Fate of Astrocytes in The Gerbil Hippocampus After Transient Global Cerebral Ischemia

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
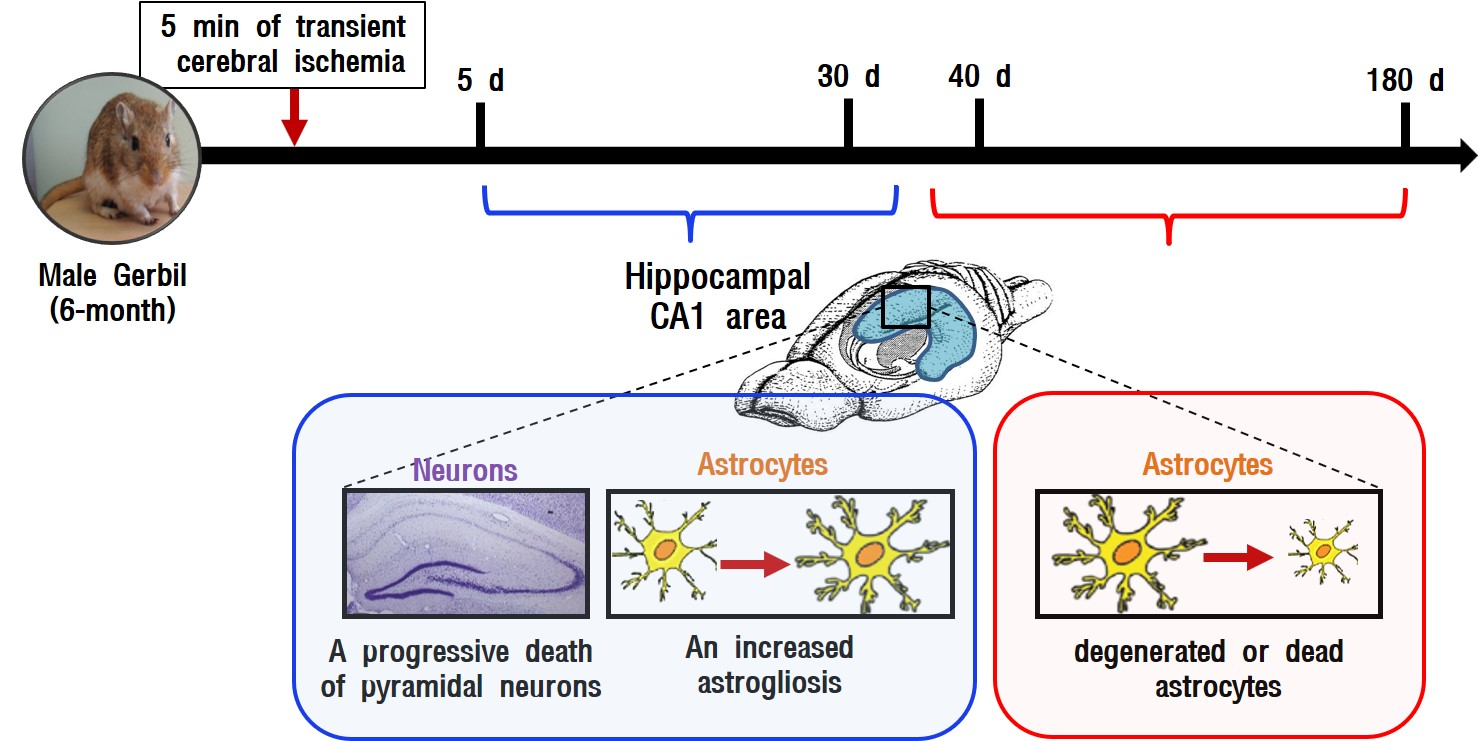
1. Introduction
2. Results
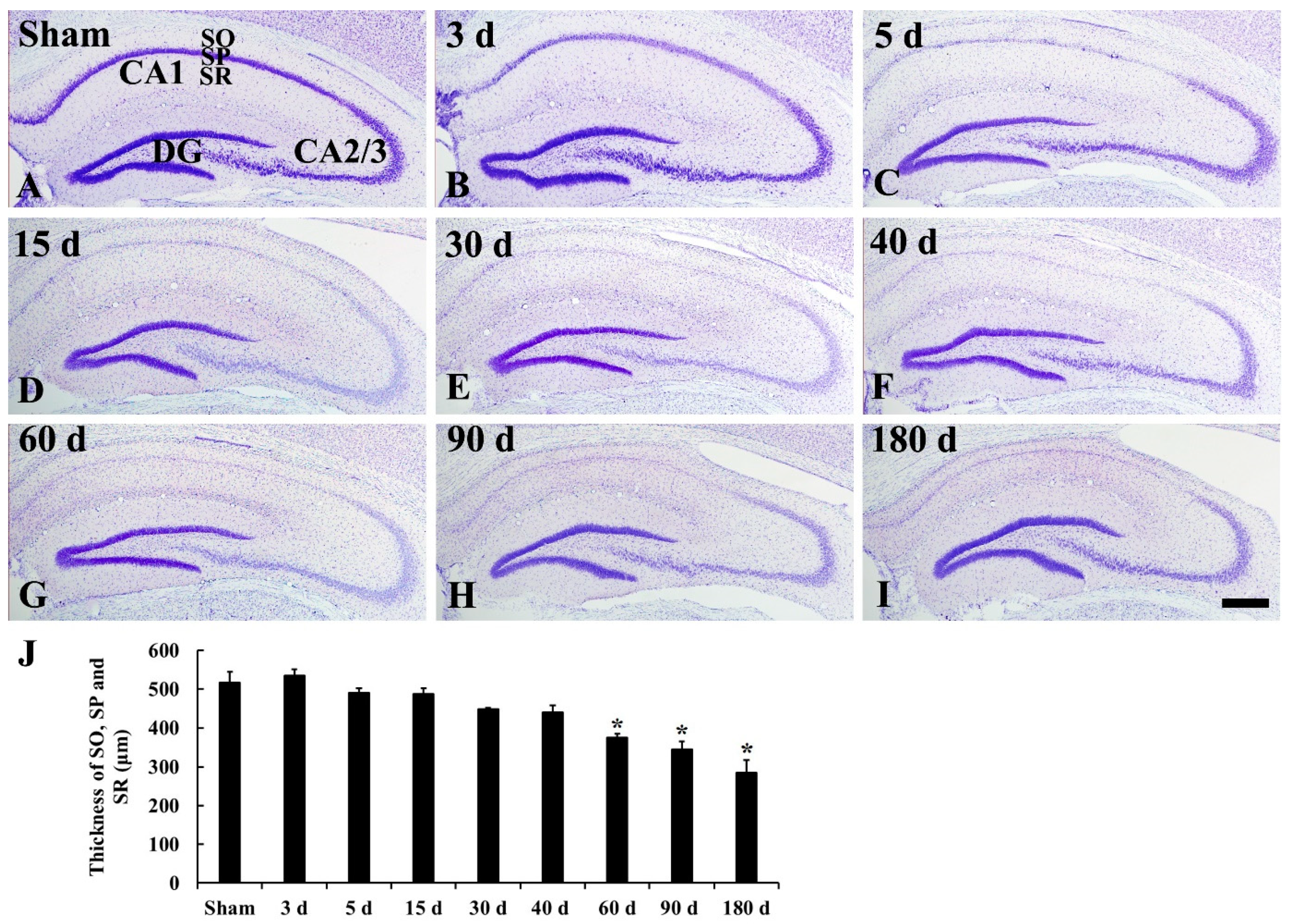
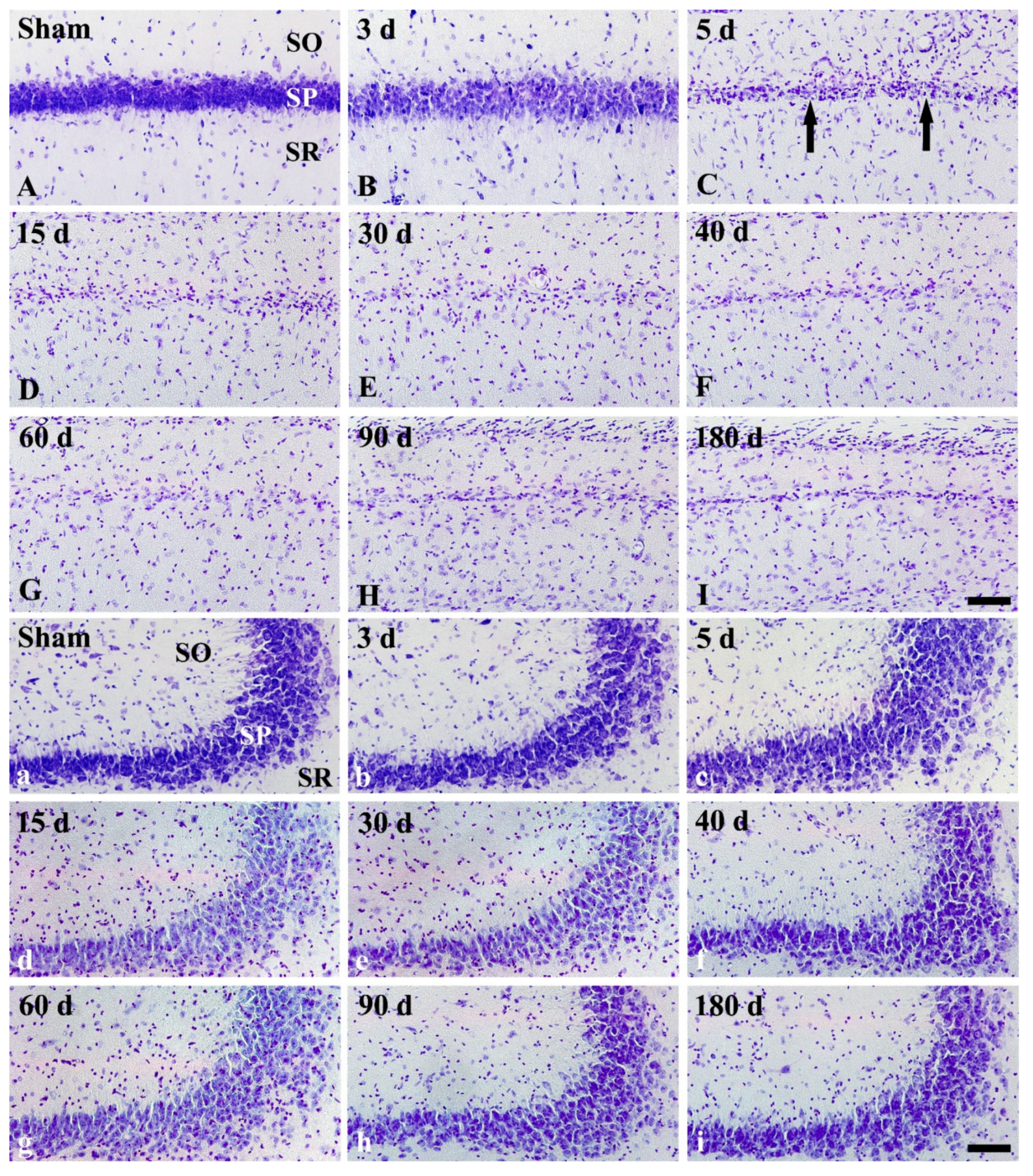
2.1. CV Staining
2.2. FJB Histofluorescence Staining
2.3. GFAP Immunoreactive Astrocytes
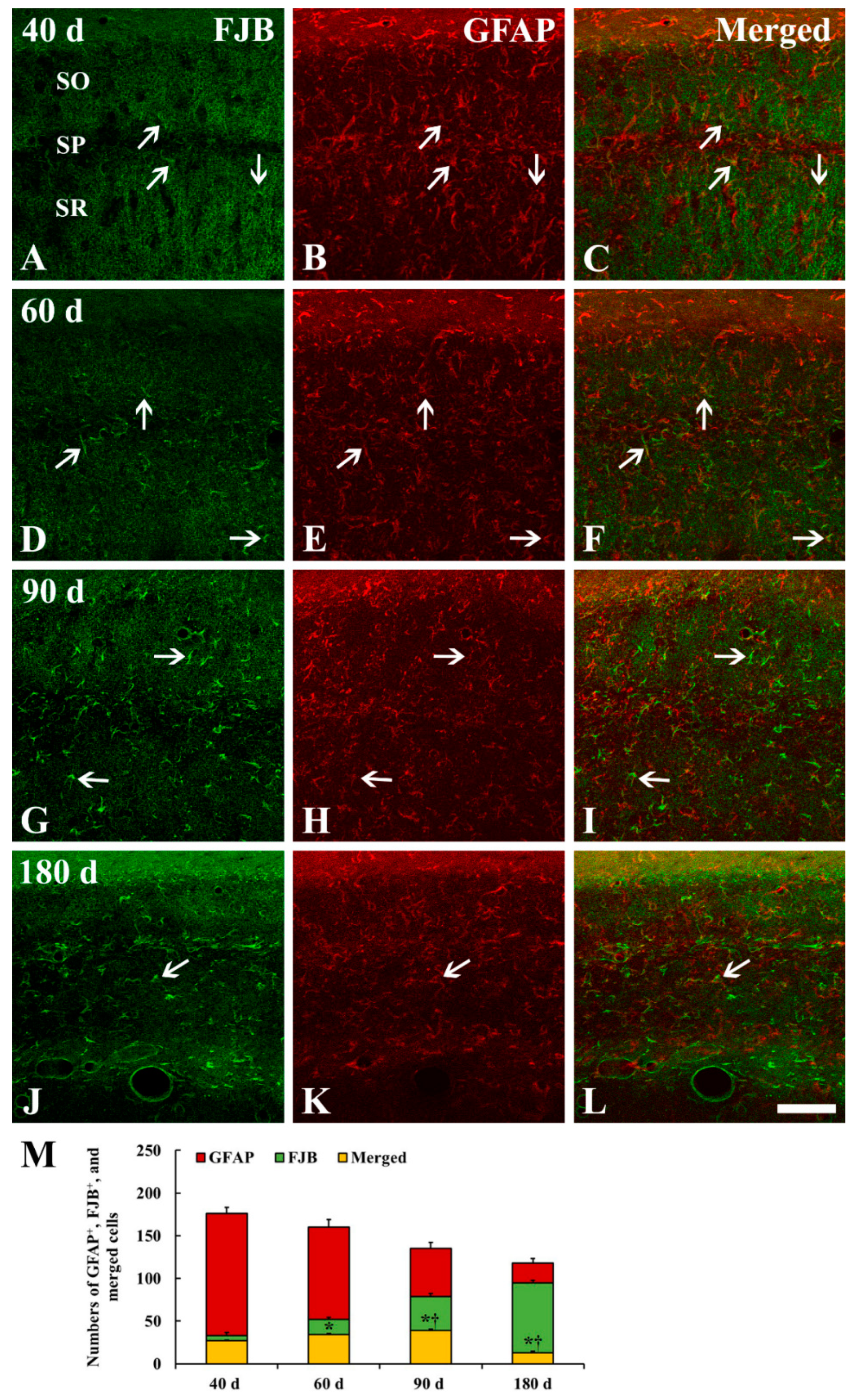
2.4. FJB+/GFAP+ Cells
3. Discussion
3.1. Ischemic Damage of CA1 Pyramidal Neurons and Shrinkage of CA1 Area
3.2. Numerical and Morphological Changes of GFAP+ Astrocytes in Ischemic Hippocampus
3.3. Degeneration/Death of GFAP+ Astrocytes in Ischemic Hippocampus
3.4. Summary
4. Materials and Methods
4.1. Experimental Animals
4.2. tgCI Induction
4.3. Tissue Preparation for Histology
4.4. CV Staining
4.5. FJB Histofluorescence Staining
4.6. Immunohistochemistry
4.7. Double Fluorescence Staining
4.8. Data Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kirino, T.; Sano, K. Selective vulnerability in the gerbil hippocampus following transient ischemia. Acta Neuropathol. 1984, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T. Delayed neuronal death in the gerbil hippocampus following ischemia. Brain Res. 1982, 239, 57–69. [Google Scholar] [CrossRef]
- Lee, C.H.; Yoo, K.-Y.; Choi, J.H.; Park, O.K.; Hwang, I.K.; Kim, S.K.; Kang, I.-J.; Kim, Y.-M.; Won, M.-H. Neuronal damage is much delayed and microgliosis is more severe in the aged hippocampus induced by transient cerebral ischemia compared to the adult hippocampus. J. Neurol. Sci. 2010, 294, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Lewén, A.; Noshita, N.; Gasche, Y.; Chan, P.H. Effects of Global Ischemia Duration on Neuronal, Astroglial, Oligodendroglial, and Microglial Reactions in the Vulnerable Hippocampal CA1 Subregion in Rats. J. Neurotrauma 2002, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Kimelberg, H.K. Functions of astrocytes and their potential as therapeutic targets. Neurotherapeutics 2010, 7, 338–353. [Google Scholar] [Green Version]
- Pekny, M.; Pekna, M. Astrocyte Reactivity and Reactive Astrogliosis: Costs and Benefits. Physiol. Rev. 2014, 94, 1077–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.-B.; Voloboueva, L.A.; Xu, L.-J.; Giffard, R.G. Selective Dysfunction of Hippocampal CA1 Astrocytes Contributes to Delayed Neuronal Damage after Transient Forebrain Ischemia. J. Neurosci. 2007, 27, 4253–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekny, M.; Wilhelmsson, U.; Pekna, M. The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 2014, 565, 30–38. [Google Scholar] [CrossRef]
- Becerra-Calixto, A.; Cardona-Gómez, G.P. The Role of Astrocytes in Neuroprotection after Brain Stroke: Potential in Cell Therapy. Front. Neurosci. 2017, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Cho, J.H.; Cho, J.-H.; Park, J.H.; Ahn, J.H.; Tae, H.-J.; Cho, G.-S.; Yan, B.C.; Hwang, I.K.; Lee, C.H.; et al. Impact of hyperthermia before and during ischemia–reperfusion on neuronal damage and gliosis in the gerbil hippocampus induced by transient cerebral ischemia. J. Neurol. Sci. 2015, 348, 101–110. [Google Scholar] [CrossRef]
- Giffard, R.G.; Swanson, R.A. Ischemia-induced programmed cell death in astrocytes. Glia 2005, 50, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jin, K.; Chen, M.; Pei, W.; Kawaguchi, K.; Greenberg, D.A.; Simon, R.P. Early Detection of DNA Strand Breaks in the Brain After Transient Focal Ischemia: Implications for the Role of DNA Damage in Apoptosis and Neuronal Cell Death. J. Neurochem. 2002, 69, 232–245. [Google Scholar] [CrossRef]
- Liu, D.; Smith, C.L.; Barone, F.C.; A Ellison, J.; Lysko, P.G.; Li, K.; A Simpson, I. Astrocytic demise precedes delayed neuronal death in focal ischemic rat brain. Mol. Brain Res. 1999, 68, 29–41. [Google Scholar] [CrossRef]
- Schmued, L.C.; Albertson, C.; Slikker, W. Fluoro-Jade: a novel fluorochrome for the sensitive and reliable histochemical localization of neuronal degeneration. Brain Res. 1997, 751, 37–46. [Google Scholar] [CrossRef]
- Yu, D.-K.; Yoo, K.-Y.; Na Shin, B.; Kim, I.H.; Park, J.H.; Lee, C.H.; Choi, J.H.; Cho, Y.-J.; Kang, I.-J.; Kim, Y.-M.; et al. Neuronal damage in hippocampal subregions induced by various durations of transient cerebral ischemia in gerbils using Fluoro-Jade B histofluorescence. Brain Res. 2012, 1437, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Colombo, J.A.; Puissant, V.I. Fluoro Jade Stains Early and Reactive Astroglia in the Primate Cerebral Cortex. J. Histochem. Cytochem. 2002, 50, 1135–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.J.; Fugaccia, I.; Scheff, S.W. Fluoro-Jade B Stains Quiescent and Reactive Astrocytes in the Rodent Spinal Cord. J. Neurotrauma 2003, 20, 1223–1231. [Google Scholar] [CrossRef]
- Grain, B.; Westerkam, W.; Harrison, A.; Nadler, J.; et al. Selective neuronal death after transient forebrain ischemia in the mongolian gerbil: A silver impregnation study. Neuroscience 1988, 27, 387–402. [Google Scholar] [CrossRef]
- Traystman, R.J. Animal Models of Focal and Global Cerebral Ischemia. ILAR J. 2003, 44, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Moon, S.M.; Yoo, K.-Y.; Choi, J.H.; Park, O.K.; Hwang, I.K.; Sohn, Y.; Moon, J.B.; Cho, J.H.; Won, M.-H.; et al. Long-term changes in neuronal degeneration and microglial activation in the hippocampal CA1 region after experimental transient cerebral ischemic damage. Brain Res. 2010, 1342, 138–149. [Google Scholar] [CrossRef]
- Mudrick, L.A.; Baimbridge, K.G. Long-term structural changes in the rat hippocampal formation following cerebral ischemia. Brain Res. 1989, 493, 179–184. [Google Scholar] [CrossRef]
- Kindy, M.S.; Bhat, A.N.; Bhat, N.R. Transient ischemia stimulates glial fibrillary acid protein and vimentin gene expression in the gerbil neocortex, striatum and hippocampus. Mol. Brain Res. 1992, 13, 199–206. [Google Scholar] [CrossRef]
- Kato, H.; Takahashi, A.; Itoyama, Y. Cell cycle protein expression in proliferating microglia and astrocytes following transient global cerebral ischemia in the rat. Brain Res. Bull. 2003, 60, 215–221. [Google Scholar] [CrossRef]
- Miyawaki, S.; Imai, H.; Hayasaka, T.; Masaki, N.; Ono, H.; Ochi, T.; Ito, A.; Nakatomi, H.; Setou, M.; Saito, N. Imaging mass spectrometry detects dynamic changes of phosphatidylcholine in rat hippocampal CA1 after transient global ischemia. Neuroscience 2016, 322, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivneva, T.A.; Tsupikov, O.M.; Pilipenko, M.N.; Vasilenko, D.A.; Skibo, G.G. Structural modifications of astrocytes in the hippocampus after experimental cerebral ischemia in gerbils. Neurophysiology 2005, 37, 359–364. [Google Scholar] [CrossRef]
- Yasuda, Y.; Tateishi, N.; Shimoda, T.; Satoh, S.; Ogitani, E.; Fujita, S. Relationship between S100β and GFAP expression in astrocytes during infarction and glial scar formation after mild transient ischemia. Brain Res. 2004, 1021, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Ding, S. Dynamic reactive astrocytes after focal ischemia. Neural Regen Res 2014, 9, 2048. [Google Scholar] [CrossRef]
- Sims, N.R.; Yew, W.P. Reactive astrogliosis in stroke: Contributions of astrocytes to recovery of neurological function. Neurochem. Int. 2017, 107, 88–103. [Google Scholar] [CrossRef]
- Li, Y.; Chopp, M.; Jiang, N.; Zaloga, C. In situ detection of DNA fragmentation after focal cerebral ischemia in mice. Mol. Brain Res. 1995, 28, 164–168. [Google Scholar] [CrossRef]
- Aoyama, K.; Burns, D.M.; Suh, S.W.; Garnier, P.; Matsumori, Y.; Shiina, H.; A Swanson, R. Acidosis Causes Endoplasmic Reticulum Stress and Caspase-12-Mediated Astrocyte Death. Br. J. Pharmacol. 2005, 25, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Damjanac, M.; Bilan, A.R.; Barrier, L.; Pontcharraud, R.; Anne, C.; Hugon, J.; Page, G. Fluoro-Jade® B staining as useful tool to identify activated microglia and astrocytes in a mouse transgenic model of Alzheimer’s disease. Brain Res. 2007, 1128, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Na Shin, B.; Park, J.H.; Kim, I.H.; Cho, J.H.; Chen, B.; Lee, T.-K.; Tae, H.-J.; Lee, J.-C.; Cho, J.H.; et al. Long-term observation of neuronal degeneration and microgliosis in the gerbil dentate gyrus after transient cerebral ischemia. J. Neurol. Sci. 2016, 363, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Candelario-Jalil, E.; Álvarez, D.; Merino, N.; Leon, O.S. Delayed treatment with nimesulide reduces measures of oxidative stress following global ischemic brain injury in gerbils. Neurosci. Res. 2003, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.-K.; Park, J.H.; Ahn, J.H.; Shin, M.C.; Cho, J.H.; Bae, E.J.; Kim, Y.-M.; Won, M.-H.; Lee, C.-H. Pretreated duloxetine protects hippocampal CA1 pyramidal neurons from ischemia-reperfusion injury through decreases of glial activation and oxidative stress. J. Neurol. Sci. 2016, 370, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Ehara, A.; Ueda, S.; et al. Application of Fluoro-Jade C in Acute and Chronic Neurodegeneration Models: Utilities and Staining Differences. Acta Histochem. Cytochem. 2009, 42, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, P.D.; Hall, G.; Kimura, T.; Ren, Y.; Bailey, R.M.; Lewis, J.; Febo, M.; Sahara, N. In vivo functional brain mapping in a conditional mouse model of human tauopathy (taup301l) reveals reduced neural activity in memory formation structures. Mol. Neurodegener. 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Radtke-Schuller, S.; Schuller, G.; Angenstein, F.; Grosser, O.S.; Goldschmidt, J.; Budinger, E. Brain atlas of the Mongolian gerbil (Meriones unguiculatus) in CT/MRI-aided stereotaxic coordinates. Brain Struct. Funct. 2016, 221, 1–272. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.J.; Chen, B.H.; Na Shin, B.; Cho, J.H.; Kim, I.H.; Park, J.H.; Lee, J.C.; Tae, H.J.; Choi, S.Y.; Kim, J.-D.; et al. Comparison of Immunoreactivities of Calbindin-D28k, Calretinin and Parvalbumin in the Striatum Between Young, Adult and Aged Mice, Rats and Gerbils. Neurochem. Res. 2015, 40, 864–872. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Park, J.H.; Shin, M.C.; Cho, J.H.; Lee, T.-K.; Kim, H.; Song, M.; Park, C.W.; Park, Y.E.; Lee, J.-C.; et al. Fate of Astrocytes in The Gerbil Hippocampus After Transient Global Cerebral Ischemia. Int. J. Mol. Sci. 2019, 20, 845. https://doi.org/10.3390/ijms20040845
Kim H, Park JH, Shin MC, Cho JH, Lee T-K, Kim H, Song M, Park CW, Park YE, Lee J-C, et al. Fate of Astrocytes in The Gerbil Hippocampus After Transient Global Cerebral Ischemia. International Journal of Molecular Sciences. 2019; 20(4):845. https://doi.org/10.3390/ijms20040845
Chicago/Turabian StyleKim, Hyeyoung, Joon Ha Park, Myoung Cheol Shin, Jun Hwi Cho, Tae-Kyeong Lee, Hyunjung Kim, Minah Song, Cheol Woo Park, Young Eun Park, Jae-Chul Lee, and et al. 2019. "Fate of Astrocytes in The Gerbil Hippocampus After Transient Global Cerebral Ischemia" International Journal of Molecular Sciences 20, no. 4: 845. https://doi.org/10.3390/ijms20040845