |
| Statistics | H = 106.112 df = 11 p < 0.001 | H = 101.994 df = 11 p < 0.001 | H = 101.407 df = 11 p < 0.001 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dáder, B.; Viñuela, E.; Moreno, A.; Plaza, M.; Garzo, E.; del Estal, P.; Fereres, A. Sulfoxaflor and Natural Pyrethrin with Piperonyl Butoxide Are Effective Alternatives to Neonicotinoids against Juveniles of Philaenus spumarius, the European Vector of Xylella fastidiosa. Insects 2019, 10, 225. https://doi.org/10.3390/insects10080225
Dáder B, Viñuela E, Moreno A, Plaza M, Garzo E, del Estal P, Fereres A. Sulfoxaflor and Natural Pyrethrin with Piperonyl Butoxide Are Effective Alternatives to Neonicotinoids against Juveniles of Philaenus spumarius, the European Vector of Xylella fastidiosa. Insects. 2019; 10(8):225. https://doi.org/10.3390/insects10080225
Chicago/Turabian StyleDáder, Beatriz, Elisa Viñuela, Aránzazu Moreno, María Plaza, Elisa Garzo, Pedro del Estal, and Alberto Fereres. 2019. "Sulfoxaflor and Natural Pyrethrin with Piperonyl Butoxide Are Effective Alternatives to Neonicotinoids against Juveniles of Philaenus spumarius, the European Vector of Xylella fastidiosa" Insects 10, no. 8: 225. https://doi.org/10.3390/insects10080225