
The View of Pediatric Nephrotic Syndrome as a Podocytopathy
by
 ,
,
Pedro Alves Soares Vaz de Castro
*,
Thomas Fujihara Ide
,
Fernando Crespo Torres
and
Ana Cristina Simões e Silva
* Interdisciplinary Laboratory of Medical Investigation, Unit of Pediatric Nephrology, Faculty of Medicine, Federal University of Minas Gerais (UFMG), Belo Horizonte 30130-100, Brazil
*
Authors to whom correspondence should be addressed.
Kidney Dial. 2023, 3(4), 346-372; https://doi.org/10.3390/kidneydial3040030
Submission received: 30 September 2023
/
Revised: 16 November 2023
/
Accepted: 4 December 2023
/
Published: 12 December 2023
Abstract
:Nephrotic syndrome (NS) is a complex clinical entity characterized by proteinuria, hypoalbuminemia, and edema. In this review, we propose the view of NS as a podocytopathy, highlighting the importance of understanding the role of podocytes in the development of this condition. We discuss the various etiologies of NS, ranging from congenital to primary renal diseases, as well as secondary forms due to systemic diseases. We also delve into the mechanisms underlying podocyte injury, which plays a crucial role in the development of NS. By viewing NS as a podocytopathy, we suggest potential implications for the diagnosis and treatment of this condition, including the use of podocyte-specific biomarkers and targeted therapies. Our review provides a comprehensive overview of NS and its underlying mechanisms, emphasizing the importance of a multidisciplinary approach to the diagnosis and management of this condition. Further research is essential to better understand the complex interplay between podocyte injury and the development of NS, with the ultimate goal of improving patient outcomes.
1. Introduction
Nephrotic syndrome (NS) is a clinical entity characterized by the triad of proteinuria, hypoalbuminemia, and edema [1]. It is a group of diseases composed of numerous etiologies, ranging from congenital, such as Finnish Type Nephropathy and Diffuse Mesangial Sclerosis, to primary renal diseases, including Minimal Change Nephrotic Syndrome (MCNS) and Focal Segmental Glomerulosclerosis (FSGS), or secondary forms due to systemic diseases such as, for instance, diabetes mellitus and lupus erythematosus [2]. The reported incidence of childhood NS is 2–7 per 100,000 children, with a prevalence of about 16 cases per 100,000 and variability among ethnic groups. The peak age of onset of childhood NS occurs at 2–3 years in most cases [3]. MCNS is the most common histological variant, accounting for approximately 80% of NS cases in children, followed by FSGS, which, although less common, is more related to poor long-term outcomes [4,5].
The main pathophysiological alteration in NS is the impairment of the glomerular filtration barrier. Therefore, its permeability becomes non-selective, and urinary protein loss occurs. All components of the glomerular barrier may interfere with the permeability of molecules. These components include the fenestrated endothelium, the glomerular basement membrane (GBM), and the podocytes. In NS patients, the effacement of the foot processes of the podocytes is a common finding in the electronic microscopy of the kidney tissue [6]. Podocytes are highly differentiated epithelial cells with a large cell body and elongated cellular extensions, the foot processes, that interdigitate along the outer wall of the glomerular capillary and, among other functions, provide structural support and control the filtration process [7].
Being the final barrier to protein loss in the glomerulus, alterations in podocyte structure can explain why injuries in these cells, such as the effacement of their foot processes, are typically related to proteinuria and hypoalbuminemia and, therefore, to the conditions under the NS umbrella [6]. In this context, this review aims to summarize the current understanding of NS as a podocytopathy and to discuss the main molecular, genetic, and histological markers and their influence on the onset and outcome of the disease.
2. Pathophysiological Mechanisms of the Podocytopathies
2.1. Pathophysiology of Nephrotic Syndrome and the Glomerular Filtration
Despite often occurring rapidly, NS can have an insidious presentation, with proteinuria before the onset of symptoms. The proteinuria indicates an altered permeability of the glomerular filtration barrier [8] and contributes to edema and hypoalbuminemia. Hypoalbuminemia induces lipoprotein synthesis, which causes hyperlipidemia [2,3]. The three components of the glomerular filtration barrier—the fenestrated endothelium, the GBM, and the podocytes—control the size of filtered molecules [9]. The podocytes are visceral glomerular epithelial cells, and their injury plays an important role in the development of the NS. Therefore, the main pathophysiological alterations in NS justify the view of this syndrome as a podocytopathy [2].
Podocytes are highly specialized and polarized epithelial cells related to the synthesis of extracellular matrix components of the GBM. Furthermore, these cells have a structural role in supporting the glomerular capillaries with their interdigitating foot processes (FP), which are linked to each other through a special cell–cell junction, the slit diaphragm (SD). The podocyte also synthesizes many proteins from the SD. The unique zipper-like architecture of the SD is essential to the glomerulus permeability; therefore, podocyte depletion due to detachment, apoptosis, or necrosis is a critical step in the development of NS [7].
Electron microscopy shows the effacement of the foot processes as a major factor related to the glomerular lesion [7]. The podocytes and the SD zipper-like array are composed of an extensive group of molecules, including an actin cytoskeleton and proteins such as ZO-1, nephrin, podocin, and CD2AP, in both the podocyte itself and the GBM. Some etiologies of congenital and steroid-resistant NS can be linked to mutations in genes encoding some of these components, altering the glomerular filtration barrier and leading to intense urinary protein loss [1].
The mechanisms that underlie FP effacement and podocytes’ response to injury are important to determine the clinical manifestations of NS and prognosis [6]. Furthermore, the understanding of podocyte-specific protein alterations and the dynamic changes of glomerular permeability and selectivity are also relevant for the management and treatment of NS [2,3].
2.2. Podocyte Foot Processes: Update on Molecular Anatomy and Effacement
Each podocyte FP contains a contractile system composed of actin, myosin-II, actinin-4, talin, and vinculin [10]. The cytoskeleton molecular composition in podocyte FP is critical to podocyte function and effacement processes. The foot processes (FPs) are anchored to the GBM because of 31 integrin complexes, which, consisting of heterodimeric transmembrane receptors, mediate cell attachment to the extracellular matrix and are crucial for cell signaling. The adjacent FPs of the same podocyte are bundled using actin filaments, which form arches between these adjacent FPs. The connection between neighboring FPs is the SD [6].
The SD is the main size-selective filter barrier in the kidney. The protein composition of SD justifies the importance of this cellular junction for glomerulus permeability. The main proteins of the SD are nephrin, P-cadherin, CD2AP, ZO-1, FAT, podocin, and Neph1 (Figure 1). These contractile proteins allow the maintenance of glomerular filtration and also indicate that the fusion of podocyte FPs and the obliteration of SD are highly related to their alterations, leading to NS [6,11].
There is another important protein associated with actin microfilaments in FPs, synaptopodin [12], and its expression can indicate alterations in podocytes and also in the response to therapy in many forms of NS [13]. Therefore, synaptopodin likely plays an important role in the podocyte FP molecular composition, interacting with membrane-associated guanylate kinase, WW, and PDZ domain-containing protein 1 (MAGI-1) such as actinin-4 [14]. This may indicate an association between its expression and alterations in the podocyte.
As mentioned, the molecular composition of the cytoskeleton in podocyte FPs is dynamic and determines the maintenance of glomerular filtration. This has been demonstrated by using the acute protamine sulfate (PS)/heparin perfusion model, which dramatically reduces FPs in rodents [15,16]. Interference with one of the three membrane domains of the basolateral portion of FPs, including the apical membrane domain, the SD complex, and the basal membrane domain, can lead to alterations in the actin cytoskeleton. The consequences of these alterations are the fusion and effacement of podocyte FPs and the obliteration of the SD.
2.3. Histopathology of Common Types of Nephrotic Syndrome in Childhood
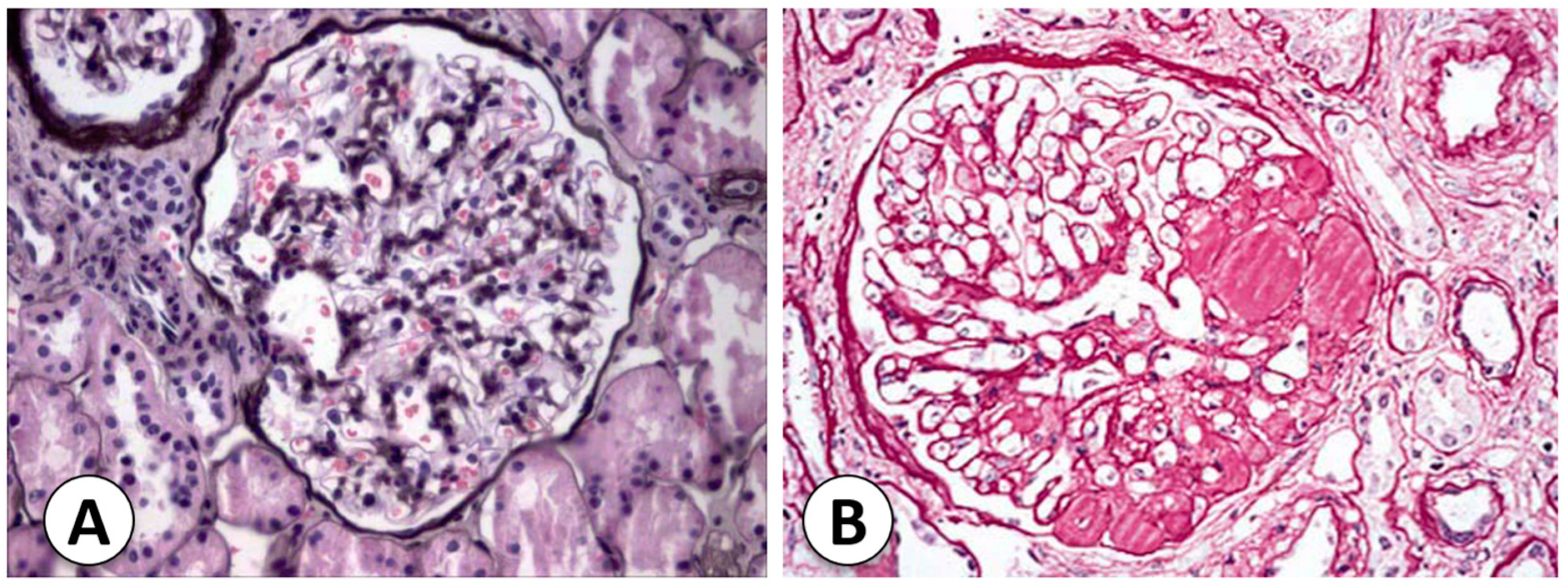
The types and causes of NS in childhood vary according to age. The most frequent type of NS in pediatric patients is primary or idiopathic disease, in which the etiology is not defined. When the NS starts before 1 year of age, the possibility of congenital NS should be considered, mostly in cases that occur before 6 months of age. Congenital NS is related to mutations in proteins related to SD. When nephrotic syndrome first appears during the school-age or adolescent years, it is important to consider the possibility of secondary NS due to systemic diseases [17]. The two most common histological forms of primary NS are MCNS and FSGS (Figure 2). Understanding histopathology is important to guide correct molecular and morphologic diagnoses and posterior management of NS [18].
2.3.1. Congenital Nephrotic Syndrome (Finnish Type)
This form of NS is highly related to prematurity in neonatal and prenatal presentations as well. In Finland, the incidence reaches 1:8200 births and is an autosomal-recessive disorder with heavy proteinuria in the neonatal period. The Finnish nephropathy gene is located on the long arm of chromosome 19 (19q13.1), which codes for nephrin [2]. The Finnish nephropathy gene is located on the long arm of chromosome 19 (19q13.1), which codes for nephrin. Around 70 changes to nephrin have been found in different parts of the body. These changes can be single nucleotide missense mutations, splices, insertions, deletions, or nonsense. Some mutations, such as Fin-major (deletion in exon 2) and Fin-minor (nonsense mutation in exon 26), are the most common. The kidney histopathology may help with congenital NS diagnosis, especially by using electron microscopy and immunohistochemistry [18]. A renal biopsy shows many glomeruli with mesangial hypercellularity and hyperlobulated capillary tufts. Furthermore, proximal and distal tubules may present microcystic dilatation, and the podocytes present FP effacement and/or villous transformations under electron microscopy. Immunohistochemical staining for nephrin is an important method to differentiate FN from other causes of congenital NS [2].
2.3.2. Diffuse Mesangial Sclerosis
The presentation of diffuse mesangial sclerosis (DMS) is similar to that of FN. However, its presentation can be later, up to 4 years of age, but usually persistent NS is described within the first 9 months of life. DMS is the second-most common cause of congenital NS. [1] The glomerular phenotype of the disease is associated with other disorders, such as Galloway–Mowat syndrome, Pierson syndrome, Frasier syndrome, and others. The pathology of the kidneys displays prominent, closely spaced podocytes in the early stages, and the glomeruli might be enlarged and exhibit hyaline casts. In immunofluorescence, nonspecific mesangial IgM, C3, and C1q deposits are found, while electron microscopy also shows podocyte hypertrophy and irregular FP effacement. In the late stages of DMS, the glomeruli are damaged and have thickened capillary loops, smaller capillary lumens, and a sclerotic mass in the mesangium [18].
2.3.3. Focal Segmental Glomerular Sclerosis
Focal segmental glomerular sclerosis (FSGS) is responsible for 10 to 20% of NS cases in children [2]. The podocyte is the main site of injury in FSGS. The damage occurs through a variety of mechanisms, including immune, toxic, and viral, mechanical injuries, and genetic dysfunction [19]. Injured podocytes are missed in the urinary space, resulting in podocyte effacement. The remaining podocytes hypertrophy and cover the glomerular capillary surface in response to this lack of podocytes. Intracapillary hypertension can cause damage to podocytes and endothelial cells. It can also cause changes in the mesangial cells that lead to sclerosis [20].
In primary FSGS, a circulating factor may be involved in the pathophysiology of NS. It is probably associated with some cytokines that disturb podocyte function [21]; therefore, it can reoccur after kidney transplantation [22]. In fact, the recurrence of idiopathic FSGS occurs in around one-third of patients after transplant [23]. Some molecules have been associated with the recurrence of FSGS, including cardiotrophin-like cytokine factor 1 [24], apoA1b (an isoform of ApoA1) [25], anti-CD40 antibody [26], and serum urine-type plasminogen activator receptor (suPAR) [27]. The association with suPAR is still controversial [28]. However, up to now, no molecule has been definitively established as the circulating factor in all cases of primary FSGS.
More than 50 genes have been presented as potential factors for monogenic forms of FSGS. The group of genes includes those involved in slit diaphragm structure, actin cytoskeleton, and cell-signaling apparatus [29,30]. FSGS can also be drug-induced, as IFN-α, -β, or -γ therapy has been linked to the occurrence of collapsing glomerulopathy in a case series of 11 subjects [31]. Anabolic steroid abuse has also been related to the development of FSGS. The association may be due to a combination of a direct nephrotoxic effect and adaptive glomerular changes to the increased lean body mass [32].
2.4. Histological Classifications for Focal Segmental Glomerular Sclerosis
2.4.1. Cellular Focal Segmental Glomerular Sclerosis
The description of this variant resembles focal proliferative glomerulonephritis (Table 1). On light microscopy, segmental hypercellularity and endocapillary proliferation are reported, with a luminal obliteration of capillaries [2]. Severe FP effacement is also common but with intact basement membranes [2,7]. Furthermore, there is an increase in mesangial and inflammatory cells such as neutrophils, foam cells, and monocytes. Although this variant shows marked podocyte hyperplasia [2], it is the least common one and is thought to be an early stage in the evolution of sclerotic lesions [7].
2.4.2. Primary Focal Segmental Glomerular Sclerosis with Mesangial Hypercellularity
This variant of FSGS reveals mainly mesangial hypercellularity in the non-sclerotic glomeruli, including the presence of IgM and C3 in these glomeruli on immunofluorescence patterns. Electron microscopy shows the classic segmental sclerosing lesions, severe FP effacement without any electron-dense deposits, and podocyte hyperplasia [2].
2.4.3. Familial Focal Segmental Glomerular Sclerosis
Familial FSGS is highly associated with genetic mutations, especially in genes that encode podocyte proteins [7]. Several defects in the proteins podocin (NPHS2 gene, chromosome 1q25-31) and actinin 4 (ACTN4, chromosome 19q13) cause autosomal dominant FSGS [2,18]. TRPC6 gene mutations lead to dysregulation of the cell cycle machinery with consequent apoptosis [18], while CD2AP mutations alter the bond between SD and the actin cytoskeleton [33]. Recent studies have shown an association between phospholipase C epsilon gene mutations (PLCE1, chromosome 10q23-33) and early onset NS [34]. The familial FSGS can manifest at any age and accounts for approximately 20% of FSGS cases [35].
2.4.4. Secondary Focal Segmental Glomerular Sclerosis
Other diseases may also trigger FSGS in pediatric patients, including IgA nephropathy [36], hereditary nephritis (Alport’s syndrome) [37], and lupus nephritis [38]. These diseases reveal different histological features, which can be traced using immunofluorescence: mesangial hypercellularity and mesangial IgA in IgA nephropathy, basement membrane abnormalities in Alport syndrome, and concurrent positive staining for IgA, IgM, IgG, C3, and C1q (full-house pattern) in lupus nephritis. Adults and children can develop secondary FSGS [2].
2.4.5. Collapsing Glomerulopathy
Severe and quick loss of renal function is highly associated with collapsing glomerulopathy [18]. This variant of FSGS is characterized by an implosive collapse of the capillary loops with alterations of the basement membrane, especially its wrinkling and contraction, and hypertrophy and hyperplasia of podocytes (Figure 3) [2]. There is no mesangial sclerosis, but the proliferating visceral podocytes and parietal epithelial cells fill Bowman’s space, resembling crescents (pseudocrescents) [18]. FP effacement is notorious even in glomeruli without collapsing lesions. In children, mitochondrial abnormalities have been reported to affect glomerular epithelial cells. COQ2 mutations inherited trigger an autosomal-recessive condition, which is the most common genetic variant of these mitochondrial disorders [39]. Ubiquinone replacement therapy is typical to treat this variant, and early recognition may prevent neurologic complications [18].
Collapsing glomerulopathy is linked to other causes like infections, drugs, and autoimmune diseases. The original description of this variant occurred in the 1980s during the HIV epidemic, but later it was described in other infections such as malaria and visceral leishmaniasis as well [40]. There is some evidence that other viral infections (cytomegalovirus, parvovirus B19, hepatitis C virus, dengue virus, and Zika virus) and systemic immune responses (tuberculosis-induced collapsing glomerulopathy) target podocytes and their proliferative machinery [41,42,43,44]. Patients with this variant are usually steroid-resistant and progress to renal failure quickly [2].
2.4.6. C1q Nephropathy
The description of this form of NS was first written by Jennette and Hipp and is still inconclusive [45]. Histological patterns in C1q nephropathy may vary from MCNS to mild mesangial proliferation and to FSGS. Electron microscopy usually shows mesangial electron-dense deposits, especially immunofluorescence staining for C1q. IgG and/or IgM are commonly present as well [46]. These deposits are predominantly located in paramesangial segments, and FP fusion is also reported. Some light microscopic findings of this entity are similar to the findings seen in lupus nephritis, which is an important differential diagnosis of C1q nephropathy. Furthermore, IgA nephropathy and the membranoproliferative pattern of histology may also distinguish the diagnosis via immunofluorescence. However, additional studies are necessary to understand the pathophysiology of C1q nephropathy and improve diagnosis and treatment [46].
2.5. Hypotheses Linking Glomerulosclerosis to Podocyte-Derived Alterations
The glomerular development is highly regulated with the podocyte, while its altered biology is a determinant of progression to glomerulosclerosis. Different glomerular diseases have been directly associated with alterations in podocyte functions and biology. Therefore, the central role of these specialized and polarized epithelial cells supports the concept of glomerular diseases as podocytopathies [47]. Although podocyte dysfunction, injury, or loss determine a common factor in the development of NS, many hypotheses propose that the pathogenesis of idiopathic NS is either immune-mediated or due to a genetic variant. The presence of a systemic circulating factor, likely podocyte-derived, has also been suggested [1,6].
2.5.1. Immune-Mediated
The immune system may be involved in the pathogenesis of NS, particularly a dysfunction or dysregulation of T lymphocytes [48]. Some evidence, such as the development of NS after allergic reactions to poisons and stings, reinforces the hypothesis. Furthermore, immunosuppressive agents play a role in NS response and even resolution [1]. In this sense, Lin et al. demonstrated spontaneous NS remission after infection with measles, which triggers prolonged depression of the immune system cells [49]. Chemotherapy for Hodgkin’s and other T-cell lymphomas in NS patients also affects the disease outcome. The literature supports an association between NS, especially MCNS, and classical Hodgkin’s lymphoma (cHL). MCNS associated with cHL is often dependent on or resistant to steroids, but the remission of NS is linked to the cure of the lymphoma [50]. MCNS can also occur in non-Hodgkin lymphoid disorders [51].
The antigen-presenting cells are responsible for the primary co-stimulatory signal for T-cell activation. CD80 (B7-1) is a protein expressed on those antigen-presenting cells and is the main molecular candidate for the immune-mediated feature in podocytopathies. CD80 binds to CTLA-4, a protein receptor expressed on the T-cell surface, and an increase in podocyte B7-1 expression may cause consequent dysfunction or dysregulation of T lymphocytes [52]. Clinical trials tested CTLA-4 as a therapeutic agent, like abatacept [53] and belatacept [54], in FSGS, but results on efficacy are still conflicting.
2.5.2. Systemic Circulating Factors
A hypothetical cause of NS is the existence of a circulating permeability factor that disturbs the glomerular permeability. It has been suggested that this supposed factor acts on the endothelial cell or podocyte, especially in steroid-resistant NS (SRNS) and FSGS [1]. A study developed in 1996 evaluated in vitro the glomerular permeability to albumin of recurrent FSGS patients. The investigation included the response to treatment and recurrence of FSGS after kidney transplantation in 30 to 40 percent of patients. This study sustains the hypothesis that a circulating factor is related to the recurrence of the disease and to the beginning of renal injury [55]. The existence of a circulating factor was further corroborated by the possibility of maternal transmission of FSGS. However, in utero exposure did not cause chronic glomerular disease, suggesting that the supposed factor would be smaller than IgG with a molecular weight between 30 and 50 kd and would not remain present in the fetus’s circulation [56].
Some molecules have been investigated as potential permeability factors, including hemopexin, heparanase, angiopoietin-like 4 (ANGPTL4), cardiotrophin-like cytokine-1, and suPAR [57,58,59]. The suPAR is found in the plasma of patients with recurrent FSGS and may lead to FP effacement and posterior proteinuria due to an interaction with α5β3-integrin receptors on the surface of podocytes [59]. ANGPTL4 is a glycoprotein expressed in the heart, adipose tissue, and skeletal muscle. Its concentration is elevated in NS. ANGPTL4 has two molecular forms, hyposialylated and sialylated, that trigger different mechanisms to regulate proteinuria [58]. The hyposialylated form is secreted via podocytes and affects GBM and endothelial cells, leading to damage to the filtration barrier and proteinuria [58]. On the other hand, the sialylated form binds to α5β3-integrins on the glomerular endothelium and reduces proteinuria. This mechanism has been explored as a potential treatment for NS. However, α5β3-integrin activation is lipid-dependent, and it causes hypertriglyceridemia in NS via inhibition of lipoprotein lipase [58].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Brief summary of the common histological types of childhood nephrotic syndrome.
| Nephrotic Syndrome | Characteristics | Causes | References |
|---|---|---|---|
| Congenital nephrotic syndrome (Finnish type) | Congenital NS is an autosomal-recessive disorder with heavy proteinuria in the neonatal period. A renal biopsy shows many glomeruli with mesangial hypercellularity and hyperlobulated capillary tufts. Proximal and distal tubules may present microcystic dilatation, and the podocytes present FP effacement. | The mutated gene related to Finnish nephropathy (FN) is NPHS1, mapped to the long arm of chromosome 19 (19q13.1), which codes for nephrin. | [2] |
| Diffuse mesangial sclerosis | It presents nonspecific mesangial IgM, C3, and C1q deposits, detected via immunofluorescence. Electron microscopy shows podocyte hypertrophy and irregular FP effacement. Late stages of DMS exhibit thickened capillary loops, a decrease in capillary lumens, and the formation of a sclerotic mass in the mesangium. | WT1 gene mutations in exons 8 or 9 are possible causes. | [2,18] |
| Focal segmental glomerular sclerosis | FSGS is related to the presence of segmental sclerotic lesions within the glomeruli. | Some molecules: cardiotrophin-like cytokine factor 1, apoA1b (an isoform of ApoA1), anti-CD40 antibody, and serum urine-type plasminogen activator receptor (suPAR). More than 50 genes are potential factors for monogenic forms of FSGS. Drugs and anabolic steroid abuse are other factors. | [2,6,24,25,26,27,29,30,31,32] |
| Cellular FSGS | It presents hypercellularity and endocapillary proliferation with luminal obliteration of capillaries. | ||
| Primary FSGS with mesangial hypercellularity | This type is associated with mesangial hypercellularity in the non-sclerotic glomeruli. | ||
| Familial FSGS | It exhibits loss of podocin staining, detected using immunohistochemistry methods. | Mutations in the NPHS2 gene, which codes for podocin. | [2,18] |
| Secondary FSGS | This type presents mesangial hypercellularity, mesangial IgA, and basement membrane abnormalities. | IgA nephropathy, hereditary nephritis (Alport’s syndrome), and lupus nephritis. | [2,51,52,53] |
| Collapsing glomerulopathy | It is related to the implosive collapse of the capillary loops with alterations of the basement membrane and hypertrophy and hyperplasia of podocytes. | COQ2 mutations that cause mitochondrial disorders. HIV, malaria, visceral leishmaniasis, cytomegalovirus, parvovirus B19, hepatitis C virus, dengue virus, and Zika virus are possible factors. | [2,55,56,57,58,59] |
NS: Nephrotic Syndrome; DMS: Diffuse Mesangial Sclerosis; FSGS: Focal Segmental Glomerulosclerosis; FP: Foot Process; GBM: Glomerular Basement Membrane; WT1: Wilms Tumor 1 Gene; NPHS1: Nephrin Gene; NPHS2: Podocin Gene; COQ2: Coenzyme Q2 Gene; suPAR: Soluble Urokinase-type Plasminogen Activator Receptor; HIV: Human Immunodeficiency Virus; Alport’s syndrome: Hereditary Nephritis; and IgA: Immunoglobulin A.
2.5.3. Genetic Variants
Mutations on genes encoding some components of the glomerular barrier, such as the GBM, podocyte, its mitochondria or lysosomes, slit diaphragm, actin cytoskeleton, or its molecular composition, can lead to proteinuria and NS. An understanding of the genes related to NS is important for the management of patients, especially considering the genetic variants and mutations associated with SRNS.
The early identification of genetic causes for SRNS allows for more appropriate management of the disease. Specific mutations in the genes NPHS1, NPHS2, LAMB2, and WT1 explain 69 to 85% of NS cases starting in the first three months of life. The presence of mutations in these genes decreases to 50 to 66% when NS starts between 4 and 12 months of life. If NS begins after 1 year of age, only 25% of cases are associated with genetic mutations [60]. The early identification of genetic causes supports the discontinuation of immunosuppressive agents and provides more information for prenatal counseling. In addition, the risk of recurrence after kidney transplant is lower in genetic forms of NS than in non-genetic-related syndromes. A multicentric study followed 1340 children with SRNS over 1 year of age and found that 14% of cases were associated with a genetic mutation [61], including polymorphisms of the major histocompatibility complex (HLA). HLA loci mutations represent a genetic risk for SSNS of 4 to 6 percent. However, the association between SSNS and these mutations is found only in specific ethnicities [62,63].
3. Molecular and Genetic Markers
3.1. Genetic Mutations
In recent years, studies have identified more than 50 genes as causes of NS in different populations. Mutations in the NPHS1 gene that codes for nephrin, described in 1998, led to an increase in studies searching for mutations in numerous components of the glomerular filtration barrier, including slit diaphragm structure and function, organization of podocyte actin cytoskeleton, biosynthesis of co-enzyme Q, lysosomal pathways, and adhesion to the GBM [33]. To date, the described mutations that caused NS can be divided into different categories, as summarized in Table 2.
3.2. Molecular Markers
The clinical characteristics of NS are commonly not predictive of patient outcomes [196]. The response to steroids has been associated with better outcomes in NS [197]. For patients with SRNS, renal biopsy is recommended to evaluate the prognosis and histopathological subtype, but it is an invasive procedure not free of complications, especially in children [198]. Furthermore, the incorrect interpretation and inadequate preparation of kidney tissue may lead to the wrong evaluation. Nevertheless, it is considered a generally safe procedure, especially when performed by trained nephrologists [199]. Therefore, new and less invasive biomarkers are necessary to predict disease outcomes, mainly for the pediatric age group. Indeed, urine is considered the ideal sample to search for biomarkers in renal diseases [200], but biomarkers in serum or plasma samples have also been evaluated in patients with NS.
3.2.1. MicroRNAs
Current studies have shown that microRNAs are a potential non-invasive biomarker that can help in the diagnosis of NS [200,201]. Some microRNAs, including hsa-miR-181a, hsa-miR-210, hsa-miR-30a, hsa-miR-942, hsa-miR-192, and hsa-miR-586, are up-regulated and detected in the serum of patients with NS. These markers might be associated with clinical findings in NS [201]. Another study showed that exosomal microRNAs, including miR-194-5p, miR-146b-5p, miR-378a-3p, miR-23b-3p, and miR-30a-5p, were found in the urine of patients with NS and might serve as novel biomarkers for diagnosing, monitoring, and stratifying pediatric patients with NS [200].
More recently, Bayomy et al. [202] showed in a cohort of children with NS that the measurement of miR-142a-5p in blood samples was capable of predicting SRNS with high accuracy (AUC = 0.965), suggesting the possible role of autoimmunity in the pathogenesis of NS and the resistance to steroids. Similarly, Zhang et al. [203] reported that miR-17-5p was significantly more expressed in the peripheral blood of pediatric patients with NS when compared to healthy controls. Nevertheless, most studies explored microRNAs in only one ethnicity, making multicenter studies necessary.
3.2.2. Proteomics
The study of urinary proteomics holds promise for identifying and predicting markers of renal disease in children [204]. Additionally, urinary proteomics is more stable than the blood proteome and closely matches pathological changes in the kidney and urogenital tract [205]. Some proteins that can be reliable biomarkers are only found in the urine. The pathogenesis of NS is strongly associated with alterations in the glomerular barrier and an increase in permeability to albumin. Additionally, a number of renal membrane proteins and receptors are subject to regulation via glycosylation [206].
Alfa 1-B glycoprotein (A1BG) is part of the immunoglobulin superfamily, but its function is still unclear. Piyaphanee et al. [207] used a proteomics approach to evaluate the urine of patients with idiopathic NS and healthy controls. A fragment of A1BG was exclusively identified in patients diagnosed with NS. A 13.8 kd fragment was only seen in patients with SRNS but not in patients with SSNS [207]. Nonetheless, no further studies were performed to validate these results, though A1BG seems to be a promising biomarker. In a similar approach, Bennett et al. [208] used a proteomics panel of 10 biomarkers (including A1BG) in the urine of 50 children with idiopathic NS. This panel showed a high capacity for predicting steroid responsiveness (AUC of 0.92). Nonetheless, multicenter studies are needed to validate the results found with this proteomics panel.
3.2.3. Inflammatory Markers, Cytokines, and Chemokines
One of the most commonly evaluated markers is inflammatory molecules. To date, urinary and blood levels of cytokines and chemokines often differ in NS patients if compared to healthy controls. Some studies have provided evidence that upregulation of interleukin (IL)-2 and its soluble receptor (sIL2R) may be involved not only in the pathophysiology of NS but also in steroid resistance to treatment by increasing the MDR1 gene expression of its product, P-glycoprotein [209]. Another biomarker that can help in the detection of steroid-resistant patients is suPAR. Even though this marker cannot distinguish the histopathological type of NS, it can be helpful in predicting steroid resistance in children with primary NS [210].
Chemokines, an extensive category of low-molecular-weight cytokines, predominantly function in the recruitment of leukocyte subsets during both homeostatic conditions and inflammation. These molecules hold promise as molecular markers owing to their pivotal roles in the pathophysiology of pediatric renal diseases [211]. The assessment of these inflammatory mediators presents potential utility in managing diverse renal conditions in children and identifying individuals at heightened risk for chronic kidney disease (CKD). The prevailing evidence underscores that the infiltration of leukocytes, orchestrated via inflammatory chemokines emanating from various cellular sources, has the potential to augment inflammatory responses within the kidney [212].
Multiple methodologies are available for quantifying chemokine levels, encompassing both protein and mRNA, in tissues and body fluids. Direct measurement of chemokines in renal tissue can be accomplished through immunohistochemical or immunofluorescent techniques, or their concentrations can be gauged in the supernatants of homogenized tissues. Conversely, less invasive approaches, such as enzyme-linked immunosorbent assays (ELISA) or flow cytometry-based techniques employing plasma and serum, prove to be more clinically useful for assessing chemokine levels in urine or blood samples [211].
The measurement of specific cytokines and chemokines emerges as a valuable tool in discerning between histopathological types of NS. Notably, patients with MCNS exhibit compromised T regulatory cells, accompanied by diminished levels of IL-10 [213]. Furthermore, distinctions between FSGS and MCNS may be drawn through urinary levels of the fibrogenic cytokine transforming growth factor beta (TGF-β), although its applicability as a biomarker for steroid responsiveness remains inconclusive [214]. Significantly elevated urinary levels of the chemokine monocyte chemoattractant protein 1 (MCP-1/CCL2) were observed in FSGS patients relative to counterparts with uropathies at equivalent CKD stages [71]. Additionally, urinary MCP-1/CCL2 levels exhibited positive correlations with serum total cholesterol and triglyceride concentrations [215]. These concentrations demonstrated noteworthy positive associations with occult blood in urine and inverse relationships with the estimated glomerular filtration rate. Moreover, urinary MCP-1/CCL2 concentrations displayed substantial correlations with histological chronicity indices in patients diagnosed with lupus nephritis and IgA nephropathy [216]. This supports the contention that the assessment of this chemokine holds promise as a noninvasive method for predicting the activity of glomerular diseases in the pediatric population.
Cardiotrophin-like cytokine factor 1 (CLCF-1) is a cytokine present in the IL-6 family. Savin et al. showed that a single intraperitoneal injection of CLCF-1 in C57Bl6 mice increased the urine albumin/creatinine ratio, and the chronic infusion of the cytokine induced segmental sclerosis in rare glomeruli [24]. These findings suggest that CLFC-1 might be involved with the development of FSGS in humans. Nonetheless, in patients with FSGS and MCNS, serum levels of CLCF-1 were not associated with renal function nor with histological markers of kidney injury [217]. Therefore, its role as a biomarker for NS is still unclear.
3.2.4. Other Potential Markers
Proteins found in the urine of NS patients named R6703, R7210, R11820, and R14356 are differentially expressed in SRNS and can be used to suggest treatment for NS patients [218]. Similarly, lower levels of APOL1 in urine are suggestive of SRNS. Other urinary biomarkers of NS, which were specifically associated with FSGS, included alpha 2 macroglobulin, retinol binding protein 4, and orosomucoid 2. Alpha 2 macroglobulin had the highest predictive power for the diagnosis of FSGS [219].
The vitamin D-binding protein (VDB) and APOL1 were regarded as candidate predictive biomarkers for steroid-resistant nephrotic syndrome (SRNS). Similarly, hemopexin (HPX), adiponectin (ADIPOQ), and sex hormone-binding globulin (SHBG) have been proposed as candidate biomarkers for mechanistically differentiating SRNS from SSNS [220]. VDB deserves special attention since vitamin D may play an essential role in the pathophysiology of NS. Patients with NS have high levels of vitamin D in their urine but low serum levels [221,222]. Urinary VDB levels were significantly higher in patients with SRNS in comparison to those with SSNS [223].
Neutrophil gelatinase-associated lipocalin (NGAL) is expressed at low levels in the kidney [224] and is considered an initial marker of renal injury [225]. In addition, NGAL has been considered a notable predictor of disease evolution in patients with chronic kidney disease [226,227]. In that sense, Bennet et al. [228] compared urinary NGAL levels between patients with SSNS, SRNS, and healthy controls. Not only did the authors find that urine NGAL levels were clearly higher in patients with SRNS when compared to the other two groups, but urinary NGAL was a good biomarker for predicting steroid-responsiveness (AUC of 0.91) [228]. As a result, NGAL levels may be a good predictor for steroid resistance in patients with NS. Nevertheless, it is relevant to consider that these results need to be validated in a larger cohort of patients.
3.3. Metabolomics
Metabolomics comprises a robust set of tools for phenotype analysis, hypothesis generation, and subsequent testing. The execution of metabolomics involves employing analytical technologies capable of generating diagnostic patterns through fingerprinting and achieving absolute quantitation of specific targeted metabolites [229]. Metabolomics has become an invaluable analytical tool for discerning urinary and serum biomarkers associated with renal diseases [230]. The thorough exploration of the metabolome stands as a pivotal method for investigating the phenotype and its alterations induced by environmental factors, pathological conditions, or variations in genotype.
Studies employing a combined proteomics and metabolomics approach, utilizing label-free mass spectrometry, have unveiled, for the first time, the implication of the glycoprotein afamin and several specific metabolites—hydroxyphenylacetate, uridine, glutamate, and phenylalanine—in the pathogenesis of primary NS [231]. Importantly, analogous patterns were evident in both protein and metabolite datasets, underscoring the potential of this integrated approach for advancing non-invasive diagnostic strategies for NS and its possible incorporation into routine clinical screening procedures for the condition.
Using a metabolomic discovery approach [232], Gooding et al. found a small group of candidate biomarkers predictive of SRNS in pediatric patients, such as glutamine and malonate. Through paired-sample analyses, a broader set of potential candidate metabolite biomarkers has been identified, offering insights into specific mechanistic molecular pathways and targets associated with steroid resistance. This expanded group includes lipoproteins, adipate, pyruvate, creatine, glucose, tyrosine, valine, and glutamine [232]. Additionally, glucocorticoid treatment in children with SRNS has been found to induce changes in a singular endogenous metabolite, sn-glycero-3-phosphocholine, distinguishing it from individuals with SSNS. Collectively, these findings pinpoint specific candidate metabolite biomarkers capable of predicting SRNS at the onset of the disease, while also highlighting potential therapeutic target pathways that could pave the way for the future development of more precise and effective treatments for NS.
3.4. Biomarkers Associated with Histological Features
The morphological picture of glomerulopathies is of great practical importance since, among all kidney diseases, they are the most diagnosable with a renal biopsy. The same form of clinical presentation can relate to very different morphological lesions with different pathogenic mechanisms, prognosis, and therapeutic approaches. Similarly, a morphological lesion pattern can be associated with different etiologies, involving specific prognosis and treatment.
Although kidney biopsy is the gold standard for NS diagnosis, this kind of invasive procedure is not routinely performed in children [233]. It is usually requested in certain specific situations, such as when there is the possibility of a coexisting nephritic syndrome or when the patient is evolving with symptoms that are not typically associated with MCNS, such as hypertension, altered renal function, gross hematuria, hypocomplementemia, and steroid resistance [234].
Generally, patients with FSGS present a low percentage (20%) of remission with steroid therapy. Thus, these patients are more likely to be submitted for a renal biopsy. On the other hand, about 90% of patients with MCNS are steroid-sensitive, and, for this reason, renal biopsies are not indicated [11]. With the purpose of minimizing the number of unnecessary biopsies, especially in children, there are some biomarkers associated with histological features that can help establish whether this procedure is really needed.
3.4.1. Transforming Growth Factor Beta (TGF-β)
It is known that TGF-β is overexpressed by podocytes in patients who are diagnosed with progressive podocytopathies, especially FSGS. This cytokine regulates the hypertrophy, proliferation, and apoptosis of renal cells. TGF-β also stimulates extracellular matrix (ECM) protein synthesis, GBM thickening, and mesangial matrix expansion [235]. Considering these actions, TGF-β could serve as a potential diagnostic biomarker in NS. In addition, this cytokine is correlated with hypoalbuminemia and dyslipidemia, which are typical clinical features of NS [236].
3.4.2. CD44
Another molecule that can play an important role as a histopathological marker is CD44. The expression of this glycoprotein, which is considered a marker of activated podocytes and parietal epithelial cells (PECs), has been implicated in the progression of renal fibrosis and the pathogenesis of FSGS. Therefore, CD44 immunostaining in PECs allows the differentiation of FSGS from MCNS. Patients with significantly increased PEC-CD44-positive renal tissue exhibited faster deterioration of renal function and the characteristics of typical FSGS [237,238].
3.4.3. CD80
CD80 (B7-1) is a co-stimulatory molecule that has been reported to be expressed in patients who have been diagnosed with glomerular diseases [239], especially those with FSGS. Expressed on antigen-presenting cells, this histological marker significantly contributes to T-cell activation. CD80, a key player in this process, is modulated via the cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4), primarily found on regulatory T cells (Tregs) [240]. Therefore, the detection of CD80 in the renal tissue of patients with SRNS indicates lower amounts of Tregs [241].
3.4.4. Extracellular Vesicles
Extracellular vesicles (EVs) are small, membrane-enclosed structures released via cells into the extracellular environment that play a crucial role in intercellular communication by carrying and transferring various bioactive molecules, including proteins, lipids, and nucleic acids (such as RNA and DNA), between cells. EVs have been implicated in various physiological and pathological processes, including immune response regulation, tissue repair, and the progression of diseases. In that sense, Eroglu et al. (2021) have found that patients with SSNS during relapse had higher protein content in circulating plasma EVs, especially proteins that involved actin cytoskeleton rearrangement, such as Ras-related C3 botulinum toxin substrate 1 (RAC-1) [242]. EVs have also been shown to induce mesangial cell proliferation in pediatric patients with FSGS through the phospho-STAT-3 pathway [243]. This topic has been recently reviewed elsewhere [244].
4. Conclusions
NS is a very complex disease with different outcomes, therapeutic responses, and histopathological patterns. The most characteristic finding of NS is the presence of podocytopathy. The function and/or the structure of the components of the podocytes can be altered through several mechanisms, including gene mutations, immunologic alterations, infectious diseases, and circulating factors. The pathophysiology of NS is still not fully understood.
There are potential biomarkers that can help with understanding the pathophysiology of NS. The advances in this research field might improve the clinical outcomes and therapeutic approaches of patients with NS. Furthermore, recent studies showing that different methods of assessing urinary molecules are less invasive and potentially more accurate for diagnosing SRNS are of particular importance for pediatric patients.
Author Contributions
Conceptualization, P.A.S.V.d.C. and A.C.S.e.S.; writing—original draft preparation, P.A.S.V.d.C., T.F.I., F.C.T. and A.C.S.e.S.; writing—review and editing, P.A.S.V.d.C., T.F.I., F.C.T. and A.C.S.e.S.; visualization, P.A.S.V.d.C., T.F.I., F.C.T. and A.C.S.e.S.; supervision, P.A.S.V.d.C. and A.C.S.e.S.; project administration, P.A.S.V.d.C. and A.C.S.e.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by the Brazilian National Council of Research Development (CNPq—Grant # 302153/2019-5), the Coordination of High Education Level Personnel (CAPES), and the Foundation of Research of Minas Gerais (FAPEMIG—CDS—APQ-02541-17).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Noone, D.G.; Iijima, K.; Parekh, R. Idiopathic Nephrotic Syndrome in Children. Lancet 2018, 392, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S. Pathology of Podocytopathies Causing Nephrotic Syndrome in Children. Front. Pediatr. 2016, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Roth, K.S.; Amaker, B.H.; Chan, J.C.M. Nephrotic Syndrome: Pathogenesis and Management. Pediatr. Rev. 2002, 23, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Banh, T.H.M.; Hussain-Shamsy, N.; Patel, V.; Vasilevska-Ristovska, J.; Borges, K.; Sibbald, C.; Lipszyc, D.; Brooke, J.; Geary, D.; Langlois, V.; et al. Ethnic Differences in Incidence and Outcomes of Childhood Nephrotic Syndrome. Clin. J. Am. Soc. Nephrol. 2016, 11, 1760–1768. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A.; Symons, J.M. Nephrotic Syndrome in Childhood. Lancet 2003, 362, 629–639. [Google Scholar] [CrossRef]
- Mundel, P.; Shankland, S.J. Podocyte Biology and Response to Injury. J. Am. Soc. Nephrol. 2002, 13, 3005–3015. [Google Scholar] [CrossRef]
- D’Agati, V.D.; Kaskel, F.J.; Falk, R.J. Focal Segmental Glomerulosclerosis. N. Engl. J. Med. 2011, 365, 2398–2411. [Google Scholar] [CrossRef]
- Downie, M.L.; Gallibois, C.; Parekh, R.S.; Noone, D.G. Nephrotic Syndrome in Infants and Children: Pathophysiology and Management. Paediatr. Int. Child. Health 2017, 37, 248–258. [Google Scholar] [CrossRef]
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The Glomerulus: The Sphere of Influence. Clin. J. Am. Soc. Nephrol. 2014, 9, 1461–1469. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Nishinakamura, R. Podocyte Development, Disease, and Stem Cell Research. Kidney Int. 2019, 96, 1077–1082. [Google Scholar] [CrossRef]
- Donoviel, D.B.; Freed, D.D.; Vogel, H.; Potter, D.G.; Hawkins, E.; Barrish, J.P.; Mathur, B.N.; Turner, C.A.; Geske, R.; Montgomery, C.A.; et al. Proteinuria and Perinatal Lethality in Mice Lacking NEPH1, a Novel Protein with Homology to NEPHRIN. Mol. Cell Biol. 2001, 21, 4829–4836. [Google Scholar] [CrossRef] [PubMed]
- Blaine, J.; Dylewski, J. Regulation of the Actin Cytoskeleton in Podocytes. Cells 2020, 9, 1700. [Google Scholar] [CrossRef]
- Yanagida-Asanuma, E.; Asanuma, K.; Kim, K.; Donnelly, M.; Young Choi, H.; Hyung Chang, J.; Suetsugu, S.; Tomino, Y.; Takenawa, T.; Faul, C.; et al. Synaptopodin Protects Against Proteinuria by Disrupting Cdc42:IRSp53:Mena Signaling Complexes in Kidney Podocytes. Am. J. Pathol. 2007, 171, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, T.; Garola, R.E.; Whiting, J.M.; Alon, U.S. Synaptopodin Expression in Idiopathic Nephrotic Syndrome of Childhood. Kidney Int. 2001, 59, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Seiler, M.W.; Venkatachalam, M.A.; Cotran, R.S. Glomerular Epithelium: Structural Alterations Induced by Polycations. Science 1975, 189, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Kerjaschki, D. Polycation-Induced Dislocation of Slit Diaphragms and Formation of Cell Junctions in Rat Kidney Glomeruli: The Effects of Low Temperature, Divalent Cations, Colchicine, and Cytochalasin B. Lab. Investig. 1978, 39, 430–440. [Google Scholar] [PubMed]
- da Silva Filha, R.; Burini, K.; Pires, L.G.; Brant Pinheiro, S.V.; Simões E Silva, A.C. Idiopathic Nephrotic Syndrome in Pediatrics: An Up-to-Date. Curr. Pediatr. Rev. 2022, 18, 251–264. [Google Scholar] [CrossRef]
- Liapis, H. Molecular Pathology of Nephrotic Syndrome in Childhood: A Contemporary Approach to Diagnosis. Pediatr. Dev. Pathol. 2008, 11, 154–163. [Google Scholar] [CrossRef]
- Kriz, W.; Elger, M.; Nagata, M.; Kretzler, M.; Uiker, S.; Koeppen-Hageman, I.; Tenschert, S.; Lemley, K.V. The Role of Podocytes in the Development of Glomerular Sclerosis. Kidney Int. Suppl. 1994, 45, S64–S72. [Google Scholar]
- Kriz, W.; Endlich, K. Hypertrophy of Podocytes: A Mechanism to Cope with Increased Glomerular Capillary Pressures? Kidney Int. 2005, 67, 373–374. [Google Scholar] [CrossRef]
- Königshausen, E.; Sellin, L. Circulating Permeability Factors in Primary Focal Segmental Glomerulosclerosis: A Review of Proposed Candidates. Biomed. Res. Int. 2016, 2016, 3765608. [Google Scholar] [CrossRef] [PubMed]
- Gallon, L.; Leventhal, J.; Skaro, A.; Kanwar, Y.; Alvarado, A. Resolution of Recurrent Focal Segmental Glomerulosclerosis after Retransplantation. N. Engl. J. Med. 2012, 366, 1648–1649. [Google Scholar] [CrossRef] [PubMed]
- Uffing, A.; Pérez-Sáez, M.J.; Mazzali, M.; Manfro, R.C.; Bauer, A.C.; de Drumond, F.S.; O’Shaughnessy, M.M.; Cheng, X.S.; Chin, K.-K.; Ventura, C.G.; et al. Recurrence of FSGS after Kidney Transplantation in Adults. Clin. J. Am. Soc. Nephrol. CJASN 2020, 15, 247. [Google Scholar] [CrossRef] [PubMed]
- Savin, V.J.; Sharma, M.; Zhou, J.; Gennochi, D.; Fields, T.; Sharma, R.; McCarthy, E.T.; Srivastava, T.; Domen, J.; Tormo, A.; et al. Renal and Hematological Effects of CLCF-1, a B-Cell-Stimulating Cytokine of the IL-6 Family. J. Immunol. Res. 2015, 2015, 714964. [Google Scholar] [CrossRef]
- Lopez-Hellin, J.; Cantarell, C.; Jimeno, L.; Sanchez-Fructuoso, A.; Puig-Gay, N.; Guirado, L.; Vilariño, N.; Gonzalez-Roncero, F.M.; Mazuecos, A.; Lauzurica, R.; et al. A Form of Apolipoprotein A-I Is Found Specifically in Relapses of Focal Segmental Glomerulosclerosis Following Transplantation. Am. J. Transplant. 2013, 13, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Delville, M.; Sigdel, T.K.; Wei, C.; Li, J.; Hsieh, S.-C.; Fornoni, A.; Burke, G.W.; Bruneval, P.; Naesens, M.; Jackson, A.; et al. A Circulating Antibody Panel for Pretransplant Prediction of FSGS Recurrence after Kidney Transplantation. Sci. Transl. Med. 2014, 6, 256ra136. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; El Hindi, S.; Li, J.; Fornoni, A.; Goes, N.; Sageshima, J.; Maiguel, D.; Karumanchi, S.A.; Yap, H.-K.; Saleem, M.; et al. Circulating Urokinase Receptor as a Cause of Focal Segmental Glomerulosclerosis. Nat. Med. 2011, 17, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Maas, R.J.H.; Deegens, J.K.J.; Wetzels, J.F.M. Serum suPAR in Patients with FSGS: Trash or Treasure? Pediatr. Nephrol. 2013, 28, 1041–1048. [Google Scholar] [CrossRef]
- Rood, I.M.; Deegens, J.K.J.; Wetzels, J.F.M. Genetic Causes of Focal Segmental Glomerulosclerosis: Implications for Clinical Practice. Nephrol. Dial. Transplant. 2012, 27, 882–890. [Google Scholar] [CrossRef]
- Santín, S.; Bullich, G.; Tazón-Vega, B.; García-Maset, R.; Giménez, I.; Silva, I.; Ruíz, P.; Ballarín, J.; Torra, R.; Ars, E. Clinical Utility of Genetic Testing in Children and Adults with Steroid-Resistant Nephrotic Syndrome. Clin. J. Am. Soc. Nephrol. 2011, 6, 1139–1148. [Google Scholar] [CrossRef]
- Markowitz, G.S.; Appel, G.B.; Fine, P.L.; Fenves, A.Z.; Loon, N.R.; Jagannath, S.; Kuhn, J.A.; Dratch, A.D.; D’Agati, V.D. Collapsing Focal Segmental Glomerulosclerosis Following Treatment with High-Dose Pamidronate. J. Am. Soc. Nephrol. 2001, 12, 1164–1172. [Google Scholar] [CrossRef]
- Herlitz, L.C.; Markowitz, G.S.; Farris, A.B.; Schwimmer, J.A.; Stokes, M.B.; Kunis, C.; Colvin, R.B.; D’Agati, V.D. Development of Focal Segmental Glomerulosclerosis after Anabolic Steroid Abuse. J. Am. Soc. Nephrol. 2010, 21, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Rheault, M.N.; Gbadegesin, R.A. The Genetics of Nephrotic Syndrome. J. Pediatr. Genet. 2016, 5, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Hinkes, B.; Wiggins, R.C.; Gbadegesin, R.; Vlangos, C.N.; Seelow, D.; Nürnberg, G.; Garg, P.; Verma, R.; Chaib, H.; Hoskins, B.E.; et al. Positional Cloning Uncovers Mutations in PLCE1 Responsible for a Nephrotic Syndrome Variant That May Be Reversible. Nat. Genet. 2006, 38, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- D’Agati, V.D. Pathobiology of Focal Segmental Glomerulosclerosis: New Developments. Curr. Opin. Nephrol. Hypertens. 2012, 21, 243–250. [Google Scholar] [CrossRef]
- Paranhos, R.M.; De Souza Figueiredo, G.A.; De Abreu, G.R.; Ferreira, G.C.; Fonseca, G.G.; Simões E Silva, A.C. Immunoglobulin A Nephropathy in Paediatrics: An up-to-Date. Nephrology 2022, 27, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, A.L.; Bitencourt, L.; Paranhos, R.M.; Leitáo, C.A.; Ferreira, G.C.; Simões E Silva, A.C. Alport Syndrome: A Comprehensive Review on Genetics, Pathophysiology, Histology, Clinical and Therapeutic Perspectives. Curr. Med. Chem. 2021, 28, 5602–5624. [Google Scholar] [CrossRef]
- Pinheiro, S.V.B.; Dias, R.F.; Fabiano, R.C.G.; de Araujo, S.A.; Silva, A.C.S.E. Pediatric Lupus Nephritis. J. Bras. Nefrol. 2019, 41, 252–265. [Google Scholar] [CrossRef]
- Diomedi-Camassei, F.; Di Giandomenico, S.; Santorelli, F.M.; Caridi, G.; Piemonte, F.; Montini, G.; Ghiggeri, G.M.; Murer, L.; Barisoni, L.; Pastore, A.; et al. COQ2 Nephropathy: A Newly Described Inherited Mitochondriopathy with Primary Renal Involvement. J. Am. Soc. Nephrol. 2007, 18, 2773–2780. [Google Scholar] [CrossRef]
- Albaqumi, M.; Barisoni, L. Current Views on Collapsing Glomerulopathy. J. Am. Soc. Nephrol. 2008, 19, 1276–1281. [Google Scholar] [CrossRef]
- Tanji, N.; Ross, M.D.; Tanji, K.; Bruggeman, L.A.; Markowitz, G.S.; Klotman, P.E.; D’Agati, V.D. Detection and Localization of HIV-1 DNA in Renal Tissues by in Situ Polymerase Chain Reaction. Histol. Histopathol. 2006, 21, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Albaqumi, M.; Soos, T.J.; Barisoni, L.; Nelson, P.J. Collapsing Glomerulopathy. J. Am. Soc. Nephrol. 2006, 17, 2854–2863. [Google Scholar] [CrossRef] [PubMed]
- Coventry, S.; Shoemaker, L.R. Collapsing Glomerulopathy in a 16-Year-Old Girl with Pulmonary Tuberculosis: The Role of Systemic Inflammatory Mediators. Pediatr. Dev. Pathol. 2004, 7, 166–170. [Google Scholar] [CrossRef]
- de Araújo, S.A.; Cordeiro, T.M.E.; Belisário, A.R.; Araújo, R.F.; Marinho, P.E.S.; Kroon, E.G.; de Oliveira, D.B.; Teixeira, M.M.; Simões E Silva, A.C. First Report of Collapsing Variant of Focal Segmental Glomerulosclerosis Triggered by Arbovirus: Dengue and Zika Virus Infection. Clin. Kidney J. 2019, 12, 355–361. [Google Scholar] [CrossRef]
- Jennette, J.C.; Hipp, C.G. C1q Nephropathy: A Distinct Pathologic Entity Usually Causing Nephrotic Syndrome. Am. J. Kidney Dis. 1985, 6, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, G.S.; Schwimmer, J.A.; Stokes, M.B.; Nasr, S.; Seigle, R.L.; Valeri, A.M.; D’Agati, V.D. C1q Nephropathy: A Variant of Focal Segmental Glomerulosclerosis. Kidney Int. 2003, 64, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, R.C. The Spectrum of Podocytopathies: A Unifying View of Glomerular Diseases. Kidney Int. 2007, 71, 1205–1214. [Google Scholar] [CrossRef]
- de Pereira, W.F.; Brito-Melo, G.E.A.; Guimarães, F.T.L.; Carvalho, T.G.R.; Mateo, E.C.; Simões e Silva, A.C. The Role of the Immune System in Idiopathic Nephrotic Syndrome: A Review of Clinical and Experimental Studies. Inflamm. Res. 2014, 63, 1–12. [Google Scholar] [CrossRef]
- Lin, C.Y.; Hsu, H.C. Histopathological and Immunological Studies in Spontaneous Remission of Nephrotic Syndrome after Intercurrent Measles Infection. Nephron 1986, 42, 110–115. [Google Scholar] [CrossRef]
- Audard, V.; Larousserie, F.; Grimbert, P.; Abtahi, M.; Sotto, J.-J.; Delmer, A.; Boue, F.; Nochy, D.; Brousse, N.; Delarue, R.; et al. Minimal Change Nephrotic Syndrome and Classical Hodgkin’s Lymphoma: Report of 21 Cases and Review of the Literature. Kidney Int. 2006, 69, 2251–2260. [Google Scholar] [CrossRef] [PubMed]
- Kofman, T.; Zhang, S.-Y.; Copie-Bergman, C.; Moktefi, A.; Raimbourg, Q.; Francois, H.; Karras, A.; Plaisier, E.; Painchart, B.; Favre, G.; et al. Minimal Change Nephrotic Syndrome Associated with Non-Hodgkin Lymphoid Disorders: A Retrospective Study of 18 Cases. Medicine 2014, 93, 350–358. [Google Scholar] [CrossRef]
- Reiser, J.; von Gersdorff, G.; Loos, M.; Oh, J.; Asanuma, K.; Giardino, L.; Rastaldi, M.P.; Calvaresi, N.; Watanabe, H.; Schwarz, K.; et al. Induction of B7-1 in Podocytes Is Associated with Nephrotic Syndrome. J. Clin. Investig. 2004, 113, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.W.; Chandar, J.; Sageshima, J.; Ortigosa-Goggins, M.; Amarapurkar, P.; Mitrofanova, A.; Defreitas, M.J.; Katsoufis, C.P.; Seeherunvong, W.; Centeno, A.; et al. Benefit of B7-1 Staining and Abatacept for Treatment-Resistant Post-Transplant Focal Segmental Glomerulosclerosis in a Predominantly Pediatric Cohort: Time for a Reappraisal. Pediatr. Nephrol. 2023, 38, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Delville, M.; Baye, E.; Durrbach, A.; Audard, V.; Kofman, T.; Braun, L.; Olagne, J.; Nguyen, C.; Deschênes, G.; Moulin, B.; et al. B7-1 Blockade Does Not Improve Post-Transplant Nephrotic Syndrome Caused by Recurrent FSGS. J. Am. Soc. Nephrol. 2016, 27, 2520–2527. [Google Scholar] [CrossRef]
- Savin, V.J.; Sharma, R.; Sharma, M.; McCarthy, E.T.; Swan, S.K.; Ellis, E.; Lovell, H.; Warady, B.; Gunwar, S.; Chonko, A.M.; et al. Circulating Factor Associated with Increased Glomerular Permeability to Albumin in Recurrent Focal Segmental Glomerulosclerosis. N. Engl. J. Med. 1996, 334, 878–883. [Google Scholar] [CrossRef]
- Kemper, M.J.; Wolf, G.; Müller-Wiefel, D.E. Transmission of Glomerular Permeability Factor from a Mother to Her Child. N. Engl. J. Med. 2001, 344, 386–387. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, P.E.C. Vascular Permeability Factors in Steroid-Sensitive Nephrotic Syndrome and Focal Segmental Glomerulosclerosis. Nephrol. Dial. Transplant. 2003, 18 (Suppl. S6), vi21–vi25. [Google Scholar] [CrossRef] [PubMed]
- Clement, L.C.; Macé, C.; Avila-Casado, C.; Joles, J.A.; Kersten, S.; Chugh, S.S. Circulating Angiopoietin-like 4 Links Proteinuria with Hypertriglyceridemia in Nephrotic Syndrome. Nat. Med. 2014, 20, 37–46. [Google Scholar] [CrossRef]
- McCarthy, E.T.; Sharma, M.; Savin, V.J. Circulating Permeability Factors in Idiopathic Nephrotic Syndrome and Focal Segmental Glomerulosclerosis. Clin. J. Am. Soc. Nephrol. 2010, 5, 2115–2121. [Google Scholar] [CrossRef]
- Sadowski, C.E.; Lovric, S.; Ashraf, S.; Pabst, W.L.; Gee, H.Y.; Kohl, S.; Engelmann, S.; Vega-Warner, V.; Fang, H.; Halbritter, J.; et al. A Single-Gene Cause in 29.5% of Cases of Steroid-Resistant Nephrotic Syndrome. J. Am. Soc. Nephrol. 2015, 26, 1279–1289. [Google Scholar] [CrossRef]
- Trautmann, A.; Bodria, M.; Ozaltin, F.; Gheisari, A.; Melk, A.; Azocar, M.; Anarat, A.; Caliskan, S.; Emma, F.; Gellermann, J.; et al. Spectrum of Steroid-Resistant and Congenital Nephrotic Syndrome in Children: The PodoNet Registry Cohort. Clin. J. Am. Soc. Nephrol. 2015, 10, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Gbadegesin, R.A.; Adeyemo, A.; Webb, N.J.A.; Greenbaum, L.A.; Abeyagunawardena, A.; Thalgahagoda, S.; Kale, A.; Gipson, D.; Srivastava, T.; Lin, J.-J.; et al. HLA-DQA1 and PLCG2 Are Candidate Risk Loci for Childhood-Onset Steroid-Sensitive Nephrotic Syndrome. J. Am. Soc. Nephrol. 2015, 26, 1701–1710. [Google Scholar] [CrossRef] [PubMed]
- Lagueruela, C.C.; Buettner, T.L.; Cole, B.R.; Kissane, J.M.; Robson, A.M. HLA Extended Haplotypes in Steroid-Sensitive Nephrotic Syndrome of Childhood. Kidney Int. 1990, 38, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Shih, N.Y.; Li, J.; Karpitskii, V.; Nguyen, A.; Dustin, M.L.; Kanagawa, O.; Miner, J.H.; Shaw, A.S. Congenital Nephrotic Syndrome in Mice Lacking CD2-Associated Protein. Science 1999, 286, 312–315. [Google Scholar] [CrossRef]
- Gigante, M.; Pontrelli, P.; Montemurno, E.; Roca, L.; Aucella, F.; Penza, R.; Caridi, G.; Ranieri, E.; Ghiggeri, G.M.; Gesualdo, L. CD2AP Mutations Are Associated with Sporadic Nephrotic Syndrome and Focal Segmental Glomerulosclerosis (FSGS). Nephrol. Dial. Transplant. 2009, 24, 1858–1864. [Google Scholar] [CrossRef] [PubMed]
- Löwik, M.M.; Groenen, P.J.T.A.; Pronk, I.; Lilien, M.R.; Goldschmeding, R.; Dijkman, H.B.; Levtchenko, E.N.; Monnens, L.A.; van den Heuvel, L.P. Focal Segmental Glomerulosclerosis in a Patient Homozygous for a CD2AP Mutation. Kidney Int. 2007, 72, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Philippe, A.; Nevo, F.; Esquivel, E.L.; Reklaityte, D.; Gribouval, O.; Tête, M.-J.; Loirat, C.; Dantal, J.; Fischbach, M.; Pouteil-Noble, C.; et al. Nephrin Mutations Can Cause Childhood-Onset Steroid-Resistant Nephrotic Syndrome. J. Am. Soc. Nephrol. 2008, 19, 1871–1878. [Google Scholar] [CrossRef]
- Li, X.; Chuang, P.Y.; D’Agati, V.D.; Dai, Y.; Yacoub, R.; Fu, J.; Xu, J.; Taku, O.; Premsrirut, P.K.; Holzman, L.B.; et al. Nephrin Preserves Podocyte Viability and Glomerular Structure and Function in Adult Kidneys. J. Am. Soc. Nephrol. 2015, 26, 2361–2377. [Google Scholar] [CrossRef]
- Kestilä, M.; Lenkkeri, U.; Männikkö, M.; Lamerdin, J.; McCready, P.; Putaala, H.; Ruotsalainen, V.; Morita, T.; Nissinen, M.; Herva, R.; et al. Positionally Cloned Gene for a Novel Glomerular Protein—Nephrin—Is Mutated in Congenital Nephrotic Syndrome. Mol. Cell 1998, 1, 575–582. [Google Scholar] [CrossRef]
- Bouchireb, K.; Boyer, O.; Gribouval, O.; Nevo, F.; Huynh-Cong, E.; Morinière, V.; Campait, R.; Ars, E.; Brackman, D.; Dantal, J.; et al. NPHS2 Mutations in Steroid-Resistant Nephrotic Syndrome: A Mutation Update and the Associated Phenotypic Spectrum. Hum. Mutat. 2014, 35, 178–186. [Google Scholar] [CrossRef]
- Boute, N.; Gribouval, O.; Roselli, S.; Benessy, F.; Lee, H.; Fuchshuber, A.; Dahan, K.; Gubler, M.C.; Niaudet, P.; Antignac, C. NPHS2, Encoding the Glomerular Protein Podocin, Is Mutated in Autosomal Recessive Steroid-Resistant Nephrotic Syndrome. Nat. Genet. 2000, 24, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Ebarasi, L.; Ashraf, S.; Bierzynska, A.; Gee, H.Y.; McCarthy, H.J.; Lovric, S.; Sadowski, C.E.; Pabst, W.; Vega-Warner, V.; Fang, H.; et al. Defects of CRB2 Cause Steroid-Resistant Nephrotic Syndrome. Am. J. Hum. Genet. 2015, 96, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. Identification and Characterization of Crumbs Homolog 2 Gene at Human Chromosome 9q33.3. Int. J. Oncol. 2004, 24, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Cil, O.; Perwad, F. Monogenic Causes of Proteinuria in Children. Front. Med. 2018, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, J.; Liang, X.; Chen, J.; Hong, J.; Li, L.; He, Q.; Cai, X. History and Progression of Fat Cadherins in Health and Disease. Onco. Targets Ther. 2016, 9, 7337–7343. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.Y.; Sadowski, C.E.; Aggarwal, P.K.; Porath, J.D.; Yakulov, T.A.; Schueler, M.; Lovric, S.; Ashraf, S.; Braun, D.A.; Halbritter, J.; et al. FAT1 Mutations Cause a Glomerulotubular Nephropathy. Nat. Commun. 2016, 7, 10822. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.; Hanby, A.M.; Poulsom, R.; Jones, T.A.; Sheer, D.; Chin, W.G.; Da, S.M.; Zhao, Q.; Beverley, P.C.; Owen, M.J. Molecular Cloning and Tissue Expression of FAT, the Human Homologue of the Drosophila Fat Gene That Is Located on Chromosome 4q34-Q35 and Encodes a Putative Adhesion Molecule. Genomics 1995, 30, 207–223. [Google Scholar] [CrossRef]
- Katoh, Y.; Katoh, M. Comparative Integromics on FAT1, FAT2, FAT3 and FAT4. Int. J. Mol. Med. 2006, 18, 523–528. [Google Scholar] [CrossRef]
- Yun, S.; Hong, W.-P.; Choi, J.H.; Yi, K.S.; Chae, S.-K.; Ryu, S.H.; Suh, P.-G. Phospholipase C-Epsilon Augments Epidermal Growth Factor-Dependent Cell Growth by Inhibiting Epidermal Growth Factor Receptor down-Regulation. J. Biol. Chem. 2008, 283, 341–349. [Google Scholar] [CrossRef]
- Gbadegesin, R.; Hinkes, B.G.; Hoskins, B.E.; Vlangos, C.N.; Heeringa, S.F.; Liu, J.; Loirat, C.; Ozaltin, F.; Hashmi, S.; Ulmer, F.; et al. Mutations in PLCE1 Are a Major Cause of Isolated Diffuse Mesangial Sclerosis (IDMS). Nephrol. Dial. Transplant. 2008, 23, 1291–1297. [Google Scholar] [CrossRef]
- Kanda, S.; Harita, Y.; Shibagaki, Y.; Sekine, T.; Igarashi, T.; Inoue, T.; Hattori, S. Tyrosine Phosphorylation–Dependent Activation of TRPC6 Regulated by PLC-Γ1 and Nephrin: Effect of Mutations Associated with Focal Segmental Glomerulosclerosis. Mol. Biol. Cell 2011, 22, 1824–1835. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Kim, E.Y.; Hagmann, H.; Benzing, T.; Dryer, S.E. Opposing Effects of Podocin on the Gating of Podocyte TRPC6 Channels Evoked by Membrane Stretch or Diacylglycerol. Am. J. Physiol. Cell Physiol. 2013, 305, C276–C289. [Google Scholar] [CrossRef] [PubMed]
- Gigante, M.; Caridi, G.; Montemurno, E.; Soccio, M.; d’Apolito, M.; Cerullo, G.; Aucella, F.; Schirinzi, A.; Emma, F.; Massella, L.; et al. TRPC6 Mutations in Children with Steroid-Resistant Nephrotic Syndrome and Atypical Phenotype. Clin. J. Am. Soc. Nephrol. 2011, 6, 1626–1634. [Google Scholar] [CrossRef]
- D’Esposito, M.; Strazzullo, M.; Cuccurese, M.; Spalluto, C.; Rocchi, M.; D’Urso, M.; Ciccodicola, A. Identification and Assignment of the Human Transient Receptor Potential Channel 6 Gene TRPC6 to Chromosome 11q21-->q22. Cytogenet. Cell Genet. 1998, 83, 46–47. [Google Scholar] [CrossRef]
- Feng, D.; Kumar, M.; Muntel, J.; Gurley, S.B.; Birrane, G.; Stillman, I.E.; Ding, L.; Wang, M.; Ahmed, S.; Schlondorff, J.; et al. Phosphorylation of ACTN4 Leads to Podocyte Vulnerability and Proteinuric Glomerulosclerosis. J. Am. Soc. Nephrol. 2020, 31, 1479–1495. [Google Scholar] [CrossRef]
- Dai, S.; Wang, Z.; Pan, X.; Chen, X.; Wang, W.; Ren, H.; Feng, Q.; He, J.C.; Han, B.; Chen, N. ACTN4 Gene Mutations and Single Nucleotide Polymorphisms in Idiopathic Focal Segmental Glomerulosclerosis. Nephron Clin. Pract. 2009, 111, c87–c94. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.M.; Kim, S.H.; North, K.N.; Rennke, H.; Correia, L.A.; Tong, H.Q.; Mathis, B.J.; Rodríguez-Pérez, J.C.; Allen, P.G.; Beggs, A.H.; et al. Mutations in ACTN4, Encoding Alpha-Actinin-4, Cause Familial Focal Segmental Glomerulosclerosis. Nat. Genet. 2000, 24, 251–256. [Google Scholar] [CrossRef]
- Gbadegesin, R.A.; Hall, G.; Adeyemo, A.; Hanke, N.; Tossidou, I.; Burchette, J.; Wu, G.; Homstad, A.; Sparks, M.A.; Gomez, J.; et al. Mutations in the Gene That Encodes the F-Actin Binding Protein Anillin Cause FSGS. J. Am. Soc. Nephrol. 2014, 25, 1991–2002. [Google Scholar] [CrossRef]
- Hall, G.; Lane, B.M.; Khan, K.; Pediaditakis, I.; Xiao, J.; Wu, G.; Wang, L.; Kovalik, M.E.; Chryst-Stangl, M.; Davis, E.E.; et al. The Human FSGS-Causing ANLN R431C Mutation Induces Dysregulated PI3K/AKT/mTOR/Rac1 Signaling in Podocytes. J. Am. Soc. Nephrol. 2018, 29, 2110–2122. [Google Scholar] [CrossRef]
- Katoh, M.; Katoh, M. Identification and Characterization of ARHGAP24 and ARHGAP25 Genes in Silico. Int. J. Mol. Med. 2004, 14, 333–338. [Google Scholar] [CrossRef]
- Akilesh, S.; Suleiman, H.; Yu, H.; Stander, M.C.; Lavin, P.; Gbadegesin, R.; Antignac, C.; Pollak, M.; Kopp, J.B.; Winn, M.P.; et al. Arhgap24 Inactivates Rac1 in Mouse Podocytes, and a Mutant Form Is Associated with Familial Focal Segmental Glomerulosclerosis. J. Clin. Investig. 2011, 121, 4127–4137. [Google Scholar] [CrossRef]
- Gupta, I.R.; Baldwin, C.; Auguste, D.; Ha, K.C.H.; El Andalousi, J.; Fahiminiya, S.; Bitzan, M.; Bernard, C.; Akbari, M.R.; Narod, S.A.; et al. ARHGDIA: A Novel Gene Implicated in Nephrotic Syndrome. J. Med. Genet. 2013, 50, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.; Tommerup, N.; Wirth, J.; Leffers, H.; Zimmer, J.; Back, E.; Weissenbach, J.; Scherer, G. A Somatic Cell Hybrid Panel for Distal 17q: GDIA1 Maps to 17q25.3. Cytogenet. Cell Genet. 1997, 76, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, E.S.; Higgs, H.N. INF2 Is a WASP Homology 2 Motif-Containing Formin That Severs Actin Filaments and Accelerates Both Polymerization and Depolymerization. J. Biol. Chem. 2006, 281, 26754–26767. [Google Scholar] [CrossRef] [PubMed]
- Boyer, O.; Nevo, F.; Plaisier, E.; Funalot, B.; Gribouval, O.; Benoit, G.; Huynh Cong, E.; Arrondel, C.; Tête, M.-J.; Montjean, R.; et al. INF2 Mutations in Charcot-Marie-Tooth Disease with Glomerulopathy. N. Engl. J. Med. 2011, 365, 2377–2388. [Google Scholar] [CrossRef] [PubMed]
- Mele, C.; Iatropoulos, P.; Donadelli, R.; Calabria, A.; Maranta, R.; Cassis, P.; Buelli, S.; Tomasoni, S.; Piras, R.; Krendel, M.; et al. MYO1E Mutations and Childhood Familial Focal Segmental Glomerulosclerosis. N. Engl. J. Med. 2011, 365, 295–306. [Google Scholar] [CrossRef]
- Krendel, M.; Kim, S.V.; Willinger, T.; Wang, T.; Kashgarian, M.; Flavell, R.A.; Mooseker, M.S. Disruption of Myosin 1e Promotes Podocyte Injury. J. Am. Soc. Nephrol. 2009, 20, 86–94. [Google Scholar] [CrossRef]
- Hasson, T.; Skowron, J.F.; Gilbert, D.J.; Avraham, K.B.; Perry, W.L.; Bement, W.M.; Anderson, B.L.; Sherr, E.H.; Chen, Z.Y.; Greene, L.A.; et al. Mapping of Unconventional Myosins in Mouse and Human. Genomics 1996, 36, 431–439. [Google Scholar] [CrossRef]
- Gee, H.Y.; Zhang, F.; Ashraf, S.; Kohl, S.; Sadowski, C.E.; Vega-Warner, V.; Zhou, W.; Lovric, S.; Fang, H.; Nettleton, M.; et al. KANK Deficiency Leads to Podocyte Dysfunction and Nephrotic Syndrome. J. Clin. Investig. 2015, 125, 2375–2384. [Google Scholar] [CrossRef]
- Sarkar, S.; Roy, B.C.; Hatano, N.; Aoyagi, T.; Gohji, K.; Kiyama, R. A Novel Ankyrin Repeat-Containing Gene (Kank) Located at 9p24 Is a Growth Suppressor of Renal Cell Carcinoma. J. Biol. Chem. 2002, 277, 36585–36591. [Google Scholar] [CrossRef]
- Pei, Q.; Ni, W.; Yuan, Y.; Yuan, J.; Zhang, X.; Yao, M. HSP70 Ameliorates Septic Lung Injury via Inhibition of Apoptosis by Interacting with KANK2. Biomolecules 2022, 12, 410. [Google Scholar] [CrossRef]
- Anjanappa, R.M.; Nayak, S.; Moily, N.S.; Manduva, V.; Nadella, R.K.; Viswanath, B.; Reddy, Y.C.J.; Jain, S.; Anand, A. A Linkage and Exome Study Implicates Rare Variants of KANK4 and CAP2 in Bipolar Disorder in a Multiplex Family. Bipolar Disord. 2020, 22, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.J.; Vazquez Fonseca, L.; Desbats, M.A.; Cerqua, C.; Zordan, R.; Trevisson, E.; Salviati, L. Coenzyme Q Biosynthesis in Health and Disease. Biochim. Biophys. Acta 2016, 1857, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Widmeier, E.; Yu, S.; Nag, A.; Chung, Y.W.; Nakayama, M.; Fernández-Del-Río, L.; Hugo, H.; Schapiro, D.; Buerger, F.; Choi, W.-I.; et al. ADCK4 Deficiency Destabilizes the Coenzyme Q Complex, Which Is Rescued by 2,4-Dihydroxybenzoic Acid Treatment. J. Am. Soc. Nephrol. 2020, 31, 1191–1211. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Gee, H.Y.; Woerner, S.; Xie, L.X.; Vega-Warner, V.; Lovric, S.; Fang, H.; Song, X.; Cattran, D.C.; Avila-Casado, C.; et al. ADCK4 Mutations Promote Steroid-Resistant Nephrotic Syndrome through CoQ10 Biosynthesis Disruption. J. Clin. Investig. 2013, 123, 5179–5189. [Google Scholar] [CrossRef]
- Forsgren, M.; Attersand, A.; Lake, S.; Grünler, J.; Swiezewska, E.; Dallner, G.; Climent, I. Isolation and Functional Expression of Human COQ2, a Gene Encoding a Polyprenyl Transferase Involved in the Synthesis of CoQ. Biochem. J. 2004, 382, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Quinzii, C.; Naini, A.; Salviati, L.; Trevisson, E.; Navas, P.; Dimauro, S.; Hirano, M. A Mutation in Para-Hydroxybenzoate-Polyprenyl Transferase (COQ2) Causes Primary Coenzyme Q10 Deficiency. Am. J. Hum. Genet. 2006, 78, 345–349. [Google Scholar] [CrossRef]
- Doimo, M.; Desbats, M.A.; Cerqua, C.; Cassina, M.; Trevisson, E.; Salviati, L. Genetics of Coenzyme Q10 Deficiency. Mol. Syndromol. 2014, 5, 156–162. [Google Scholar] [CrossRef]
- Gigante, M.; Diella, S.; Santangelo, L.; Trevisson, E.; Acosta, M.J.; Amatruda, M.; Finzi, G.; Caridi, G.; Murer, L.; Accetturo, M.; et al. Further Phenotypic Heterogeneity of CoQ10 Deficiency Associated with Steroid Resistant Nephrotic Syndrome and Novel COQ2 and COQ6 Variants. Clin. Genet. 2017, 92, 224–226. [Google Scholar] [CrossRef]
- Park, E.; Ahn, Y.H.; Kang, H.G.; Yoo, K.H.; Won, N.H.; Lee, K.B.; Moon, K.C.; Seong, M.-W.; Gwon, T.R.; Park, S.S.; et al. COQ6 Mutations in Children With Steroid-Resistant Focal Segmental Glomerulosclerosis and Sensorineural Hearing Loss. Am. J. Kidney Dis. 2017, 70, 139–144. [Google Scholar] [CrossRef]
- Heeringa, S.F.; Chernin, G.; Chaki, M.; Zhou, W.; Sloan, A.J.; Ji, Z.; Xie, L.X.; Salviati, L.; Hurd, T.W.; Vega-Warner, V.; et al. COQ6 Mutations in Human Patients Produce Nephrotic Syndrome with Sensorineural Deafness. J. Clin. Investig. 2011, 121, 2013–2024. [Google Scholar] [CrossRef]
- Li, Y.; Lin, S.; Li, L.; Tang, Z.; Hu, Y.; Ban, X.; Zeng, T.; Zhou, Y.; Zhu, Y.; Gao, S.; et al. PDSS2 Deficiency Induces Hepatocarcinogenesis by Decreasing Mitochondrial Respiration and Reprogramming Glucose Metabolism. Cancer Res. 2018, 78, 4471–4481. [Google Scholar] [CrossRef]
- Gasser, D.L.; Winkler, C.A.; Peng, M.; An, P.; McKenzie, L.M.; Kirk, G.D.; Shi, Y.; Xie, L.X.; Marbois, B.N.; Clarke, C.F.; et al. Focal Segmental Glomerulosclerosis Is Associated with a PDSS2 Haplotype and, Independently, with a Decreased Content of Coenzyme Q10. Am. J. Physiol. Renal Physiol. 2013, 305, F1228–F1238. [Google Scholar] [CrossRef] [PubMed]
- Desbats, M.A.; Lunardi, G.; Doimo, M.; Trevisson, E.; Salviati, L. Genetic Bases and Clinical Manifestations of Coenzyme Q10 (CoQ 10) Deficiency. J. Inherit. Metab. Dis. 2015, 38, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Mariyama, M.; Leinonen, A.; Mochizuki, T.; Tryggvason, K.; Reeders, S.T. Complete Primary Structure of the Human Alpha 3(IV) Collagen Chain. Coexpression of the Alpha 3(IV) and Alpha 4(IV) Collagen Chains in Human Tissues. J. Biol. Chem. 1994, 269, 23013–23017. [Google Scholar] [CrossRef] [PubMed]
- Hostikka, S.L.; Eddy, R.L.; Byers, M.G.; Höyhtyä, M.; Shows, T.B.; Tryggvason, K. Identification of a Distinct Type IV Collagen Alpha Chain with Restricted Kidney Distribution and Assignment of Its Gene to the Locus of X Chromosome-Linked Alport Syndrome. Proc. Natl. Acad. Sci. USA 1990, 87, 1606–1610. [Google Scholar] [CrossRef] [PubMed]
- Lemmink, H.H.; Schröder, C.H.; Monnens, L.A.; Smeets, H.J. The Clinical Spectrum of Type IV Collagen Mutations. Hum. Mutat. 1997, 9, 477–499. [Google Scholar] [CrossRef]
- Mochizuki, T.; Lemmink, H.H.; Mariyama, M.; Antignac, C.; Gubler, M.C.; Pirson, Y.; Verellen-Dumoulin, C.; Chan, B.; Schröder, C.H.; Smeets, H.J. Identification of Mutations in the Alpha 3(IV) and Alpha 4(IV) Collagen Genes in Autosomal Recessive Alport Syndrome. Nat. Genet. 1994, 8, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Morrison, K.E.; Mariyama, M.; Yang-Feng, T.L.; Reeders, S.T. Sequence and Localization of a Partial cDNA Encoding the Human Alpha 3 Chain of Type IV Collagen. Am. J. Hum. Genet. 1991, 49, 545–554. [Google Scholar]
- Wang, D.; Shan, C.; Jing, X.; Zhang, Q.; Chang, H.; Lin, Y. Clinical Features and Familial Mutations in an Autosomal-Inherited Alport Syndrome Patient With the Presentation of Nephrotic Syndrome. Front. Pediatr. 2021, 9, 678633. [Google Scholar] [CrossRef]
- Takada, Y.; Murphy, E.; Pil, P.; Chen, C.; Ginsberg, M.H.; Hemler, M.E. Molecular Cloning and Expression of the cDNA for Alpha 3 Subunit of Human Alpha 3 Beta 1 (VLA-3), an Integrin Receptor for Fibronectin, Laminin, and Collagen. J. Cell Biol. 1991, 115, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Has, C.; Spartà, G.; Kiritsi, D.; Weibel, L.; Moeller, A.; Vega-Warner, V.; Waters, A.; He, Y.; Anikster, Y.; Esser, P.; et al. Integrin A3 Mutations with Kidney, Lung, and Skin Disease. N. Engl. J. Med. 2012, 366, 1508–1514. [Google Scholar] [CrossRef]
- Jones, S.D.; van der Flier, A.; Sonnenberg, A. Genomic Organization of the Human Alpha 3 Integrin Subunit Gene. Biochem. Biophys. Res. Commun. 1998, 248, 896–898. [Google Scholar] [CrossRef] [PubMed]
- Nakano, A.; Pulkkinen, L.; Murrell, D.; Rico, J.; Lucky, A.W.; Garzon, M.; Stevens, C.A.; Robertson, S.; Pfendner, E.; Uitto, J. Epidermolysis Bullosa with Congenital Pyloric Atresia: Novel Mutations in the Beta 4 Integrin Gene (ITGB4) and Genotype/Phenotype Correlations. Pediatr. Res. 2001, 49, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Kambham, N.; Tanji, N.; Seigle, R.L.; Markowitz, G.S.; Pulkkinen, L.; Uitto, J.; D’Agati, V.D. Congenital Focal Segmental Glomerulosclerosis Associated with Beta4 Integrin Mutation and Epidermolysis Bullosa. Am. J. Kidney Dis. 2000, 36, 190–196. [Google Scholar] [CrossRef]
- Ashton, G.H.; Sorelli, P.; Mellerio, J.E.; Keane, F.M.; Eady, R.A.; McGrath, J.A. Alpha 6 Beta 4 Integrin Abnormalities in Junctional Epidermolysis Bullosa with Pyloric Atresia. Br. J. Dermatol. 2001, 144, 408–414. [Google Scholar] [CrossRef]
- Zenker, M.; Aigner, T.; Wendler, O.; Tralau, T.; Müntefering, H.; Fenski, R.; Pitz, S.; Schumacher, V.; Royer-Pokora, B.; Wühl, E.; et al. Human Laminin Beta2 Deficiency Causes Congenital Nephrosis with Mesangial Sclerosis and Distinct Eye Abnormalities. Hum. Mol. Genet. 2004, 13, 2625–2632. [Google Scholar] [CrossRef] [PubMed]
- Wewer, U.M.; Gerecke, D.R.; Durkin, M.E.; Kurtz, K.S.; Mattei, M.G.; Champliaud, M.F.; Burgeson, R.E.; Albrechtsen, R. Human Beta 2 Chain of Laminin (Formerly S Chain): cDNA Cloning, Chromosomal Localization, and Expression in Carcinomas. Genomics 1994, 24, 243–252. [Google Scholar] [CrossRef]
- Isojima, T.; Harita, Y.; Furuyama, M.; Sugawara, N.; Ishizuka, K.; Horita, S.; Kajiho, Y.; Miura, K.; Igarashi, T.; Hattori, M.; et al. LMX1B Mutation with Residual Transcriptional Activity as a Cause of Isolated Glomerulopathy. Nephrol. Dial. Transplant. 2014, 29, 81–88. [Google Scholar] [CrossRef]
- Dreyer, S.D.; Zhou, G.; Baldini, A.; Winterpacht, A.; Zabel, B.; Cole, W.; Johnson, R.L.; Lee, B. Mutations in LMX1B Cause Abnormal Skeletal Patterning and Renal Dysplasia in Nail Patella Syndrome. Nat. Genet. 1998, 19, 47–50. [Google Scholar] [CrossRef]
- Iannotti, C.A.; Inoue, H.; Bernal, E.; Aoki, M.; Liu, L.; Donis-Keller, H.; German, M.S.; Permutt, M.A. Identification of a Human LMX1 (LMX1.1)-Related Gene, LMX1.2: Tissue-Specific Expression and Linkage Mapping on Chromosome 9. Genomics 1997, 46, 520–524. [Google Scholar] [CrossRef]
- Herold, A.; Suyama, M.; Rodrigues, J.P.; Braun, I.C.; Kutay, U.; Carmo-Fonseca, M.; Bork, P.; Izaurralde, E. TAP (NXF1) Belongs to a Multigene Family of Putative RNA Export Factors with a Conserved Modular Architecture. Mol. Cell Biol. 2000, 20, 8996–9008. [Google Scholar] [CrossRef]
- Esposito, T.; Lea, R.A.; Maher, B.H.; Moses, D.; Cox, H.C.; Magliocca, S.; Angius, A.; Nyholt, D.R.; Titus, T.; Kay, T.; et al. Unique X-Linked Familial FSGS with Co-Segregating Heart Block Disorder Is Associated with a Mutation in the NXF5 Gene. Hum. Mol. Genet. 2013, 22, 3654–3666. [Google Scholar] [CrossRef] [PubMed]
- Bétous, R.; Glick, G.G.; Zhao, R.; Cortez, D. Identification and Characterization of SMARCAL1 Protein Complexes. PLoS ONE 2013, 8, e63149. [Google Scholar] [CrossRef] [PubMed]
- Elizondo, L.I.; Huang, C.; Northrop, J.L.; Deguchi, K.; Clewing, J.M.; Armstrong, D.L.; Boerkoel, C.F. Schimke Immuno-Osseous Dysplasia: A Cell Autonomous Disorder? Am. J. Med. Genet. A 2006, 140, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.A.; Eisen, J.A.; Mohrenweiser, H.W. Cloning and Characterization of HARP/SMARCAL1: A Prokaryotic HepA-Related SNF2 Helicase Protein from Human and Mouse. Genomics 2000, 65, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.-D.; Wagner, N.; Schley, G.; Theres, H.; Scholz, H. The Wilms’ Tumor Suppressor Wt1 Encodes a Transcriptional Activator of the Class IV POU-Domain Factor Pou4f2 (Brn-3b). Gene 2003, 305, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Bruening, W.; Li, F.P.; Haber, D.A.; Glaser, T.; Housman, D.E. WT1 Mutations Contribute to Abnormal Genital System Development and Hereditary Wilms’ Tumour. Nature 1991, 353, 431–434. [Google Scholar] [CrossRef]
- Pelletier, J.; Bruening, W.; Kashtan, C.E.; Mauer, S.M.; Manivel, J.C.; Striegel, J.E.; Houghton, D.C.; Junien, C.; Habib, R.; Fouser, L. Germline Mutations in the Wilms’ Tumor Suppressor Gene Are Associated with Abnormal Urogenital Development in Denys-Drash Syndrome. Cell 1991, 67, 437–447. [Google Scholar] [CrossRef]
- Little, M.H.; Williamson, K.A.; Mannens, M.; Kelsey, A.; Gosden, C.; Hastie, N.D.; van Heyningen, V. Evidence That WT1 Mutations in Denys-Drash Syndrome Patients May Act in a Dominant-Negative Fashion. Hum. Mol. Genet. 1993, 2, 259–264. [Google Scholar] [CrossRef]
- Salazar, H.; Kanbour, A.; Burgess, F. Ultrastructure and Observations on the Histogenesis of Mesotheliomas, “Adenomatoid Tumors”, of the Female Genital Tract. Cancer 1972, 29, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Rose, E.A.; Glaser, T.; Jones, C.; Smith, C.L.; Lewis, W.H.; Call, K.M.; Minden, M.; Champagne, E.; Bonetta, L.; Yeger, H. Complete Physical Map of the WAGR Region of 11p13 Localizes a Candidate Wilms’ Tumor Gene. Cell 1990, 60, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Edvardson, S.; Prunetti, L.; Arraf, A.; Haas, D.; Bacusmo, J.M.; Hu, J.F.; Ta-Shma, A.; Dedon, P.C.; de Crécy-Lagard, V.; Elpeleg, O. tRNA N6-Adenosine Threonylcarbamoyltransferase Defect Due to KAE1/TCS3 (OSGEP) Mutation Manifest by Neurodegeneration and Renal Tubulopathy. Eur. J. Hum. Genet. 2017, 25, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Tsai, J.D.; Lin, S.P.; Tzen, C.Y.; Shen, E.Y.; Shih, C.S. Galloway-Mowat Syndrome: A Glomerular Basement Membrane Disorder? Pediatr. Nephrol. 2001, 16, 653–657. [Google Scholar] [CrossRef]
- Seki, Y.; Ikeda, S.; Kiyohara, H.; Ayabe, H.; Seki, T.; Matsui, H. Sequencing Analysis of a Putative Human O-Sialoglycoprotein Endopeptidase Gene (OSGEP) and Analysis of a Bidirectional Promoter between the OSGEP and APEX Genes. Gene 2002, 285, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.A.; Rao, J.; Mollet, G.; Schapiro, D.; Daugeron, M.-C.; Tan, W.; Gribouval, O.; Boyer, O.; Revy, P.; Jobst-Schwan, T.; et al. Mutations in KEOPS-Complex Genes Cause Nephrotic Syndrome with Primary Microcephaly. Nat. Genet. 2017, 49, 1529–1538. [Google Scholar] [CrossRef]
- Liu, T.-L.; Lin, S.-P.; Zenker, M.; Chen, T.-Y.; Chang, J.-H.; Lin, C.-C.; Tsai, J.-D. X-Linked Recessive Galloway-Mowat Syndrome 2 Caused by a Specific LAGE3 Variant. Pediatr. Neonatol. 2023, 64, 208–209. [Google Scholar] [CrossRef]
- Jinks, R.N.; Puffenberger, E.G.; Baple, E.; Harding, B.; Crino, P.; Fogo, A.B.; Wenger, O.; Xin, B.; Koehler, A.E.; McGlincy, M.H.; et al. Recessive Nephrocerebellar Syndrome on the Galloway-Mowat Syndrome Spectrum Is Caused by Homozygous Protein-Truncating Mutations of WDR73. Brain 2015, 138, 2173–2190. [Google Scholar] [CrossRef]
- Colin, E.; Huynh Cong, E.; Mollet, G.; Guichet, A.; Gribouval, O.; Arrondel, C.; Boyer, O.; Daniel, L.; Gubler, M.-C.; Ekinci, Z.; et al. Loss-of-Function Mutations in WDR73 Are Responsible for Microcephaly and Steroid-Resistant Nephrotic Syndrome: Galloway-Mowat Syndrome. Am. J. Hum. Genet. 2014, 95, 637–648. [Google Scholar] [CrossRef]
- Ben-Omran, T.; Fahiminiya, S.; Sorfazlian, N.; Almuriekhi, M.; Nawaz, Z.; Nadaf, J.; Khadija, K.A.; Zaineddin, S.; Kamel, H.; Majewski, J.; et al. Nonsense Mutation in the WDR73 Gene Is Associated with Galloway-Mowat Syndrome. J. Med. Genet. 2015, 52, 381–390. [Google Scholar] [CrossRef]
- Abe, Y.; Matsumoto, S.; Wei, S.; Nezu, K.; Miyoshi, A.; Kito, K.; Ueda, N.; Shigemoto, K.; Hitsumoto, Y.; Nikawa, J.; et al. Cloning and Characterization of a P53-Related Protein Kinase Expressed in Interleukin-2-Activated Cytotoxic T-Cells, Epithelial Tumor Cell Lines, and the Testes. J. Biol. Chem. 2001, 276, 44003–44011. [Google Scholar] [CrossRef]
- Grandi, P.; Dang, T.; Pané, N.; Shevchenko, A.; Mann, M.; Forbes, D.; Hurt, E. Nup93, a Vertebrate Homologue of Yeast Nic96p, Forms a Complex with a Novel 205-kDa Protein and Is Required for Correct Nuclear Pore Assembly. Mol. Biol. Cell 1997, 8, 2017–2038. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.A.; Sadowski, C.E.; Kohl, S.; Lovric, S.; Astrinidis, S.A.; Pabst, W.L.; Gee, H.Y.; Ashraf, S.; Lawson, J.A.; Shril, S.; et al. Mutations in Nuclear Pore Genes NUP93, NUP205 and XPO5 Cause Steroid-Resistant Nephrotic Syndrome. Nat. Genet. 2016, 48, 457–465. [Google Scholar] [CrossRef]
- Boehmer, T.; Enninga, J.; Dales, S.; Blobel, G.; Zhong, H. Depletion of a Single Nucleoporin, Nup107, Prevents the Assembly of a Subset of Nucleoporins into the Nuclear Pore Complex. Proc. Natl. Acad. Sci. USA 2003, 100, 981–985. [Google Scholar] [CrossRef]
- Miyake, N.; Tsukaguchi, H.; Koshimizu, E.; Shono, A.; Matsunaga, S.; Shiina, M.; Mimura, Y.; Imamura, S.; Hirose, T.; Okudela, K.; et al. Biallelic Mutations in Nuclear Pore Complex Subunit NUP107 Cause Early-Childhood-Onset Steroid-Resistant Nephrotic Syndrome. Am. J. Hum. Genet. 2015, 97, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Weinberg-Shukron, A.; Renbaum, P.; Kalifa, R.; Zeligson, S.; Ben-Neriah, Z.; Dreifuss, A.; Abu-Rayyan, A.; Maatuk, N.; Fardian, N.; Rekler, D.; et al. A Mutation in the Nucleoporin-107 Gene Causes XX Gonadal Dysgenesis. J. Clin. Investig. 2015, 125, 4295–4304. [Google Scholar] [CrossRef] [PubMed]
- Marquez, J.; Bhattacharya, D.; Lusk, C.P.; Khokha, M.K. Nucleoporin NUP205 Plays a Critical Role in Cilia and Congenital Disease. Dev. Biol. 2021, 469, 46–53. [Google Scholar] [CrossRef]
- Brownawell, A.M.; Macara, I.G. Exportin-5, a Novel Karyopherin, Mediates Nuclear Export of Double-Stranded RNA Binding Proteins. J. Cell Biol. 2002, 156, 53–64. [Google Scholar] [CrossRef]
- Kozyraki, R.; Kristiansen, M.; Silahtaroglu, A.; Hansen, C.; Jacobsen, C.; Tommerup, N.; Verroust, P.J.; Moestrup, S.K. The Human Intrinsic Factor-Vitamin B12 Receptor, Cubilin: Molecular Characterization and Chromosomal Mapping of the Gene to 10p within the Autosomal Recessive Megaloblastic Anemia (MGA1) Region. Blood 1998, 91, 3593–3600. [Google Scholar] [CrossRef]
- Nykjaer, A.; Fyfe, J.C.; Kozyraki, R.; Leheste, J.R.; Jacobsen, C.; Nielsen, M.S.; Verroust, P.J.; Aminoff, M.; de la Chapelle, A.; Moestrup, S.K.; et al. Cubilin Dysfunction Causes Abnormal Metabolism of the Steroid Hormone 25(OH) Vitamin D(3). Proc. Natl. Acad. Sci. USA 2001, 98, 13895–13900. [Google Scholar] [CrossRef]
- Kalantry, S.; Manning, S.; Haub, O.; Tomihara-Newberger, C.; Lee, H.G.; Fangman, J.; Disteche, C.M.; Manova, K.; Lacy, E. The Amnionless Gene, Essential for Mouse Gastrulation, Encodes a Visceral-Endoderm-Specific Protein with an Extracellular Cysteine-Rich Domain. Nat. Genet. 2001, 27, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Atienza-Manuel, A.; Castillo-Mancho, V.; De Renzis, S.; Culi, J.; Ruiz-Gómez, M. Endocytosis Mediated by an Atypical CUBAM Complex Modulates Slit Diaphragm Dynamics in Nephrocytes. Development 2021, 148, dev199894. [Google Scholar] [CrossRef] [PubMed]
- Gräsbeck, R. Imerslund-Gräsbeck Syndrome (Selective Vitamin B(12) Malabsorption with Proteinuria). Orphanet J. Rare Dis. 2006, 1, 17. [Google Scholar] [CrossRef]
- Christ, A.; Herzog, K.; Willnow, T.E. LRP2, an Auxiliary Receptor That Controls Sonic Hedgehog Signaling in Development and Disease. Dev. Dyn. 2016, 245, 569–579. [Google Scholar] [CrossRef]
- Kantarci, S.; Al-Gazali, L.; Hill, R.S.; Donnai, D.; Black, G.C.M.; Bieth, E.; Chassaing, N.; Lacombe, D.; Devriendt, K.; Teebi, A.; et al. Mutations in LRP2, Which Encodes the Multiligand Receptor Megalin, Cause Donnai-Barrow and Facio-Oculo-Acoustico-Renal Syndromes. Nat. Genet. 2007, 39, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, B.P.; Lundgren, S.; Johansson, M.; Hjälm, G.; Akerström, G.; Gustavsson, I.; Rask, L. In Situ Hybridization Mapping of a 500-kDa Calcium-Sensing Protein Gene (LRP2) to Human Chromosome Region 2q31-->q32.1 and Porcine Chromosome Region 15q22-->q24. Cytogenet. Cell Genet. 1995, 71, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Ault, B.H. Factor H and the Pathogenesis of Renal Diseases. Pediatr. Nephrol. 2000, 14, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Ying, L.; Katz, Y.; Schlesinger, M.; Carmi, R.; Shalev, H.; Haider, N.; Beck, G.; Sheffield, V.C.; Landau, D. Complement Factor H Gene Mutation Associated with Autosomal Recessive Atypical Hemolytic Uremic Syndrome. Am. J. Hum. Genet. 1999, 65, 1538–1546. [Google Scholar] [CrossRef]
- Zipfel, P.F.; Wiech, T.; Stea, E.D.; Skerka, C. CFHR Gene Variations Provide Insights in the Pathogenesis of the Kidney Diseases Atypical Hemolytic Uremic Syndrome and C3 Glomerulopathy. J. Am. Soc. Nephrol. 2020, 31, 241–256. [Google Scholar] [CrossRef]
- Kömpf, J.; Luckenbach, C.; Kloor, D.; Schunter, F.; Wernet, P.; Ritter, H. Human Factor H (Beta 1H-Globulin): Linkage Analysis. Hum. Genet. 1988, 79, 181–182. [Google Scholar] [CrossRef]
- Ozaltin, F.; Li, B.; Rauhauser, A.; An, S.-W.; Soylemezoglu, O.; Gonul, I.I.; Taskiran, E.Z.; Ibsirlioglu, T.; Korkmaz, E.; Bilginer, Y.; et al. DGKE Variants Cause a Glomerular Microangiopathy That Mimics Membranoproliferative GN. J. Am. Soc. Nephrol. 2013, 24, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.C.; Price, J.A.; Bobby, P.L.; Pettenati, M.J.; Shashi, V.; Von Kap Herr, C.; Van Dyke, T.E. Cytogenetic Assignment and Physical Mapping of the Human DGKE Gene to Chromosome 17q22. Genomics 1999, 56, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Matthijs, G.; Schollen, E.; Pardon, E.; Veiga-Da-Cunha, M.; Jaeken, J.; Cassiman, J.J.; Van Schaftingen, E. Mutations in PMM2, a Phosphomannomutase Gene on Chromosome 16p13, in Carbohydrate-Deficient Glycoprotein Type I Syndrome (Jaeken Syndrome). Nat. Genet. 1997, 16, 88–92. [Google Scholar] [CrossRef]
- Thakor, J.M.; Parmar, G.; Mistry, K.N.; Gang, S.; Rank, D.N.; Joshi, C.G. Mutational Landscape of TRPC6, WT1, LMX1B, APOL1, PTPRO, PMM2, LAMB2 and WT1 Genes Associated with Steroid Resistant Nephrotic Syndrome. Mol. Biol. Rep. 2021, 48, 7193–7201. [Google Scholar] [CrossRef] [PubMed]
- Banderali, G.; Salvatici, E.; Rovelli, V.; Jaeken, J. PMM2-CDG and Nephrotic Syndrome: A Case Report. Clin. Case Rep. 2022, 10, e05347. [Google Scholar] [CrossRef]
- Ozaltin, F.; Ibsirlioglu, T.; Taskiran, E.Z.; Baydar, D.E.; Kaymaz, F.; Buyukcelik, M.; Kilic, B.D.; Balat, A.; Iatropoulos, P.; Asan, E.; et al. Disruption of PTPRO Causes Childhood-Onset Nephrotic Syndrome. Am. J. Hum. Genet. 2011, 89, 139–147. [Google Scholar] [CrossRef]
- Dai, W.; Xiang, W.; Han, L.; Yuan, Z.; Wang, R.; Ma, Y.; Yang, Y.; Cai, S.; Xu, Y.; Mo, S.; et al. PTPRO Represses Colorectal Cancer Tumorigenesis and Progression by Reprogramming Fatty Acid Metabolism. Cancer Commun. 2022, 42, 848–867. [Google Scholar] [CrossRef]
- Heybrock, S.; Kanerva, K.; Meng, Y.; Ing, C.; Liang, A.; Xiong, Z.-J.; Weng, X.; Ah Kim, Y.; Collins, R.; Trimble, W.; et al. Lysosomal Integral Membrane Protein-2 (LIMP-2/SCARB2) Is Involved in Lysosomal Cholesterol Export. Nat. Commun. 2019, 10, 3521. [Google Scholar] [CrossRef]
- Berkovic, S.F.; Dibbens, L.M.; Oshlack, A.; Silver, J.D.; Katerelos, M.; Vears, D.F.; Lüllmann-Rauch, R.; Blanz, J.; Zhang, K.W.; Stankovich, J.; et al. Array-Based Gene Discovery with Three Unrelated Subjects Shows SCARB2/LIMP-2 Deficiency Causes Myoclonus Epilepsy and Glomerulosclerosis. Am. J. Hum. Genet. 2008, 82, 673–684. [Google Scholar] [CrossRef]
- Badhwar, A.; Berkovic, S.F.; Dowling, J.P.; Gonzales, M.; Narayanan, S.; Brodtmann, A.; Berzen, L.; Caviness, J.; Trenkwalder, C.; Winkelmann, J.; et al. Action Myoclonus-Renal Failure Syndrome: Characterization of a Unique Cerebro-Renal Disorder. Brain 2004, 127, 2173–2182. [Google Scholar] [CrossRef]
- Xiao, C.; Ahn, H.; Kibrom, S.; Toro, C. SCARB2-Related Action Myoclonus—Renal Failure Syndrome. In GeneReviews®; Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Freije, J.M.; Blay, P.; Pendás, A.M.; Cadiñanos, J.; Crespo, P.; López-Otín, C. Identification and Chromosomal Location of Two Human Genes Encoding Enzymes Potentially Involved in Proteolytic Maturation of Farnesylated Proteins. Genomics 1999, 58, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.K.; Fryns, J.-P.; Auchus, R.J.; Garg, A. Zinc Metalloproteinase, ZMPSTE24, Is Mutated in Mandibuloacral Dysplasia. Hum. Mol. Genet. 2003, 12, 1995–2001. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Thiel, C.; Lübbehusen, J.; Dorland, B.; de Koning, T.; von Figura, K.; Lehle, L.; Körner, C. Deficiency of GDP-Man:GlcNAc2-PP-Dolichol Mannosyltransferase Causes Congenital Disorder of Glycosylation Type Ik. Am. J. Hum. Genet. 2004, 74, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Harshman, L.A.; Ng, B.G.; Freeze, H.H.; Trapane, P.; Dolezal, A.; Brophy, P.D.; Brumbaugh, J.E. Congenital Nephrotic Syndrome in an Infant with ALG1-Congenital Disorder of Glycosylation. Pediatr. Int. 2016, 58, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhao, Y.; Wu, B.; Shu, J.; Yan, D.; Li, D.; Yu, X.; Cai, C. A Novel Variant in ALG1 Gene Associated with Congenital Disorder of Glycosylation: A Case Report and Short Literature Review. Mol. Genet. Genomic Med. 2023, 11, e2197. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Chen, Z.; Choi, W.-I.; Gee, H.Y.; Hildebrandt, F.; Zhou, W. Loss of Epithelial Membrane Protein 2 Aggravates Podocyte Injury via Upregulation of Caveolin-1. J. Am. Soc. Nephrol. 2016, 27, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.Y.; Ashraf, S.; Wan, X.; Vega-Warner, V.; Esteve-Rudd, J.; Lovric, S.; Fang, H.; Hurd, T.W.; Sadowski, C.E.; Allen, S.J.; et al. Mutations in EMP2 Cause Childhood-Onset Nephrotic Syndrome. Am. J. Hum. Genet. 2014, 94, 884–890. [Google Scholar] [CrossRef]
- Liehr, T.; Kuhlenbäumer, G.; Wulf, P.; Taylor, V.; Suter, U.; Van Broeckhoven, C.; Lupski, J.R.; Claussen, U.; Rautenstrauss, B. Regional Localization of the Human Epithelial Membrane Protein Genes 1, 2, and 3 (EMP1, EMP2, EMP3) to 12p12.3, 16p13.2, and 19q13.3. Genomics 1999, 58, 106–108. [Google Scholar] [CrossRef]
- Tran, P.V.; Haycraft, C.J.; Besschetnova, T.Y.; Turbe-Doan, A.; Stottmann, R.W.; Herron, B.J.; Chesebro, A.L.; Qiu, H.; Scherz, P.J.; Shah, J.V.; et al. THM1 Negatively Modulates Mouse Sonic Hedgehog Signal Transduction and Affects Retrograde Intraflagellar Transport in Cilia. Nat. Genet. 2008, 40, 403–410. [Google Scholar] [CrossRef]
- Davis, E.E.; Zhang, Q.; Liu, Q.; Diplas, B.H.; Davey, L.M.; Hartley, J.; Stoetzel, C.; Szymanska, K.; Ramaswami, G.; Logan, C.V.; et al. TTC21B Contributes Both Causal and Modifying Alleles across the Ciliopathy Spectrum. Nat. Genet. 2011, 43, 189–196. [Google Scholar] [CrossRef]
- Awazu, M.; Yamada, M.; Asada, N.; Hashiguchi, A.; Kosaki, K.; Matsumura, K. A Girl with a Mutation of the Ciliary Gene CC2D2A Presenting with FSGS and Nephronophthisis. CEN Case Rep. 2022, 11, 116–119. [Google Scholar] [CrossRef]
- McInerney-Leo, A.M.; Harris, J.E.; Leo, P.J.; Marshall, M.S.; Gardiner, B.; Kinning, E.; Leong, H.Y.; McKenzie, F.; Ong, W.P.; Vodopiutz, J.; et al. Whole Exome Sequencing Is an Efficient, Sensitive and Specific Method for Determining the Genetic Cause of Short-Rib Thoracic Dystrophies. Clin. Genet. 2015, 88, 550–557. [Google Scholar] [CrossRef]
- Prasad, R.; Hadjidemetriou, I.; Maharaj, A.; Meimaridou, E.; Buonocore, F.; Saleem, M.; Hurcombe, J.; Bierzynska, A.; Barbagelata, E.; Bergadá, I.; et al. Sphingosine-1-Phosphate Lyase Mutations Cause Primary Adrenal Insufficiency and Steroid-Resistant Nephrotic Syndrome. J. Clin. Investig. 2017, 127, 942–953. [Google Scholar] [CrossRef]
- Zhou, J.; Saba, J.D. Identification of the First Mammalian Sphingosine Phosphate Lyase Gene and Its Functional Expression in Yeast. Biochem. Biophys. Res. Commun. 1998, 242, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Tarshish, P.; Tobin, J.N.; Bernstein, J.; Edelmann, C.M. Prognostic Significance of the Early Course of Minimal Change Nephrotic Syndrome: Report of the International Study of Kidney Disease in Children. J. Am. Soc. Nephrol. 1997, 8, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, A.C.Q.; Oliveira, E.A.; Fróes, B.P.; Faria, L.D.C.; Pinto, J.S.; Nogueira, M.M.I.; Lima, G.O.; Resende, P.I.; Assis, N.S.; Simões E Silva, A.C.; et al. A Predictive Model of Progressive Chronic Kidney Disease in Idiopathic Nephrotic Syndrome. Pediatr. Nephrol. 2015, 30, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Poggio, E.D.; McClelland, R.L.; Blank, K.N.; Hansen, S.; Bansal, S.; Bomback, A.S.; Canetta, P.A.; Khairallah, P.; Kiryluk, K.; Lecker, S.H.; et al. Systematic Review and Meta-Analysis of Native Kidney Biopsy Complications. Clin. J. Am. Soc. Nephrol. 2020, 15, 1595–1602. [Google Scholar] [CrossRef] [PubMed]
- Tøndel, C.; Vikse, B.E.; Bostad, L.; Svarstad, E. Safety and Complications of Percutaneous Kidney Biopsies in 715 Children and 8573 Adults in Norway 1988–2010. Clin. J. Am. Soc. Nephrol. CJASN 2012, 7, 1591. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, C.; Yu, H.; Ding, M.; Zhang, C.; Lu, X.; Zhang, C.-Y.; Zhang, C. Increased Urinary Exosomal microRNAs in Children with Idiopathic Nephrotic Syndrome. EBioMedicine 2019, 39, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.; Sun, F.; Yu, P.-F.; Li, J.-X.; Yuan, D.; Chang, J.; Lin, S.-H. Differential microRNA Expression in the Serum of Patients with Nephrotic Syndrome and Clinical Correlation Analysis. Int. J. Clin. Exp. Pathol. 2015, 8, 7282–7286. [Google Scholar]
- Bayomy, N.R.; Abo Alfottoh, W.M.; Ali Eldeep, S.A.; Ibrahim Mabrouk Mersal, A.M.S.; Abd El-Bary, H.M.A.; Abd El Gayed, E.M. Mir-142-5p as an Indicator of Autoimmune Processes in Childhood Idiopathic Nephrotic Syndrome and as a Part of MicroRNAs Expression Panels for Its Diagnosis and Prediction of Response to Steroid Treatment. Mol. Immunol. 2022, 141, 21–32. [Google Scholar] [CrossRef]
- Zhang, Y.-R.; Wu, Y.-F.; Wang, H.; Lin, X.-M.; Zhang, X.-M. Role of microRNA-17-5p in the pathogenesis of pediatric nephrotic syndrome and related mechanisms. Zhongguo Dang Dai Er Ke Za Zhi 2020, 22, 958–963. [Google Scholar] [CrossRef]
- Traum, A.Z.; Schachter, A.D. Proteomic Analysis in Pediatric Renal Disease. Semin. Nephrol. 2007, 27, 652–657. [Google Scholar] [CrossRef]
- Decramer, S.; de Peredo, A.G.; Breuil, B.; Mischak, H.; Monsarrat, B.; Bascands, J.-L.; Schanstra, J.P. Urine in Clinical Proteomics. Mol. Cell. Proteom. 2008, 7, 1850–1862. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.J. Glomerular Disease: Gain-of-Glycosylation Mutation in ITGA3 Causes Nephrotic Syndrome. Nat. Rev. Nephrol. 2013, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Piyaphanee, N.; Ma, Q.; Kremen, O.; Czech, K.; Greis, K.; Mitsnefes, M.; Devarajan, P.; Bennett, M.R. Discovery and Initial Validation of α 1-B Glycoprotein Fragmentation as a Differential Urinary Biomarker in Pediatric Steroid-Resistant Nephrotic Syndrome. Proteomics Clin. Appl. 2011, 5, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Pleasant, L.; Haffner, C.; Ma, Q.; Haffey, W.D.; Ying, J.; Wagner, M.; Greis, K.D.; Devarajan, P. A Novel Biomarker Panel to Identify Steroid Resistance in Childhood Idiopathic Nephrotic Syndrome. Biomark. Insights 2017, 12, 1177271917695832. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.M.; Elbehidy, R.M.; Abdelhalim, H.S.; Amr, G.E. Soluble Interleukine-2 Receptor and MDR1 Gene Expression Levels as Inflammatory Biomarkers for Prediction of Steroid Response in Children with Nephrotic Syndrome. Iran. J. Kidney Dis. 2011, 5, 154–161. [Google Scholar]
- Peng, Z.; Mao, J.; Chen, X.; Cai, F.; Gu, W.; Fu, H.; Shen, H.; Wang, J.; Jin, X.; Zhu, X.; et al. Serum suPAR Levels Help Differentiate Steroid Resistance from Steroid-Sensitive Nephrotic Syndrome in Children. Pediatr. Nephrol. 2015, 30, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Simões e Silva, A.C.; Pereira, A.B.; Teixeira, M.M.; Teixeira, A.L. Chemokines as Potential Markers in Pediatric Renal Diseases. Dis. Markers 2014, 2014, 278715. [Google Scholar] [CrossRef]
- Segerer, S.; Alpers, C.E. Chemokines and Chemokine Receptors in Renal Pathology. Curr. Opin. Nephrol. Hypertens. 2003, 12, 243–249. [Google Scholar] [CrossRef]
- Araya, C.; Diaz, L.; Wasserfall, C.; Atkinson, M.; Mu, W.; Johnson, R.; Garin, E. T Regulatory Cell Function in Idiopathic Minimal Lesion Nephrotic Syndrome. Pediatr. Nephrol. 2009, 24, 1691–1698. [Google Scholar] [CrossRef]
- Woroniecki, R.P.; Shatat, I.F.; Supe, K.; Du, Z.; Kaskel, F.J. Urinary Cytokines and Steroid Responsiveness in Idiopathic Nephrotic Syndrome of Childhood. Am. J. Nephrol. 2008, 28, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Vianna, H.R.; Soares, C.M.B.M.; Silveira, K.D.; Elmiro, G.S.; Mendes, P.M.; de Sousa Tavares, M.; Teixeira, M.M.; Miranda, D.M.; Simões E Silva, A.C. Cytokines in Chronic Kidney Disease: Potential Link of MCP-1 and Dyslipidemia in Glomerular Diseases. Pediatr. Nephrol. 2013, 28, 463–469. [Google Scholar] [CrossRef]
- Aizawa, T.; Imaizumi, T.; Tsuruga, K.; Watanabe, S.; Yoshida, H.; Kumagai, N.; Ito, E.; Tanaka, H. Urinary Fractalkine and Monocyte Chemoattractant Protein-1 as Possible Predictors of Disease Activity of Childhood Glomerulonephritis. Tohoku J. Exp. Med. 2013, 231, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Chebotareva, N.; Vinogradov, A.; Cao, V.; Gindis, A.; Berns, A.; Alentov, I.; Sergeeva, N. Serum Levels of Plasminogen Activator Urokinase Receptor and Cardiotrophin-like Cytokine Factor 1 in Patients with Nephrotic Syndrome. Clin. Nephrol. 2022, 97, 103–110. [Google Scholar] [CrossRef]
- Bai, Y.; Liu, W.; Guo, Q.; Zou, Y. Screening for Urinary Biomarkers of Steroid-Resistant Nephrotic Syndrome in Children. Exp. Ther. Med. 2013, 5, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Suresh, C.P.; Saha, A.; Kaur, M.; Kumar, R.; Dubey, N.K.; Basak, T.; Tanwar, V.S.; Bhardwaj, G.; Sengupta, S.; Batra, V.V.; et al. Differentially Expressed Urinary Biomarkers in Children with Idiopathic Nephrotic Syndrome. Clin. Exp. Nephrol. 2016, 20, 273–283. [Google Scholar] [CrossRef]
- Agrawal, S.; Merchant, M.L.; Kino, J.; Li, M.; Wilkey, D.W.; Gaweda, A.E.; Brier, M.E.; Chanley, M.A.; Gooding, J.R.; Sumner, S.J.; et al. Predicting and Defining Steroid Resistance in Pediatric Nephrotic Syndrome Using Plasma Proteomics. Kidney Int. Rep. 2020, 5, 66–80. [Google Scholar] [CrossRef]
- Barragry, J.M.; France, M.W.; Carter, N.D.; Auton, J.A.; Beer, M.; Boucher, B.J.; Cohen, R.D. Vitamin-D Metabolism in Nephrotic Syndrome. Lancet 1977, 2, 629–632. [Google Scholar] [CrossRef]
- Selewski, D.T.; Chen, A.; Shatat, I.F.; Pais, P.; Greenbaum, L.A.; Geier, P.; Nelson, R.D.; Kiessling, S.G.; Brophy, P.D.; Quiroga, A.; et al. Vitamin D in Incident Nephrotic Syndrome: A Midwest Pediatric Nephrology Consortium Study. Pediatr. Nephrol. 2016, 31, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Pordal, A.; Haffner, C.; Pleasant, L.; Ma, Q.; Devarajan, P. Urinary Vitamin D-Binding Protein as a Biomarker of Steroid-Resistant Nephrotic Syndrome. Biomark. Insights 2016, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cowland, J.B.; Borregaard, N. Molecular Characterization and Pattern of Tissue Expression of the Gene for Neutrophil Gelatinase-Associated Lipocalin from Humans. Genomics 1997, 45, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Ma, Q.; Prada, A.; Mitsnefes, M.; Zahedi, K.; Yang, J.; Barasch, J.; Devarajan, P. Identification of Neutrophil Gelatinase-Associated Lipocalin as a Novel Early Urinary Biomarker for Ischemic Renal Injury. J. Am. Soc. Nephrol. 2003, 14, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Bolignano, D.; Lacquaniti, A.; Coppolino, G.; Donato, V.; Campo, S.; Fazio, M.R.; Nicocia, G.; Buemi, M. Neutrophil Gelatinase-Associated Lipocalin (NGAL) and Progression of Chronic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Nickolas, T.L.; Forster, C.S.; Sise, M.E.; Barasch, N.; Solá-Del Valle, D.; Viltard, M.; Buchen, C.; Kupferman, S.; Carnevali, M.L.; Bennett, M.; et al. NGAL (Lcn2) Monomer Is Associated with Tubulointerstitial Damage in Chronic Kidney Disease. Kidney Int. 2012, 82, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Piyaphanee, N.; Czech, K.; Mitsnefes, M.; Devarajan, P. NGAL Distinguishes Steroid Sensitivity in Idiopathic Nephrotic Syndrome. Pediatr. Nephrol. 2012, 27, 807–812. [Google Scholar] [CrossRef]
- Dubin, R.F.; Rhee, E.P. Proteomics and Metabolomics in Kidney Disease, Including Insights into Etiology, Treatment, and Prevention. Clin. J. Am. Soc. Nephrol. 2020, 15, 404–411. [Google Scholar] [CrossRef]
- Pereira, P.R.; Carrageta, D.F.; Oliveira, P.F.; Rodrigues, A.; Alves, M.G.; Monteiro, M.P. Metabolomics as a Tool for the Early Diagnosis and Prognosis of Diabetic Kidney Disease. Med. Res. Rev. 2022, 42, 1518–1544. [Google Scholar] [CrossRef]
- Sedic, M.; Gethings, L.A.; Vissers, J.P.C.; Shockcor, J.P.; McDonald, S.; Vasieva, O.; Lemac, M.; Langridge, J.I.; Batinić, D.; Pavelić, S.K. Label-Free Mass Spectrometric Profiling of Urinary Proteins and Metabolites from Paediatric Idiopathic Nephrotic Syndrome. Biochem. Biophys. Res. Commun. 2014, 452, 21–26. [Google Scholar] [CrossRef]
- Gooding, J.R.; Agrawal, S.; McRitchie, S.; Acuff, Z.; Merchant, M.L.; Klein, J.B.; Smoyer, W.E.; Sumner, S.J. Midwest Pediatric Nephrology Consortium Predicting and Defining Steroid Resistance in Pediatric Nephrotic Syndrome Using Plasma Metabolomics. Kidney Int. Rep. 2020, 5, 81–93. [Google Scholar] [CrossRef]
- Stone, H.; Magella, B.; Bennett, M.R. The Search for Biomarkers to Aid in Diagnosis, Differentiation, and Prognosis of Childhood Idiopathic Nephrotic Syndrome. Front. Pediatr. 2019, 7, 404. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.K.; Arif, M.; Amjad, N. A Histopathological Outlook on Nephrotic Syndrome: A Pediatric Perspective. Indian. J. Nephrol. 2016, 26, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S. Mechanisms and Consequences of TGF-ß Overexpression by Podocytes in Progressive Podocyte Disease. Cell Tissue Res. 2012, 347, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.Z.; Youssef, D.M.; El-Shal, A.S.; Abdelsalam, A.A. Transforming Growth Factor-Beta in Nephrotic Syndrome and Its Correlation with Albuminemia and Hyperlipidemia. Biochem. Lett. 2019, 15, 186–195. [Google Scholar] [CrossRef]
- Froes, B.P.; de Almeida Araújo, S.; Bambirra, E.A.; Oliveira, E.A.; Simões E Silva, A.C.; Pinheiro, S.V.B. Is CD44 in Glomerular Parietal Epithelial Cells a Pathological Marker of Renal Function Deterioration in Primary Focal Segmental Glomerulosclerosis? Pediatr. Nephrol. 2017, 32, 2165–2169. [Google Scholar] [CrossRef]
- Guimarães, F.T.L.; Ferreira, R.N.; Brito-Melo, G.E.A.; Rocha-Vieira, E.; de Pereira, W.F.; Pinheiro, S.V.B.; Miranda, A.S.; Simões E Silva, A.C. Pediatric Patients With Steroid-Sensitive Nephrotic Syndrome Have Higher Expression of T Regulatory Lymphocytes in Comparison to Steroid-Resistant Disease. Front. Pediatr. 2019, 7, 114. [Google Scholar] [CrossRef]
- Gonzalez Guerrico, A.M.; Lieske, J.; Klee, G.; Kumar, S.; Lopez-Baez, V.; Wright, A.M.; Bobart, S.; Shevell, D.; Maldonado, M.; Troost, J.P.; et al. Urinary CD80 Discriminates Among Glomerular Disease Types and Reflects Disease Activity. Kidney Int. Rep. 2020, 5, 2021–2031. [Google Scholar] [CrossRef]
- Sansom, D.M.; Manzotti, C.N.; Zheng, Y. What’s the Difference between CD80 and CD86? Trends Immunol. 2003, 24, 314–319. [Google Scholar] [CrossRef]
- Eroglu, F.K.; Orhan, D.; İnözü, M.; Duzova, A.; Gulhan, B.; Ozaltin, F.; Topaloglu, R. CD80 Expression and Infiltrating Regulatory T Cells in Idiopathic Nephrotic Syndrome of Childhood. Pediatr. Int. 2019, 61, 1250–1256. [Google Scholar] [CrossRef]
- Eroglu, F.K.; Yazar, V.; Guler, U.; Yıldırım, M.; Yildirim, T.; Gungor, T.; Celikkaya, E.; Karakaya, D.; Turay, N.; Ciftci Dede, E.; et al. Circulating extracellular vesicles of patients with steroid-sensitive nephrotic syndrome have higher RAC1 and induce recapitulation of nephrotic syndrome phenotype in podocytes. Am. J. Physiol. Renal Physiol. 2021, 321, F659–F673. [Google Scholar] [CrossRef] [PubMed]
- Pekkucuksen, N.T.; Liu, L.P.; Aly, R.; Shoemaker, L.R.; Alli, A.A. Extracellular vesicles from focal segmental glomerulosclerosis pediatric patients induce STAT3 activation and mesangial cell proliferation. PLoS ONE 2022, 17, e0274598. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Candiano, G.; Angeletti, A.; Lugani, F.; Panfoli, I. Extracellular Vesicles as Source of Biomarkers in Glomerulonephritis. Int. J. Mol. Sci. 2023, 24, 13894. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the glomerular slit diaphragm, a pivotal structure within the glomerulus that is essential for the filtration barrier’s integrity. Positioned between podocyte foot processes and the glomerular basement membrane (GBM), this intricate assembly involves a network of molecules crucial for maintaining structural and functional integrity. Key components include transmembrane proteins like nephrin (a structural protein forming the backbone of the slit diaphragm), podocin (interacting with nephrin, crucial for slit diaphragm function), NEPH1/2/3 (members of the neph family proteins contributing to slit diaphragm molecular architecture), Cadherin 3 (involved in cell adhesion within podocytes), and podoplanin (a transmembrane protein expressed in podocytes). Additionally, molecules such as laminin (a component of the GBM providing structural support) and dystroglycan complex proteins (α-DG, β-DG) (linking the cytoskeleton to the extracellular matrix) are integral to the GBM. The schema also highlights other essential players, including utrophin (participating in cytoskeletal organization), synaptopodin (involved in actin cytoskeleton regulation), ezrin (linking actin filaments to the plasma membrane), NHERF-2 (a scaffold protein interacting with nephrin and podocin), podocalyxin (a sialoprotein contributing to the glycocalyx), FAT1 (a cell adhesion and signaling protein), ZO-1 (a tight junction-associated protein), TRCP6 (an ion channel implicated in slit diaphragm function), actin (a cytoskeletal component crucial for podocyte structure), and megalin (a receptor involved in protein reabsorption in the proximal tubule).
Figure 1.
Schematic representation of the glomerular slit diaphragm, a pivotal structure within the glomerulus that is essential for the filtration barrier’s integrity. Positioned between podocyte foot processes and the glomerular basement membrane (GBM), this intricate assembly involves a network of molecules crucial for maintaining structural and functional integrity. Key components include transmembrane proteins like nephrin (a structural protein forming the backbone of the slit diaphragm), podocin (interacting with nephrin, crucial for slit diaphragm function), NEPH1/2/3 (members of the neph family proteins contributing to slit diaphragm molecular architecture), Cadherin 3 (involved in cell adhesion within podocytes), and podoplanin (a transmembrane protein expressed in podocytes). Additionally, molecules such as laminin (a component of the GBM providing structural support) and dystroglycan complex proteins (α-DG, β-DG) (linking the cytoskeleton to the extracellular matrix) are integral to the GBM. The schema also highlights other essential players, including utrophin (participating in cytoskeletal organization), synaptopodin (involved in actin cytoskeleton regulation), ezrin (linking actin filaments to the plasma membrane), NHERF-2 (a scaffold protein interacting with nephrin and podocin), podocalyxin (a sialoprotein contributing to the glycocalyx), FAT1 (a cell adhesion and signaling protein), ZO-1 (a tight junction-associated protein), TRCP6 (an ion channel implicated in slit diaphragm function), actin (a cytoskeletal component crucial for podocyte structure), and megalin (a receptor involved in protein reabsorption in the proximal tubule).

Figure 2.
Histological comparison of Minimal Change Nephrotic Syndrome (MCNS) and Focal Segmental Glomerulosclerosis (FSGS). (A) Representative histological slide of kidney tissue from a patient diagnosed with Minimal Change Nephrotic Syndrome (MCNS). The glomerulus exhibits minimal structural alterations, characterized by a lack of significant changes in the glomerular basement membrane, mesangium, and podocytes (Jones silver stain). Electronic microscopy can reveal diffuse effacement of podocyte foot processes, a hallmark feature of MCNS. (B) In contrast, kidney tissue from a patient diagnosed with Focal Segmental Glomerulosclerosis (FSGS) is depicted in this section (h&E staining). The histological examination reveals focal and segmental areas of sclerosis within the glomerulus, indicating regions of glomerular injury and scarring. The presence of hyalinosis, adhesions, and segmental obliteration of capillary lumina is evident, reflecting the characteristic histopathological changes associated with FSGS. Both micrographs were captured at a magnification of 400× (personal archive of the authors).
Figure 2.
Histological comparison of Minimal Change Nephrotic Syndrome (MCNS) and Focal Segmental Glomerulosclerosis (FSGS). (A) Representative histological slide of kidney tissue from a patient diagnosed with Minimal Change Nephrotic Syndrome (MCNS). The glomerulus exhibits minimal structural alterations, characterized by a lack of significant changes in the glomerular basement membrane, mesangium, and podocytes (Jones silver stain). Electronic microscopy can reveal diffuse effacement of podocyte foot processes, a hallmark feature of MCNS. (B) In contrast, kidney tissue from a patient diagnosed with Focal Segmental Glomerulosclerosis (FSGS) is depicted in this section (h&E staining). The histological examination reveals focal and segmental areas of sclerosis within the glomerulus, indicating regions of glomerular injury and scarring. The presence of hyalinosis, adhesions, and segmental obliteration of capillary lumina is evident, reflecting the characteristic histopathological changes associated with FSGS. Both micrographs were captured at a magnification of 400× (personal archive of the authors).

Figure 3.
Histological examination of Collapsing Focal Segmental Glomerulosclerosis (cFSGS). (A) Low magnification view (100×) of kidney tissue depicting the characteristic features of collapsing FSGS (cFSGS). The image reveals widespread collapse and obliteration of glomerular capillaries, accompanied by prominent hyperplasia and hypertrophy of podocytes. The collapsing variant is characterized by the involvement of the entire glomerulus, leading to marked changes in the glomerular architecture. (B) Higher magnification (400×) of a representative section illustrating the detailed histopathological changes in cFSGS. At this level of magnification, the severe podocyte hyperplasia, segmental capillary collapse, and visceral epithelial cell hyperplasia become more evident. The presence of prominent protein resorption droplets within dilated tubules is also notable, highlighting the severity of glomerular injury in cFSGS. Both micrographs were obtained using Jones silver staining (personal archive of the authors).
Figure 3.
Histological examination of Collapsing Focal Segmental Glomerulosclerosis (cFSGS). (A) Low magnification view (100×) of kidney tissue depicting the characteristic features of collapsing FSGS (cFSGS). The image reveals widespread collapse and obliteration of glomerular capillaries, accompanied by prominent hyperplasia and hypertrophy of podocytes. The collapsing variant is characterized by the involvement of the entire glomerulus, leading to marked changes in the glomerular architecture. (B) Higher magnification (400×) of a representative section illustrating the detailed histopathological changes in cFSGS. At this level of magnification, the severe podocyte hyperplasia, segmental capillary collapse, and visceral epithelial cell hyperplasia become more evident. The presence of prominent protein resorption droplets within dilated tubules is also notable, highlighting the severity of glomerular injury in cFSGS. Both micrographs were obtained using Jones silver staining (personal archive of the authors).

Table 2.
Summary of genes related to the development of childhood nephrotic syndrome.
| Gene | Function/Description | Mutations and Associated Conditions | Chromosome Location | Mode of Inheritance | References |
|---|---|---|---|---|---|
| Slit-diaphragm-associated mutations | |||||
| CD2AP | Acts as a bridge between the SD and the actin cytoskeleton. Lack of expression causes mesangial cell proliferation, glomerulosclerosis, and NS. | Mutations alter the SD structure. CD2AP-/- mice develop severe NS. | 6p12.3 | Autosomal Recessive and Dominant | [64,65,66] |
| NPHS1 | Encodes nephrin. Mutations account for 40–60% of infants with congenital NS. Malfunction leads to massive protein loss. | Mutations associated with Finnish-type nephrotic syndrome. | 19q13.12 | Autosomal Recessive | [67,68,69] |
| NPHS2 | Encodes podocin. Plays a role in recruiting nephrin to the SD. Mutations lead to congenital/infantile NS and SRNS. | Mutations lead to childhood- or adult-onset SRNS. | 1q25.2 | Autosomal Recessive | [70,71] |
| CRB2 | Encodes the Crumbs cell polarity complex protein | Mutations lead to SRNS with FSGS. | 9q33.3 | Autosomal Recessive | [72,73,74] |
| FAT1 | Encodes the tumor suppressor of the cadherin superfamily. | Mutations lead to a combination of SRNS, tubular ectasia, hematuria, and neurological involvement. | 4q35.2 | Autosomal Recessive | [75,76,77,78] |
| PLCE1 | Encodes the Phospholipase C epsilon 1 (PLCε1) protein. | Mutations in PLCE1 are responsible for a major part of cases of Mesangial Sclerosis. | 10q23.33 | Autosomal Recessive | [79,80] |
| TRPC6 | Encodes the Transient Receptor Potential Cation Channel Subfamily C member 6 receptor. | Mutations lead to late-onset FSGS. | 11q22.1 | Autosomal Dominant | [81,82,83,84] |
| Actin-cytoskeleton-associated mutations | |||||
| ACTN4 | Encodes alpha-actin 4, an actin-binding protein in the FPs of podocytes. | Mutations associated with adult-onset SRNS with FSGS. | 19q13.2 | Autosomal Recessive | [85,86,87] |
| ANLN | Encodes anilin, an actin-binding protein. Mutations cause a reduction in CD2AP binding, dysregulating signaling in podocytes. | Mutations lead to disarrangement in the SD. | 7p14.2 | Autosomal Dominant | [88,89] |
| ARHGAP24 | Encodes the Rho GTPase-activating protein 24. | Mutations associated with familial SRNS in the second and third decades of life. | 4q21.3 | Autosomal Dominant | [90,91] |
| ARHGDIA | Encodes the RhoGDP dissociation inhibitor α. | Mutations associated with congenital NS or SRNS within the first 2 years. | 17q25.3 | Autosomal Recessive | [92,93] |
| INF2 | Encodes inverted formin 2 and regulates actin polymerization. Mutations related to Charcot–Marie–Tooth disease and FSGS. | Mutations related to Charcot–Marie–Tooth disease and FSGS. | 14q32.33 | Autosomal Dominant | [94,95] |
| MYO1E | Encodes non-muscular myosin 1E, an actin-binding molecular motor in the FPs of podocytes. | Mutations lead to SRNS with focal thickening, disorganization, and multilamination of GBM. | 15q22.2 | Autosomal Recessive | [96,97,98] |
| KANK1 | Encodes kidney ankyrin repeat-containing protein 1. | Mutations cause congenital and early childhood onset NS. | 9p24.3 | Autosomal Recessive | [99,100] |
| KANK2 | Encodes kidney ankyrin repeat-containing protein 2. | Mutations cause congenital and early childhood onset NS. | 19p13.2 | Autosomal Recessive | [99,101] |
| KANK4 | Encodes kidney ankyrin repeat-containing protein 4. | Mutations cause congenital and early childhood onset NS. | 1p31.3 | Autosomal Recessive | [99,102] |
| Mitochondrial protein mutations | |||||
| ADCK4 | Has a role in the synthesis of Coenzyme Q10 (CoQ10), a lipid-soluble ubiquinone. | Mutations cause SRNS. | 19q13.1 | Autosomal Recessive | [103,104,105] |
| COQ2 | Encodes parahydroxybenzoate-polyprenyltransferase, involved in the biosynthesis of ubiquinone CoQ10. | Mutations lead to CoQ10 deficiency, resulting in SRNS. | 4q21.23 | Autosomal Recessive | [106,107] |
| COQ6 | The product is a mono-oxygenase required for CoQ10 biosynthesis. Mutations lead to primary CoQ10 deficiency. | Mutations result in primary CoQ10 deficiency, leading to SRNS. | 14q24.3 | Autosomal Recessive | [108,109,110,111] |
| PDSS2 | Encodes decaprenyl diphosphate synthase, subunit 2, a component of a heterotetrameric decaprenyl synthase. | Mutations lead to primary CoQ10 deficiency, resulting in SRNS. | 6q21 | Autosomal Recessive | [112,113,114] |
| Glomerular-basement-membrane-related mutations | |||||
| COL4A3 | Encodes Collagen Type IV alpha-3 chain. | Mutations related to Alport syndrome type 2 and autosomal dominant or recessive modes of inheritance for benign familial hematuria. | 2q36.3 | Autosomal Dominant (Alport syndrome type 3), Autosomal Recessive (Alport syndrome type 2) | [115,116,117,118,119] |
| COL4A4 | Encodes Collagen Type IV alpha-4 chain. | Mutations related to Alport syndrome type 2 and autosomal dominant or recessive modes of inheritance for benign familial hematuria. | 2q36.3 | Autosomal Dominant (Alport syndrome type 3), Autosomal Recessive (Alport syndrome type 2) | [115,116,117,118,119] |
| COL4A5 | Encodes Collagen Type IV alpha-5 chain. | Mutations related to Alport syndrome type 1, exhibiting X-linked dominant mode of inheritance. | Xq22 | X-Linked Dominant | [115,116,118,119,120] |
| ITGA3 | Encodes integrin alpha-3 chain and forms an integrin-family molecule, VLA-3. | Mutations associated with interstitial lung disease, NS, and congenital epidermolysis bullosa. | 17q21.33 | Autosomal Recessive | [121,122,123] |
| ITGB4 | Encodes integrin beta-4 chain and forms the alpha-6-beta-4 integrin molecule. | Mutations associated with congenital FSGS and epidermolysis bullosa with pyloric atresia. | 17q25.1 | Autosomal Recessive | [124,125,126] |
| LAMB2 | Encodes laminin beta-2 chain. Mutations associated with Pierson syndrome. | Mutations cause Pierson syndrome, presenting as congenital NS with mesangial sclerosis and eye abnormalities. | 3p21 | Autosomal Recessive | [127,128] |
| Nuclear transcription factors and protein mutations | |||||
| LMX1B | Encodes LIM homeobox transcription factor. | Mutations cause FSGS and nail–patella syndrome. | 9q32-34.1 | Autosomal Dominant | [129,130,131] |
| NXF5 | Belongs to a multigene family of nuclear RNA export factors. Role in FSGS is not well understood. | Mutations might contribute to FSGS, but their significance is unknown. | X chromosome | X-Linked Recessive | [132,133] |
| SMARCAL1 | Encodes SWI/SNF-related matrix-associated, actin-dependent regulator of chromatin. | Mutations result in Schimke immunosseous dysplasia and NS. | 2q34.36 | Autosomal Recessive | [134,135,136] |
| WT1 | Encodes a zinc finger DNA-binding protein. Mutations associated with Wilms Tumor, Denys–Drash Syndrome with NS, somatic mesothelioma, and other diseases. | Mutations associated with Wilms Tumor, Denys–Drash Syndrome with NS, somatic mesothelioma, and other diseases. | 11p13 | Autosomal Dominant (Wilms Tumor, Denys–Drash Syndrome), Somatic Mutation | [137,138,139,140,141,142] |
| OSGEP | Encodes O-sialoglycoprotein endopeptidase. Mutations lead to Galloway–Mowat syndrome and GBM disorder. | Mutations cause Galloway–Mowat syndrome and GBM disorder. | 14q11.2-12 | Autosomal Recessive | [143,144,145] |
| LAGE3 | Encodes L-antigen family member 3. | Mutations cause X-linked Galloway–Mowat syndrome with NS and primary microcephaly. | Xq28 | X-Linked Recessive | [146,147] |
| WDR73 | Encodes WD repeat-containing protein 73. | Mutations cause Galloway–Mowat syndrome with microcephaly and SRNS. | 15q25.2 | Autosomal Recessive | [148,149,150] |
| TP53RK | Encodes p-53-related protein kinase. | Mutations cause Galloway–Mowat syndrome. | 20q13.2 | Autosomal Recessive | [146,151] |
| NUP93 | Encodes nucleoporin 93 kd, a subunit of the 12 million Da nuclear pore complex. | Mutations cause SRNS. | 16q13 | Autosomal Recessive | [152,153] |
| NUP107 | Encodes nucleoporin 107 kd, a subunit of the 120 million Da nuclear pore complex. | Mutations cause SRNS and ovarian dysgenesis. | 12q15 | Autosomal Recessive | [154,155,156] |
| NUP205 | Encodes nucleoporin 205 kd, a subunit of the 120 million Da nuclear pore complex. | Mutations cause SRNS. | 7q33 | Autosomal Recessive | [153,157] |
| XPO5 | Encodes exportin 5. | Mutations associated with early onset NS. | 6p21.1 | Autosomal Recessive | [153,158] |
| Proximal tubule protein reabsorption mutations | |||||
| CUBN | Encodes cubilin, the intestinal receptor for endocytosis of intrinsic factor, vitamin B12, and a receptor in epithelial apoA-I/HDL metabolism. | Mutations cause Imerslund–Grasbeck syndrome, low molecular weight proteinuria, and megaloblastic anemia. | 10p13 | Autosomal Recessive | [159,160] |
| AMN | Encodes amnion-associated transmembrane protein. Along with cubilin, it forms a receptor complex called “cubam.” | Mutations cause Imerslund–Grasbeck syndrome with B12 deficiency and proteinuria. | 14q32 | Autosomal Recessive | [161,162,163] |
| LRP2 | Encodes low-density lipoprotein receptor-related protein 2 or megalin. | Mutations cause Donnai–Barrow syndrome with facial anomalies, ocular alterations, sensorineural hearing loss, and NS. | 2q31.1 | Autosomal Recessive | [164,165,166] |
| Other mutations | |||||
| CFH | Encodes complement factor H, a serum glycoprotein that regulates the function of the alternative complement pathway. | Mutations associated with atypical hemolytic uremic syndrome and C3 glomerulopathy. | 1q31.3 | Autosomal Dominant | [167,168,169,170] |
| DGKE | Encodes diacylglycerol kinase epsilon, an intracellular molecule that phosphorylates diacylglycerol (DAG) to phosphatidic acid. | Mutations associated with susceptibility to atypical hemolytic uremic syndrome and NS. | 17q22 | Autosomal Recessive | [171,172] |
| PMM2 | Encodes phosphomannomutase, an enzyme necessary for the synthesis of GDP-mannose. | Mutations cause congenital disorders of glycosylation type I and SRNS. | 16p13.2 | Autosomal Recessive | [173,174,175] |
| PTPRO | Encodes protein-tyrosine phosphatase receptor-type O or glomerular epithelial protein 1. | Mutations cause childhood-onset NS with varying severities. | 12p12.3 | Autosomal Recessive | [176,177] |
| SCARB2 | Encodes scavenger receptor class B, member 2, a lysosomal integral membrane glycoprotein. | Mutations cause progressive myoclonic epilepsy, with or without renal failure and SRNS. | 4q21.1 | Autosomal Recessive | [178,179,180,181] |
| ZMPSTE24 | Encodes zinc metalloproteinase STE24, a protein involved in the metabolism of farnesylated proteins. | Mutations cause mandibuloacral dysplasia type B, increasing the risk for FSGS. | 1p34.2 | Autosomal Recessive | [182,183] |
| ALG1 | Encodes chitobiosyldiphosphodolichol beta-mannosyltransferase, a protein that acts in the glycosylation process. | Mutations cause congenital disorder of glycosylation type Ik, producing congenital NS. | 16p13.3 | Autosomal Recessive | [184,185,186] |
| EMP2 | Encodes epithelial membrane protein 2, which regulates cell membrane composition and plays a role in glomerular filtration. | Mutations cause childhood-onset NS. | 16p13.2 | Autosomal Recessive | [187,188,189] |
| TTC21B | Encodes tetratricopeptide repeat domain-containing protein 21B, involved in ciliary function. | Mutations cause nephronophthisis, FSGS, and short-rib thoracic dysplasia. | 2q24.3 | Autosomal Recessive | [190,191,192,193] |
| SGPL1 | Encodes sphingosine-1-phosphate lyase 1, involved in sphingolipid catabolism. | Mutations cause primary adrenal insufficiency and SRNS. | 10q22.1 | Autosomal Recessive | [194,195] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vaz de Castro, P.A.S.; Fujihara Ide, T.; Crespo Torres, F.; Simões e Silva, A.C. The View of Pediatric Nephrotic Syndrome as a Podocytopathy. Kidney Dial. 2023, 3, 346-372. https://doi.org/10.3390/kidneydial3040030
AMA Style
Vaz de Castro PAS, Fujihara Ide T, Crespo Torres F, Simões e Silva AC. The View of Pediatric Nephrotic Syndrome as a Podocytopathy. Kidney and Dialysis. 2023; 3(4):346-372. https://doi.org/10.3390/kidneydial3040030
Chicago/Turabian StyleVaz de Castro, Pedro Alves Soares, Thomas Fujihara Ide, Fernando Crespo Torres, and Ana Cristina Simões e Silva. 2023. "The View of Pediatric Nephrotic Syndrome as a Podocytopathy" Kidney and Dialysis 3, no. 4: 346-372. https://doi.org/10.3390/kidneydial3040030