Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Collection and Material Preservation
2.2. Genotyping
2.3. Sequencing and Analysis of Individual Mosquito Genomes
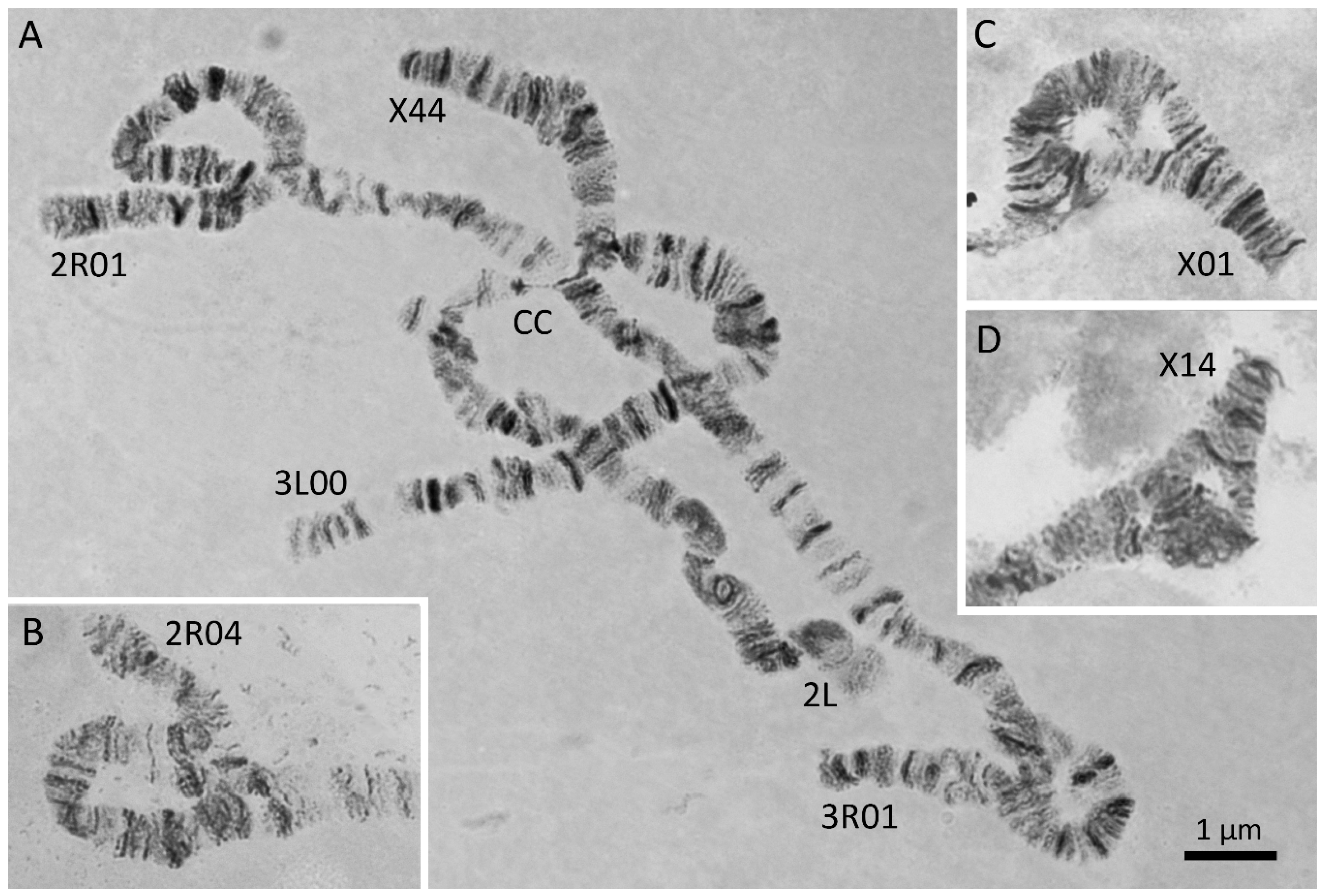
2.4. Karyotyping
2.5. Statistical Analysis of Chromosomal Inversions
3. Results
3.1. Molecular Structure of ITS2 in An. messeae and An. daciae from the Moscow Region
3.2. Species Compositions in Moscow Populations
3.3. Chromosomal Inversions in An. messeae and An. daciae
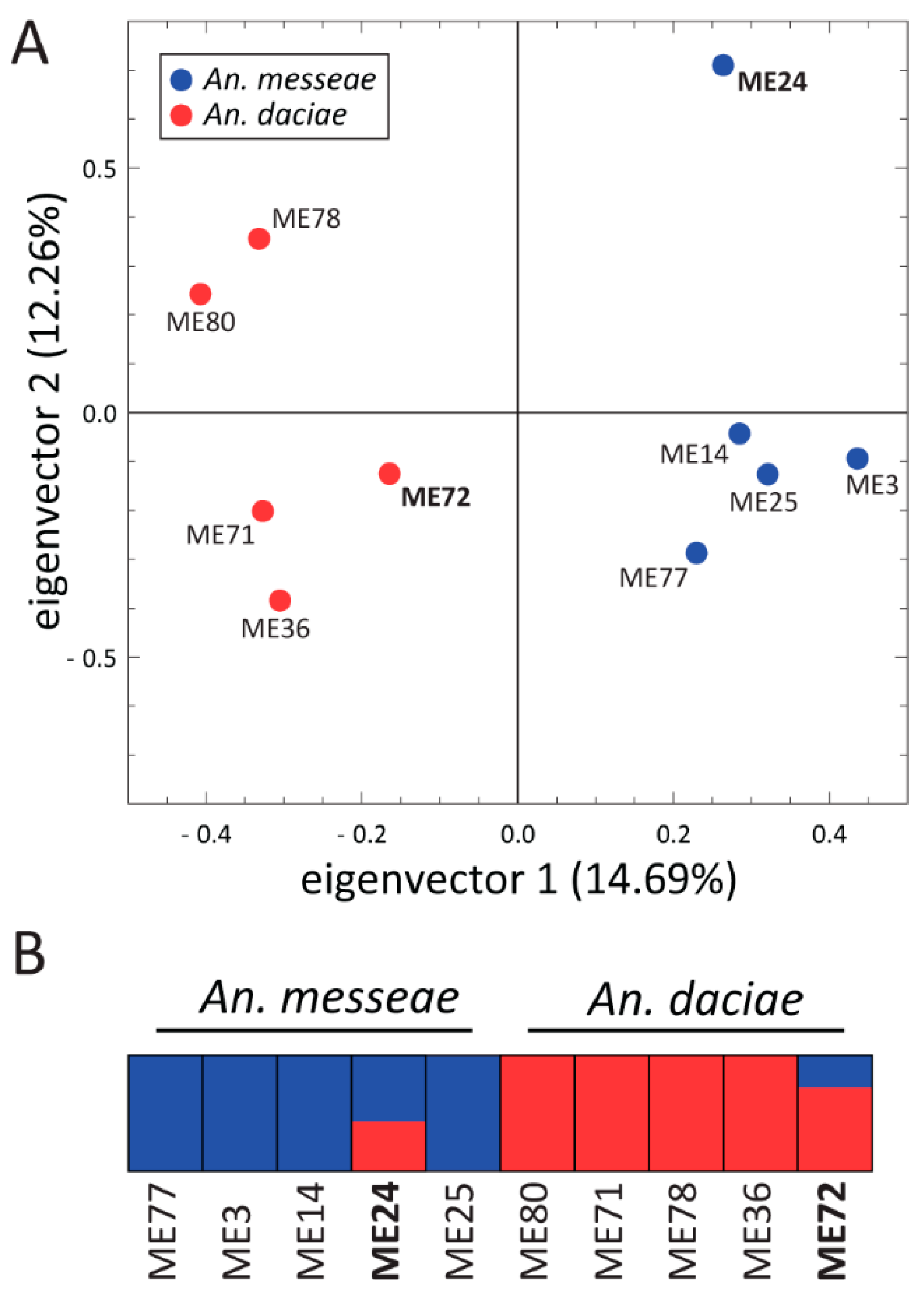
3.4. Genome Sequence Analysis of An. messeae and An. daciae
4. Discussion
4.1. Chromosomal Inversions Differentiate An. messeae and An. daciae
4.2. A Whole-Genome Analysis Determines the Genome-Wide Divergence between An. messeae and An. daciae
4.3. An. messeae and An. daciae Differ in Their Ecological Preferences and Behavior
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kirkpatrick, M.; Barton, N. Chromosome inversions, local adaptation and speciation. Genetics 2006, 173, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, F.J.; Coluzzi, M. Chromosome speciation: Humans, Drosophila, and mosquitoes. Proc. Natl. Acad. Sci. USA 2005, 102 (Suppl. 1), 6535–6542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krimbas, C.B.; Powell, J.R. Drosophila Inversion Polymorphism; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Kapun, M.; Fabian, D.K.; Goudet, J.; Flatt, T. Genomic Evidence for Adaptive Inversion Clines in Drosophila melanogaster. Mol. Biol. Evol. 2016, 33, 1317–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, Z.L.; Haynes, G.D.; Richards, S.; Schaeffer, S.W. Genomics of natural populations: Evolutionary forces that establish and maintain gene arrangements in Drosophila pseudoobscura. Mol. Ecol. 2017, 26, 6539–6562. [Google Scholar] [CrossRef] [PubMed]
- Entesarian, M.; Carlsson, B.; Mansouri, M.R.; Stattin, E.L.; Holmberg, E.; Golovleva, I.; Stefansson, H.; Klar, J.; Dahl, N. A chromosome 10 variant with a 12 Mb inversion [inv(10)(q11.22q21.1)] identical by descent and frequent in the Swedish population. Am. J. Med. Genet. Part A 2009, 149, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Drabova, J.; Trkova, M.; Hancarova, M.; Novotna, D.; Hejtmankova, M.; Havlovicova, M.; Sedlacek, Z. A 15 Mb large paracentric chromosome 21 inversion identified in Czech population through a pair of flanking duplications. Mol. Cytogenet. 2014, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Stefansson, H.; Helgason, A.; Thorleifsson, G.; Steinthorsdottir, V.; Masson, G.; Barnard, J.; Baker, A.; Jonasdottir, A.; Ingason, A.; Gudnadottir, V.G.; et al. A common inversion under selection in Europeans. Nat. Genet. 2005, 37, 129–137. [Google Scholar] [CrossRef]
- Gonzalez, J.R.; Caceres, A.; Esko, T.; Cusco, I.; Puig, M.; Esnaola, M.; Reina, J.; Siroux, V.; Bouzigon, E.; Nadif, R.; et al. A common 16p11.2 inversion underlies the joint susceptibility to asthma and obesity. Am. J. Hum. Genet. 2014, 94, 361–372. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Tsurusaki, Y.; Nakashima, M.; Miyake, N.; Saitsu, H.; Takeda, S.; Matsumoto, N. Precise detection of chromosomal translocation or inversion breakpoints by whole-genome sequencing. J. Hum. Genet. 2014, 59, 649–654. [Google Scholar] [CrossRef]
- Coluzzi, M.; Sabatini, A.; della Torre, A.; Di Deco, M.A.; Petrarca, V. A polytene chromosome analysis of the Anopheles gambiae species complex. Science 2002, 298, 1415–1418. [Google Scholar] [CrossRef]
- Gray, E.M.; Rocca, K.A.; Costantini, C.; Besansky, N.J. Inversion 2La is associated with enhanced desiccation resistance in Anopheles gambiae. Malar. J. 2009, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanzaro, G.C.; Toure, Y.T.; Carnahan, J.; Zheng, L.; Dolo, G.; Traore, S.; Petrarca, V.; Vernick, K.D.; Taylor, C.E. Complexities in the genetic structure of Anopheles gambiae populations in west Africa as revealed by microsatellite DNA analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 14260–14265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, F.; Ayala, D.; Kamdem, G.C.; Pombi, M.; Etouna, J.; Ose, K.; Fotsing, J.M.; Fontenille, D.; Besansky, N.J.; Costantini, C. Ecological niche partitioning between Anopheles gambiae molecular forms in Cameroon: The ecological side of speciation. BMC Ecol. 2009, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R. Genetic Variation in Insect Vectors: Death of Typology? Insects 2018, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Kabaria, C.W.; et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic precis. Parasites Vectors 2010, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Stegniy, V.N. Population Genetics and Evolution of Malaria Mosquitoes; Tomsk State University Publisher: Tomsk, Russia, 1991; p. 137. [Google Scholar]
- Gornostaeva, R.M.; Danilov, A.V. On distribution of malaria mosquitoes (Diptera, Culicidae: Anopheles) from Maculipennis Complex in Russian territory. Parsitologia 2002, 26, 33–47. [Google Scholar]
- Sokolova, M.I.; Snow, K.R. Malaria vectors in European Russia. European Mosquito Bulletin. J. Eur. Mosq. Control Assoc. 2002, 12, 1–6. [Google Scholar]
- Novikov, Y.M.; Vaulin, O.V. Expansion of Anopheles maculipennis s.s. (Diptera: Culicidae) to northeastern Europe and northwestern Asia: Causes and consequences. Parasites Vectors 2014, 7, 389. [Google Scholar] [CrossRef] [Green Version]
- Githeko, A.K.; Lindsay, S.W.; Confalonieri, U.E.; Patz, J.A. Climate change and vector-borne diseases: A regional analysis. Bull. World Health Organ. 2000, 78, 1136–1147. [Google Scholar]
- Falleroni, D. Fauna anofelicaitaliana e suo ‘habitat’ (paludi, risaie, canali). Metodi di lottacontro la malaria. Riv. Malariol. 1926, 5, 553–593. [Google Scholar]
- Harbach, R.E. The classification of genus Anopheles (Diptera: Culicidae): A working hypothesis of phylogenetic relationships. Bull. Entomol. Res. 2004, 94, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Hackett, L.W.; Missiroli, A. The varieties of Anopheles maculipennis and their relation to the distribution of malaria in Europe. Riv. Malariol. 1935, 14, 45–109. [Google Scholar]
- Gutsevich, A.V.; Monchadskii, A.S.; Shtakelberg, A.A. Fauna of the USSR. Diptera. Mosquitoes; Zoological Institute, USSR Academy of Science: Leningard, Russia, 1970. [Google Scholar]
- Stegnii, V.N.; Pestryakova, T.S.; Kabanova, V.M. Cytological identification of sibling species of the malaria mosquitoes Anopheles maculipennis and An. messeae. Zool. Zhurnal 1973, 52, 1971–1976. [Google Scholar]
- Stegnii, V.N. Revealing of chromosme races in the malaria mosquito Anopheles sacharovi (Diptera, Culicidae). Tsitologia 1976, 18, 1039–1041. [Google Scholar]
- Stegnii, V.N.; Kabanova, V.M. Cytoecological study of natural populations of malaria mosquitoes on the USSR territory. 1. Isolation of a new species of Anopheles in Maculipennis complex by the cytodiagnostic method. Med. Parazitol. 1976, 45, 192–198. [Google Scholar]
- Stegniy, V.N.; Kabanova, V.M. Cytoecological study of indigenous populanions of the malaria mosquito in the territory of the USSR. 1. Identification new species of Anopheles in the Maculipennis complex by the cytodiagnostic method. Mosq. Syst. 1978, 10, 1–12. [Google Scholar]
- Gordeev, M.I.; Zvantsov, A.B.; Goriacheva, I.I.; Shaikevich, E.V.; Ezhov, M.N. Description of the new species Anopheles artemievi sp.n. (Diptera, Culicidae). Med. Parazitol. 2005, 2005, 4–5. [Google Scholar]
- Sedaghat, M.M.; Linton, Y.M.; Oshaghi, M.A.; Vatandoost, H.; Harbach, R.E. The Anopheles maculipennis complex (Diptera: Culicidae) in Iran: Molecular characterization and recognition of a new species. Bull. Entomol. Res. 2003, 93, 527–535. [Google Scholar] [CrossRef]
- Nicolescu, G.; Linton, Y.M.; Vladimirescu, A.; Howard, T.M.; Harbach, R.E. Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull. Entomol. Res. 2004, 94, 525–535. [Google Scholar] [CrossRef]
- Weitzel, T.; Gauch, C.; Becker, N. Identification of Anopheles daciae in Germany through ITS2 sequencing. Parasitol. Res. 2012, 111, 2431–2438. [Google Scholar] [CrossRef]
- Kronefeld, M.; Werner, D.; Kampen, H. PCR identification and distribution of Anopheles daciae (Diptera, Culicidae) in Germany. Parasitol. Res. 2014, 113, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H.; Schafer, M.; Zielke, D.E.; Walther, D. The Anopheles maculipennis complex (Diptera: Culicidae) in Germany: An update following recent monitoring activities. Parasitol. Res. 2016, 115, 3281–3294. [Google Scholar] [CrossRef]
- Danabalan, R.; Monaghan, M.T.; Ponsonby, D.J.; Linton, Y.M. Occurrence and host preferences of Anopheles maculipennis group mosquitoes in England and Wales. Med. Vet. Entomol. 2014, 28, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Rydzanicz, K.; Czulowska, A.; Manz, C.; Jawien, P. First record of Anopheles daciae (Linton, Nicolescu & Harbach, 2004) in Poland. J. Vector Ecol. 2017, 42, 196–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazejova, H.; Sebesta, O.; Rettich, F.; Mendel, J.; Cabanova, V.; Miterpakova, M.; Betasova, L.; Pesko, J.; Hubalek, Z.; Kampen, H.; et al. Cryptic species Anopheles daciae (Diptera: Culicidae) found in the Czech Republic and Slovakia. Parasitol. Res. 2018, 117, 315–321. [Google Scholar] [CrossRef]
- Kavran, M.; Zgomba, M.; Weitzel, T.; Petric, D.; Manz, C.; Becker, N. Distribution of Anopheles daciae and other Anopheles maculipennis complex species in Serbia. Parasitol. Res. 2018, 117, 3277–3287. [Google Scholar] [CrossRef] [Green Version]
- Stegnii, V.N.; Kabanova, V.M.; Novikov Iu, M. Study of the karyotype of the malaria mosquito. Tsitologiia 1976, 18, 760–766. [Google Scholar]
- Stegnii, V.N.; Kabanova, V.M.; Novikov, Y.M.; Pleshkova, G.N. Inversion polymorphism in malaria mosquito Anopheles messeae. I. Distribution of the inversions in the species areal. Genetika 1976, 12, 47–55. [Google Scholar]
- Gordeev, M.I.; Sibataev, A.K. Cytogenetic and phenotypic variation in central and peripheral populations of the malaria mosquito, Anopheles messeae Fall. (Diptera, Culicidae). Genetika 1996, 32, 1199–1205. [Google Scholar]
- Gordeev, M.I.; Bezzhonova, O.V.; Moskaev, A.V. Chromosomal polymorphism in the populations of malaria mosquito Anopheles messeae (Diptera, Culicidae) at the south of Russian Plain. Genetika 2012, 48, 1124–1128. [Google Scholar]
- Perevozkin, V.P.; Gordeev, M.I.; Bondarchuk, S.S. Chromosome polymorphism and regularities of the subpopulation organization of malaria mosquitoes Anopheles (Diptera, Culicidae) in biotopes of the Tomsk oblast. Genetika 2009, 45, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, V.M.; Stegnii, V.N.; Luzhkova, A.G. Seasonal dynamics of inversion polymorphism in a natural population of the malarial mosquito Anopheles messeae (Diptera: Culicidae). Genetika 1973, 9, 78–82. [Google Scholar] [PubMed]
- Stegnii, V.N. Inversion polymorphism of the malarial mosquito Anopheles messeae. IV. The stability of the frequency distribution of the inversions by species area. Genetika 1983, 19, 466–473. [Google Scholar] [PubMed]
- Stegniy, V.N.; Pishchelko, A.O.; Sibataev, A.K.; Abylkassymova, G. Spatial and temporal variations of the chromosomal inversion frequencies across the range of malaria mosquito Anopheles messeae Fall. (Culicidae) during the 40-year monitoring period. Genetika 2016, 52, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Novikov Iu, M.; Kabanova, V.M. Adaptive association of inversions in a natural population of the malaria mosquito Anopheles messeae Fall. Genetika 1979, 15, 1033–1045. [Google Scholar]
- Gordeev, M.I.; Stegnii, V.N. Inversion polymorphism in the malaria mosquito Anopheles messeae. VII. Fertility and the population genetics structure of the species. Genetika 1987, 23, 2169–2174. [Google Scholar]
- Stegnii, V.N. Inversion polymorphism of the malarial mosquito Anopheles messeae. V. The interaction of different chromosomal inversions in the spatial area. Genetika 1983, 19, 474–482. [Google Scholar]
- Gordeev, M.I.; Troshkov, N. Inversion polymorphism of the malaria mosquito Anopheles messeae. IX. Cannibalism in larvae as a selection factor. Genetika 1990, 26, 1597–1603. [Google Scholar]
- Burlak, V.A.; Gordeev, M.I. The effect of infection by the entomopathogenic bacterium Bacillus thuringiensis on the spread of microsporidia in an inversion-polymorphic population of the malarial mosquito Anopheles messeae (Diptera: Culicidae). Parazitologiia 1998, 32, 264–267. [Google Scholar]
- Gordeev, M.I.; Perevozkin, V.P. Strategies for selection and stability to asphyxia in larvae of the malaria mosquito Anopheles messeae with various karyotypes. Genetika 1995, 31, 180–184. [Google Scholar]
- Gordeev, M.I.; Burlak, V.A. Inversion polymorphism in malaria mosquito Anopheles messeae. Part X. Resistance of larvae with different genotypes to toxins of crystal-forming bacteria Bacillus thuringiensis subsp. israelensis (serovar H14). Genetika 1991, 27, 238–246. [Google Scholar] [PubMed]
- Gordeev, M.I.; Burlak, V.A. Inversion polymorphism in the malaria mosquito Anopheles messeae. XI. The group effect of larval infection with Bacillus thuringiensis subsp. israeliensis bacteria. Genetika 1992, 28, 82–88. [Google Scholar] [PubMed]
- Novikov Iu, M.; Shevchenko, A.I. Inversion polymorphism and the divergence of two cryptic forms of Anopheles messeae (Diptera, Culicidae) at the level of genomic DNA repeats. Genetika 2001, 37, 915–925. [Google Scholar]
- Vaulin, O.V.; Novikov, Y.M. Geographic variability of ITS2 rDNA and COI mtDNA and cryptic species of malaria mosquito Anopheles messeae Fall. (Diptera: Culicidae). Vestn. VOGIS 2010, 14, 546–555. [Google Scholar]
- Vaulin, O.V.; Karagodin, D.A.; Zakharov, I.K.; Baricheva, E.M. The dynamics of the species composition of malaria mosquitoes in Siberian populations, detected using restriction analysis. Genetika 2018, 54, 832–842. [Google Scholar]
- OpenStreetMap. Available online: https://www.openstreetmap.org/ (accessed on 15 April 2019).
- Proft, J.; Maier, W.A.; Kampen, H. Identification of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol. Res. 1999, 85, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tools for High Throughput Sequnce Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 April 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Neafsey, D.E.; Waterhouse, R.M.; Abai, M.R.; Aganezov, S.S.; Alekseyev, M.A.; Allen, J.E.; Amon, J.; Arca, B.; Arensburger, P.; Artemov, G.; et al. Mosquito genomics. Highly evolvable malaria vectors: The genomes of 16 Anopheles mosquitoes. Science 2015, 347, 1258522. [Google Scholar] [CrossRef] [Green Version]
- Artemov, G.N.; Bondarenko, S.M.; Naumenko, A.N.; Stegniy, V.N.; Sharakhova, M.V.; Sharakhov, I.V. Partial-arm translocations in evolution of malaria mosquitoes revealed by high-coverage physical mapping of the Anopheles atroparvus genome. BMC Genom. 2018, 19, 278. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome, S. Project Data Processing, The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Tamazian, G.; Dobrynin, P.; Krasheninnikova, K.; Komissarov, A.; Koepfli, K.P.; O’Brien, S.J. Chromosomer: A reference-based genome arrangement tool for producing draft chromosome sequences. Gigascience 2016, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Artemov, G.N.; Sharakhova, M.V.; Naumenko, A.N.; Karagodin, D.A.; Baricheva, E.M.; Stegniy, V.N.; Sharakhov, I.V. A standard photomap of ovarian nurse cell chromosomes in the European malaria vector Anopheles atroparvus. Med. Vet. Entomol. 2015, 29, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [Green Version]
- Kabanova, V.M.; Kartashova, N.N.; Stegnii, V.N. Karyological study of natural populations of malarial mosquitoes in the Middle Ob river. I. Characteristics of the karyotype of Anopheles maculipennis messeae. Tsitologiia 1972, 14, 630–636. [Google Scholar]
- Meirmans, P.G.; Van Tienderen, P.H. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Michalakis, Y.; Excoffier, L. A generic estimation of population subdivision using distances between alleles with special reference for microsatellite loci. Genetics 1996, 142, 1061–1064. [Google Scholar] [PubMed]
- Stegniy, V.N.; Novikov Iu, M.; Kabanova, V.M. Cytogenetic analysis and distribution of Anopheles beklemishevi. Zool. J. 1978, 57, 873–876. [Google Scholar]
- Tanygina, E.; Gordeev, M.I.; Moskaev, A.V.; Ganushkina, L.A. The species and karyotype composition of malaria mosquito larvae in different water reservoirs of the city of Moscow. Med. Parazitol. 2014, 29–33. [Google Scholar]
- Gordeev, M.I.; Moskaev, A.V. Chromosomal polymorphism in the populations of malaria mosquito Anopheles messeae (Diptera, Culicidae) in the Volga region. Genetika 2016, 52, 685–690. [Google Scholar] [CrossRef]
- Kamali, M.; Xia, A.; Tu, Z.; Sharakhov, I.V. A New Chromosomal Phylogeny Supports the Repeated Origin of Vectorial Capacity in Malaria Mosquitoes of the Anopheles gambiae Complex. PLoS Pathog. 2012, 8, e1002960. [Google Scholar] [CrossRef]
- Toure, Y.T. The current state of studies of malaria vectors and the antivectorial campaign in west Africa. Trans. R. Soc. Trop. Med. Hyg. 1989, 83 (Suppl. 39–41). [Google Scholar]
- Toure, Y.T.; Petrarca, V.; Traore, S.F.; Coulibaly, A.; Maiga, H.M.; Sankare, O.; Sow, M.; Di Deco, M.A.; Coluzzi, M. Ecological genetic studies in the chromosomal form Mopti of Anopheles gambiae s.str. in Mali, west Africa. Genetica 1994, 94, 213–223. [Google Scholar] [CrossRef]
- Coetzee, M.; Hunt, R.H.; Wilkerson, R.; Della Torre, A.; Coulibaly, M.B.; Besansky, N.J. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae complex. Zootaxa 2013, 3619, 246–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- della Torre, A.; Tu, Z.; Petrarca, V. On the distribution and genetic differentiation of Anopheles gambiae s.s. molecular forms. Insect Biochem. Mol. Biol. 2005, 35, 755–769. [Google Scholar] [CrossRef] [PubMed]
- Wondji, C.; Frederic, S.; Petrarca, V.; Etang, J.; Santolamazza, F.; Della Torre, A.; Fontenille, D. Species and populations of the Anopheles gambiae complex in Cameroon with special emphasis on chromosomal and molecular forms of Anopheles gambiae s.s. J. Med. Entomol. 2005, 42, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- della Torre, A.; Costantini, C.; Besansky, N.J.; Caccone, A.; Petrarca, V.; Powell, J.R.; Coluzzi, M. Speciation within Anopheles gambiae—the glass is half full. Science 2002, 298, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Favia, G.; della Torre, A.; Bagayoko, M.; Lanfrancotti, A.; Sagnon, N.; Toure, Y.T.; Coluzzi, M. Molecular identification of sympatric chromosomal forms of Anopheles gambiae and further evidence of their reproductive isolation. Insect Mol. Biol. 1997, 6, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.L.; Hahn, M.W.; Nuzhdin, S.V. Genomic islands of speciation in Anopheles gambiae. PLoS Biol. 2005, 3, e285. [Google Scholar] [CrossRef] [PubMed]
- Sharakhova, M.V.; George, P.; Brusentsova, I.V.; Leman, S.C.; Bailey, J.A.; Smith, C.D.; Sharakhov, I.V. Genome mapping and characterization of the Anopheles gambiae heterochromatin. BMC Genom. 2010, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Lawniczak, M.K.; Emrich, S.J.; Holloway, A.K.; Regier, A.P.; Olson, M.; White, B.; Redmond, S.; Fulton, L.; Appelbaum, E.; Godfrey, J.; et al. Widespread divergence between incipient Anopheles gambiae species revealed by whole genome sequences. Science 2010, 330, 512–514. [Google Scholar] [CrossRef] [Green Version]
- Presgraves, D.C. Evaluating genomic signatures of “the large X-effect” during complex speciation. Mol. Ecol. 2018, 27, 3822–3830. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates: Sunderland, MA, USA, 2004; p. xiii. 545p. [Google Scholar]
- Turelli, M.; Orr, H.A. The dominance theory of Haldane’s rule. Genetics 1995, 140, 389–402. [Google Scholar]
- Charlesworth, B.; Coyne, J.A.; Barton, N.H. The relative rates of evolution of sex chromosomes and autosomes. Am. Nat. 1987, 130, 113–146. [Google Scholar] [CrossRef]
- Hurst, L.D.; Pomiankowski, A. Causes of sex ratio bias may account for unisexual sterility in hybrids: A new explanation of Haldane’s rule and related phenomena. Genetics 1991, 128, 841–858. [Google Scholar]
- Geiger, R. Classification of Climates after W. Koppen; Springer: Berlin/Heidelberg, Germany, 1954; Volume 3. [Google Scholar]
- Luhken, R.; Czajka, C.; Steinke, S.; Jost, H.; Schmidt-Chanasit, J.; Pfitzner, W.; Becker, N.; Kiel, E.; Kruger, A.; Tannich, E. Distribution of individual members of the mosquito Anopheles maculipennis complex in Germany identified by newly developed real-time PCR assays. Med. Vet. Entomol. 2016, 30, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Kronefeld, M.; Dittmann, M.; Zielke, D.; Werner, D.; Kampen, H. Molecular confirmation of the occurrence in Germany of Anopheles daciae (Diptera, Culicidae). Parasites Vectors 2012, 5, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 15 April 2019).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genotype | Population | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 150 | 211 | 215 | 217 | 412 | 432 | Novokosino | Noginsk | Yegoryevsk | |
| An. daciae | C | W(A+T) | W(A+T) | Y(C+T) | A | C | 24 | 31 | 60 |
| C | W | W | T | A | C | 2 | 0 | 0 | |
| M(A+C) | W | W | Y | A | C | 0 | 0 | 2 | |
| An. messeae | M(A+C) | T | T | C | G | G | 73 | 63 | 24 |
| Hybrid | M | W | W | Y | R(A+G) | S(C+G) | 0 | 0 | 1 |
| Total | 99 | 94 | 87 | ||||||
| Population, Species | X | 2R | 3R | 3L |
|---|---|---|---|---|
| Novokosino, An. messeae | 0.817 | 0.071 | 0.433 | 0.958 |
| Novokosino, An. daciae | 0.046 | Monomorphic | 0.878 | Monomorphic |
| Noginsk, An. messeae | 0.051 | 0.111 | 0.302 | 0.838 |
| Noginsk, An. daciae | 0.002 * | 1.000 | 0.242 | Monomorphic |
| Yegoryevsk, An. messeae | 0.022 * | 0.595 | 0.601 | 0.791 |
| Yegoryevsk, An. daciae | 0.001 * | 0.992 | 0.922 | Monomorphic |
| Population Species | Novokosino An. messeae | Novokosino An. daciae | Noginsk An. messeae | Noginsk An. daciae | Yegoryevsk An. messeae | Yegoryevsk An. daciae |
|---|---|---|---|---|---|---|
| Novokosino An. messeae | -- | 0.159 * | 0.003 | 0.143 * | 0.021 | 0.218 * |
| Novokosino An. daciae | 0.159 * | -- | 0.211 * | −0.017 | 0.095 * | 0.000 |
| Noginsk An. messeae | 0.003 | 0.211 * | -- | 0.197 * | 0.046 | 0.280 * |
| Noginsk An. daciae | 0.143 * | −0.017 | 0.197 * | -- | 0.076 | 0.010 |
| Yegoryevsk An. messeae | 0.021 | 0.095 * | 0.046 | 0.076 | -- | 0.164 * |
| Yegoryevsk An. daciae | 0.218 * | 0.000 | 0.280 * | 0.010 | 0.164 * | -- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naumenko, A.N.; Karagodin, D.A.; Yurchenko, A.A.; Moskaev, A.V.; Martin, O.I.; Baricheva, E.M.; Sharakhov, I.V.; Gordeev, M.I.; Sharakhova, M.V. Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae. Genes 2020, 11, 165. https://doi.org/10.3390/genes11020165
Naumenko AN, Karagodin DA, Yurchenko AA, Moskaev AV, Martin OI, Baricheva EM, Sharakhov IV, Gordeev MI, Sharakhova MV. Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae. Genes. 2020; 11(2):165. https://doi.org/10.3390/genes11020165
Chicago/Turabian StyleNaumenko, Anastasia N., Dmitriy A. Karagodin, Andrey A. Yurchenko, Anton V. Moskaev, Olga I. Martin, Elina M. Baricheva, Igor V. Sharakhov, Mikhail I. Gordeev, and Maria V. Sharakhova. 2020. "Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae" Genes 11, no. 2: 165. https://doi.org/10.3390/genes11020165