
Genome Doubling of Northern Spicebush, Lindera benzoin L.


1
Department of Horticultural Science, North Carolina State University, 2721 Founders Dr, Raleigh, NC 27695, USA
2
Mountain Crop Improvement Lab, Department of Horticultural Science, Mountain Horticultural Crops Research and Extension Center, North Carolina State University, 455 Research Drive, Mills River, NC 28759, USA
3
Department of Life Sciences, National University of Kaohsiung, Kaohsiung 881, Taiwan
*
Author to whom correspondence should be addressed.
J 2024, 7(2), 116-126; https://doi.org/10.3390/j7020007
Submission received: 26 February 2024
/
Revised: 14 March 2024
/
Accepted: 16 March 2024
/
Published: 22 March 2024
(This article belongs to the Special Issue Feature Papers of J—Multidisciplinary Scientific Journal in 2024)
Abstract
:Lindera benzoin is a dioecious understory shrub native to eastern North America. Northern spicebush is a beautiful shrub with a natural round shrub shape, golden-yellow fall foliage, attractive bright red drupes, and precocious yellow flowers in early spring; however, its market value as an ornamental value has been overlooked. To improve the ornamental values of this under-cultivated nursery crop, breeding for a better compact form, larger leaves, enlarged flower clusters and fruit, and increased stress tolerances could all be beneficial. Polyploidy manipulation is a valuable method to improve such traits for many ornamental plants. This study established the genome doubling method by oryzalin-infused solid agar treatment on young northern spicebush seedlings. The seedlings of two wild populations in North Carolina were collected and used. A total of 288 seedlings were treated with solid agar containing 150 µM oryzalin for 24, 72, and 120 h. The results were sporadic in their survival ratios and tetraploid conversion ratios between different treatments; however, a total of 16 tetraploid L. benzoin plants were produced in this study. The 24-h treatment showed the optimal result, with 7.1% of total treated seedlings or 15.2% of surviving seedlings converted into tetraploids. Tetraploid plants had visible differences in leaf morphology, a statistically significant enlarged stomata size, and reduced stomatal density compared to diploid plants. This research provides ploidy manipulation information for all future breeding processes of L. benzoin and related species.
1. Introduction
Lindera spp. are of the Lauraceae family, comprising 55 genera and over 2000 species that often have aromatic, medicinal, nutritional qualities, and essential oil contents (Azhar and Salleh, 2020). Approximately 100 Lindera species are distributed between tropical and temperate regions in Asia and North America [1]. Northern spicebush, L. benzoin, has a broad native range and environmental adaptability. The native range of northern spicebush extends across the east coast of North America and is distributed from Maine and Ontario to Florida and west to Texas and Kansas [2]. In the wild, northern spicebush can be found in “stream margins or alluvial woods and on basic rock” [3]. Northern spicebush’s natural habitats consist of stream sides, low moist woodlands, valley bottoms, drainage areas in wooded hillsides, and at the base of cliffs and bluffs [4]. The naturally high adaptability in various environments implies the potential of the northern spicebush to be an easy-to-cultivate landscape plant used in a wide range of conditions.
Lindera benzoin, a northern spicebush, has exceptional potential in gardens and landscapes because of its ornamental value [5]. Northern spicebush is a dioecious or polygamodioecious understory shrub with axillary umbellate clusters of fragrant yellow flowers that appear from March to April and glossy, bright-red, spicy drupes that are present from August to September in North Carolina [3]. The plant grows upright at first and then horizontally, resulting in a dense, round, oval outline [6]. Lindera benzoin has thin, glabrous, deciduous leaves that are sparsely pubescent on the abaxial surface [5]. The alternate-simple leaves that are light green in summer and gold-yellow in fall also contribute to the northern spicebush’s ornamental value [6]. In a suitability survey of Northeastern U.S. native shrubs as replacements for invasive plants by the uniformity of shape and density of foliage, Lindera benzoin scored the second highest in aesthetic quality among eight shrub species [6].
In addition to ornamental and landscaping values, northern spicebush is also noticed as a functional crop because of its culinary [7] and potential medicinal qualities [8]. Multiple native American nations have a long history of using northern spicebush for multiple purposes, including using it as a tonic, miscellaneous remedy, gynecological aid, sexually transmitted disease aid, respiratory disease aid, antirheumatic aid, and dermatological aid [8,9,10,11]. The active compounds for the potential medicinal effect are relatively unknown; however, 39 components were identified in the essential oil extracted from the leaves, twigs, and fruits of Lindera benzoin [7]. Several bioactive compounds were extracted from this plant, including y-lactones (isolinderanolide, isolinderenolide, and linderanolide), obtusilactones (isoobtusilactone A, obtusilactone A, isoobtusilactone, and obtusilactone), and (6Z,9Z,12Z)-pentadecatrien-2-one, (6Z,9Z)-pentadecadien-2-one, and (+)-(Z)-nerolidol [12]. Some compounds from a related species, L. obtusiloba, are notable for their therapeutic potential of antioxidative and pharmacological properties, which were also found in northern spicebush [13]. Three bisabolene sesquiterpenes isolated from L. benzoin leaves were observed to reduce the pro-inflammatory prostaglandin E2 formation in A549 cells [14]. At least 341 constituents, including sesquiterpenoids, alkaloids, butanolides, lucidones, flavonoids, and phenylpropanoids, were discovered in Lindera plants, and many of them have shown their potential in anticancer, antinociceptive, antiarthritic, and antinociceptive [15]. Although many historical usages, phytochemistry, and in vitro research indicated the medicinal potential of Lindera, more research in animal models and at the clinical level is needed.
Environmental resilience, low pest pressure, and stabilization contribute to the northern spicebush’s potential in environment-friendly gardens and landscapes [16]. It has an open form as an understory shrub; northern spicebush can grow from full sun to shady spots; however, in full sun, it takes a more compact, dense form [17]. Spicebush leaves are among the first to emerge in the spring or late winter compared to other plants, which implies that they can be one of the few food sources for many animals in late winter [16]. Ecologically, the leaves and seeds of L. benzoin are a food source for more than 20 species of animals, including birds, rabbits, raccoons, and opossums [18]. In addition, the northern spicebush is an essential larval host of the spicebush swallowtail, Papilio troilus, and the eastern tiger swallowtail, Papilio glaucus [5]. Biochemical defenses within the leaves often deter deer, contributing to their proliferation, even in populated areas [18]. Their strong, wet-tolerant root system is also believed to prevent soil erosion around streams and rivers [16]. Although northern spicebush was reported to be susceptible to white-tailed deer (Odocoileus virginianus) damage, fast recovery ability was also reported [6]. As a U.S. native plant with various ecosystem importances, wide light adaptability, and low significant disease issues, L. benzoin is excellent for U.S. landscaping and gardens. These characteristics are conducive to this plant’s widespread use in different spots, and it has a unique niche in the landscaping market.
Polyploid manipulation is a widely used tool in ornamental plant and medicinal plant breeding. Increasing ploidy frequently results in morphological changes such as “thicker, darker-colored leaves; larger, longer-lasting flowers and thicker petals; enhanced vigor; improved tolerances to environmental stresses, pests, and pathogens; increased metabolite production” [19]. In many ornamental plants, larger flower size was reported to correlate to higher ploidy levels, including Hibiscus [20,21], hybrid Euphorbia [22], Lagerstroemia indica [23], and Impatiens spp. [24]. Compared to its diploid clone, which has a more compact habit, larger leaves were also found in tetraploid plants, ex. Hibiscus acetosella [25], chinese privet, Ligustrum sinense [26], and Rhododendron [27]. Thicker and darker foliage was also a common trait of induced polyploid plants, for example, Lagerstroemia indica [23,28], petunia [29], and Taraxacum kok-saghyz [30]. Enlarged flowers were also frequently reported in polyploidy induction research, including Cyclamen [31], Plectranthus × hybrida [32], and Hibiscus × hybrida [21]. Fruit size changed through ploidy manipulation is rarely discussed in ornamental plants; however, enlarged fruit at higher ploidy was reported in many fruit crops, ex, Malus × domestica [33], Actinidia arguta [34], and Olea europaea [35]. For medicinal plants, increased ploidy commonly results in an increased concentration or yield of active compounds. For example, in hemp (Cannabis sativa), higher ploidy levels and cannabinoid (CBG) concentrations in their inflorescences are correlated [36]. Moreover, tetraploid Chinese sage (Salvia miltiorrhiza) has a higher biomass and dihydrotanshinone I yield than the diploids [37]. Overall, polyploid manipulation is a valuable tool for ornamental and medicinal plants for cultivar improvements.
Oryzalin is a commonly used mitotic inhibitor for polyploidy induction. Oryzalin, a dinitroaniline herbicide, has had higher success rates in polyploid induction than colchicine for many crops [38,39]. For example, in Hibiscus acetosella, polyploid induction research and colchicine treatment on seeds showed no success in polyploid induction, while successful polyploid plants were received from the use of a 100 to 150 μM oryzalin treatment [40]. Several types of research showed that the highest concentration of oryzalin treatment gave optimal results in polyploidization. For example, in an in vivo oryzalin treatment on common cherry laurel, Prunus laurocerasus, a higher concentration (about 150 μM) showed the best tetraploid conversion ratio compared to all lower oryzalin concentration treatments, and, notably, the duration of the treatment is crucial to the survival ratio and tetraploid conversation ratio [41]. Other research works on Hydrangea serrata indicated that the polyploid induction ratio was significantly correlated with the treatment’s longer duration; however, the survival rate was unrelated to the duration [42]. Although the result can vary between species and tissue types, higher concentrations and the appropriate treatment duration of oryzalin treatment generally gave positive results in plant polyploid inductions.
This research aims to establish a protocol for the autopolyploidy induction of L. benzoin and create tetraploid L. benzoin plants for breeding purposes. With a limited seed number, three treatment periods of 24, 72, and 120 h with a high oryzalin (150 μM) solid agar treatment were tested in this research. The seedling survival ratio and tetraploid converting ratios were measured. The morphologies of the leaves and stomata of tetraploid and diploid plants were compared.
2. Materials and Methods
2.1. Plant Materials
Seeds from two wild locations of L. benzoin were collected in fall 2022. Accession C2022-005 was collected from William B. Umstead State Park, NC, USA (35.844337, −78.726215). Accession C2022-008 was collected from the Morgan Creek trail (35.896322, −79.065192), about 2.7 km west of the North Carolina Botanical Garden in Cabarrus County, NC, USA. Seeds were manually extracted from the fruit and stratified in moistened perlite at 4 °C for six months. Seeds were sowed, and plants were grown in a greenhouse maintained at 21 °C at the Horticultural Science Field Lab, Raleigh, NC (35°47′28.9″ N 78°41′53.6″ W) in February 2023 and grown in natural light conditions within a range of 25 °C maximum and 18.3 °C minimum temperatures. Approximately 100 seeds were sowed in 20″ × 14″ × 4″ Kadon Heavy Duty Plastic Vented Trays (Kadon Corp, Dayton, OH, USA) with a Sungro potting mix (Seba Beach, AB, Canada). For each accession, three trays were used. Trays were manually irrigated to keep them constantly moist until germination and ready for the treatment of the mitotic inhibitor (oryzalin).
Seeds were germinated about 2 to 11 weeks after sowing. Some seeds germinated sporadically, and most germinated in 7 weeks. For a more straightforward operation, only seedlings that germinated 3 to 7 weeks after sowing and grew to a treatable size (1–2.5 cm, Figure 1a) were treated with oryzalin solid agar. A total of 288 seedlings were ready for chemical treatment, including 121 seedlings of C2022-008 and 167 seedlings of C2022-005 (Table 1). After mitotic inhibitor treatments, plants stayed in the same greenhouse for six months until all flow cytometry examinations, stomata measurements, and morphology observation were finished.
2.2. Seedling Solid Agar Treatment
The 0.5% agar solution was prepared for the solid agar seedling treatment and then microwaved to boil before cooling. Once the agar solution cooled to ~50 °C, oryzalin (Surflan A.S.®, Southern Agricultural Products, Palmetto, FL, USA) was added to make it a 150 μmol oryzalin solution. The solution was then filled into dark brown 1.7 mL microtubes before cooling down and placed in a rack in a 4 °C refrigerator to allow the gel to be fully set and stored. The microtube lid was removed right before being applied to the seedling. Tubes were carefully approached onto the cryptocotylar hypogeal seedling to have the apical meristem inserted into the solid agar (Figure 1b). Each seedling was evenly assigned to the three groups of the three treatment durations: 24, 72, and 120 h. After the treatment, the microtube was carefully removed using plants.
Seedlings recovered in the Kadon Heavy Duty Plastic Vented Trays until two normal morphologic leaves were grown. Lateral shoots from non-treated parts of the plant were constantly checked and removed to ensure that only shoots from the treated meristem were kept. Most seedlings stopped growing until 1–2 weeks after the treatment when new leaves began to grow. The first few new leaves were usually abnormally shaped, being wrinkled and uneven in color, after which the plant started to grow normal-shaped leaves. Some seedlings died immediately after the treatment or stopped growing up to six months before death. Only seedlings that produced leaves four months after the treatment were defined as surviving plants.
2.3. Flow Cytometry
Flow cytometry (Quantum P Ploidy Analyzer, QuantaCyte, Mullica Hill, NJ, USA) was used to investigate the ploidies of surviving plants. Ploidy tests were conducted on two young, normal-looking leaves from each seedling. For each leaf sample, 1 cm2 of tissue was chopped using a razor in 150 μL of nuclei extraction buffer (Cystain Ultraviolet Precise P Nuclei Extraction Buffer; Sysmex, Görlitz, Germany) in a 60 mm diameter petri dish. Then, 600 μL of stain buffer (Cystain Ultraviolet Precise P Staining Buffer; Sysmex, Görlitz, Germany) was added. The Petri dish was gently shaken to fully mix the buffer and sample before the chopped samples were filtered using a 50 μm green gauge filter (Celltrics, Sysmex America Inc., Lincolnshire, IL, USA). The filtered sample was then collected in a 3.5 mL plastic tube (Sarstedt Ag & Co., Nümbrecht, Germany).
Nuclei were then analyzed using the flow cytometer. Each seedling was analyzed twice using new tissue to confirm the observed ploidy. Diploid L. benzoin plant material was used for reference standard set peak placement (Figure 2). Two samples of each seedling were run independently, and the placement of the peaks determined their putative ploidy. When both leaf samples of a seedling were tested as tetraploid, the seedling was then defined as tetraploid. When at least one leaf sample showed a mixoploid or its two samples showed inconsistent results, the seedling was then defined as a tetraploid mixoploid. When both leaf samples of a seedling were tested as diploid, the seedling was defined as diploid. Each tetraploid plant was transplanted into a 1-gallon pot, and their ploidies were re-tested two months after the first test; plants that remained tetraploid were then defined as stable tetraploids.
2.4. Stomatal Density Size and Measurement and Visual Morphology
Stomatal size and density measurements were made six months after treatment, comparing a diploid and a stable tetraploid plant. A Zeiss Axio imager A2 compound light microscope (Carl Zeiss Microscopy GmbH, Jenna, Germany) was used to examine the printed abaxial leaf surface for stomatal measurements. Impressions of the abaxial leaf surface were made by applying nitrocellulose nail polish (Sally Hansen® Hard As Nails® Xtreme wear® Nail Color Invisible, New York City, NY, USA), letting it dry for approximately 5–10 min, peeling the resulting impression off with transparent tape, and mounting the impression to a microscope slide for imaging with the tape. Imaging was performed with AxioVision software for stomatal density measurements. The length and width of about 40 stomata of each sample were measured. An independent two-tail unequal variance t-test compared the average stomatal length and width.
Stomata density was calculated by counting the number of stomata in six randomly placed frames captured at 40× magnification (0.7821 nm2 in each frame) taken using ZEN PRO (Carl Zeiss Microscopy GmbH, Jenna, Germany). Stomata number/frame area = stomata density (n/nm2), the average density of diploid and tetraploid plants, was compared using an independent two-tail t-test.
3. Results
3.1. Seedling Solid Agar Treatment
A total of 288 seedlings were germinated in the treatment window; the result is listed in Table 1. For the C2023-008 seedling population, 41, 40, and 40 seedlings were treated in the 150 μmol oryzalin gel for 24, 72, and 120-h treatments. As a result, 17, 28, and 14 seedlings survived, and 5, 4, and 3 seedlings were then tested tetraploid. For the C2023-005 seedling population, 58, 56, and 53 seedlings were treated in the 150 μmol oryzalin gel for 24, 72, and 120-h treatments. As a result, 19, 18, and 20 seedlings survived, and 2, 2, and 0 seedlings were then tested as tetraploid. In summary, 99, 96, and 93 seedlings were treated; 46, 55, and 50 seedlings survived, and 7, 6, and 3 were tested as tetraploid plants. The survival ratio of those treatments was expected to be negatively related to the treatment hours; however, the results showed that the 72-h treatment showed a significantly higher survival ratio than the other two treatments in the logistic (binomial) general linear model (p-value < 0.01). The tetraploid conversion ratio of tested seedlings was 7.1%, 6.2%, and 3.2% after the 24, 72, and 120-h treatments, respectively. The tetraploid conversion ratio of surviving seedlings was 15.2%, 10.5%, and 7.5% after the 24, 72, and 120-h treatments, respectively. No statistically significant difference in the conversion ratios between the three treatments was detected.
3.2. Stomata Size, Density, and Leaf Morphology
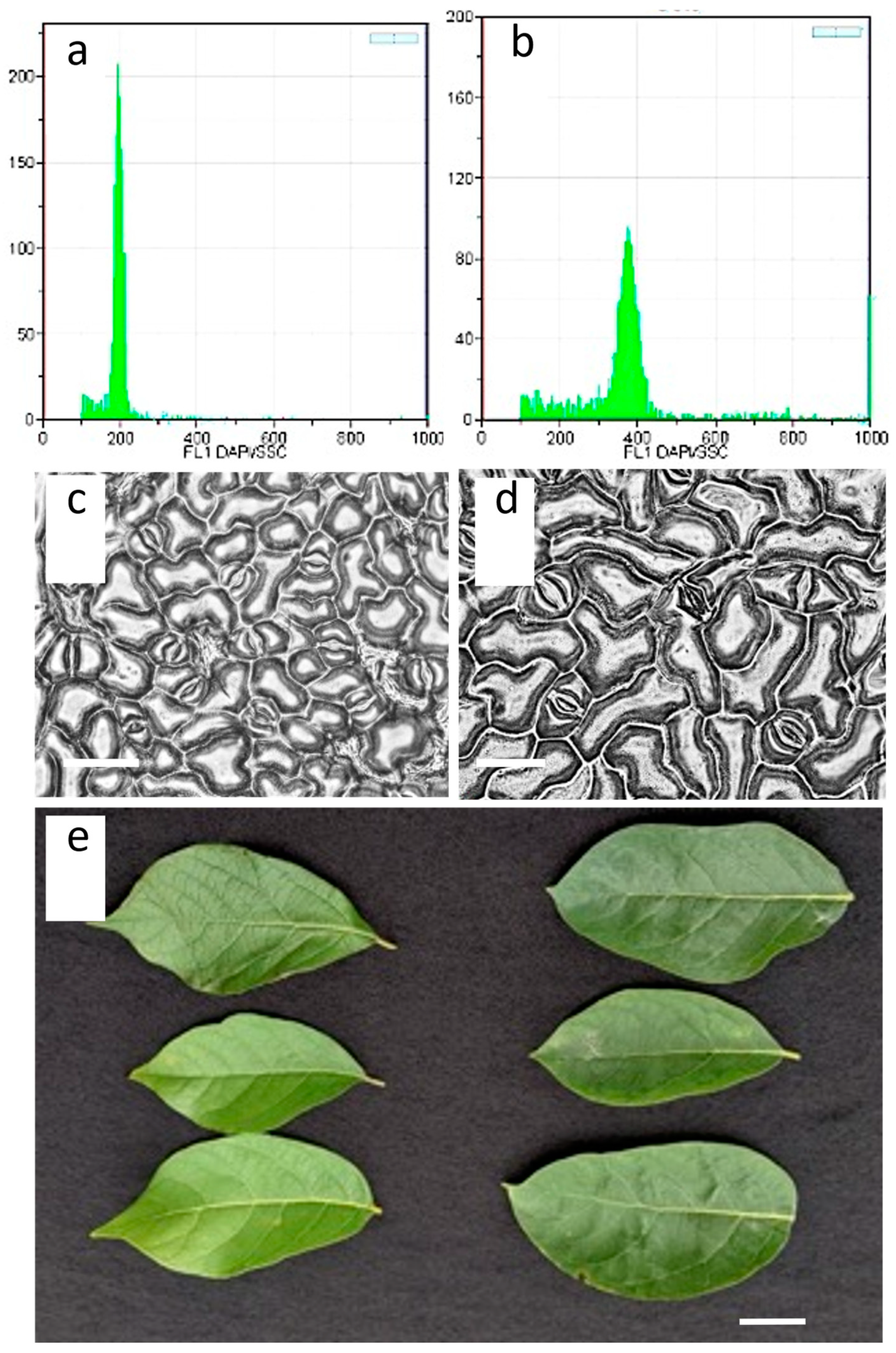
Stomata size was represented by the length and width of the 44 and 42 stomata of a standard diploid and a stable tetraploid plant, respectively. The average stomata lengths of the diploid and tetraploid plants were 18.14 µm and 29.13 µm, which was significantly different (p < 0.01). The average stomata width of the diploid and tetraploid plants was 18.97 µm and 24.19 µm, which was significantly different (p < 0.01). Stomata density was measured at 300.44 and 123.59 stomata/nm2, which was significantly different (p < 0.01). The results are listed in Table 2. The stomata images are presented in Figure 2.
The diploid and tetraploid leaf morphology showed visible differences (Figure 2e). Typical diploid Lindera benzoin leaves are simple elliptic, oblong, to obovate shapes with acuminate tips and cuneate bases to rounded bases. In contrast, the leaves of tetraploid Lindera benzoin showed elliptic leaning to oblong shape with cuspidate tips and cuneate to rounded bases. Generally, the darker leaf color of tetraploid compared to diploid leaves was primarily observed; however, a more definite measurement is needed to confirm.
4. Discussion
In this study, the 150 µM oryzalin solid agar treatment with three durations was tested, and sixteen tetraploid plants were recovered. The survival ratios of 24-h and 72-h treatments are 46% and 55%, not far from 50%. In contrast, the survival ratios of the 120-h treatment are statistically reduced compared to the other two durations. From our result, seven and six tetraploid plants were received from the 24-h and 72-h treatments, and only three were received from the 120-h treatment. Although the tetraploid conversion ratios, 7%, 6.3%, and 3.2% for the 24, 72, and 120-h treated seedlings, are not statistically or significantly different among the three durations, the tetraploid conversion ratios of the 24-h and 72-h treatments are more than double the 120-h treatment. In most mutation breeding processes, a 50% lethal dose (LD50) is usually assumed to be optimal [43]. Our result is similar to the assumption that the 24-h and 72-h treatments had about a 50% lethality ratio and gave the most tetraploid plants (7% of all treated seedlings) after the treatment. Here, we simply recommend the 24-h 150 µM oryzalin solid agar treatment as the optimal treatment for the tetraploidization protocol for L. benzoin, and it could have application in relative species. To our knowledge, no genome doubling protocol for L. benzoin or Lindera spp. has been published before this study.
Several genome doubling protocols for woody plants used the 150 µM oryzalin treatment. In sugi, Cryptomeria japonica, seedlings were sprayed with 150 μM oryzalin + 0.1% SilEnergy™ for 30 consecutive days, resulting in 32.8% of the treated seedlings being tested as tetraploids [40]. Similar to our discovery, the 150 μM oryzalin treatment resulted in tetraploid shoots in Cercis glabra [44] and C. yunnanensis [45]. In a Rhododendron research work, 150 μM oryzalin resulted in the highest tetraploid conversion rate (of surviving seedlings) compared to all other treatments [46]. For Escallonia rosea, the 50 μM oryzalin resulted in a 33% tetraploid conversion ratio, which is higher than other tested treatments [47]. Similar results were reported in other woody plants like Acacia crassicarpa [48] and Dracaena [49]. In addition to the woody plants, a 150 μM oryzalin treatment was recommended for doubling Hibiscus moscheutos [50]. Although the reactions of different mitotic inhibitors and connections vary among different species and tissue types [49,51], a 150 μM oryzalin treatment could be worth including in the first trial for woody plant genome doubling, where plant material and resources are limited.
Genome doubling could enhance North Spicebush’s ornamental or aesthetic value by enlarging leaf size, enhancing leaf color, creating a more compact form, larger fruit size, and increasing environment adaptability and vigor. Tetraploid plants received in this study primarily showed some morphological changes, including leaf shape change, less stomata density, and a visibly darker leaf color. Increasing ploidy frequently brings an enlarged leaf size and a more rounded plant shape [26,27,40,42,49,52]. Shorter internode or more compact plant shapes are commonly observed [52]. However, in some cases, genome-doubled plants show a similar internode length to their original ploidy plant [21,36,53], and some even have longer internodes [42]. Ploidy’s impact on growth vigor can vary in different situations. In hemp, Cannabis sativa, triploids show the most vigor, followed by diploid and tetraploid [36]. Polyploidy manipulation has commonly been applied to fruit size, enlarging breeding in Malus × domestica [33], Actinidia arguta [34], and Olea europaea [35]; however, the impact of genome doubling on Lindera remains unknown. More observations and research on the fruit size of tetraploid Lindera plants will be needed. Reduced stomatal density could be found in many induced polyploid plants [39,54,55], and there is an assumption that low stomatal density could be correlated with increased drought tolerance [55,56]. However, better drought tolerance in higher ploidy of plants is not necessarily correct. Some polyploid plants showed improved resistance to different stress tolerances compared to their lower ploidy counterparts but reduced environmental tolerance has also often been discovered [57]. For the tetraploid Lindera, changes in leaf and stomata morphologies were reported; however, further observations are needed to know how polyploids impact plant form, growth vigor, and environmental adaptabilities.
Native plants used in nurseries and landscaping can benefit the environment, and the demand for native plants in the green industry has increased. Research showed that native plant nurseries had higher bee species diversity than non-native plant gardens [58]. In addition, native plants were believed to have co-evolved in the local environment and might be better resistant to local abiotic and biotic stresses, resulting in fewer inputs (i.e., fertilizer, irrigation, and pesticides) than non-native species [59]. Demand for native plants has increased in the U.S. nursery market [59,60,61]. In addition, some new laws could even drive this trend; for example, a new NC law requires native plants to be used in public landscapes, state parks, public roadsides, and local projects using state landscaping funds [62]. However, the production of native plants as nursery crops has extra barriers and challenges, including low seed/liner supply, limited production, slow propagation and growth, and less aesthetic appeal than conventional nursery plants [60,63,64]. Breeding native plants can help deal with the limits of the native plant market and production. Our research used a polyploid manipulation breeding strategy to increase genetic diversity and create novel traits of the northern spicebush. We hope this research will eventually benefit the native plant industry. Further research on the propagation efficiency, adult plant aesthetics, production cost, and general environmental adaptability will need to be tested before recommending the tetraploid version of L. benzoin to the industry.
Author Contributions
Conceptualization, H.C. and R.F.A.; methodology, H.C. and J.T.N.; validation, H.C., R.F.A. and T.B.M.; formal analysis, H.C. and Y.-L.K.; investigation, H.C.; resources, H.C. and J.T.N.; data curation, R.F.A. and T.B.M.; writing—original draft preparation, R.F.A.; writing—review and editing, H.C., J.T.N., T.B.M. and Y.-L.K.; visualization, H.C.; supervision, H.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data is contained within the article.
Acknowledgments
We appreciate Kyle Freeman for providing plant materials for our research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wei, G.; Zhang, J.; Lei, J.; Ma, C.; Tong, Y.; Jiang, H. Chemical constituents from Lindera nacusua (D. Don) Merr. Biochem. Syst. Ecol. 2016, 66, 94–97. [Google Scholar] [CrossRef]
- Small, E. North American Cornucopia: Top 100 Indigenous Food Plants; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Core, E.L. Carolina Flora Manual of the Vascular Flora of the Carolinas. In Bioscience; Radford, A.E., Ahles, H.E., Bell, C.R., Eds.; The University of North Carolina Press: Chapel Hill, NK, USA, 1970; Volume 20, p. 1217. [Google Scholar]
- Hoss, G. Propagation Protocol for Spicebush (Lindera benzoin). Nativ. Plants J. 2006, 7, 134–136. [Google Scholar] [CrossRef]
- USDA-NRCS Spicebush Lindera benzoin (L.) Blume. Available online: https://plants.usda.gov/DocumentLibrary/plantguide/pdf/pg_libe3.pdf (accessed on 20 December 2023).
- Shrestha, P.; Lubell, J.D. Suitability of eight northeastern US native shrubs as replacements for invasive plants in a difficult landscape site with white-tailed deer pressure. HortTechnology 2015, 25, 171–176. [Google Scholar] [CrossRef]
- Azhar, M.A.M.; Salleh, W.M.N.H.W. Chemical composition and biological activities of essential oils of the genus Litsea (Lauraceae)—A review. Agric. Conspec. Sci. 2020, 85, 97–103. [Google Scholar]
- Tucker, A.O.; Maciarello, M.J.; Burbage, P.W.; Sturtz, G. Spicebush [Lindera benzoin (L.) Blume var. benzoin, Lauraceae]: A Tea, Spice, and Medicine. Econ. Bot. 1994, 48, 333–336. [Google Scholar] [CrossRef]
- Tantaquidgeon, G. Folk Medicine of the Delaware and Related Algonkian Indians; DIANE Publishing Inc.: Darby, PA, USA, 2007. [Google Scholar]
- Hutton, K. A Comparative Study of the Plants Used for Medicinal Purposes by the Creek and Seminoles Tribes. Master’s Thesis, University of South Florida, Tampa, FL, USA, 2010. [Google Scholar]
- Hamel, P.B.; Chiltoskey, M.U. Cherokee Plants and Their Uses: A 400 Year History; Herald Publishing Company: Sylva, NC, USA, 1975. [Google Scholar]
- Anderson, J.E.; Ma, W.; Smith, D.L.; Chang, C.; McLaughlin, J.L. Biologically active γ-lactones and methylketoalkenes from Lindera benzoin. J. Nat. Prod. 1992, 55, 71–83. [Google Scholar] [CrossRef]
- Haque, M.E.; Azam, S.; Balakrishnan, R.; Akther, M.; Kim, I. Therapeutic potential of Lindera obtusiloba: Focus on antioxidative and pharmacological properties. Plants 2020, 9, 1765. [Google Scholar] [CrossRef]
- Henz Ryen, A.; Göls, T.; Steinmetz, J.; Tahir, A.; Jakobsson, P.; Backlund, A.; Urban, E.; Glasl, S. Bisabolane sesquiterpenes from the leaves of Lindera benzoin reduce prostaglandin E2 formation in A549 cells. Phytochem. Lett. 2020, 38, 6–11. [Google Scholar] [CrossRef]
- Cao, Y.; Xuan, B.; Peng, B.; Li, C.; Chai, X.; Tu, P. The genus Lindera: A source of structurally diverse molecules having pharmacological significance. Phytochem. Rev. 2016, 15, 869–906. [Google Scholar] [CrossRef]
- Smith, R.; Pomper, K.W.; Lowe, J.D.; Botkins, J.; Crabtree, S.B. Genetic Diversity in Kentucky Spicebush Populations using Simple Sequence Repeat Markers. J. Ky. Acad. Sci. 2012, 73, 101–109. [Google Scholar] [CrossRef]
- Poston, T.; Lee, C.N.; Choie, Y.; Kwon, Y. Local minima and back propagation. In Proceedings of the IJCNN-91-Seattle International Joint Conference on Neural Networks, Seattle, WA, USA, 8–12 July 1991; Volume 2, pp. 173–176. [Google Scholar]
- Templeton, L.K.; Neel, M.C.; Groffman, P.M.; Cadenasso, M.L.; Sullivan, J.H. Changes in vegetation structure and composition of urban and rural forest patches in Baltimore from 1998 to 2015. For. Ecol. Manag. 2019, 454, 117665. [Google Scholar] [CrossRef]
- Touchell, D.H.; Palmer, I.E.; Ranney, T.G. In vitro Ploidy Manipulation for Crop Improvement. Front. Plant Sci. 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Singh, F.; Khoshoo, T.N. Chromosomal polymorphism within the Hibiscus rosa-sinensis complex. Caryologia 1970, 23, 19–27. [Google Scholar] [CrossRef]
- Chen, H.; Contreras, R.N. Near-hexaploid and near-tetraploid aneuploid progenies derived from backcrossing tetraploid parents Hibiscus syriacus × (H. syriacus × H. paramutabilis). Genes 2022, 13, 1022. [Google Scholar] [CrossRef]
- Pan, I.; Lu, Y.; Wen, P.; Chen, Y. Using colchicine to create poinsettia (Euphorbia pulcherrima × Euphorbia cornastra) mutants with various morphological traits. HortScience 2019, 54, 1667–1672. [Google Scholar] [CrossRef]
- Ye, Y.M.; Tong, J.; Shi, X.P.; Yuan, W.; Li, G.R. Morphological and cytological studies of diploid and colchicine-induced tetraploid lines of crape myrtle (Lagerstroemia indica L.). Sci. Hortic. 2010, 124, 95–101. [Google Scholar] [CrossRef]
- Defiani, M.R.; Astarini, I.A.; Kriswiyanti, E. Oryzalin and gamma radiation induced polyploidization in garden balsam plants (Impatiens balsamina L.) in vitro. Curr. Agric. Res. J. 2017, 5, 1–5. [Google Scholar] [CrossRef]
- Contreras, R.N.; Ruter, J.M.; Hanna, W.W. An oryzalin-induced autoallooctoploid of Hibiscus acetosella ‘Panama Red’. J. Am. Soc. Hort. Sci. 2009, 134, 553–559. [Google Scholar] [CrossRef]
- Fetouh, M.I.; Deng, Z.; Wilson, S.B.; Adams, C.R.; Knox, G.W. Induction and characterization of tetraploids in chinese privet (Ligustrum sinense Lour.). Sci. Hortic. 2020, 271, 109482. [Google Scholar] [CrossRef]
- Mo, L.; Chen, J.; Lou, X.; Xu, Q.; Dong, R.; Tong, Z.; Huang, H.; Lin, E. Colchicine-induced polyploidy in Rhododendron fortunei Lindl. Plants 2020, 9, 424. [Google Scholar] [CrossRef]
- Tong, J.; Ye, Y.; Feng, B.; Yuan, W. Colchicines induced polyploid plants and their identification in three species of Lagerstroemia indica. Acta Hortic. Sin. 2009, 36, 127–132. [Google Scholar]
- Ning, G.; Shi, X.; Hu, H.; Yan, Y.; Bao, M. Development of a range of polyploid lines in Petunia hybrida and the relationship of ploidy with the single-/double-flower trait. HortScience 2009, 44, 250–255. [Google Scholar] [CrossRef]
- Luo, J.; Ren, W.; Cai, G.; Huang, L.; Shen, X.; Li, N.; Nie, C.; Li, Y.; Wang, N. The chromosome-scale genome sequence of Triadica sebifera provides insight into fatty acids and anthocyanin biosynthesis. Commun. Biol. 2022, 5, 786. [Google Scholar] [CrossRef] [PubMed]
- Takamura, T.; Sugimura, T.; Tanaka, M.; Kage, T. Breeding of the tetraploid yellow-flowered cyclamen with “eye”. In Proceedings of the III International Symposium on New Floricultural Crops, Perth, Australia, 1–4 October 1996; pp. 119–126. [Google Scholar]
- Brits, G.J. Polyploid breeding of wild South African plectranthus (spurflowers) as new flowering pot plants, XXVII International, Horticultural Congress-IHC2006: International Symposium on Endogenous and Exogenous Plant. Bioregulators 2006, 774, 437–442. [Google Scholar]
- Guerra, D.; Wittmann, M.T.S.; Schwarz, S.F.; Souza, P.V.D.d.; Gonzatto, M.P.; Weiler, R.L. Comparison between diploid and tetraploid citrus rootstocks: Morphological characterization and growth evaluation. Bragantia 2014, 73, 1–7. [Google Scholar] [CrossRef]
- Kataoka, I.; Mizugami, T.; Kim, J.G.; Beppu, K.; Fukuda, T.; Sugahara, S.; Tanaka, K.; Satoh, H.; Tozawa, K. Ploidy variation of hardy kiwifruit (Actinidia arguta) resources and geographic distribution in Japan. Sci. Hortic. 2010, 124, 409–414. [Google Scholar] [CrossRef]
- Rosati, A.; Caporali, S.; Hammami, S.B.; Moreno-Alías, I.; Rapoport, H. Fruit growth and sink strength in olive (Olea europaea) are related to cell number, not to tissue size. Funct. Plant Biol. 2020, 47, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.; Rojas, B.M.; Crawford, E.; Otten, M.; Schoenenberger, T.A.; Garfinkel, A.R.; Chen, H. Characteristics of the diploid, triploid, and tetraploid versions of a cannabigerol-fominant F1 Hybrid Industrial Hemp Cultivar, Cannabis sativa ‘Stem Cell CBG’. Genes 2021, 12, 923. [Google Scholar] [CrossRef]
- Chen, E.G.; Tsai, K.; Chung, H.; Chen, J. Chromosome doubling-enhanced biomass and dihydrotanshinone I production in Salvia miltiorrhiza, a traditional Chinese medicinal plant. Moleculars 2018, 23, 3106. [Google Scholar] [CrossRef]
- Barandalla, L.; Ritter, E.; Ruiz De Galarreta, J.I. Oryzalin treatment of potato diploids yields tetraploid and chimeric plants from which euploids could be derived by callus induction. Potato Res. 2006, 49, 143–154. [Google Scholar] [CrossRef]
- Gallone, A.; Hunter, A.; Douglas, G.C. Polyploid induction in vitro using colchicine and oryzalin on Hebe ‘Oratia Beauty’: Production and characterization of the vegetative traits. Sci. Hortic. 2014, 179, 59–66. [Google Scholar] [CrossRef]
- Contreras, R.N.; Ruter, J.M.; Schwartz, B.M. Oryzalin-induced tetraploidy in Cryptomeria japonica (Cupressaceae). HortScience 2010, 45, 316–319. [Google Scholar] [CrossRef]
- Contreras, R.N.; Meneghelli, L. In vitro chromosome doubling of Prunus laurocerasus ‘Otto Luyken’and ‘Schipkaensis’. HortScience 2016, 51, 1463–1466. [Google Scholar] [CrossRef]
- Deans, L.E.; Palmer, I.E.; Touchell, D.H.; Ranney, T.G. In Vitro induction and characterization of polyploid Hydrangea macrophylla and H. serrata. HortScience 2021, 56, 709–715. [Google Scholar] [CrossRef]
- Melsen, K.; van de Wouw, M.; Contreras, R. Mutation breeding in ornamentals. HortScience 2021, 56, 1154–1165. [Google Scholar] [CrossRef]
- Nadler, J.D.; Pooler, M.; Olsen, R.T.; Coleman, G.D. In vitro induction of polyploidy in Cercis glabra Pamp. Sci. Hortic. 2012, 148, 126–130. [Google Scholar] [CrossRef]
- Nadler, J.D. In Vitro Induction of Polyploidy in Cercis yunnanensis Hu et Cheng. Master’s Thesis, University of Maryland, College Park, MD, USA, 2009. [Google Scholar]
- Väinölä, A. Polyploidization and early screening of Rhododendron hybrids. Euphytica 2000, 112, 239–244. [Google Scholar] [CrossRef]
- Denaeghel, H.; Van Laere, K.; Leus, L.; Van Huylenbroeck, J.; Van Labeke, M. Induction of Tetraploids in Escallonia spp. In Proceedings of the XXV International EUCARPIA Symposium Section Ornamentals: Crossing Borders 1087, Melle, Belgium, 28 June–2 July 2015; pp. 453–458. [Google Scholar]
- Lam, H.K.; Harbard, J.L.; Koutoulis, A. Tetraploid induction of Acacia crassicarpa using colchicine and oryzalin. J. Trop. For. Sci. 2014, 26, 347–354. [Google Scholar]
- Teng, E.S.; Leonhardt, K.W. In vitro and in vivo polyploidization of Dracaena with oryzalin. In Proceedings of the VI International Symposium on New Floricultural Crops, Funchal, Portugal, 11–15 June 2007; Volume 813, pp. 509–516. [Google Scholar]
- Li, Z.; Ruter, J.M. Development and evaluation of diploid and polyploid Hibiscus moscheutos. HortScience 2017, 52, 676–681. [Google Scholar] [CrossRef]
- Hebert, C.J.; Touchell, D.H.; Ranney, T.G.; LeBude, A.V. In Vitro Shoot Regeneration and Polyploid Induction of Rhododendron ‘Fragrantissimum Improved’. HortScience 2010, 45, 801–804. [Google Scholar] [CrossRef]
- Oates, K.M.; Ranney, T.G.; Touchell, D.H. Influence of induced polyploidy on fertility and morphology of Rudbeckia species and hybrids. HortScience 2012, 47, 1217–1221. [Google Scholar] [CrossRef]
- Lattier, J.D.; Chen, H.; Contreras, R.N. Variation in genome size, ploidy, stomata, and rDNA signals in Althea. J. Am. Soc. Hort. Sci. 2019, 144, 130–140. [Google Scholar] [CrossRef]
- Alexander, L. Production of triploid Hydrangea macrophylla via unreduced gamete breeding. HortScience 2017, 52, 221–224. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. N. Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Zhou, M.; Shabala, S. How does stomatal density and residual transpiration contribute to osmotic stress tolerance? Plants 2023, 12, 494. [Google Scholar] [CrossRef]
- Ranney, T.G. Polyploidy: From evolution to new plant development. In Combined Proceedings International Plant Propagators’ Society; IPPS: Bellefonte, PA, USA, 2006; Available online: https://ena.ipps.org/uploads/docs/56_85.pdf (accessed on 10 December 2023).
- Cecala, J.M.; Wilson Rankin, E.E. Wild bee functional diversity and plant associations in native and conventional plant nurseries. Ecol. Entomol. 2021, 46, 1283–1292. [Google Scholar] [CrossRef]
- Rihn, A.L.; Knuth, M.J.; Peterson, B.J.; Torres, A.P.; Campbell, J.H.; Boyer, C.R.; Palma, M.A.; Khachatryan, H. Investigating drivers of native plant production in the United States green industry. Sustainability 2022, 14, 6774. [Google Scholar] [CrossRef]
- Kauth, P.J.; Pérez, H.E. Industry survey of the native wildflower market in Florida. HortTechnology 2011, 21, 779–788. [Google Scholar] [CrossRef]
- Zadegan, Y.R.; Behe, B.K.; Gough, R. Consumer preferences for native plants in Montana residential landscapes and perceptions for naturalistic designs. J. Environ. Hortic. 2008, 26, 109–114. [Google Scholar] [CrossRef]
- Graham, B. Native Plants Policy, Conservation funding increases pass the Legislature. Audubon North Carol. 2023. Available online: https://nc.audubon.org/news/native-plants-policy-conservation-funding-increases-pass-legislature (accessed on 3 January 2024).
- Beck, T.B. Gardeners perceptions of the aesthetics, manageability, and sustainability of residential landscapes. Appl. Environ. Educ. Commun. 2002, 1, 163–172. [Google Scholar] [CrossRef]
- Norcini, J. Native plants: An overview. Environ. Hort. Dept. Florida Coop. Ext. Serv. Inst. Food Agr. Sci. Univ. Florida, Document ENH1045. 2006. Available online: https://edis.ifas.ufl.edu/publication/EP297 (accessed on 15 January 2024).
Figure 1.
Oryzalin solid agar treatment with different durations on Lindera benzoin seedlings: (a) A 3-day seedling that reaches the size ready for the treatment; (b) the 150 µmol oryzalin solid agar treatment on a seedling; (c) a C2023-008 seedling 19 days after a 72-h treatment; seedling showed two leaves with abnormal morphology and two new normal-looking leaves. Scale bar = 1 cm.
Figure 1.
Oryzalin solid agar treatment with different durations on Lindera benzoin seedlings: (a) A 3-day seedling that reaches the size ready for the treatment; (b) the 150 µmol oryzalin solid agar treatment on a seedling; (c) a C2023-008 seedling 19 days after a 72-h treatment; seedling showed two leaves with abnormal morphology and two new normal-looking leaves. Scale bar = 1 cm.

Figure 2.
Flow cytometry, stomata, and leave morphology of a control diploid and a stable tetraploid Lindera benzoin plant: (a,b) flow cytometry results of diploid (a) and tetraploid (b) sample; (c,d) leave epidermis images of a diploid (c) and a tetraploid (d) plant. Scale bars = 20 µm; (e) Leave morphology of a diploid (left) and tetraploid (right) plant. Scale bar = 1 cm.
Figure 2.
Flow cytometry, stomata, and leave morphology of a control diploid and a stable tetraploid Lindera benzoin plant: (a,b) flow cytometry results of diploid (a) and tetraploid (b) sample; (c,d) leave epidermis images of a diploid (c) and a tetraploid (d) plant. Scale bars = 20 µm; (e) Leave morphology of a diploid (left) and tetraploid (right) plant. Scale bar = 1 cm.


{kind=link}
{kind=link}
Table 1.
Result of 150 µmol oryzalin solid agar treatment with different durations on Lindera benzoin seedlings.
Table 1.
Result of 150 µmol oryzalin solid agar treatment with different durations on Lindera benzoin seedlings.
| Accession | Duration (H) | Treated Plants # | Survival | Diploid # | Mixoploid # | Tetraploid # |
|---|---|---|---|---|---|---|
| C2023-008 | 24 | 41 | 17 | 6 | 6 | 5 |
| 72 | 40 | 28 | 16 | 8 | 4 | |
| 120 | 40 | 14 | 5 | 6 | 3 | |
| C2023-005 | 24 | 58 | 29 | 19 | 8 | 2 |
| 72 | 56 | 27 | 18 | 7 | 2 | |
| 120 | 53 | 26 | 20 | 6 | 0 | |
| Total | 24 | 99 | 46a * | 25a * | 14a * | 7a * |
| 72 | 96 | 55b | 34a | 15a | 6a | |
| 120 | 93 | 40a | 25a | 12a | 3a |
# = Number. H = Hours. * Significant difference detected by logistic general linear model analysis with 95% confidence.
Table 2.
Stomata sizes and density of diploid and tetraploid Lindera benzoin L.
| n | Length (µm) | SD | Width (µm) | SD | n | Density (per nm2) | SD | |
|---|---|---|---|---|---|---|---|---|
| 2x | 44 | 18.14 | 1.76 | 18.97 | 2.67 | 6 | 300.44 | 28.87 |
| 4x | 42 | 29.13 | 2.50 | 24.19 | 2.22 | 6 | 123.59 | 19.25 |
| p-value * | 7.65 × 10−36 | 1.21 × 10−15 | 7.41 × 10−7 |
* t-test, Two-tailed unequal variance t-test, SD = standard deviation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arram, R.F.; Morgan, T.B.; Nix, J.T.; Kao, Y.-L.; Chen, H. Genome Doubling of Northern Spicebush, Lindera benzoin L. J 2024, 7, 116-126. https://doi.org/10.3390/j7020007
AMA Style
Arram RF, Morgan TB, Nix JT, Kao Y-L, Chen H. Genome Doubling of Northern Spicebush, Lindera benzoin L. J. 2024; 7(2):116-126. https://doi.org/10.3390/j7020007
Chicago/Turabian StyleArram, Ramsey F., Thomas B. Morgan, John T. Nix, Yu-Lin Kao, and Hsuan Chen. 2024. "Genome Doubling of Northern Spicebush, Lindera benzoin L." J 7, no. 2: 116-126. https://doi.org/10.3390/j7020007