
Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms
Museum National d’Histoire Naturelle, Paléontologie, 75005 Paris, France
Quaternary 2022, 5(3), 38; https://doi.org/10.3390/quat5030038
Submission received: 12 February 2022
/
Revised: 8 July 2022
/
Accepted: 22 August 2022
/
Published: 11 September 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Discussion of the phylogenetic relations between Plesippus, Allohippus, and Equus. Descriptions and illustrations of 30 Equid extant and fossil species younger than 2 Ma. Particular attention is given to slender forms with short protocones usually referred to ‘Equus altidens’ from Süssenborn and Untermassfeld (Germany), Akhalkalaki and Dmanisi (Georgia), Pirro (Italy), Venta Micena (Spain) and Aïn Hanech (Algeria). Occurrence of Asinine features in fossil taxa from Africa, Greece, Mongolia, and North-Eastern Siberia. Supplementary Materials include additional discussions and photographs of fossils in particular from Süssenborn (especially those referred to E. altidens and E. marxi by Reichenau) and from Dmanisi from where a new species is described.
Keywords:
Plesippus; Allohippus; Asinus; Equus; Dolichohippus; Hemionus; Hippotigris; Quagga; Sussemionus; E. altidens; E. antunesi; E. apolloniensis; E. capensis; E. chosaricus; E. coliemensis; E. ferus; E. germanicus; E. granatensis; E. graziosii; E. hipparionoides; E. hydruntinus; E. marxi; E. mauritanicus; E. melkiensis; E. nalaikhaensis; E. oldowayensis; E. ovodovi; E. przewalskii; E. cf. scotti; E. suessenbornensis; E. aff. suessenbornansis; E. tabeti; E. wuesti1. Introduction
The aim of this article is to ease the identification and, if possible, clarify and rectify some points in the taxonomy of extant Equus species, their fossil relatives, and some fossils whose relations are uncertain. This review is not exhaustive, nor balanced: paragraphs dealing with some taxa may be more developed than others; for example, Equids from Dmanisi and Süssenborn, in particular Equus altidens, are given special attention. Since fossils are the main concern of this work, the descriptions and discussions are limited to crania, teeth, and limb bones. I hope that the descriptions and the numerous photographs (partly in the Supplementary Materials) may help paleomammalogists and archeozoologists confronted with problems of determination. Aside from this practical concern, there is the more fundamental problem presented by a diagnosis of the genus Equus. Although the phylogeny and taxonomy of the entire genus Equus are beyond the scope of this work, this issue will be addressed first. Some of the existing proposals that are either incorrect, incomplete, or misleading are noted below.
In 1978, Churcher and Richardson [1] gave a diagnosis where some characteristics are not really useful. For example: ‘Bony auditory meatus variable in length and orientation; basi-cranial region with or without low longitudinal crest; grooves on mandibular incisors variably developed; canines usually absent in females’. Other elements of the diagnosis are inexact: ‘Few to no enamel plis on the pre- and postfossette mesial and distal borders’ and ‘Ascending ramus with obliquely posterior orientation’; these characters are just variable. Eventually remain the ‘hypsodonty, the ‘protocone united to protoloph’ and the ‘monodactyly’—not exclusive to Equus.
In 1994, MacFadden ([2], chapter 2, p. 11) noted that ‘A cladistic approach to the study of taxonomy and phylogeny is currently considered as a great improvement, sometimes even as a “must”, in paleontology’. Later MacFadden ([3], p. 20) wrote: ‘The (cladistic) approach… now is the dominant theoretical framewok used by vertebrate paleontologists to construct phylogenies… of extant and fossils groups’.
In the proposed cladogram ([3], Figures 5–15, p. 100), node 7 for genus Equus is defined by ‘Dorsal preorbital fossa poorly developed or absent, very high crowned and relatively straight teeth, complex enamel plications and protocones, elongated and either robust or gracile metapodials, well developed intermediate tubercule on distal (sic) humerus’. There, again, some characteristics are rather vague, ‘relatively’, ‘either robust or gracile’, or inexact: enamel plications are not always complex.
Barron-Ortiz et al. [4] asked a very good question: ‘What is Equus?’ They answered it through phylogenetic analysis including 32 characteristics (12 cranial, 16 dental, and 4 related to limb bones and body size) and 21 taxa ranging in time from the Miocene to the present, but not including extant Zebras and Asses, nor fossil Sussemiones. They found six synapomorphies for clades 6 and 7 ([4], Figure 2): three of those synapomorphies are unambiguous (meaning that they only occur in clade 6) and three synapomorphies are ambiguous (the characteristic states that are synapomorphies for clade 6 but are also present in one or a few other taxa outside of clade 6). The ambiguous synapomorphies are: absent or poorly developed buccinator fossa; P1 absent; and an oval protocone outline on P2. The unambiguous synapomorphies for clade 6 are: a short and squared lambdoidal crest (or external occipital crest); an oblong protocone outline on P3–P4; and a high, well-developed proximal intermediate tubercule on the humerus (the well-developed humeral proximal intermediate tubercule is an important characteristic albeit not absolutely ‘unique to Equids’ [2] since it is present in Hipparion heintzi of çalta, Turkey (humerus ACA 94) and also in Camels). It is stated in the conclusions ([4], p. 9) that ‘Allohippus and Plesippus should be elevated to generic rank’.
The study of Cirilli and al. [5] involves 30 taxa ranging in time from the Eocene to modern times but does not include two important taxa: the extant Ass and the fossil Sussemionus group, although the latter is well documented by a cranium and many cheek teeth with original characteristics. Among the 129 characteristics (44 cranial, 12 mandibular, 57 dental, and 16 for limb bones), several are not very useful. For example, the state of characteristics 19 and 80 are unknown except in two or three taxa; the state of characteristic 21 is the same in all.
Inside the genus Equus ([5], Figure 2), the “Zebras clade” comprises E. quagga and the Hemiones E. hemionus and E. kiang, wrongly labeled “Asses” (Asses do not figure in among the studied taxa), E. zebra, and E. grevyi.
The pairing of Quaggas and Hemiones is astonishing. The authors suppose that it may be explained by “a close morphological similarity in cranial and postcranial elements of the skeleton”. Actually, it is not possible: Quaggas have rather caballine cranial proportions and rather robust limb bones, while Hemiones have rather asinine cranial proportions and are characterized by extremely slender limb bones. The only thing they have in common is their relatively small size.
Cirilli and al. ([6], p. 4) do not recognize a generic rank for Allohippus and Plesippus, both of which they include in the genus Equus, but consider that E. simplicidens-E. stenonis-E. koobiforensis form an evolutionary lineage leading to the “Zebras” clade. The authors state that the evolution from “Equus simplicidens” includes the reduction in the vomerine length and the elongation of the post-vomerine length, but omit to mention that multivariate analyses of Plesippus, Allohippus, and Equus crania have evidenced the same more than twenty years ago [7].
The distinction between Plesippus, Alohippus, and Equus has been discussed previously [8,9,10] and summarized and illustrated again in [11]. Schematically: Equus and Plesippus have shorter naso-incisival notches and shorter vomerine lengths relative to the palatal lengths sensu stricto than Allohippus; Equus have longer post-vomerine lengths (and ipso facto, larger braincases) relative to overall palatal lengths than Plesippus and Allohippus. The schematic Figure S1 attempts to illustrate the different depths of the naso-incisival notch—longer in Allohippus (C) than in Plesippus (B), and the relatively longer braincase in Equus (A) than in Plesippus (B) and Allohippus (C).
Recently it has been proposed [12] that extant zebras derive from Plesippus simplicidens (although no other zebra than E. (Dolichohippus) grevyi was studied). A single transformation of the cranial proportions of Plesippus would thus be required to occur before the putative differentiation of the three African lineages of zebras. However, what about the other lineages of Equus? Were other Plesippus than Plesippus simplicidens at the origin of other Equus?
If Allohippus koobiforensis evolved into E. grevyi [13] and Allohippus stenonis into E. apolloniensis [14], Allohippus must have undergone at least two modifications, one affecting the post-vomerine length, and the other the length of the naso-incisival notch. While it can be supposed that the length of the naso-incisival notch is not of paramount importance, the size of the braincase most probably is. It can be supposed that the first transformation may have occurred independently in various Allohippus lineages, but was it also the case of the last, much more important one? Did distinct Allohippus species undergo the same modifications in Africa and in Europe, transforming them into distinct Equus species? Is Equus polyphyletic?
Too many points remain unknown to answer that with certitude—mostly the lack of fossil crania. However, some decision must be made before proceeding: I shall assume for now that the braincase size modification occurred just once and, in other words, that the genus Equus is monophyletic.
I believe that Plesippus (simplicidens and other), Allohippus (stenonis and other), and Equus are stades in evolution but do not form a direct lineage. All Equus share a cranial synapomorphy [7]—an important one since it is probably related to the increase in brain size. However, Allohippus alone evolved a characteristic deep naso-incisival notch, which is not found in Equus. It is probable that some Plesippus evolved in the direction of Allohippus (acquiring the deep naso-incisival notch) while another evolved in the direction of Equus (acquiring a larger brain). The former seems confirmed by comparisons of crania from lower and upper levels of Longdan, China ([15], p. 1363).
2. Material and Methods
2.1. Material
Skeletons of extant Equus and Equid fossils were studied in the Collections listed below.
Museum Acronyms used in this article
AM: Zoologisch Museum, Amsterdam, Nederland.
DD: Dehra Dun, India, Dr. Nita Shah collections.
DSTF: findings in the northern escarpment of Cava Pirro, made by researchers of the University of Florence
HUJ-ESE: Section of Ecology, Systematic and Evolution, Hebrew University, Jerusalem
IA: Geological Institute, Yakutsk, Russia.
IGF: Istituto di Geologia, Firenze, Florence, Italy
IPH: Institut de Paléontologie Humaine, Paris.
IVCM: Imperial Valley College Museum, California, U.S.A.
KNM: Kenya National Museums, Nairobi, Kenya.
LACM: Los Angeles County Museum.
LGPUT: Laboratory of Geology and Paleontology, Aristotle University Thessaloniki, Thessaloniki, Greece.
MB: Museum für Naturkunde, Berlin, Germany.
MCZ: Harvard University, Cambridge, USA.
MGU and MS: Zoological Museum of the Moscow University, Moscow, Russia.
MNHL: Muséum d’Histoire naturelle (ex Musée Guimet), Lyon, France.
MNHN-F: Laboratoire de Paléontologie du MNHN, Paris, France.
MNP-Bonifay: collection of M.F. Bonifay.
MS: Zoological Museum of the Moscow University, Moscow, Russia.
NHMUK -ZD (zoology): British Museum (Natural History), London, Great Britain.
PH: Academy of Natural Sciences, Philadelphia, U.S.A.
PIN: Paleontological Institute, Moscow, Russia.
SAM: South African Museum, Cape Town, RSA.
SAP: Service Géologique du Portugal, Lisbon, Portugal
SI: Severtsov Institute, Moscow, Russia.
TB: Janashia Museum of Georgia.
Windhoek: Windhoek Sciences Museum collections, Namibia.
ZIN: Zoological Institute, Sankt Petersburg, Russia.
Abbreviations
dP1, dP2, dP3, dP4: upper first, second, third, fourth decidual premolars.
dp2, dp3, dp4: lower second, third, fourth decidual premolars.
I1, I2, I3: upper first, second, third incisors.
i1, i2, i3: lower first, second, third incisors.
M1, M2, M3: upper first, second, third molars.
m1, m2, m3: lower first, second, third molars.
MC: third metacarpal.
MT: third metatarsal.
P1, P2, P3, P4: upper first, second, third, fourth premolars.
p1, p2, p3, p4: lower first, second, third, fourth premolars.
Ph1, Ph2, Ph3: first, second, third, fourth phalanges.
Prot.: protocone.
Detailed data on cheek teeth and limb bones of Hemiones were published by Eisenmann and Mashkour [16]. For other extant species, numerical data and photographs may be found at Equidae monodactyles> Equus actuels et récemment éteints (https://vera-eisenmann.com/, accessed on 1 January 2009).
For fossil species numerical data and photographs may be found at Equidés monodactyles fossiles (Equus, Allohippus, (...) https://vera-eisenmann.com accessed on 1 January 2009).
2.2. Methods
2.2.1. System of Measurements
The detailed system of measurements used in this article may be found in Eisenmann 1986 [17] with slight modifications and additions on my website at: https://vera-eisenmann.com/-system-of-measurementments-for-Equus-bones-and-teeth-english (accessed on 10 October 2009).
Click on the element for which measurement details are needed.
2.2.2. Simpson’s Diagrams
Simpson’s ratio diagrams [18] are used for comparisons. Although they are not absolutely reliable to draw definitive conclusions, they do offer rapid and easy comparisons, both of size and shape for a single bone or a group of bones. The reference is provided by a single bone (or a group of bones) or the means of a bone sample, the dimensions of which are converted into decimal logarithms. By convention, logarithms of these dimensions are placed on the “0” line of the graph. For convenience, I always use the extant E. hemionus onager as a reference but using another reference would not change the observations at all. The dimensions of the material under study are also converted into decimal logarithms. Arithmetic differences between the reference logarithms and the logarithms of the studied dimensions are placed above the “0” line if they are positive (larger dimensions) or below if they are negative (smaller dimensions). In such a logarithmic diagram, the proportions remain unchanged whatever the absolute dimensions: the diagrams of two bones differing by their size but identical by their proportions will appear one above the other but on parallel lines. Details and examples are given at: https://vera-eisenmann.com/simpson-ratio-diagrams (accessed on 30 March 2009).
3. Taxonomy
Order Perissodactyla Owen, 1848
Family Equidae Gray, 1821
Subfamily Equinae Gray, 1821
Tribe Equini Gray, 1821
Genus Equus Linnaeus, 1758
Table S1 illustrates the taxonomic scheme followed in this article.
3.1. Diagnosis of Genus Equus
The long post-vomerine length (Basion to Hormion) relative to the overall palatal length (from Prosthion to Hormion), ([7], Figure S2).
Naso-incisival notch not reaching farther back than the level of P4.
Well-developed humeral proximal intermediate tubercule.
Shared with Allohippus and Plesippus are: protocone united to protoloph, monodactyly.
As far as possible, the subgenera listed below are based on associated cranial, dental, and limb bone characteristics. Preeminence is given to cranial features. Although this leads to uncertainties in the case of extinct taxa when no crania are available, species with markedly different cranial characteristics are not referred to the same subgenus even if their teeth and limb features are similar.
Descriptions and discussions of the various subgenera and taxa addressed here will be given in the following order:
1. Hemiones: E. (Hemionus) including E. hemionus, E. hydruntinus, and other fossil Hemiones.
2. Asses: E. (Asinus) including E. africanus, E. atlanticus, E. melkiensis, E. aff. africanus, E. graziosii, and E. apolloniensis.
3. Grevy’s zebras: E. (Dolihohippus) grevyi.
4. Plain’s zebras: E. (Quagga) including E. quagga, E. burchelli, E. mauritanicus, E. capensis, and E. oldowayensis.
5. Mountain zebras: E. (Hippotigris) zebra.
6. Horses: E. (Equus) including E. ferus, E. przewalskii, E. cf. scotti, E. chosaricus, E. germanicus, and E. antunesi.
7. Sussemiones: E. (Sussemionus) including E. coliemensis, E. verae, E. suessenbornensis, and E. aff. suessenbornensis from Akhalkalaki, Georgia.
8. E. altidens and the species group ‘Pseudohydruntines’ including: E. granatensis and E. aff. granatensis from Pirro, Italy, E. wuesti, E. hipparionoides, E. ovodovi, and a new species from Dmanisi.
9. Incertae sedis including, E. altidens, E. marxi, E. tabeti, E. nalaikhaensis, and Equus sp. from Konso, Ethiopia.
3.2. Affinities between Extant Subgenera
Affinities between extant subgenera may be schematically illustrated by two figures. The first one (Figure 1A) proposed by Bourdelle [19] is based on his anatomical observations of external and internal characters [20,21]. For example, Asses and Mountain zebras share some common characteristics: short intestines (especially small intestines); the anatomy of the false nostril, the larynx, and the thyroid. It is interesting that unexpected resemblances between Asses and Mountain zebras were also noted in a study of Equid DNA [22]. Figure 1B is a schematic representation of the first plane of multivariate analyses of 23 measurements taken on 342 crania of extant Equus [23]. Here, again, Mountain zebras appear close to Asses. The closeness between Plain’s zebras and Horses is illustrated by both Figure 1A,B. The affinity between Asses and Hemiones has been observed by all researchers and has led Groves and Grubb to unite them in the single genus Asinus ([24], pp. 13–15). Genetic studies indicate that Asses and Hemiones separated as long as 1.7 Ma ago [25,26], which may give some ground to consider them as distinct subgenera as indeed Groves himself has previously concluded at the end of a very detailed article [27].
3.3. Affinities between Sussemiones and Pseudohydruntines
In several past articles, I have referred several slender extinct Equus to the subgenus Sussemionus because of such particular teeth features as the frequency of stylids on the lower cheek teeth. These resemblances may, however, only mean that these ‘Pseudohydruntines’ are related to the subgenus Sussemionus without actually belonging to it.
4. Descriptions and Discussions
Since my Simpson’s diagrams always have E. (Hemionus) hemionus onager for reference, it is fitting to begin with a description of E. (Hemionus).
4.1. Hemiones
The extant and recently extinct hemiones range is limited to Asia and the Middle East. Fossil species reached Europe and possibly Africa. Neither fossil nor modern ones were found in the New World [29].
4.1.1. Extant Taxa
E. (Hemionus Cuvier 1823) hemionus Pallas, 1775 hemionus Pallas, 1775
E. (Hemionus) hemionus kulan Groves and Mazak, 1967
E. (Hemionus) hemionus onager Boddaert, 1795
E. (Hemionus) hemionus khur Lesson, 1827
E. (Hemionus) hemionus hemippus Saint Hilaire, 1855
E. (Hemionus) hemionus kiang Moorcroft, 1841
E. (Hemionus) kiang is considered by Vilstrup et al. 2013 ([30], p. 9) as an evolutionary distinct species but Bennett et al. ([26], p. 25) consider it ‘as a distinct population or even a metapopulation’ of E. hemionus hemionus; to my knowledge, their osteological characteristics do not differ much.
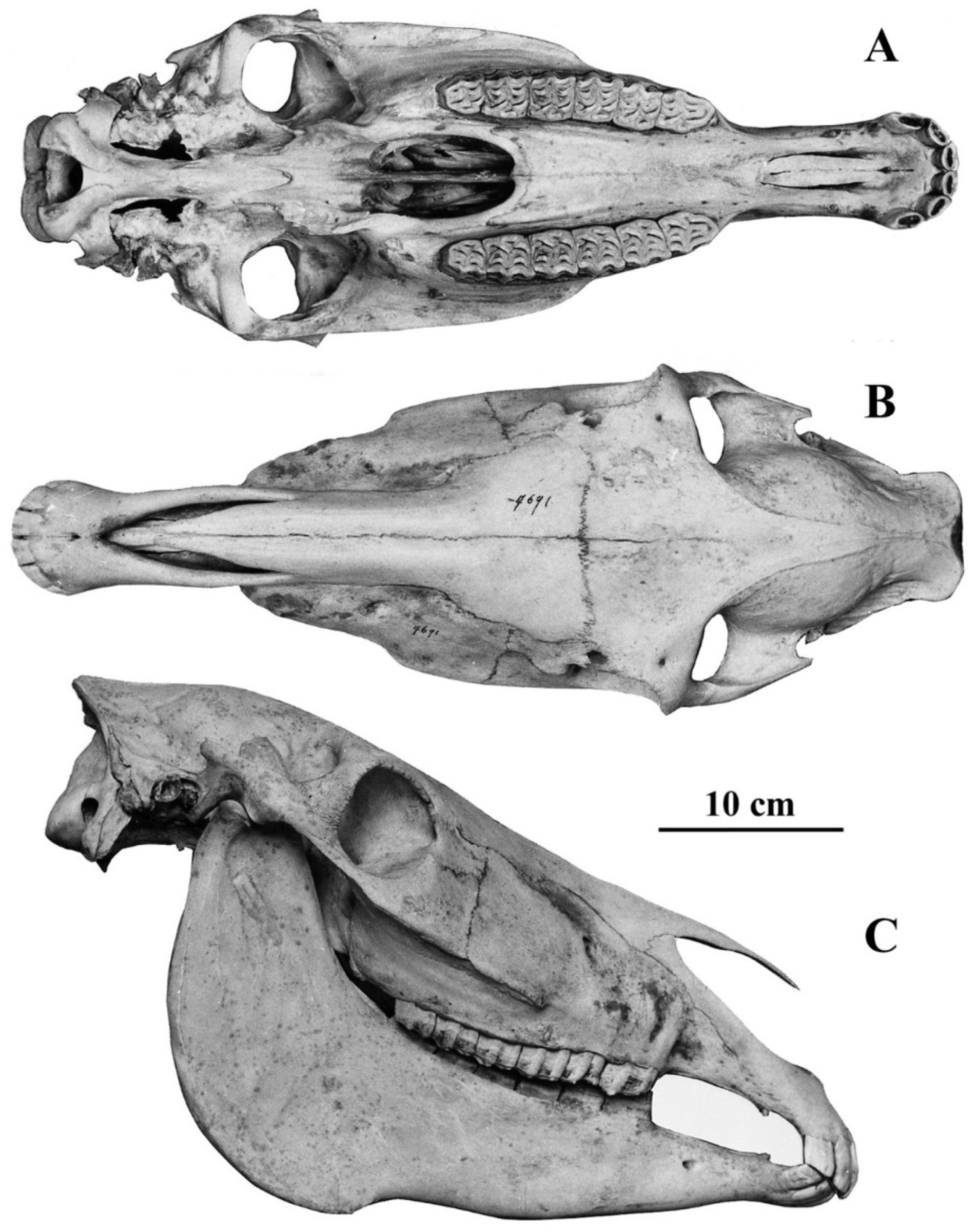
Figure 2.
E. (Hemionus) hemionus hemippus skull MCZ 6345, Lateral view.

Extremely high face; frontal breadth larger than bizygomatic breadth; broad muzzle both at the posterior borders of the third incisors and between the inter-alveolar borders; asinine Franck’s Index: vomerine length (distance from Staphylion to Hormion) longer than the post-vomerine length (distance between Staphylion and Basion); short choanae; narrow external occipital protuberance (= lambdoidal crest). Extreme hypsodonty. On the upper cheek teeth: usually long protocones and deep post-protoconal valleys. On the lower cheek teeth: shallow ectoflexids on molars as well as on premolars, with variable morphology of the double knot (see variants in ([29], Figures 3 and 4)). Well-developed infundibula in almost all the i1 and i2 and about 60% of the i3 [31]. Very slender limb bones, with deep diaphyses on the metapodials. An excellent cursorial adaptation is testified by the proportions of their segments: short proximal (humerus and femur) long distal (radius, tibia, and metapodials). In E. (Hemionus) hemionus and E. (Hemionus) hemippus, the third phalanges are especially narrow.
4.1.2. E. (Hemionus) hydruntinus Regalia, 1907
E. (Hemionus) hydruntinus Regalia, 1907 hydruntinus Regalia, 1907
E. (Hemionus) hydruntinus minor Bonifay, 1991
E. (Hemionus) hydruntinus petralonensis Tsoukala, 1991
E. hydruntinus is molecularly close to extant hemiones [32] but differs by some cranial and dental characters [33,34] The oldest cranium, referred to E. hydruntinus minor, found at Lunel Viel, France, is ca. 300 Ka [35]; unfortunately, it is fragmentary (Figure 5).
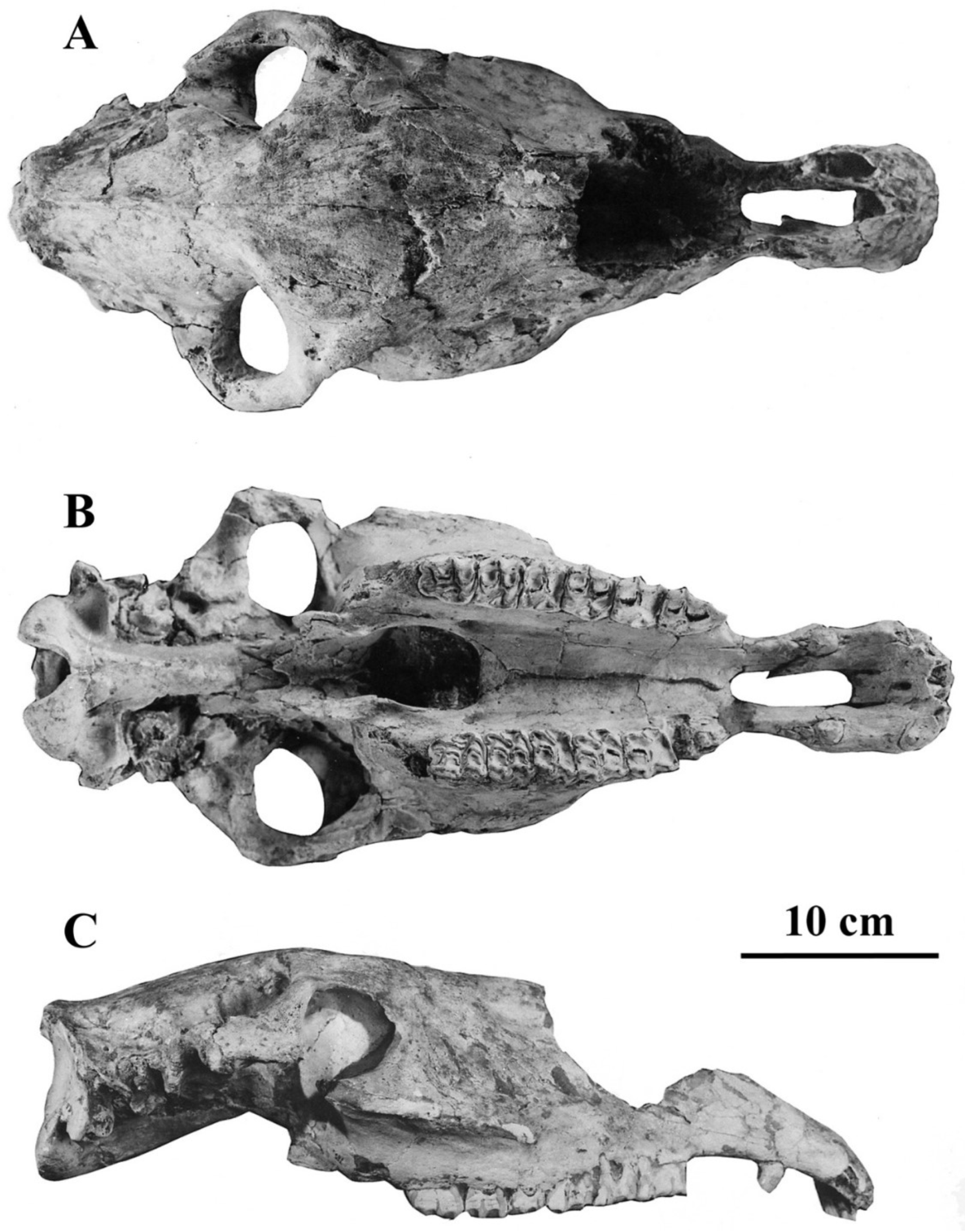
Figure 5.
E. (Hemionus) hydruntinus minor type cranium MNP-Bonifay LVIV 18698 from Lunel-Viel, France. Ventral view. Modified from Bonifay, M.-F. Equus hydruntinus Regalia minor n.ssp. from the caves of Lunel-Viel (Hérault, France) In Equids in the ancient world, vol. II, Eds. R.H. Meadow and H.-P. Uerpmann, Beihefte zum Tübinger Atlas des Vorderen Orients, Reihe A (Naturwissenschaften), Nr. 19/2, Dr Ludwig Reichert Verlag, Wiesbaden, Germany. 1991.
Figure 5.
E. (Hemionus) hydruntinus minor type cranium MNP-Bonifay LVIV 18698 from Lunel-Viel, France. Ventral view. Modified from Bonifay, M.-F. Equus hydruntinus Regalia minor n.ssp. from the caves of Lunel-Viel (Hérault, France) In Equids in the ancient world, vol. II, Eds. R.H. Meadow and H.-P. Uerpmann, Beihefte zum Tübinger Atlas des Vorderen Orients, Reihe A (Naturwissenschaften), Nr. 19/2, Dr Ludwig Reichert Verlag, Wiesbaden, Germany. 1991.

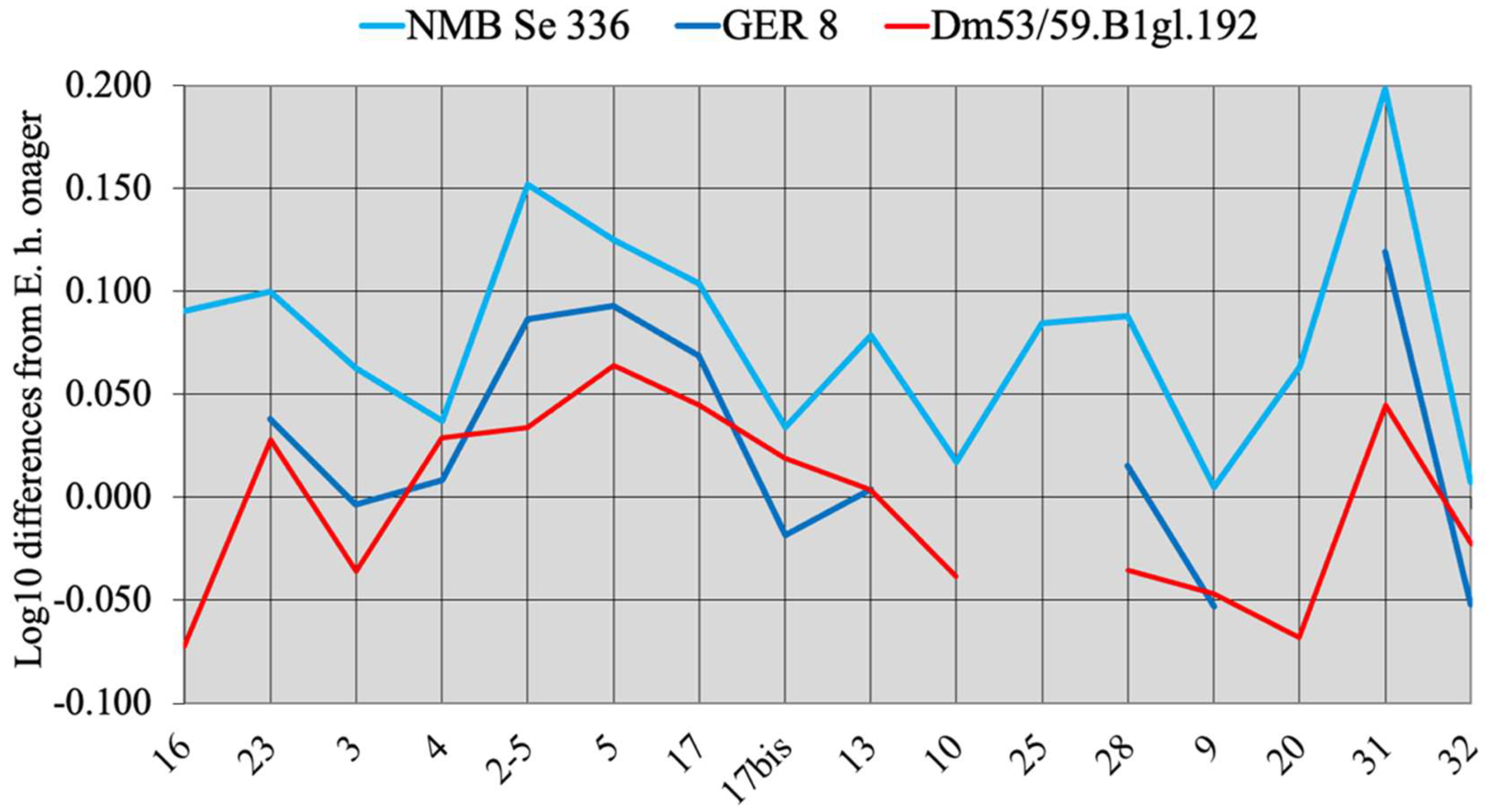
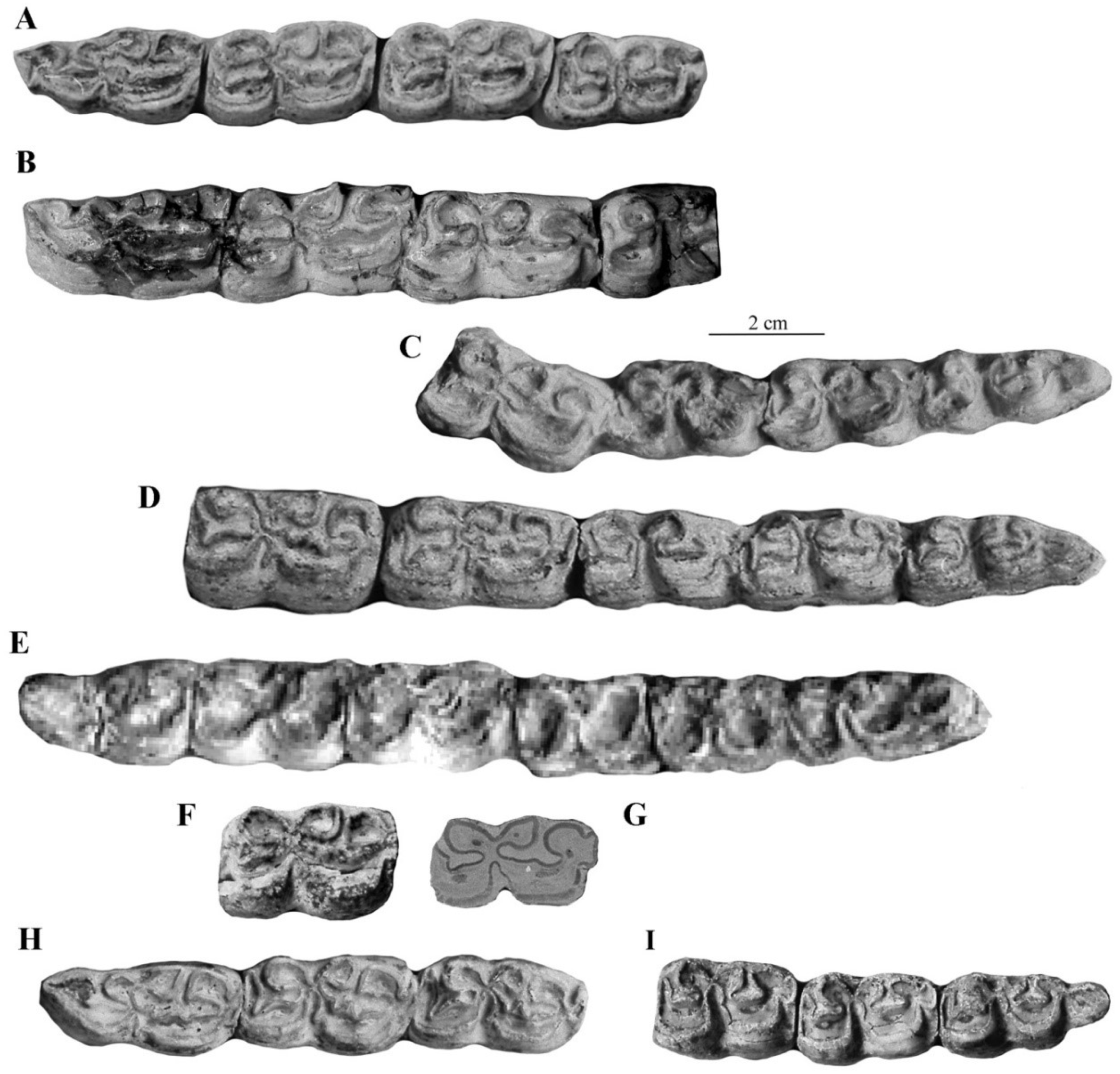
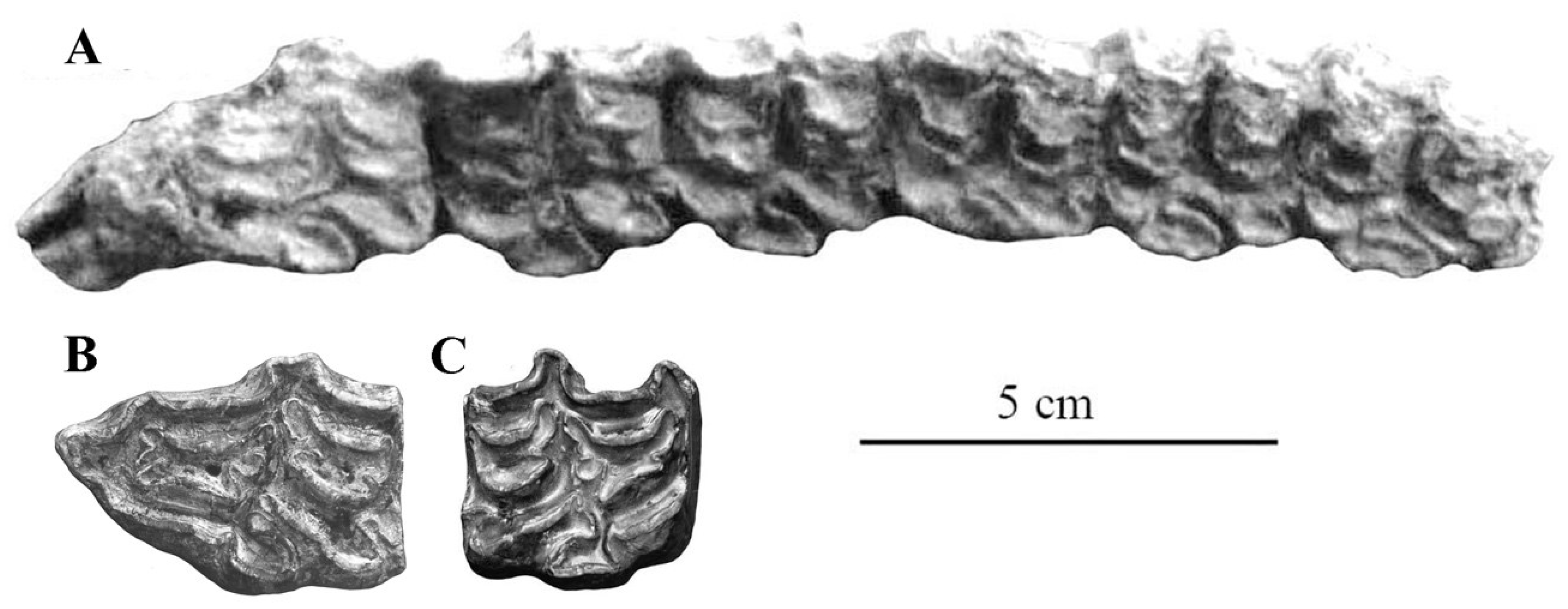
The best-preserved cranium was found in the Late Pleistocene Emine-Bair-Khosar Cave, Crimea, Ukraine [36]. Another cranium (Figure 6) originates also from the Late Pleistocene of Crimea (Kabazi). Both are remarkable by their wide muzzle (17, 17bis on Figure 6) and by their very short naso-incisival notch (31 in Figure 7) ([37], Figure 11)).
Most characteristic of E. hydruntinus are the short protocones of the upper cheek teeth and the deep ectoflexids of the lower molars (Figure 8).
Figure 6.
E. (Hemionus) hydruntinus cranium 1-05-21 from Kabazi, Russia, (A) Left lateral view. (B) Ventral view. Kindly communicated by Ariane Burke.
Figure 6.
E. (Hemionus) hydruntinus cranium 1-05-21 from Kabazi, Russia, (A) Left lateral view. (B) Ventral view. Kindly communicated by Ariane Burke.

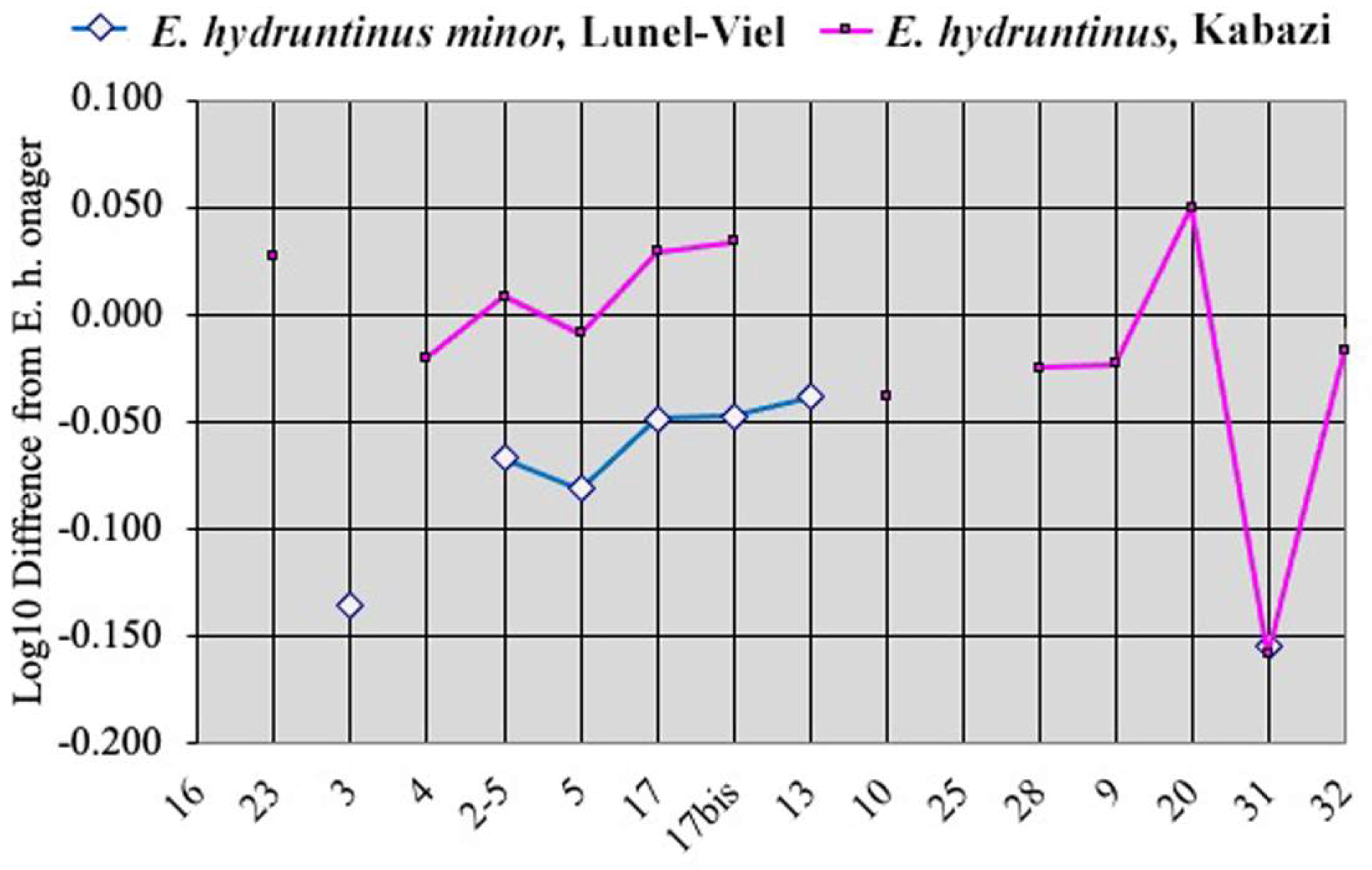
Figure 7.
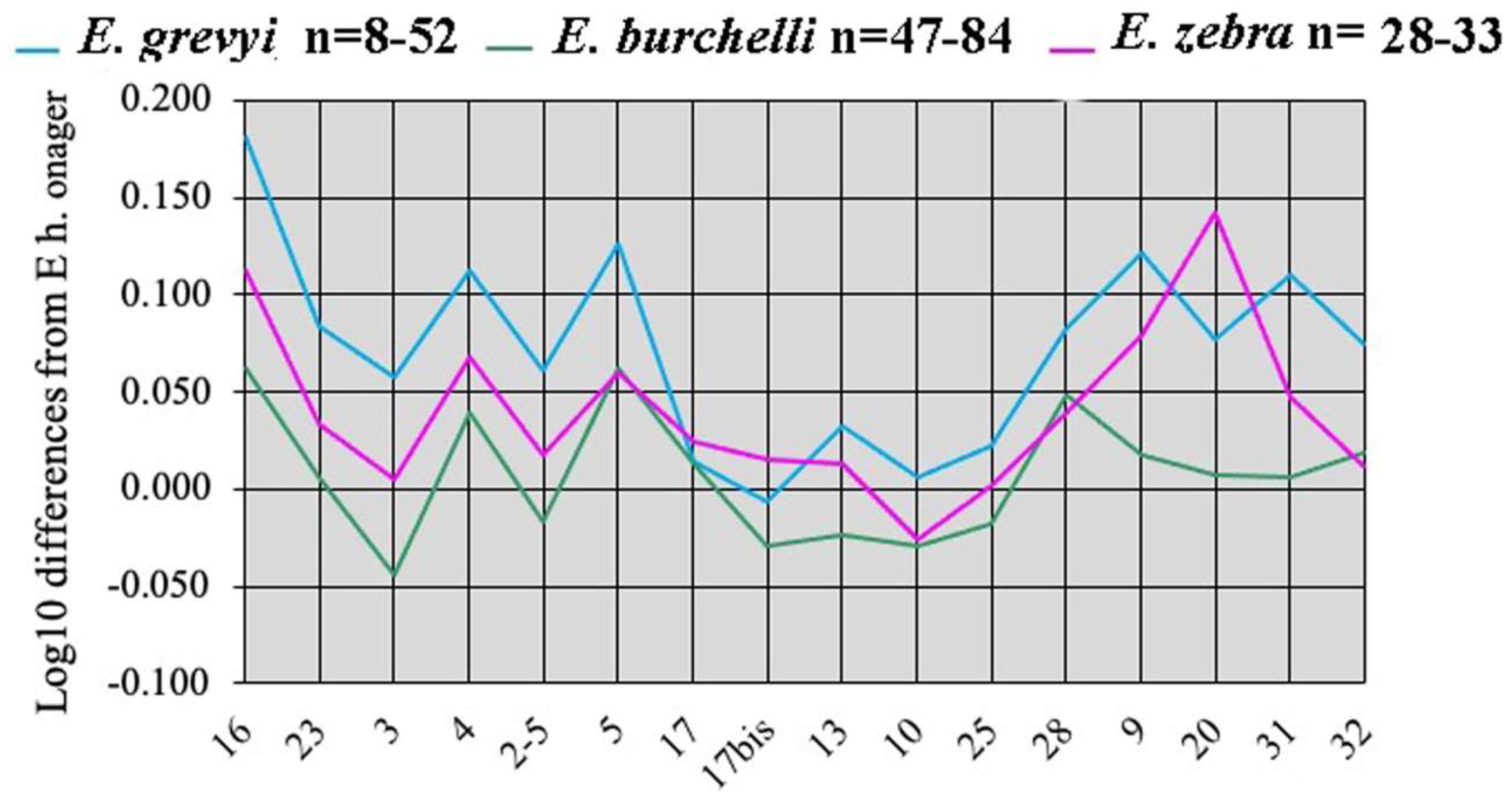
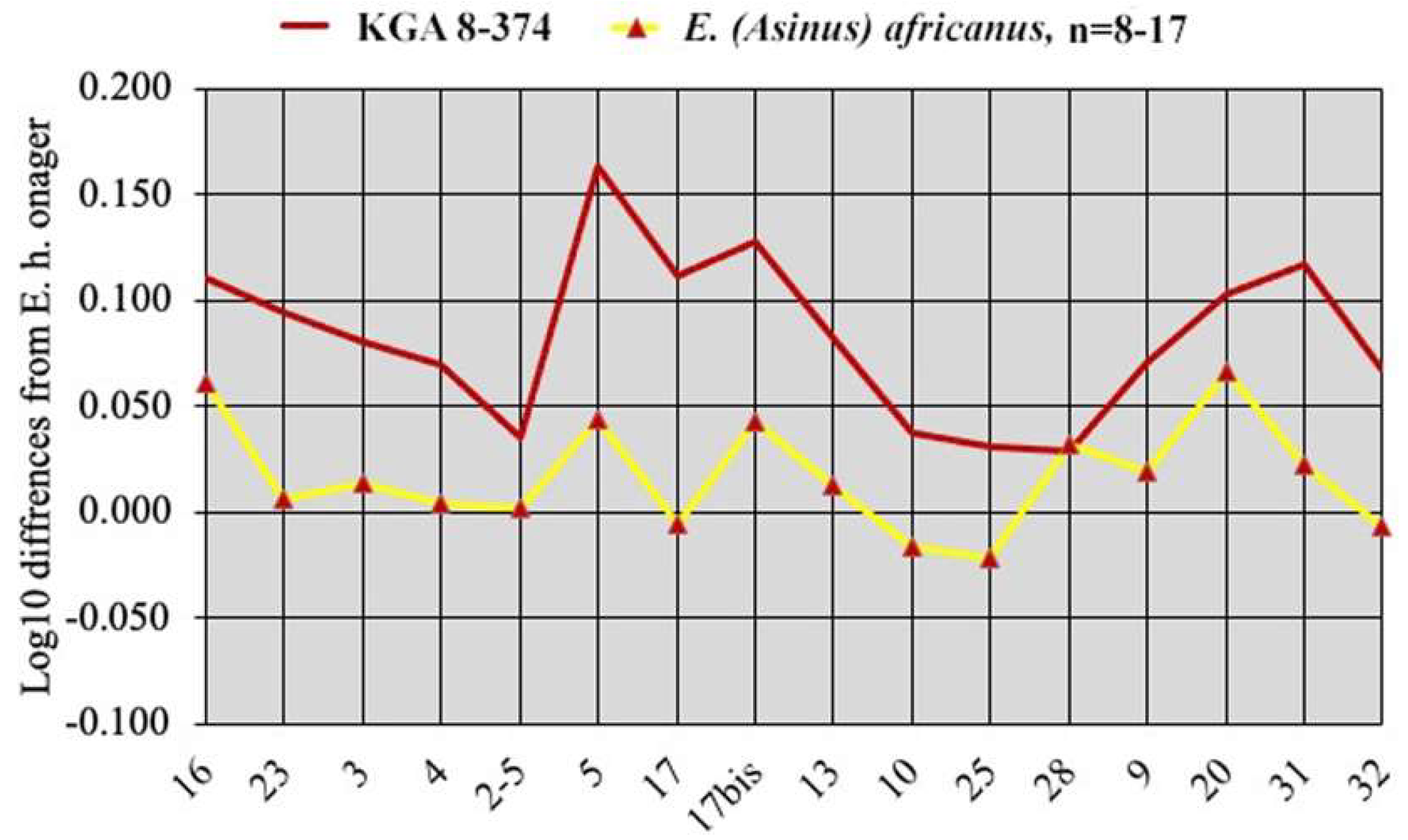
Simpson’s diagrams of E. (Hemionus) hydruntinus minor from Lunel-Viel and E. (Hemionus) hydruntinus) from Kabazi crania. 16: Breadth of the supra-occipital (lambdoidal) crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length.
Figure 7.
Simpson’s diagrams of E. (Hemionus) hydruntinus minor from Lunel-Viel and E. (Hemionus) hydruntinus) from Kabazi crania. 16: Breadth of the supra-occipital (lambdoidal) crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length.

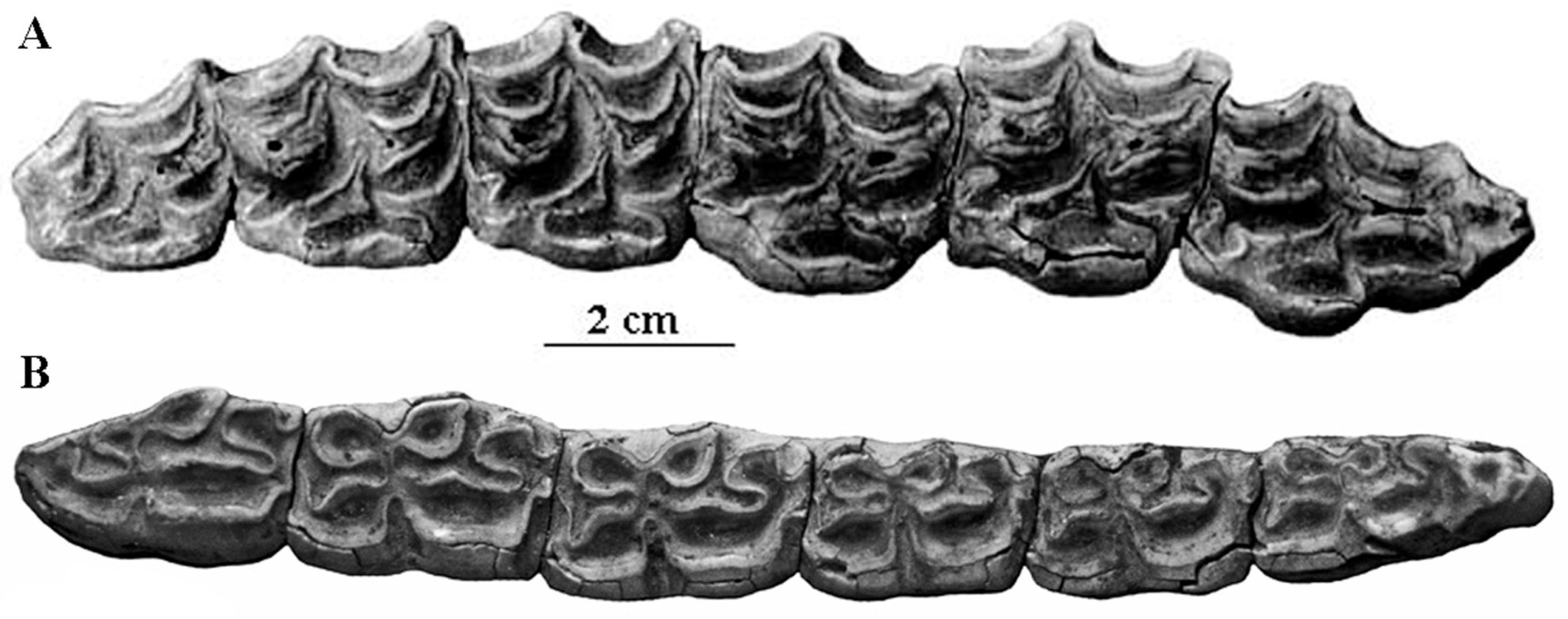
Figure 8.
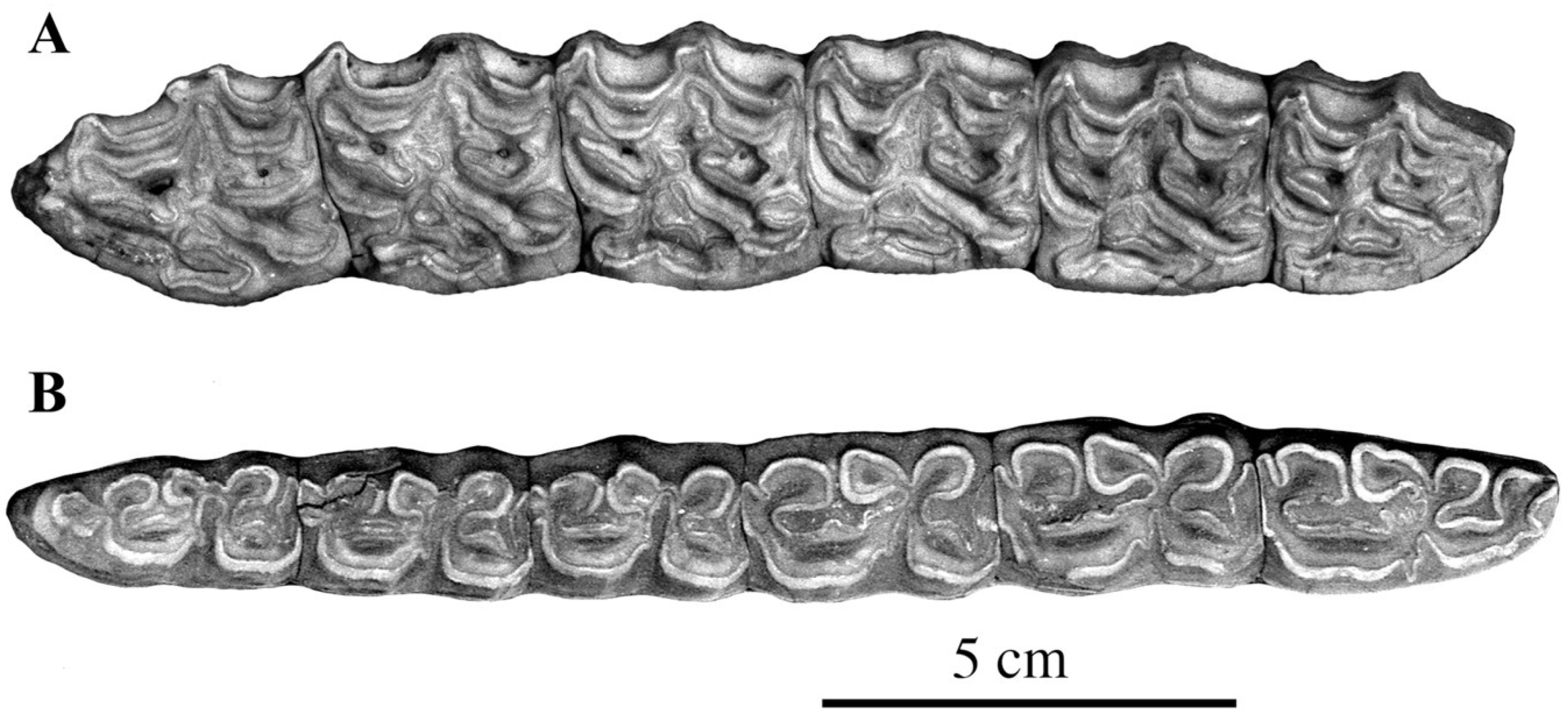
Occlusal views of cheek teeth of E. (Hemionus) hydruntinus from San Teodoro, Italy. (A) P3–M2 ST 98 PL 179; (B) p2–m3 ST04 PL 754. Courtesy by Gabriella Mangano.
Figure 8.
Occlusal views of cheek teeth of E. (Hemionus) hydruntinus from San Teodoro, Italy. (A) P3–M2 ST 98 PL 179; (B) p2–m3 ST04 PL 754. Courtesy by Gabriella Mangano.

Another characteristic is the microdonty (Figure 9): compared to the length of metapodials (MC1 and MT1), the dimensions of occlusal surfaces (occlusal length + occlusal width)/2 of P3–P4/(P size) and of M1–M2/(M size) are small except in the oldest form of Lunel-Viel. The small size of the protocones (P prot. and M prot.) is also well illustrated in the same figure.
Figure 9.
Simpson’s diagrams of E. (Hemionus) hemionus and E. (Hemionus) hydruntinus metapodials and teeth proportions. MC: third metacarpal; MT: third metatarsal; 11: Distal articular breadth; 4: Diaphysis depth; P: Upper P3 and P4; M: Upper M1 and M2; size: (occlusal length + occlusal width)/2; prot.: Protocone length. The number of specimens may be found in Table S2.
Figure 9.
Simpson’s diagrams of E. (Hemionus) hemionus and E. (Hemionus) hydruntinus metapodials and teeth proportions. MC: third metacarpal; MT: third metatarsal; 11: Distal articular breadth; 4: Diaphysis depth; P: Upper P3 and P4; M: Upper M1 and M2; size: (occlusal length + occlusal width)/2; prot.: Protocone length. The number of specimens may be found in Table S2.

It must be noted that all these dental characteristics are shared by some quite older—Late Villafranchian–Early Galerian—Equids such as those of Pirro, Italy and Aïn Hanech, Africa. Morphologically, E. hydruntinus could easily be considered a close relative or even a descendant of these taxa if they belonged to Equus instead of Allohippus or Plesippus as is commonly accepted [38]. However, the crania from Pirro and Aïn Hanech are unknown, and the cranium from Kabazi is close to the extant Equus (Hemionus).
There were several subspecies of E. hydruntinus.
The best represented are the smallest, oldest (and less typical) E. hydruntinus minor of Lunel Viel, France, one of the largest—E. hydruntinus of Petralona, Greece [39] and the ‘type’ E. hydruntinus hydruntinus of Romanelli, Italy [40]. E. hydruntinus was widely distributed over Eurasia during the Pleistocene: Roterberg and Senzig (Germany), Agios Georgios, Petralona (Greece), Dorog (Hungary), Tabun, Quneitra (Israel), Romanelli, San Teodoro (Italy), Prolom, Staroselie (Russia), and many other localities in France, Great Britain, Portugal, and Spain. It survived in the Holocene and perhaps may have been present at historical times in Portugal [41]. In Africa, a few cheek teeth very probably belonging to E. hydruntinus were described from the Hagfet et-Tera Cave, Lybia by Blanc [42], and an MC from Salé, Morocco is almost identical to the one from Pair-non-Pair, Würm II, France (Figure 10).
Figure 10.
Simpson’s diagrams of third metacarpals of E. hydruntinus from Pair-non-Pair, France and from Salé, Morocco. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.
Figure 10.
Simpson’s diagrams of third metacarpals of E. hydruntinus from Pair-non-Pair, France and from Salé, Morocco. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.

4.1.3. Other Fossil Hemiones
As they were described in detail in [29] I will just list them here with brief remarks.
E. (Hemionus) binagadensis [34] from the final Riss or Riss-Würm of Azerbaidjan differs so much from other E. (Hemionus) that it may belong to a true species ([29], p. 172). The cranium is small with a very short muzzle, and the post-vomerine distance is longer than in hemiones. The metacarpals are slender and deep in the diaphysis and the proximal epiphysis; the metatarsals have wide distal articular breadths.
Some fossils somehow intermediate between extant hemiones and E. (Hemionus) hydruntinus were found in the Paleolithic of Transbaikalia [43,44] at Tologoj.
Hemione-like metapodials were found in the Late Pleistocene of Kurtak, South-Central Siberia [45].
Several skulls, an associated skeleton, and some limb bones were found in the middle or late Paleolithic levels at Sjara-osso-gol, Mongolia [46]. They may have belonged to E. (Hemionus) hemionus hemionus.
A fragmentary cranium, much smaller than that of Sjara-osso-gol, comes from Jiling, Yushu, China. It has a very high face. The post-protoconal valleys are very deep.
Metacarpals from the Late Pleistocene Gulongshan Cave, Dalian, China were studied by Zhou et al. [47]. They are similar to E. (Hemionus) hydruntinus but slightly larger.
4.2. Asses
E. (Asinus) Gray, 1824 (Asses)
E. (Asinus) asinus Linné, 1758
E. (Asinus) africanus africanus Heuglin and Fitzinger, 1866
E. (Asinus) africanus Heuglin and Fitzinger, 1866 somaliensis Noack, 1884
E. (Asinus) atlanticus Thomas, 1884
E. (Asinus) melkiensis Bagtache, Hadjouis and Eisenmann, 1984
E. (Asinus) graziosii Azzaroli,1966
E. (Asinus) lauracensis Astre, 1948
E. (Asinus) apolloniensis Koufos et al., 1997
The extant wild Ass E. (Asinus) africanus is strictly limited to Africa. However, more or less similar fossils were found in Europe, such as E. (Asinus) graziosii and E. (Asinus) lauracensis, in the Middle East and even in Asia. Asinine characteristics occur frequently in fossil species.
A thorough review of North-African Ass-like forms [48] discusses the Atlas wild Asses (E. atlanticus, E. melkiensis, and E. tabeti). I do not think that E. tabeti, which will be addressed later, is an Ass. However, it seems that there were in North Africa many Ass-like fossils. Moreover, there is a not yet formally described Ethiopian species possibly related to Asses and the enigmatic South African E. lylei.
The first Asses really resembling the extant ones are poorly documented and of Holocene age: one metatarsal at Tell Muraibit, Syria [49] and one at Maysar, Oman [50].
In France, Astre [51] described a new species of Ass, E. lauracensis based on a very well-preserved and fossilized metacarpal most similar to an MC of the small domestic Asses of Cameroon [28], p. 73. It may have belonged to the enigmatic Zebro of the Iberic Peninsula [41].
4.2.1. E. (Asinus) africanus Heuglin and Fitzinger, 1866 (Figure 11)
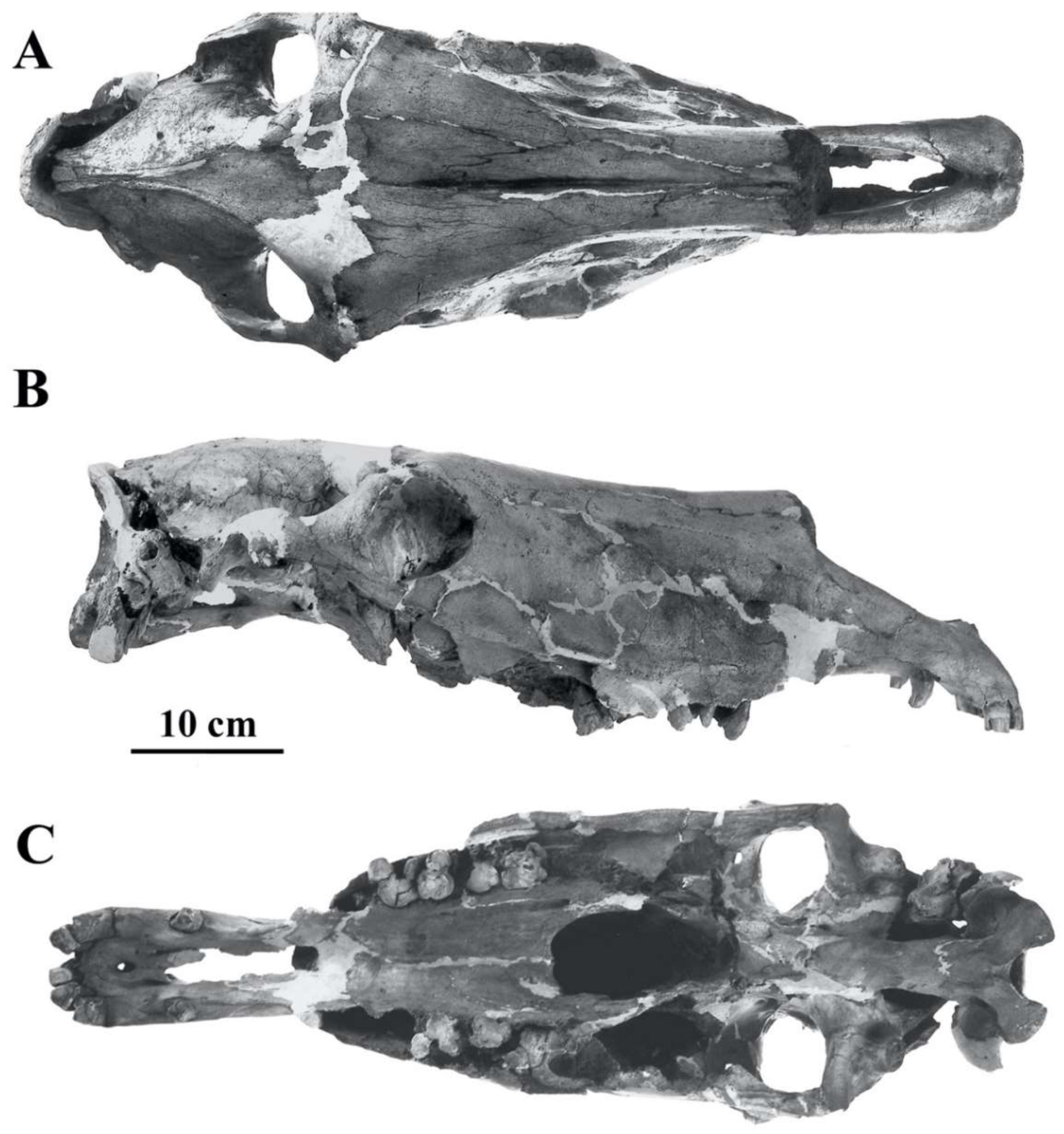
Description
Very wide supra-occipital crest (measurement 16), frontal breadth larger (measurement 13) than bizygomatic breadth (measurement 14), vomerine length (measurement 3) longer than the post-vomerine length (measurement 4), very wide muzzle between the inter-alveolar borders (measurement 17bis), cranium high behind the orbits (measurement 28), and very large external auditory meatus (measurement 20). In the somaliensis subspecies, the muzzle (measurement 5) is longer than in the africanus one.
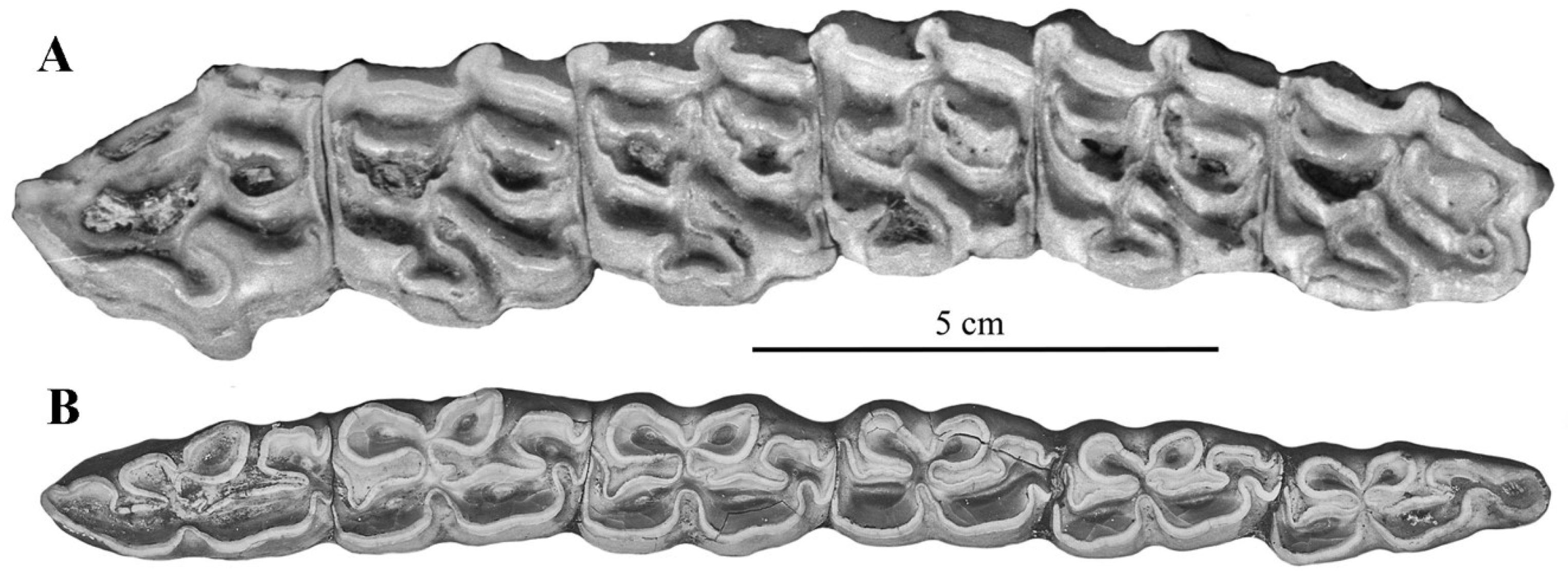
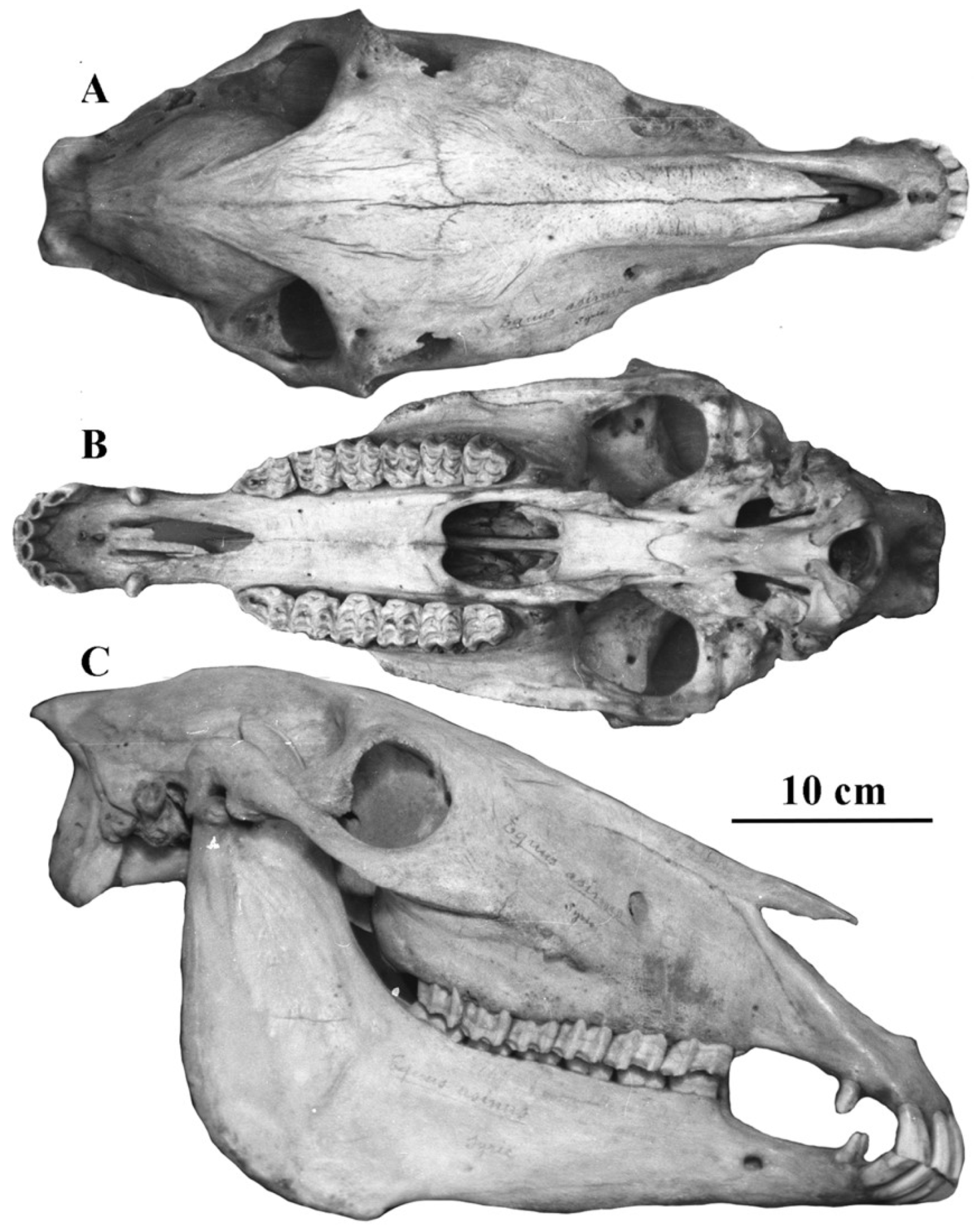
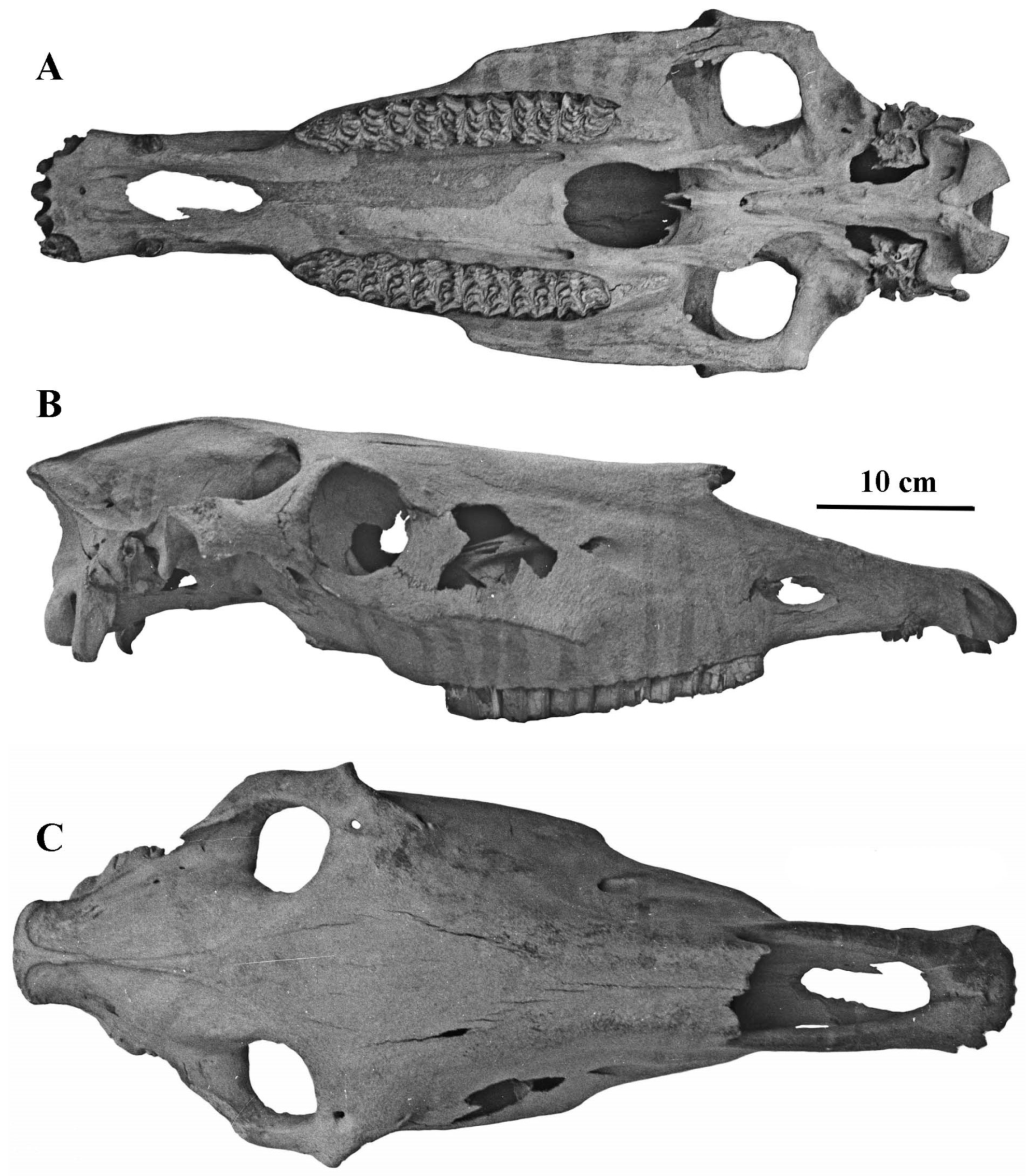
Figure 11.
E. (Asinus) africanus somaliensis skull ZIN 7204, (A) dorsal view, (B) Ventral view, (C) Right lateral view.
Figure 11.
E. (Asinus) africanus somaliensis skull ZIN 7204, (A) dorsal view, (B) Ventral view, (C) Right lateral view.

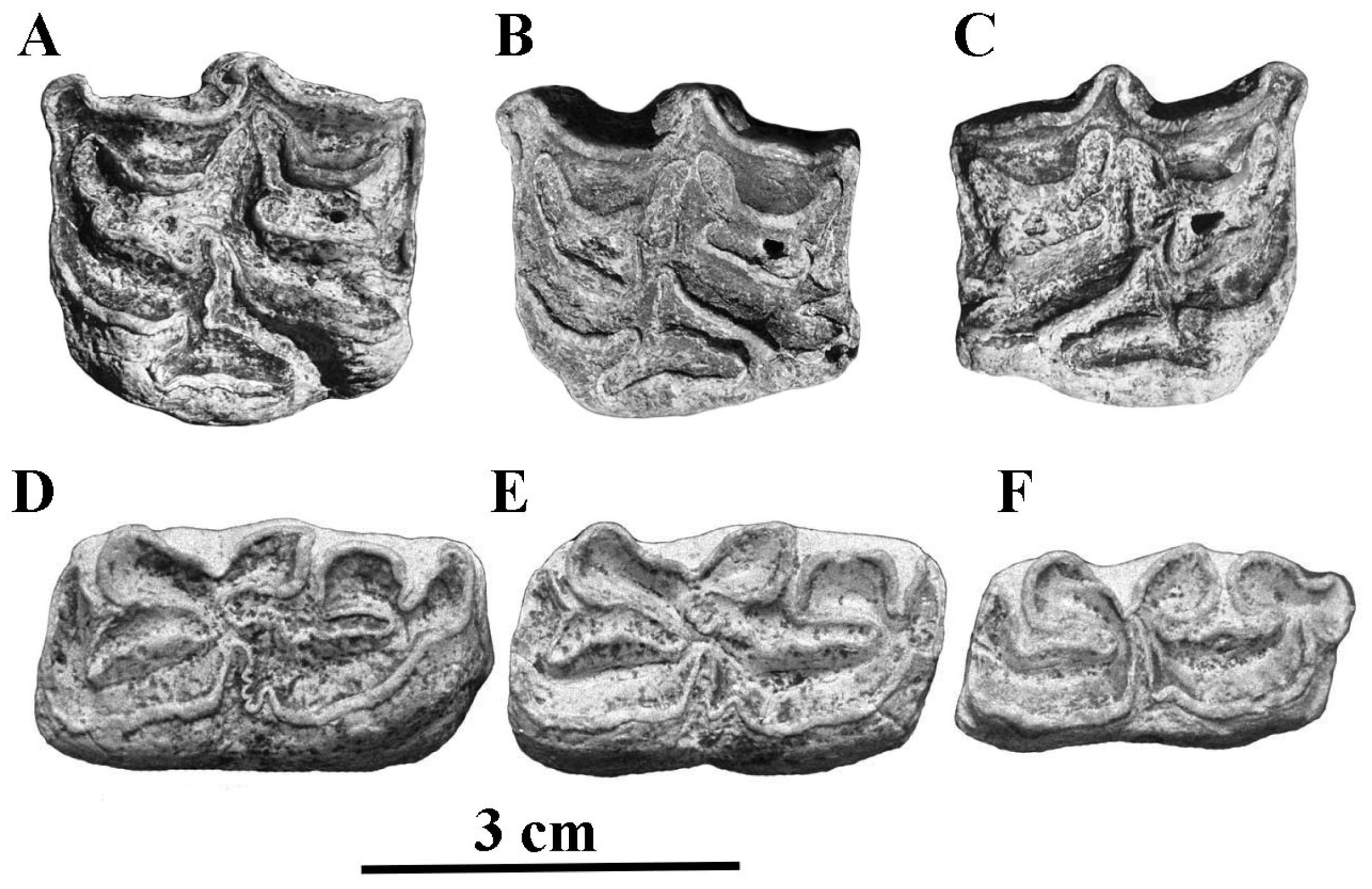
On the upper cheek teeth, the post-protoconal valleys are shallow, unlike in hemiones; the protocones are shorter in E. africanus than E. hemionus (Figure 12) and shorter in E. africanus africanus than in E. africanus somaliensis. Plis caballin occur in around 50% of P3–P4 and 33% of M1–M2. Hypoconal islets (Figure 13) occur in 7 M3 out of 11 [52].
Figure 12.
Scatter diagram of upper P3–P4 and M1–M2 occlusal dimensions in E. (Asinus) africanus and E. (Hemionus) hemionus.
Figure 12.
Scatter diagram of upper P3–P4 and M1–M2 occlusal dimensions in E. (Asinus) africanus and E. (Hemionus) hemionus.

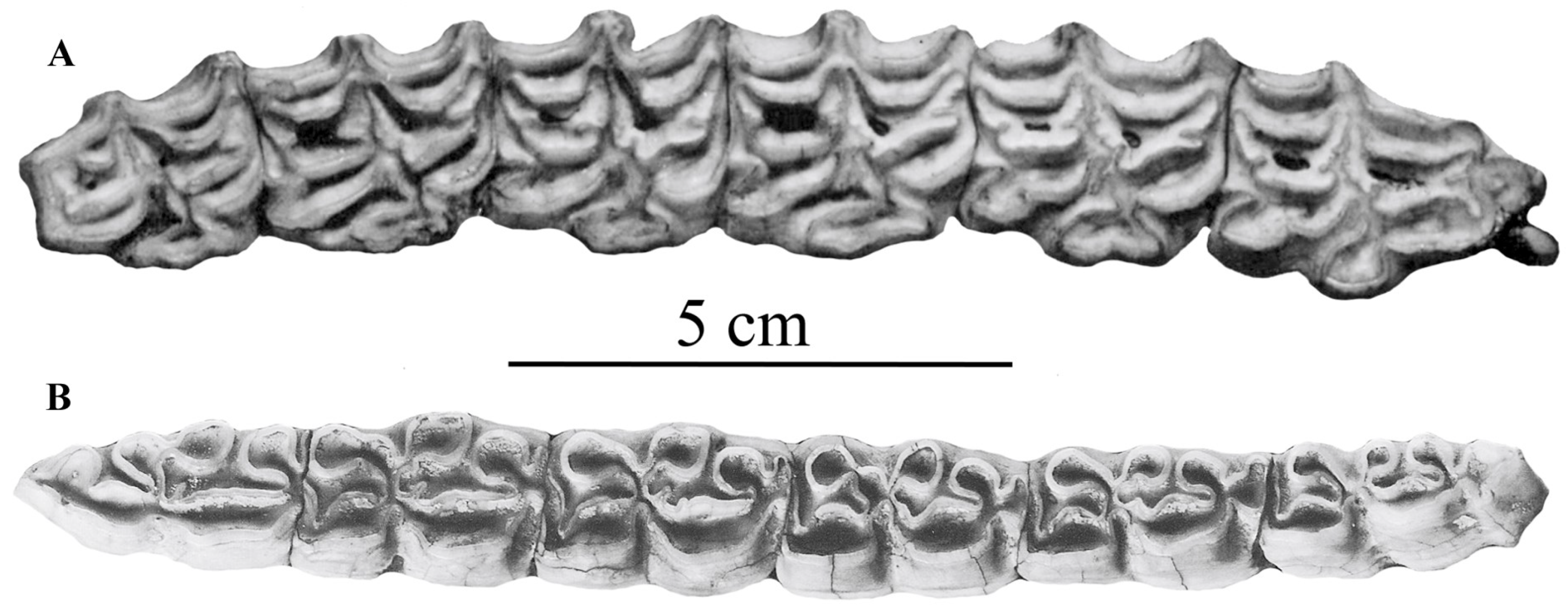
Figure 13.
Occlusal views of upper and lower cheek series of E. (Asinus) africanus africanus. (A) NHMUK-ZD 1939.4780: P2–M3. (B) NHMUK-ZD 1935.5.7.1: p2–m3.
Figure 13.
Occlusal views of upper and lower cheek series of E. (Asinus) africanus africanus. (A) NHMUK-ZD 1939.4780: P2–M3. (B) NHMUK-ZD 1935.5.7.1: p2–m3.

On the lower cheek teeth, the metaconids are more developed than metastylids, especially in E. (Asinus) africanus somaliensis (Figure 14). The ectoflexids (vestibular grooves) are shallow both in lower premolars and molars. Plis caballinid are more frequent in premolars than in molars. [53]. The ‘bridge’ between the metaconid and metatstylid noted by Groves [54] is frequent, especially on M1. Well-developed infundibula appear in lower i1, less so in i2, and even less in i3 [31], altogether less developed than in hemiones.
Figure 14.
Occlusal views of upper (A) and lower (B) cheek teeth series of E. (Asinus) africanus somaliensis MNHN–MO 1977-65.
Figure 14.
Occlusal views of upper (A) and lower (B) cheek teeth series of E. (Asinus) africanus somaliensis MNHN–MO 1977-65.

Limb bones (see vera-eisenmann.com accessed on 3 November 2007): MC and MT less slender than in E. (Hemionus) hemionus onager; MC with deeper diaphyses, and larger proximal epiphyses. MT with wider proximal epiphyses and larger distal articular breadths. The proportions of the limb bone segments are similar to those of E. (Hemionus) hemionus onager except that the metapodials are shorter, and the third phalanges are as narrow as in E. (Hemionus) hemionus hemippus.
4.2.2. E. (Asinus) atlanticus Thomas, 1884
The type specimen [55] is a lower juvenile mandible (Figure 15) found inside the lower clay of Oued Seguen (near Constantine, Algeria) close to a cranium of Bos primigenius mauritanicus.
Figure 15.
E. (Asinus) atlanticus, type mandible from Oued Seguen, Algeria. Modified from [55], occlusal view.
Figure 15.
E. (Asinus) atlanticus, type mandible from Oued Seguen, Algeria. Modified from [55], occlusal view.

The dp2–dp4 series is approximately 92 mm long. A well-developed pli protostylid is present on the dp2. An isolated stylid is also present at the postero-vestibular corner of the dp3. On the dp3 and dp4, the metaconids are elongated and more developed than the metastylids. On the dp3, the metaconid is bilobated. On all the teeth ectoflexids are shallow and plis caballinid are present. The referral of this mandible to an Ass was contested by Boule, who observed a similar enamel islet on the dp3 of an E. burchelli ([56], Figure 4) and proposed an attribution to the latter.
I have no knowledge of the frequency of enamel islets on the dP3 in either species. According to my data [53], the presence of a pli protostylid is as frequent in E. africanus as in E. burchelli, and the dimensions of the dp2–dp4 are also the same (approximately 85 mm). However, two features seem to support Thomas’ attribution: the elongated metaconids and the shallow ectoflexids. Still from Oued Seguen, Thomas also refers to Asses some limb bones and two adult lower cheek teeth series. One of them is 160 mm long; the other (from a very old individual) is only 148 mm long. These dimensions are quite larger than the average of E. africanus (mean: 104.5 mm, max:115) or E. burchelli (101 mm).
Since neither the adult teeth nor the limb bones of this Ass were figured nor measured in detail, it seems difficult to consider E atlanticus as a valid species, at least until the fossils mentioned by Thomas are found and studied.
4.2.3. E. (Asinus) melkiensis Bagtache, Hadjouis, Eisenmann, 1984
E. (Asinus) melkiensis was described from fossils found at the Aterian site of Les Phacocères, usually known as ‘Les Allobroges’, in Algeria near Algiers [57].
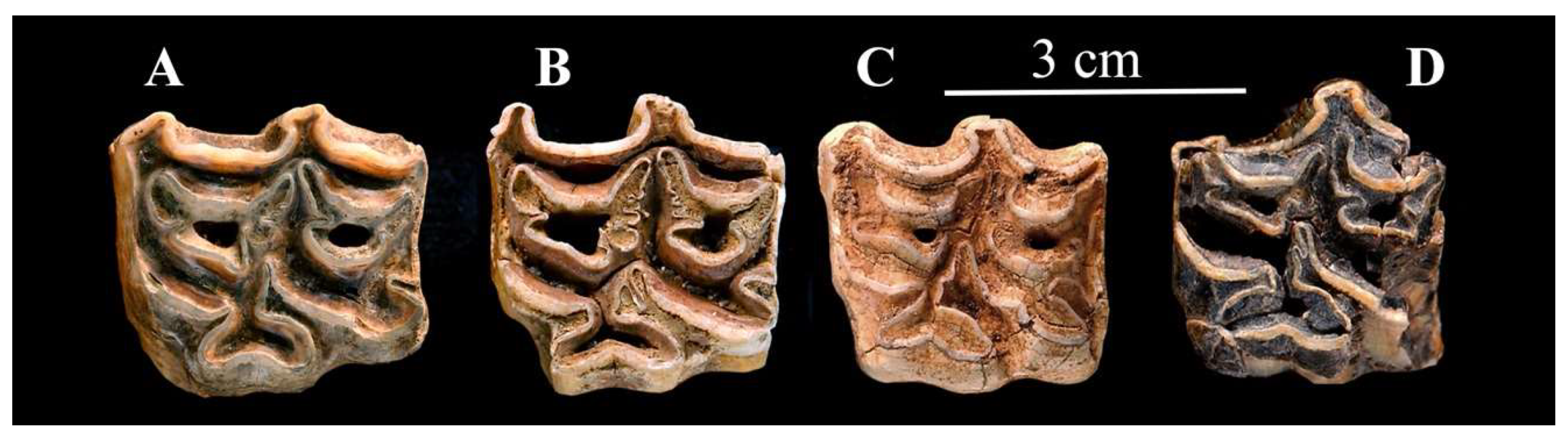
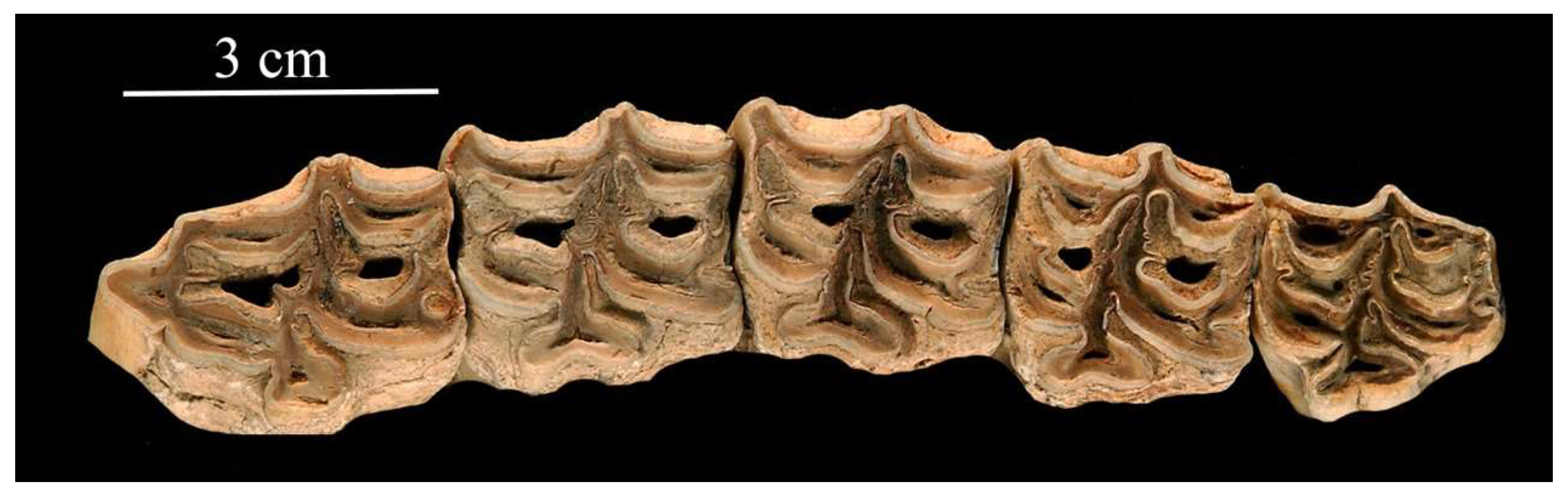
In his remarkable review, Sam gave a list of north-western localities where Ass-like fossils, in particular E. (Asinus) melkiensis, have been found ([48], Table S1). I will illustrate here some fossils morphologically similar to E. (Asinus) melkiensis from Filfila [58] and Aïn Benian [59], Algeria, from Bou-Knadel [60] and Mugharet El Alya [61], Morocco, and also from Yemen, Oumm Qatafa, Israel [62,63] and Petralona, Greece [39].
The type specimen is a third metacarpal Allo. 61-1314, (Figure 16A,B); the paratypes a third metatarsal (Allo. 61-1837, Figure 16D) and a lower M2 (Allo. 61-1969, Figure 17F), all preserved in the IPH collections. A few more cheek teeth very probably belong to the same species while other specimens belong to a smaller Ass (see below) and some to the caballine E. algericus. There are also several indifferently preserved limb bones.
The post-protoconal groove is deep, especially on the P2 (Figure 17A) the protocone of which is very short. On the other upper cheek teeth (Figure 17B–D) the symmetrical protocones bear a pronounced lingual groove; small plis caballin are present.
Similar enamel patterns may be seen on upper premolars in Algeria at Filfila, (Figure 18A), in Morocco at Aïn Tit Mellil, Sidi Bou Knadel, Sidi Abderahmane (Figure 18B,C, Figure 19 and Figure 20). They are also present on upper premolars from an unknown locality in Yemen (Figure 18D), Aïn el Guettar, Tunisia (Figure 21C), from Oum Qatafa, Israel (Figure 21B), and from Petralona, Greece (Figure 21A).
Associated upper and lower cheek teeth series were found by Yves Coppens in Tchad (#282 in the MNHN-F collections). The lowers (Figure 22A) have elongated metaconids but not the ‘bridges’ found in E. (Asinus) africanus somaliensis and E. (Asinus) melkiensis. The upper cheek teeth (Figure 22B) have deep post-prorotoconal valleys, thus resembling E. (Asinus) melkiensis.
The metaconid of the lower molar from Les Allobroges (Figure 17F) is very elongated. On the other lower molar, the ‘bridge’ described by Groves [54] is present between the metaconid and metastylid. On both teeth, the ectoflexids are shallow (Figure 17E,F). A lower premolar from Aïn Tit Mellil (Figure 23) shows a similar ectoflexid pattern.
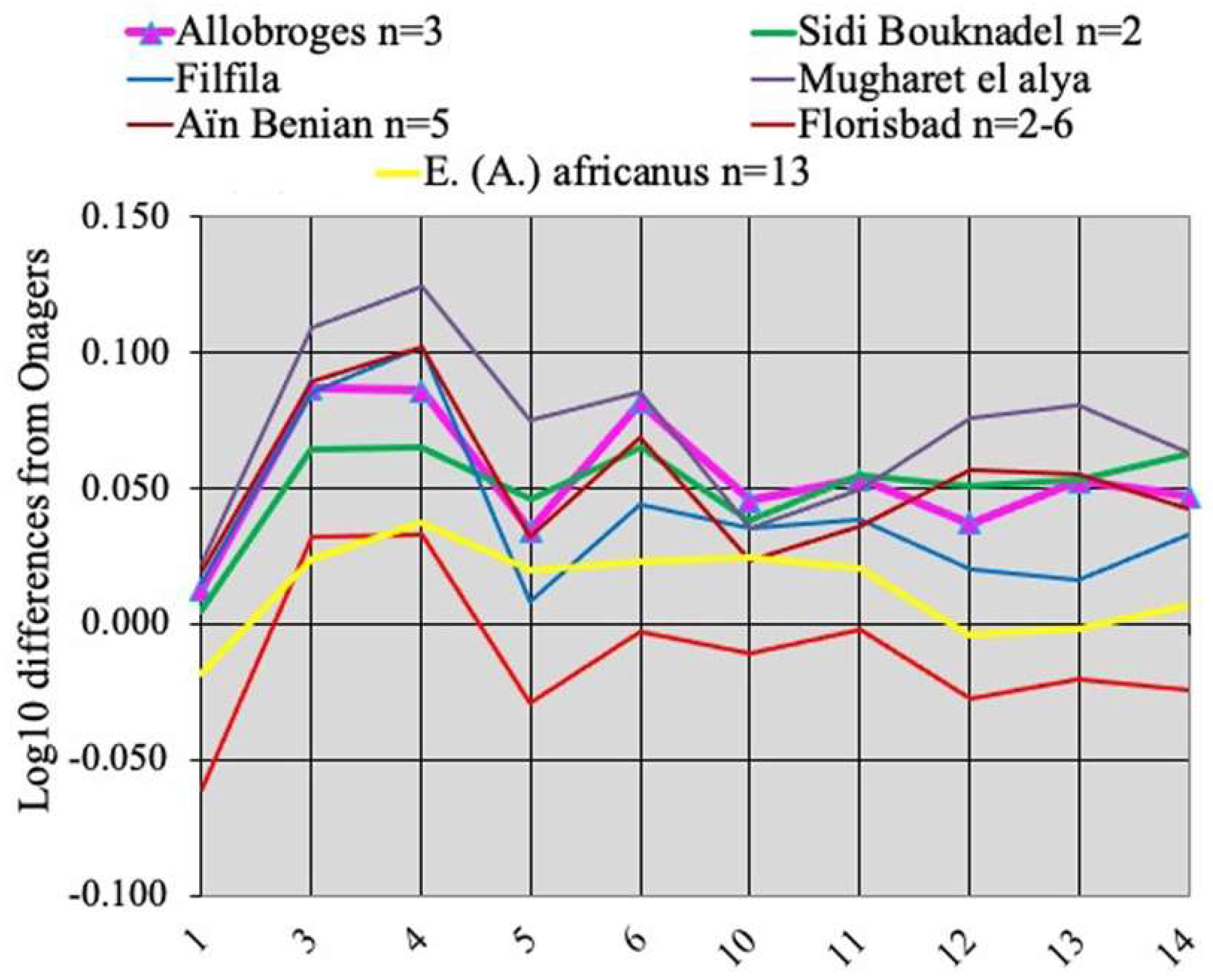
The metapodials of E. melkiensis do not resemble those of extant Wild Asses; they are much more robust and have smaller proximal articular breadths (measurement 5) and deeper proximal epiphyses (measurement 6). Filfila and Aïn Benian (Algeria), and Sidi Bou Kndel, and Mugharet El Alya (Morocco) have similar MCs; so do the MCs of E. lylei from Florisbad, South Africa but their size is smaller (Figure 24). The best-preserved MT from Les Allobroges has a deep proximal articular surface but a large distal articular breadth like E. (Asinus) africanus. Unfortunately, the fossil MTs from Aïn Benian, Algeria) and Aïn Tit Mellil, Morocco (see data at vera-eisenmann.com (accessed on 13 March 2020)) are too badly preserved to be compared with E. (Asinus) melkiensis.
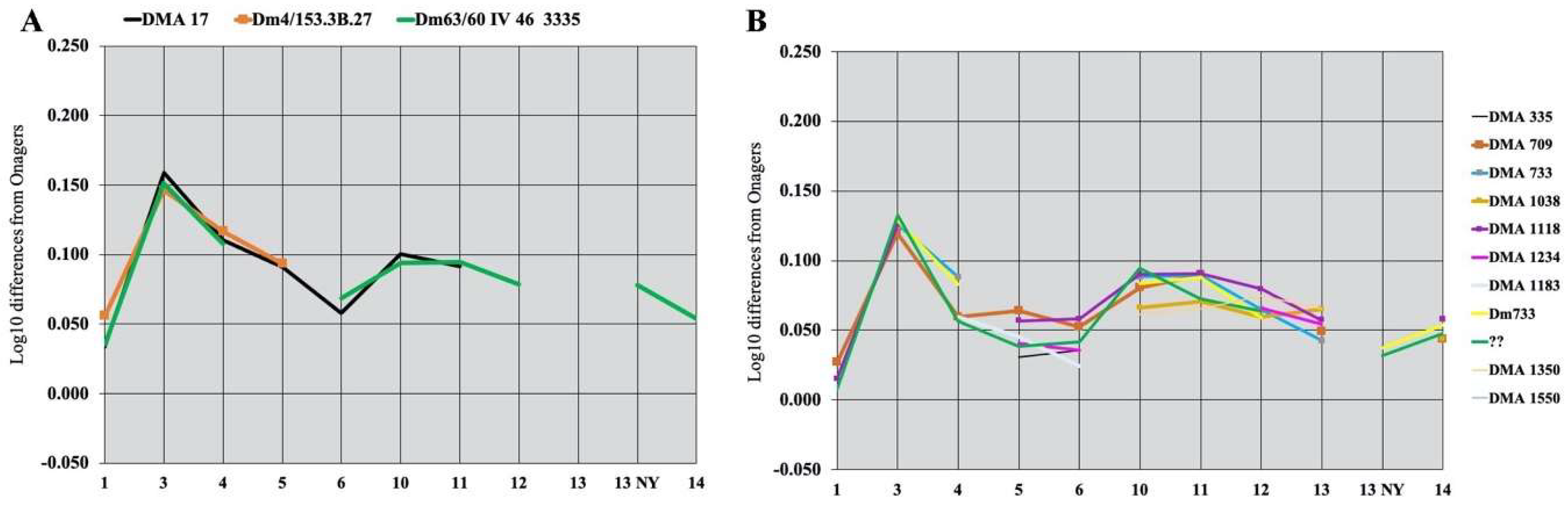
The earliest evidence in Africa for a form possibly ancestral to E. melkiensis (E. cf. melkiensis) is found at Tighenif, Algeria: one third metacarpal (MNHN-F Ter 404) has the proportions of E. melkiensis, but one third metatarsal resembles more Wild Asses. In Asia, upper cheek teeth resembling E. melkiensis (Figure 25) were found at Lakhuti II (Loc. 67), Tadjikistan, believed to belong just below the Brunhes–Matuyama boundary [64,65].
4.2.4. E. (Asinus) asinus Linné, 1758 and E. (Asinus) sp.
Although like all domestic animals, domestic Asses are very polymorph, they have some common features.
Description
The average cranium (Figure 27) is smaller than in E. (Asinus) africanus but has similar proportions (Figure 28). In the somaliensis subspecies, the muzzle (measurement 5) is longer than in the africanus and the asinus ones. The same figure shows also the differences between Asses and hemiones since the reference zero line is E. hemionus onager.
The upper and lower cheek teeth (Figure 29) are not very different from E. (Asinus) africanus. However, on the lower cheek teeth, the metaconids are more rounded and not larger than the metastylids. Well-developed infundibula appear in the lower i1, less so in the i2, and even less in the i3 [31].
In North Africa, many fossils present these characteristics in the enamel pattern of cheek teeth and the proportions of limb bones, not quite the same as in E. africanus or E. melkiensis.
A lower cheek series from Aïn Benian (= Guyotville), MNHN-F Guy 118 is similar to E. (Asinus) asinus: the ectoflexids are shallow and the metaconids rounded (Figure 30); the age may be Aterian [67].
Several lower cheek teeth from the Aterian of Les Allobroges have the same pattern (Figure 31).
From Aïn Metherchem, Tunisia, Middle Paleolithic [68], there are associated upper and lower cheek teeth series (Figure 32). The upper resemble E. (Asinus) asinus but the lower have more elongated metaconids. I refer them to E. (Asinus) sp.
A few badly preserved MTs from the Aterian of Aïn Tit Mellil [67] and Tihodaïne (ca 0.4 Ma, according to Van Couvering and Delson [69]), differ from E. (Asinus) africanus and E. (Asinus) asinus by their deep diaphyses (Figure 33). From Aïn Metherchem, a slender MC preserved at the IPH has also a deep diaphysis ([63], Figure Supplement C). More detailed studies of North African Equids are needed to understand where these Ass-like forms belong.
Figure 33.
Simpson’s diagrams of E. (Asinus) sp. MT. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.
Figure 33.
Simpson’s diagrams of E. (Asinus) sp. MT. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.

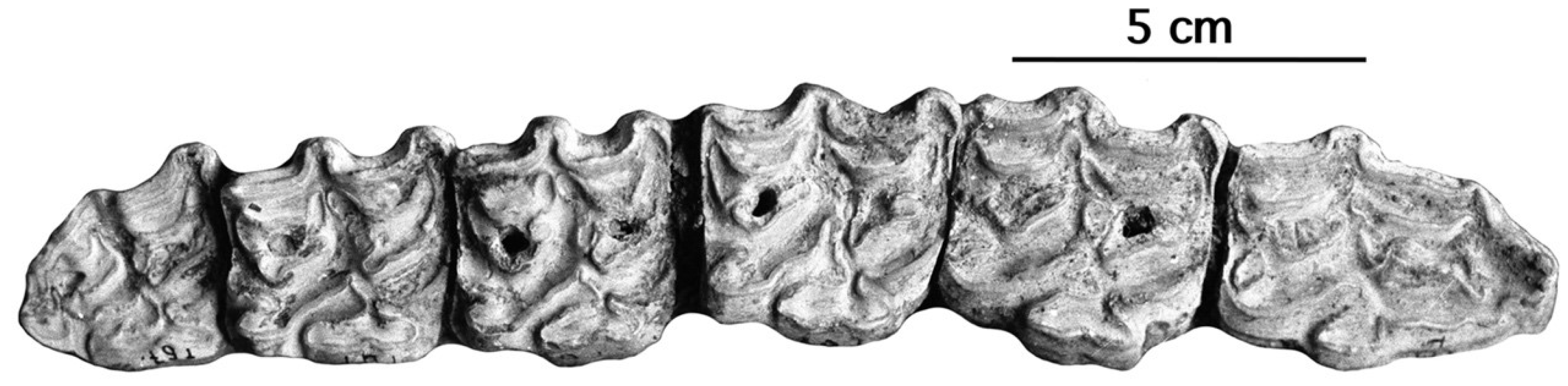
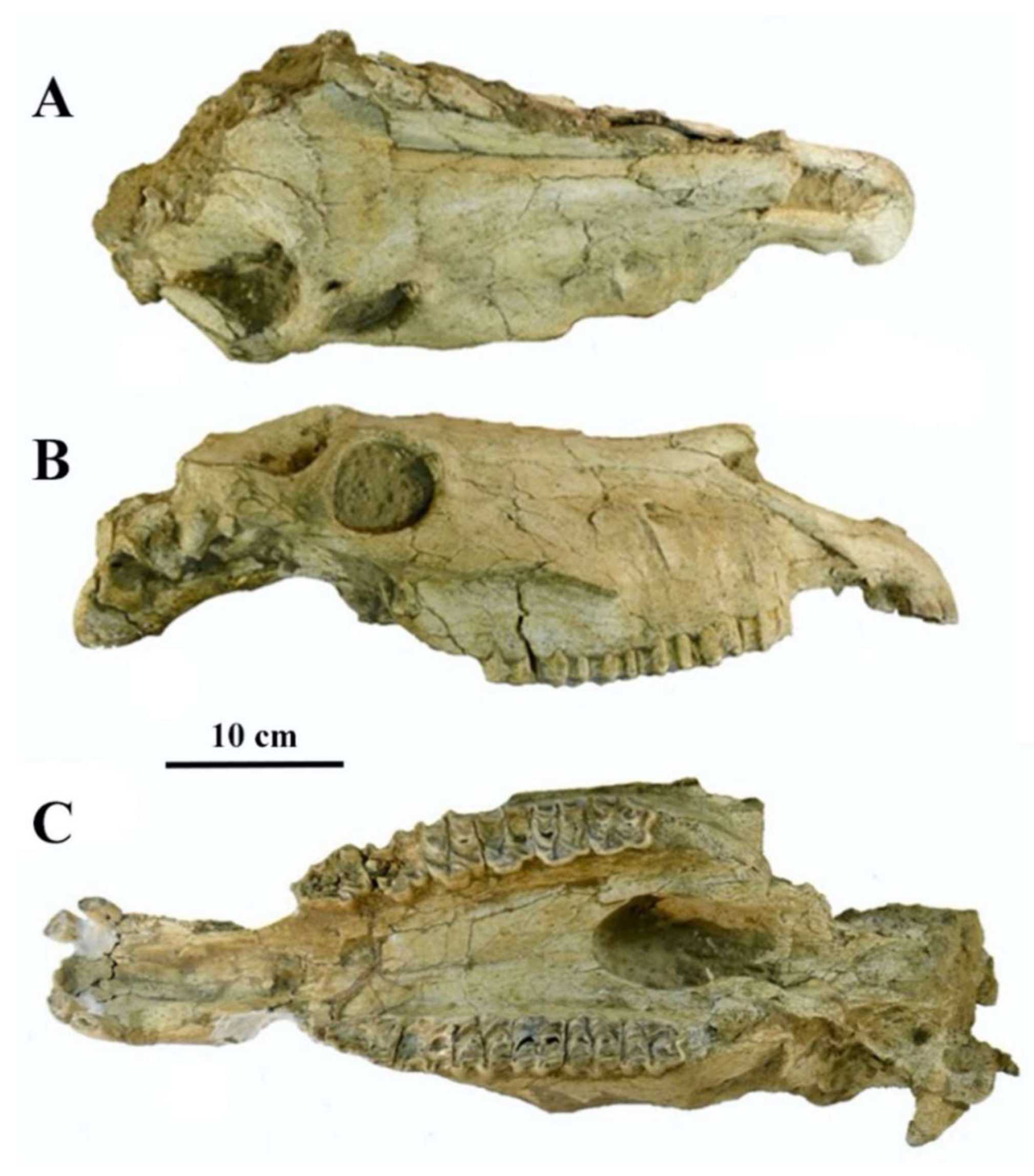
4.2.5. E. graziosii Azzaroli, 1966
From the Late Pleistocene of Maspino, Italy, Azzaroli [70,71] described E. graziosii as a new species of fossil Ass, based on the anterior part of a cranium, IGF 192V (Figure 34).
The cranium of E. graziosii shares with domestic and wild extant Asses most of its proportions except the muzzle breadth between the inter-alveolar borders (Figure 35 measurement 17bis).
Except for the P2, the cheek teeth (Figure 36) have deep post-protoconal grooves and symmetric bilobated protoconids, just like E. melkiensis, but the protocones are much longer.
The enamel pattern of the lower cheek teeth of the mandible referred to E. graziosii ([71], p. 7) is that of a horse.
It may be that E. melkiensis will be found to be a junior synonym of E. graziosii. However, pending lower cheek teeth and metapodials are found for the latter and/or cranium for the former I prefer to consider them as sister species, both possibly related to the more ancient Asiatic form from Lakhuti II (Figure 24).
4.2.6. E. (Asinus) apolloniensis Koufos et al., 1997
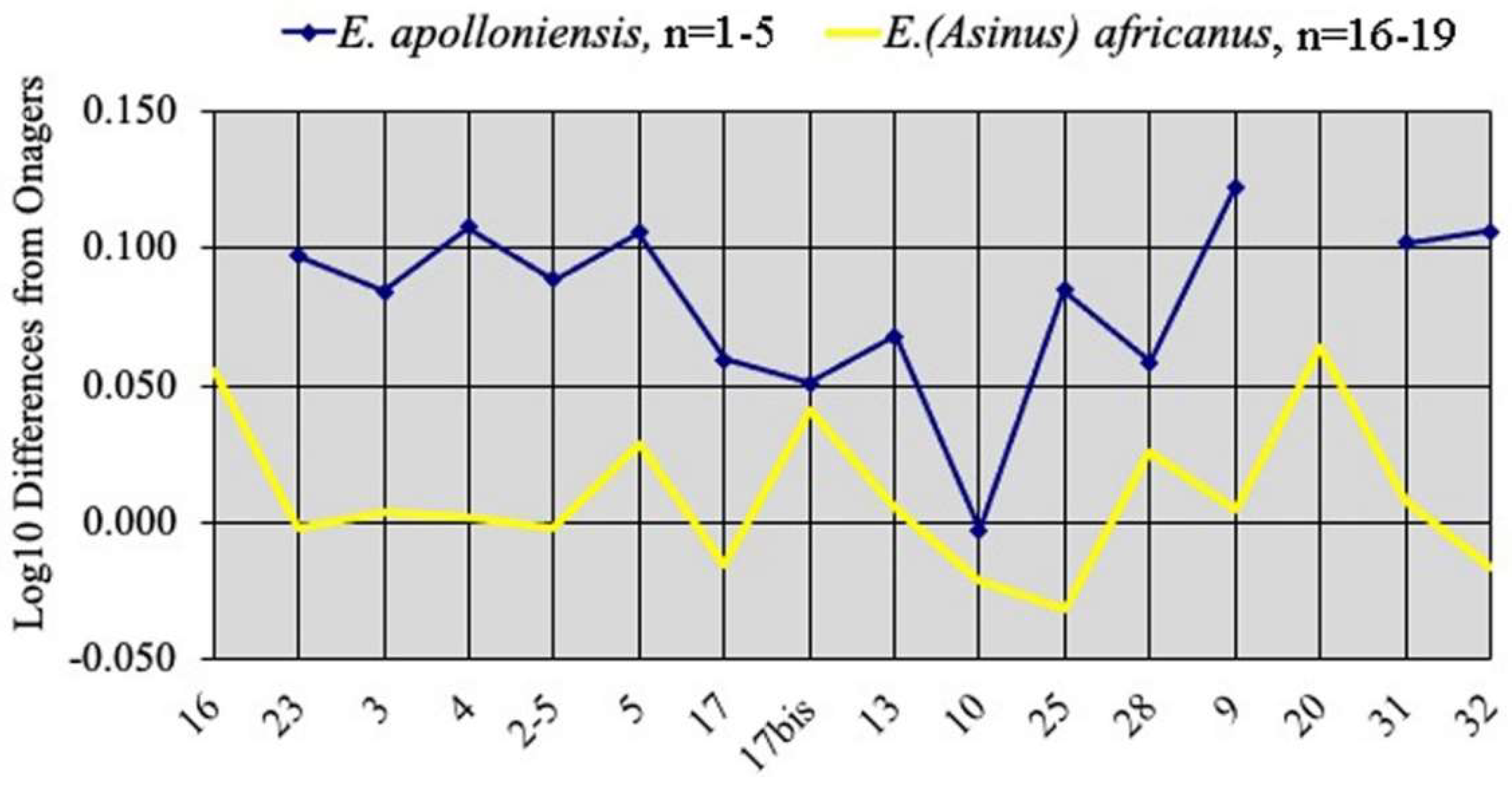
E. apolloniensis was described [72] from the Apollonia P-1 of the Platanochori Formation of Mygdonia Basin, Greece. The large mammal assemblage is believed to be circa 1.2 Ma old ([73], Figure 1). Equids are well represented; unfortunately, most crania are distorted, rendering measurements difficult to make and to trust. I have palliated this problem as much as I could by collating the data published [14,73] and checking them with the photographs (Table S3). Figure 37 shows two of the less distorted crania. Another two specimens APL 518 and APL 129 seem too large to belong with the rest.
Although comparisons by way of Simpson’s diagrams are difficult, some important features may, however, be observed (Figure 38). The average cranium of E. apolloniensis is mostly characterized by its narrow choanae (measurement 10). The palate length–muzzle length–muzzle breadth at the I3 posterior borders (measurements 2-5, 5, 17) proportions are similar to those of Asses. However, E. apolloniensis differs from Asses by a smaller inter-alveolar breadth of the muzzle, a higher face, and a smaller cranial height at the posterior borders of the orbits (measurements 17bis, 25 and 28).
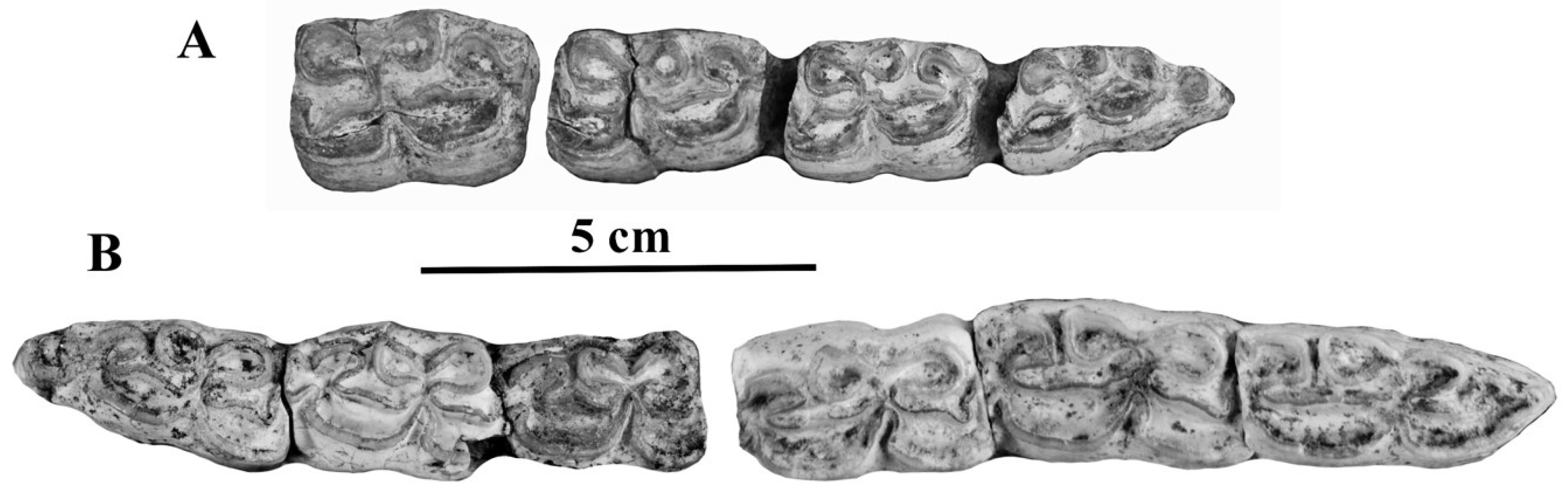
The upper cheek teeth (Figure 39) resemble those of E. (Asinus) africanus africanus: short protocones, some of them symmetric and bilobated.
The lower cheek teeth of APL 103 and 171 (Figure 40) have also asinine features: elongated metaconid on p3 and shallow ectoflexids, at least on m2 and m3.
In an article describing new fossil Equids from Apollonia and revising and comparing the whole material [14], it was suggested that ‘possibly E. apolloniensis, evolved in Europe and belonged to stenonoid lineage’. I have explained at the beginning of this article why Equus could not evolve from Allohippus, whether locally in Europe or elsewhere. However, whatever its origin, E. apolloniensis was a true Equus and somehow related to Asses.
4.3. E. (Dolichohippus) grevyi Oustalet, 1882
Description
The largest extant wild Equus. Very elongated cranium due to a very long muzzle and long choanae, convex forehead, naso-incisival notch slightly shorter than cheek length, long and large external auditory meatus, and very wide supra-occipital crest (Figure 41).
Compared to other zebras’ crania (Figure 42), the E. grevyi cranium is larger, with a narrower muzzle (measurements 17 and 17bis). From E. burchelli, it differs by longer choanae (measurement 9); from E. zebra, by a smaller external auditory meatus (measurement 20).
Upper cheek teeth plicated with well-developed plis caballin, long, asymmetric, and indented protocones. Lower molars with rounded metaconids and metastylids and deep vestibular valleys; lower premolars with rounded metaconids, shallow vestibular valleys, and plis caballinid; very frequent plis protostylids on p2 [74], (Figure 43B).
Well-developed cups, at least on i1 and i2 [31]. The metapodials are slender and deep in the diaphysis. More cursorial limb bone proportions than Asses and other Zebras (Figure 44).
O’Brien et al. [13] have described a cranium of E. (Dolichohippus) grevyi from the Middle Pleistocene Kapthurin Formation, Kenya, aged between 547 and 396 Ka. They also recorded and discussed the “proposed fossil occurrences of E. grevyi” and conclusively argued that most of them are not characteristic enough to be referred to that taxon and certainly not those older than ca. 500 Ka.
4.4. E. (Quagga) Shortridge, 1934 (Plain’s Zebras)
Of all zebras, extant Plain’s zebras are the ones most resembling Horses [19,23] apart from the pattern of the cheek teeth.
E. (Quagga) quagga Boddaert, 1785, South Africa, recently extinct
E. (Quagga) burchelli Gray 1824, East, South, and West Africa, extant
E. (Quagga) mauritanicus Pomel 1897, North Africa, Middle Pleistocene
E. (Quagga) capensis Broom 1909, South Africa, Middle Pleistocene
E. (Quagga) oldowayensis Hopwood, 1937, East Africa, Lower Pleistocene.
4.4.1. Extant Quaggas
Description
Convex forehead, subequal naso-incisival notch and cheek lengths, short choanae, and small auditory meatus (Figure 45). Premaxilla more developed in E. (Quagga) quagga than in E. (Quagga) burchelli.
The upper cheek teeth have moderately long asymmetric protocones and moderate plications; plis caballin are usually present, at least on premolars; post-protoconal valleys are shallow. On lower cheek teeth, the vestibular valleys (ectoflexids) are shallow on premolars and usually, but not always, deep on molars; metaconids are rounded, metastylids may be rounded or pointed; lingual valleys (linguaflexids) are always ‘V’-shaped, unlike the ‘U’-shaped pattern of caballines (Figure 46).
The infundibula in the lower incisors are variably developed, often lacking in northern subspecies of extant Plain’s zebras [31].
Robust metapodials.
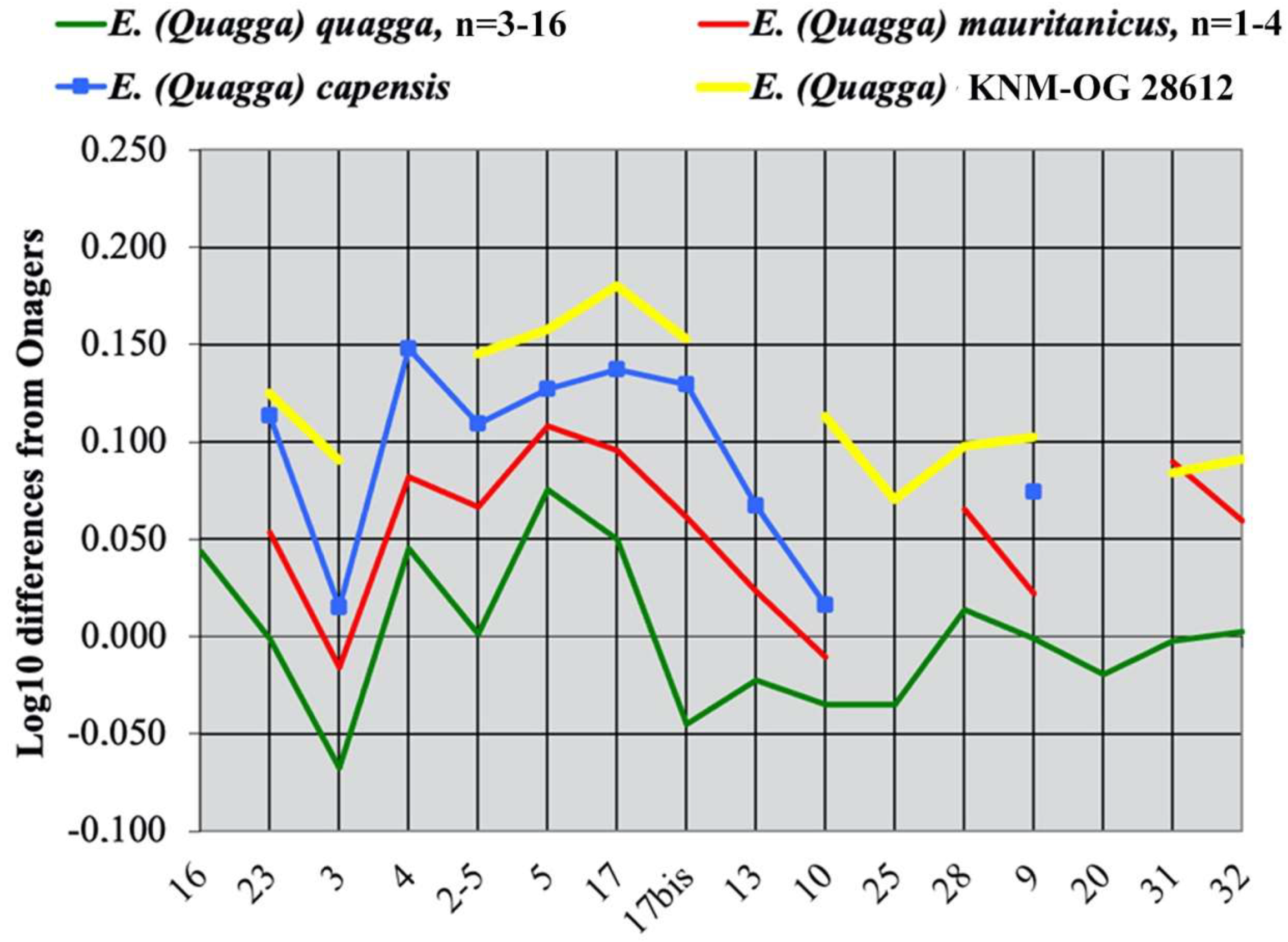
Biomolecular studies [22,30] have established the conspecificity of the recently extinct quagga and the extant Plain’s zebras (granti, boehmi, burchelli, chapmanni). Orlando et al. [22] have also found that ‘the extinct DNA sequences of the Cape zebra (E. capensis Broom, 1909) clustered among the two southern subspecies E. (Quagga) quagga, E. (Quagga) burchelli)’ confirming the observations of Eisenmann [75] on cranial proportions.
4.4.2. E. (Quagga) mauritanicus Pomel, 1897
4.4.3. E. (Quagga) capensis Broom, 1909
4.4.4. E. (Quagga) oldowayensis Hopwood, 1937
A very large cranium, lacking its posterior part but otherwise very well preserved, was found in member 1 of the Olorgesailie Formation, Kenya inside the Acheulean levels circa 1 Ma old [77]. This specimen (Figure 50), KNM-OG (Kenya National Museums, OG 22833: Olorgesailie, Nairobi, Kenya), was referred to E. oldowayensis [12]. The holotype of this species was lost during the war; the paratype—a lower incisive region—is inadequate, as pointed out by Bernor et al. [12]. The neotype chosen by Churcher and Hooijer [78] is a mandible whose precise provenance is unfortunately not known. The banal stenonine morphology of its teeth does not allow us to infer its belonging to the rather slender Equus of Bed I–lower Bed II or to the robust Equus of the Bed II species. Thus, E. oldowayensis remains badly defined.
4.5. E. (Hippotigris) Smith, 1841 (Mountain Zebras)
Very wide supra-occipital crest, very long and large external auditory meatus, long choanae, flat forehead, rectilinear naso-frontal suture, very narrow infraorbital bar: the suture between the premaxillar and nasal bones is perpendicular to the naso-maxillary suture; on the ascending ramus of the mandible, the surface for the insertion of the masseter has a peculiar rectangular shape ([79], Figures 2–5).
Short protocones, especially on premolars; plis caballin weak or absent. Banal pattern of lower cheek teeth; plis caballinid weak or absent (Figure 53).
Infundibula present on most lower incisors [31]. Very short metapodials; very narrow third phalanges with flat plantar surfaces.
E. (Hippotigris) zebra zebra are smaller than E. (Hippotigris) zebra hartmannae and have shorter post-vomerine lengths.
A fossil cranium found near Norval’s Pont on the Orange River was described by Lundholm [80] as a new subspecies of E. (Hippotigris): E. zebra greatheadi. Unfortunately, its age is unknown.
4.6. E. (Equus) Linnaeus, 1758
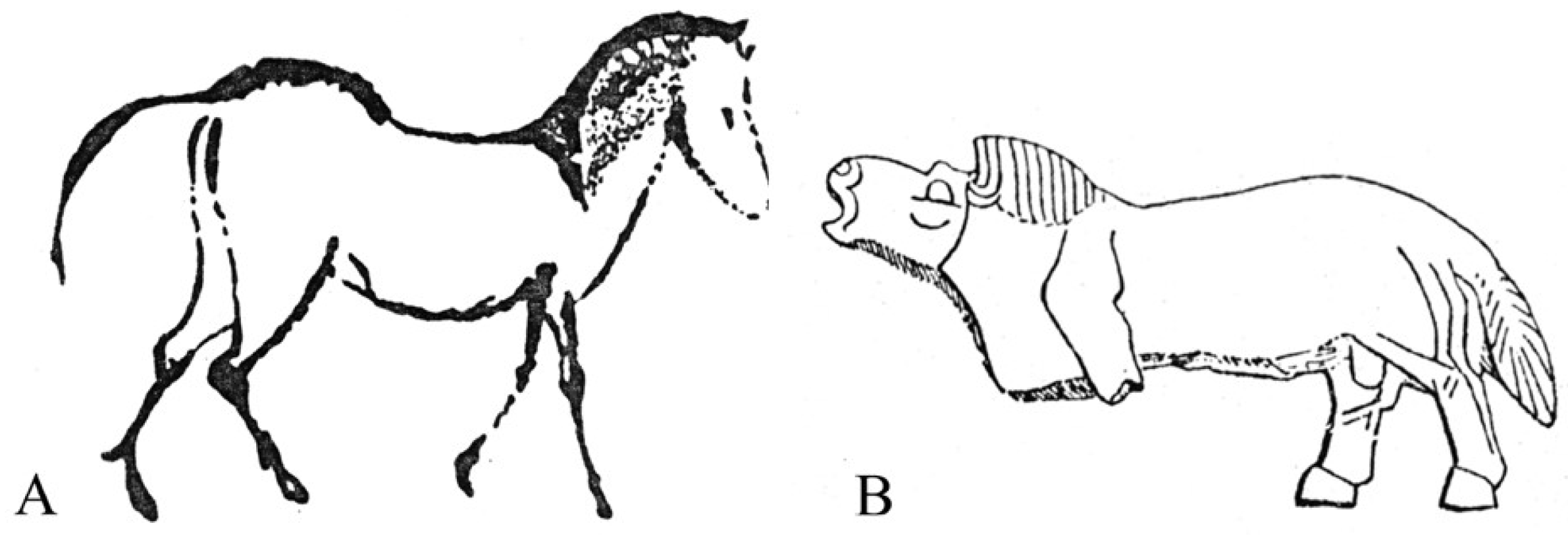
The fossils belonging to the subgenus Equus had the widest range of all Equus. In the Old World, during the late Pleistocene, caballine horses may be found from 75° N [81] to 35° N [57], and from 130° E [82] to 10° W [83]. Obviously, their environments were extremely different, leading to different adaptations evidenced by cranial, dental, and limb bone morphologies [84,85]. In particular, size follows Bergmann’s law, muzzles tend to be short in cold biotopes (Allen’s law), cursoriality is best developed in open landscapes, and wide third phalanges are an adaption to heavy grounds. Ecomorphomogical patterns were recently discussed by Boulbes and van Asperen [37] and the morphological variability in the subgenus may be schematically represented by the juxtaposition of two prehistoric artworks (Figure 54).
It should be noted that the decrease in the size during the Pleistocene [37], or rather its fluctuating decrease [87], is not absolute. One of the largest skeletons of a fossil horse was found at San Sidero, Puglie, Italy, and it is of Tardiglacial age [88]; the distal supra-articular and articular breadths of the MT MM 1404-5 from San Sidero 3 (dated at 10.280 BP) are, respectively, 58.5 and 60 mm.
Size, cranial, dental, and limb bone features may be combined in different ways, characterizing many different forms. There are, however, more or less constant characteristics that may be defined as ‘caballine’.
Caballine characteristics
- Cranium characteristics: Franck’s and Palatal indices [89].
In Caballines, unlike in Asses, the distance from the Staphylion (the posterior border of the vomer) to the Basion (the anterior border of the foramen magnum) is longer than the distance from the Staphylion (the posterior border of the palate) to the Hormion (S2); the Palatal length sensu stricto is always longer than the distance between the Staphylion and the Hormion (S2). Unfortunately, a great number of Equus are wrongly placed among Caballines.
The external auditive meatus is small.
- 2.
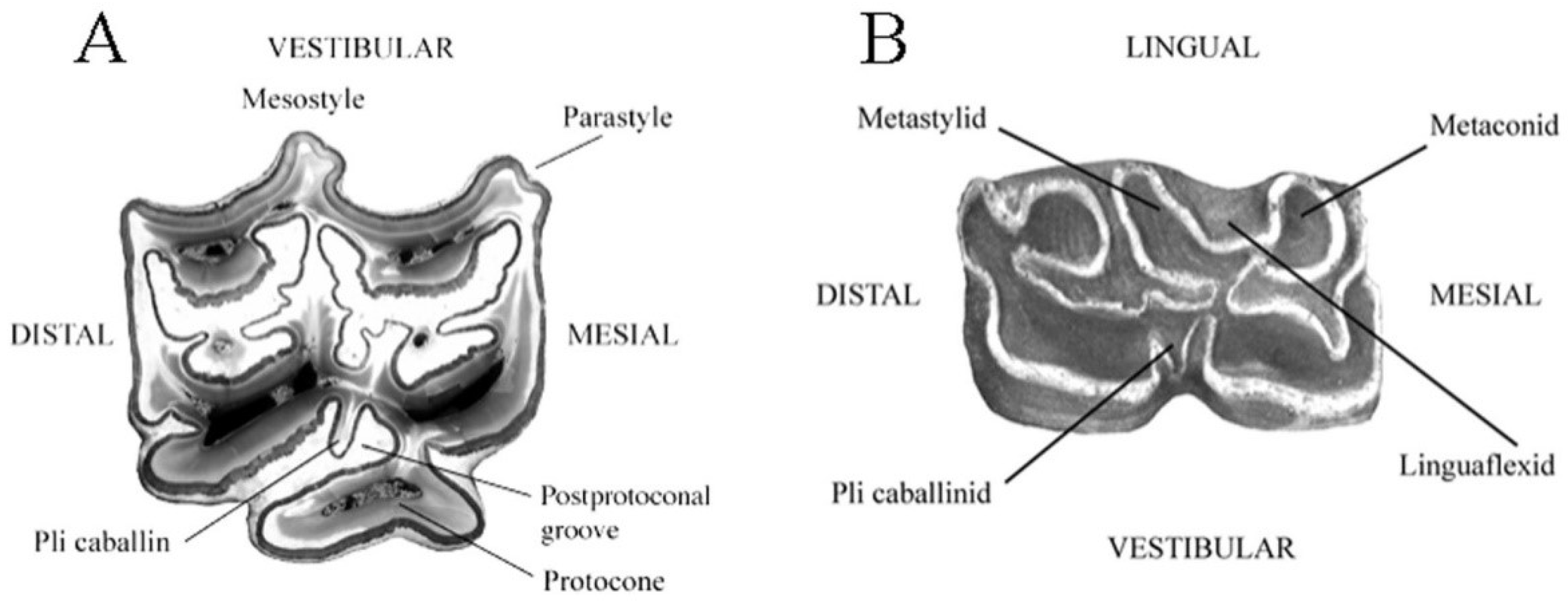
- Cheek teeth (Figure 55)
In typical upper cheek teeth:
- -
- The parastyle and mesostyle are grooved at least on premolars.
- -
- The vestibular enamel in the front and back of the mesostyle is concave.
- -
- The pli caballin is present at least on premolars.
- -
- The post-protoconal groove is not very deep.
- -
- The protocone is long and asymmetric, being less developed medially than distally.
In typical lower cheek teeth:
- -
- The linguaflexid is angular.
- -
- The double knot (metaconid + linguaflexid + metastylid) is asymmetric.
- -
- The metastylid is pointed.
- -
- The pli caballinid is present at least on premolars.
The ectoflexid (vestibular groove) of the molars may be either shallow or deep. The lower molars are often more typical than the premolars.
- 3.
- Metapodials
Usually, the distal articular breadths are larger than the distal supra-articular ones.
Groves and Grubb [24] recognize only two species more or less recently surviving: E. przewalskii and E. ferus. Boulbes and Asperen [37] refer to all fossil forms as subspecies of E. ferus. There were many fossil subspecies of horses; which of them are ‘good’, ‘bad’, or ‘ugly’ (according to the definitions of Groves and Grubb) remains open to discussion. Only the most representative of some epochs and groups will be addressed here.
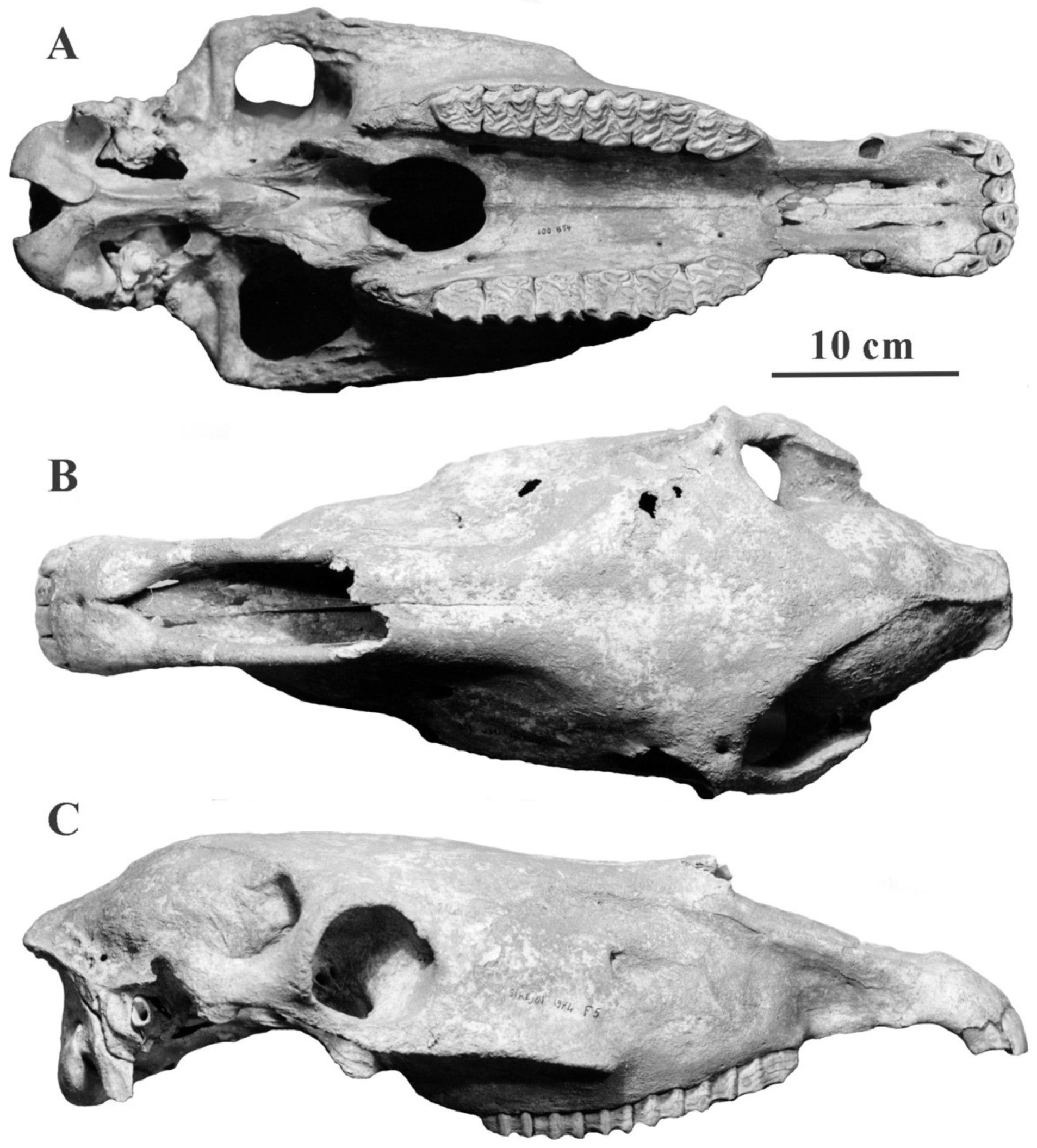
4.6.1. E. (Equus) ferus Boddaert, 1785
The available osteological remains of the extinct Tarpan are limited to one complete skeleton: St Petersburg: ZIN 521 (Figure 56) and one isolated cranium (Moscow: MGU 94535), both from Ukraine. The latter is in a very poor state of preservation and belonged to a very old male. Both individuals were castrated. Other specimens labeled ‘tarpans’ are the result of tentative genetic reconstructions by crossing domestic horses and selecting which individuals appear in their exterior morphology more like the available descriptions of extinct Tarpans.
Description
Probably because of the castration, the cranium lateral view reminds of juvenile specimens (Figure 56C). The Basilar length is not very large: 470 mm. The muzzle is short and wide. The choanae are short.
On the upper cheek teeth, the protocones are long on the molars; the plis caballin are well developed on premolars; the enamel is moderately plicated (Figure 57A).
On the lower cheek teeth, the double knot is caballine; the plis caballinid poorly developed; the ectoflexids are deep on the molars (Figure 57B).
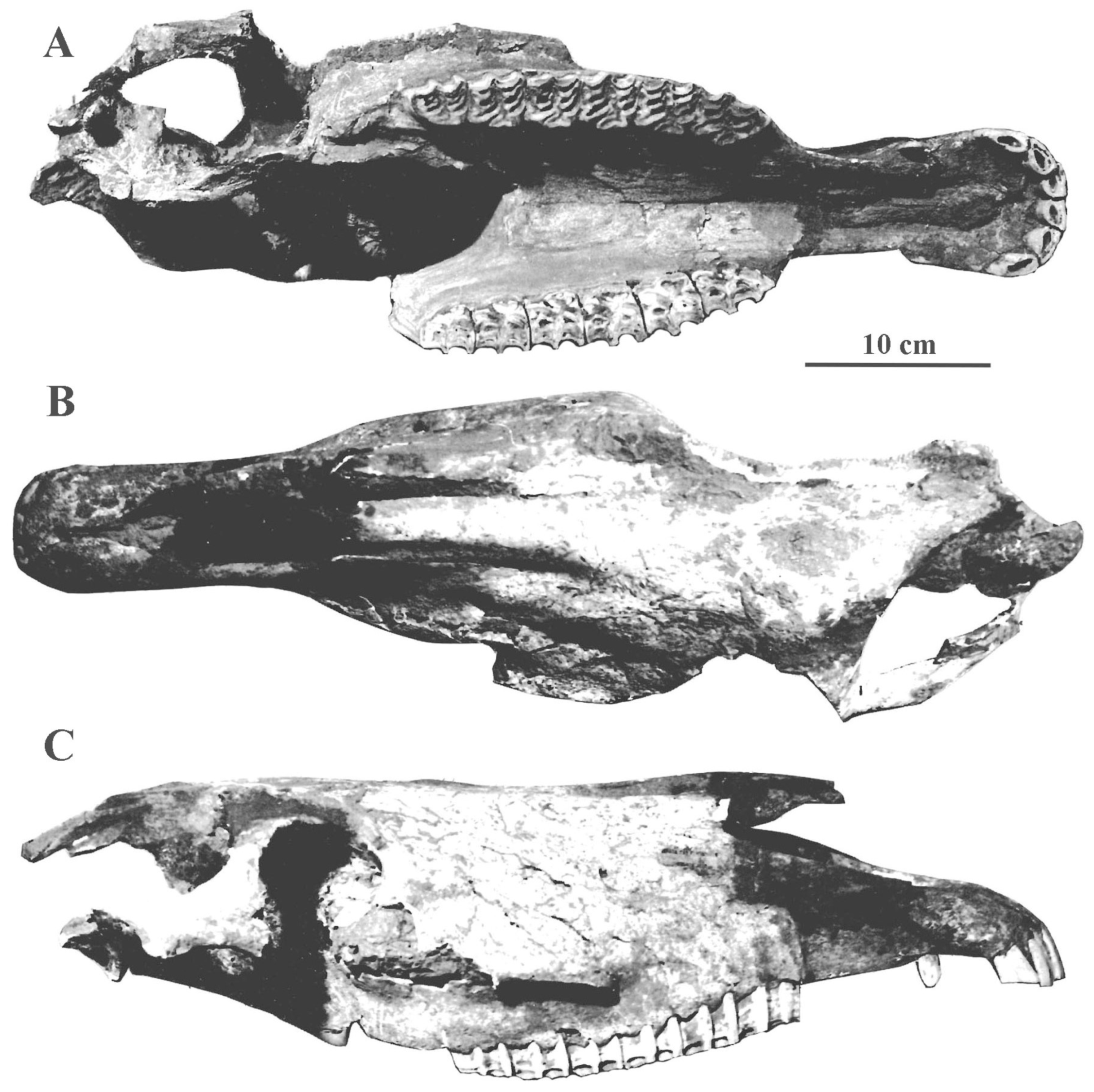
4.6.2. E. (Equus) przewalskii Polyakoff, 1881
The all but extinct E. przewalskii was confined to Mongolia in a semi-desertic envi-ronment and harsh climate.
Description
The cranium has a short and broad muzzle, a much longer distance from the Staphylion (the posterior border of the palate) to the Hormion, and larger facial and cranial heights than the Tarpan (Figure 58).
On the upper cheek teeth, the protocones are long, and the enamel is plicated; the plis caballin are better developed on the premolars than on the molars (Figure 59A). On the lower cheek teeth (Figure 59B), the double knot is caballine; the plis caballinid are poorly developed or absent; the ectoflexids may occasionally be shallow on the molars.
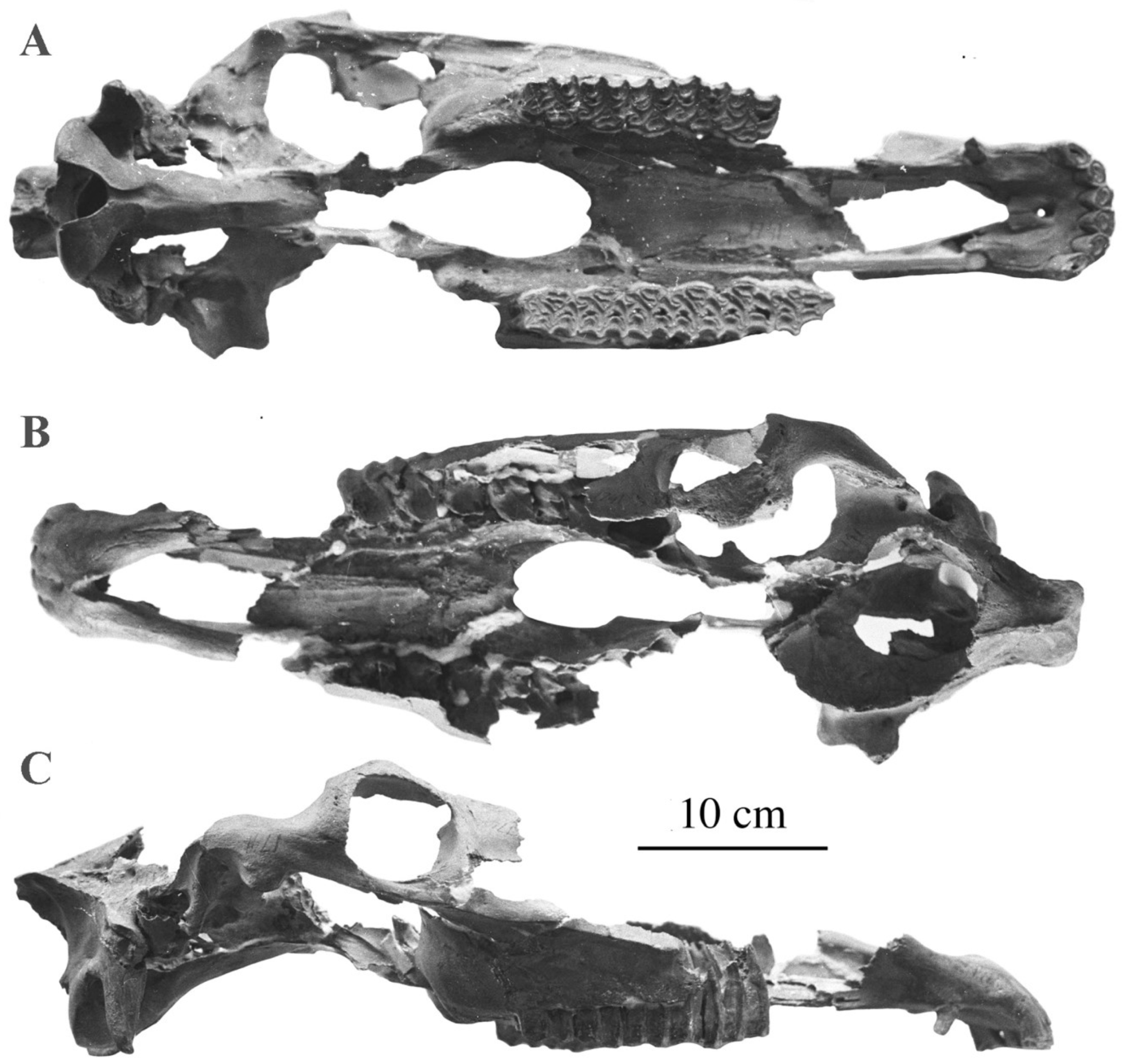
4.6.3. E. (Equus) ferus cf. scotti Gidley, 1900
The well-preserved cranium SI 160-455 (ex Bet 55) was found in magneto-positive deposits referred to as the Lower Brunhes at Ulakhan Sular on the banks of the river Adycha, North Eastern Siberia (Sher, personal communication).
Description
The cranium belonged to an adult male (Figure 60). It is very large (Basilar length: 585 mm); the choanae are small, and the muzzle wide. By all characteristics except its flat forehead, this cranium resembles E. scotti crania from Rock Creek, Texas, believed to be ca. 0.7 Ma old [90].
The upper cheek teeth have a plicated enamel, long protocones, and small plis caballin (Figure 61). There are no lower cheek teeth associated.
4.6.4. E. (Equus) ferus chosaricus Gromova, 1949
Tunguz peninsula on the Volga, Riss [91].
Description
The cranium MGRI 113-165 belonged to an adult female (Figure 62). It is smaller than E. (Equus) ferus cf. scotti (Basilar length = 511 mm) and has a short and wide muzzle.
The upper cheek teeth have a very plicated enamel, long plis caballin, and may have very long protocones (Figure 63).
The limb bones are robust. The MC length = 244 mm, and the diaphysis breadth = 42 mm.
The short muzzle (also present in E. (Equus) przewalskii) and the robustness, suggesting a cold and humid environment, are present also in two other younger European subspecies, E. (Equus) ferus germanicus and its smaller successor E. (Equus) ferus gallicus. Both (unlike E. przewalskii) are heavily built, not cursorial, and have very wide third phalanges ([85], Figure 1).
4.6.5. E. (Equus) ferus germanicus Nehring, 1884
According to Forsten and Ziegler, who compared E. (Equus) ferus germanicus to many Pleistocene horses [92], its age is believed to be Early to mid-Würmian.
From Remagen, Germany, Nehring [93] described the nearly-complete skeleton of a mare about 10 years old.
Unfortunately, the cranium (Figure 64) is nowadays too damaged to be measured, but some dimensions were given by Nehring ([93], pp. 100–114): the Basilar length = 528 mm, and the breadth of the supra-occipital crest = 58 mm. The specimen has a very short and very wide muzzle.
The upper cheek teeth (Figure 65A) are typically caballine. The protocones are less long and the enamel less plicated than in E. (Equus) ferus chosaricus, corresponding probably to less humid conditions.
The MC MBMa 16697 is robust (Length = 233 mm, the diaphysis breadth = 39 mm, robustness index = 167.3, but less so than in E. (Equus) ferus chosaricus (172.1) and in the Mousterian of Tournal, France [37], where one specimen has a length of 223.1 mm and a diaphysis breadth of 43.7 mm, giving it a robustness index of 195.9 (Boulbes, personal communication). The MT dimensions ([93], p. 137) are: maximal length = 285 mm, proximal breadth = 60 mm, breadth at the middle of diaphysis = 39 mm, and distal breadth = 57. The posterior first phalanx is 87 mm long and the posterior third phalanx is 85 mm wide ([93], p. 140, 141).
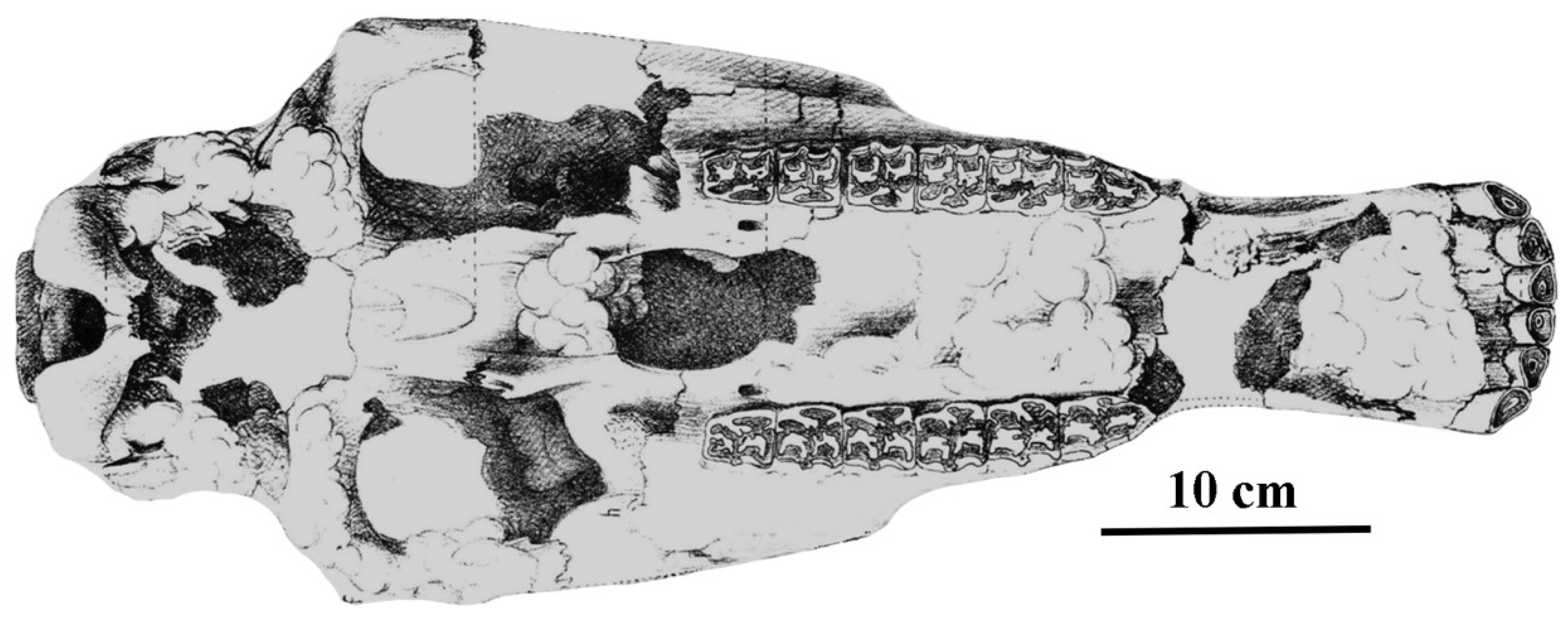
The horse from Siréjol, France, [94] dated to 27.100–31.500 Ka. may be referred to this subspecies.
Description
The cranium MNHL 100-854 (Figure 66) is perfectly preserved. It belonged to a middle-aged male. The Basilar length is 490 mm. The choanae are short, and the muzzle is not as short nor as wide as in the Remagen cranium.
The upper cheek teeth (Figure 65B) are similar to those from Remagen (Figure 65A). The lower cheek teeth are caballine.
The metacarpals are slightly smaller than the specimen from Remagen and larger than those of E. (Equus) ferus gallicus (respectively: maximal length = 229.1, 233, 220.5 mm; breadth at the middle of the diaphysis = 37.8, 39, 37.7 mm). The metatarsals are also slightly smaller than the specimen from Remagen and larger than those of E. (Equus) ferus gallicus (respectively: maximal length = 270.1, 285, 263 mm; breadth at the middle of the diaphysis = 39, 39, 38 mm).
4.6.6. E. (Equus) antunesi Cardoso and Eisenmann, 1989
Unlike E. (Equus) ferus chosaricus and E. (Equus) ferus germanicus this Late Pleistocene South-European species [95] was slender and rather cursorial.
Description
The cranium belonged to an adult male (Figure 67). It is large and narrow (Basilar length: ca. 520 mm; Frontal breadth: ca. 250 mm). The muzzle is longer than in E. (Equus) ferus and E. (Equus) przewalskii; it is broad at the posterior end of the incisors (66 mm) but constricted in the middle (43.2 mm). It resembles some crania from Valdichiana, Italy, to which it is probably related, and the extant Arab horses ([95], Figure 3).
The protocones are long.
The upper and lower cheek teeth are typically caballine. The limb bones are slender; the MCs are deep in the diaphysis.
4.7. Sussemionus Eisenmann, 2010 [96]
All species are extinct.
Type species: Equus (Sussemionus) coliemensis, [81], Kolyma, NE Siberia, Russia, Late Olyorian (600–450 Ka).
Type specimen: cranium IA 1741. The type cranium will be described below, but since it is the only one cranium known at the moment, the diagnosis of the subgenus must be limited to dental characteristics. Fortunately, they are quite distinctive.
Diagnosis.
Upper cheek teeth with extraordinarily developed and shaped plis caballin; very plicated fosettes. Lower cheek teeth with very developed and sometimes isolated stylids; premolar double knots with elongated, sometimes bilobated metaconids; lingual valleys shallow, nearly absent at times; frequent deep vestibular valleys on molars and sometimes in premolars too. Robust limb bones. Altogether, the Sussemiones seem adapted to humid climates.
Differential diagnosis
The enamel development, the frequent occurrence of strange plis caballin on the upper cheek teeth, as well as the occurrence of plis protostylid in the p2 and of the isolated ectostylids are characteristic of the subgenus Sussemionus.
Detailed descriptions, photos, and discussions are available in [28] and at vera-eisenmann.com (accessed on 26 september 2008).
In the Old World, the Sussemionus species ranged from northeastern Siberia to Germany and Ethiopiaa from just before Jaramillo to ca. 0.6 Ma. They are often associated with smaller and slenderer species. At first view, it appears surprising since the first seem to be adapted to humid conditions while the opposite is probable for the latter. A recent study of body mass and diet ([97], p. 11) offers an explanation: if sympatric ‘the smaller species had a more grass-dominated mesowear signal... whereas the large species… had a mixed-feeding one, even including a significant component of browse in the diet’.
4.7.1. E. (Sussemionus) coliemensis, Lazarev, 1980
The only Sussemione cranium known (IA 1741, Figure 68) was collected in Kolyma, Russia, near the Chukochya river (155° E, 70° S) in Late Olyorian deposits (600–450 Ka) [81].
Description
The cranium is the size of a Dolichohippus; the basi-cranial proportions are those of an Equus; the muzzle is long and narrow; the frontal breadth is large; the supra-occipital crest (lambdoid crest) is very narrow. Altogether, the proportions are close to African Wild Asses except for the narrow supra-occipital crest (Figure 69).
The upper P2–M3 series is 180 mm long. The teeth have a very plicated enamel and moderately long protocones. On the premolars, the plis caballin are wide at their base (Figure 70A). The lower cheek teeth (Figure 70B) illustrated by Lazarev may or may not belong to the same species. On the m3, the enamel is plicated, the hypostylid is very developed, the ectoflexid is shallow, and the metaconid is elongated. On the rather worn p2–p4, the lingual valleys are shallow, the metaconids are elongated, and the enamel is simple. The vestibular valleys (ectoflexids) are shallow.
The cranium was not associated with limb bones of corresponding size.
4.7.2. E. (Sussemionus) verae Sher, 1971
The type of E. verae [98] (PIN 835-123) was also found in Kolyma, Russia, at Chukochya Locality 21—the base of the type section of the Olyor Formation. The Formation extends between just under Jaramillo and around 0.6 Ma. [99]. This p2–m3 series is believed to be as old as or even older than Jaramillo, thus being several hundreds of Ka older than the cranium of E. coliemensis.
Description
The series is 208 mm long; the cheek teeth have a very plicated enamel, a pli protostylid on the p2, and deep ectoflexids on the molars (Figure 71A). They differ from E. coliemensis by a larger size, rounded metaconids, deep lingual valleys, more plicated enamel, and much deeper ectoflexids on the molars.
The material from Locality 21 is not well preserved nor homogeneous: teeth size as well as metapodial size and morphology are variable. There is, however, a well-preserved upper premolar (Figure 71B) that, although rather small to belong to the type, shows interesting characteristics: extremely plicated enamel and a pli caballin wide at its base.
The MC III PIN 3100-801 is supposed to be of the same age; it resembles, however, very much the specimen found in the younger deposits at Locality 37.
Locality 37 is dated to the beginning of Brunhes, thus being contemporary with E. coliemensis’ age. It is one of the most interesting localities because of the associated remains of a large Equus (PIN 3100-333): upper and lower cheek teeth and several limb bones.
The upper cheek teeth resemble the premolar from Locality 21 (Figure 72A). The enamel is very plicated (at times multiple plis caballin) and has deep post-protoconal valleys. There are only two lower cheek teeth. The m1 or m2 has a stenonine double knot and an extremely deep ectoflexid. On the m3 (at an early stage of wear), the ectoflexid is very shallow (Figure 72B).
The associated MC and MT III are large, robust, and flat in the diaphysis. The MC dimensions are: length = 261 mm; breadth proximal = 66.5 mm, at the mid-diaphysis = 44 mm, distal supra-articular = 62.2, articular = 64.2; depth proximal = 42.5, at the mid-diaphysis = 32 mm, distal at the keel = 43.4.
The MT dimensions are: length = 312 mm; breadth proximal = 57.5 mm, at the mid-diaphysis = 41 mm, distal supra-articular = 62.5, articular = 60.5; depth proximal = 48, at the mid-diaphysis = 36.5 mm, distal at the keel = 43. The third anterior phalanx is very wide: 111 mm. If the classical interpretations of plicated enamel and wide third phalanges are correct, this Equid lived in very humid conditions and on heavy ground.
Concluding remarks.
Inside the whole sample collected by Sher at Chukochya, there is no upper cheek tooth quite similar to those of IA 1741, although many have also the plicated enamel and the bizarre, wide-at-the-base plis caballin on P3 and P4. In addition, the IA 1741 teeth are smaller than most of the other Chukochya teeth. I think that E. verae and the younger E. coliemensis may be considered different species.
4.7.3. E. (Sussemionus) suessenbornensis Wüst, 1901
The Süssenborn quarry is 23 m high, and deposits may have been accumulating for as long as 100 Ka. The horizons with fossils are covered by a moraine of the Elsterian (Mindelian) glacial so the fossils are older than 450 Ka. The fauna of Süssenborn is even older than ca. 600 Ka because it contains Mimomys savini, the phylogenetic precursor of Arvicola, whose oldest Central European representatives have been recorded in Mauer ca. 600 Ka old [100]. On the other hand, the Süssenborn fauna is younger than that of Voigtstedt/Lehmzone, whose age is probably ca. 700 Ka [101].
Fossils were collected a long time ago without stratigraphic information. The collection is mainly composed of teeth and is very heterogeneous. The size and morphology of the taxa have probably changed during the time of accumulation, and there are several ‘morphs’ that may or may not have been true species.
Description
E. (Sussemionus) suessenbornensis is a large Equid. It is mainly characterized by the hypertrophy, even exuberancy of its cheek teeth enamel. On the upper, plis caballin are constant but vary in size and shape; the postprotoconal groove is shallow; the parastyles are wide but not grooved on premolars and are less wide on molars; the size and shape of the protocones are very variable.
The lectotype chosen by Musil ([102], Pl. XXXVII, Figure 1) is a rather worn upper cheek P2–M3 series IQW 1964/1177 (S 514); the enamel is plicated but there are nearly no plis caballin; the protocones are very variable in size (Figure 73A). The upper moderately worn series figured by Wüst ([103], Plate VI, Figure 9) is unfortunately lost. It is larger (Figure 73B) and illustrates what I believe are more typical characters: extremely plicated enamel, deep postprotoconal valleys, complicated plis caballin, and bilobated protocones. Figure 73C–E further illustrates the peculiar morphologies of E. sussenbornensis upper cheek teeth.
The p2–m3 S 9279 series is the size of Wüst’s upper series; it has deep ectoflexids on all cheek teeth (Figure 74B). On the m3 Ha E 23 (Figure 74C), there is an ectostylid at the base of the crown. The lower p2–m3 was figured by Wüst ([103], Plate VII, Figure 1)—this article, Figure 74D, also seems lost. The paralectotype [103] IQW 1964/1302 (S 9279) is smaller (Figure 74A).
In the Süssenborn collection, there are a few teeth, smaller than E. suessenbornensis but sharing some enamel patterns with it, which may be younger, ‘evolved’ forms (Figure 75); I refer to them as E. cf. suessenbornensis.
More detailed information can be found in the Supplementary Materials (Figures S3 and S4).
4.7.4. E. (Sussemionus) aff. sussenbornensis Vekua, 1962
The Akhalkalaki (Georgia) fauna was described in English in 1986 [104]. It is believed to be 0.8–0.9 Ka old. There are two Equus: one rather similar to E. suessenbornensis and the other—a new species, E. hipparionoides—much smaller, with many strange features that will be discussed later.
Description
The upper cheek teeth (Figure 76A) are the size of E. verae and resemble them as well as those of E. suessenbornensis: deep post-protoconal valleys and long plis caballin on the premolars. The enamel is, however, less plicated. The lower cheek teeth also have a plicated enamel, unless they are too worn, but never as much as E. verae and E. suessenbornensis. An m3 and an m2 (TB Akha 4), both very worn, have an isolated ectostylid (Figure 76B,C). One p2 without a number (Figure 76D) has a pli protostylid.
4.7.5. Concluding Remarks
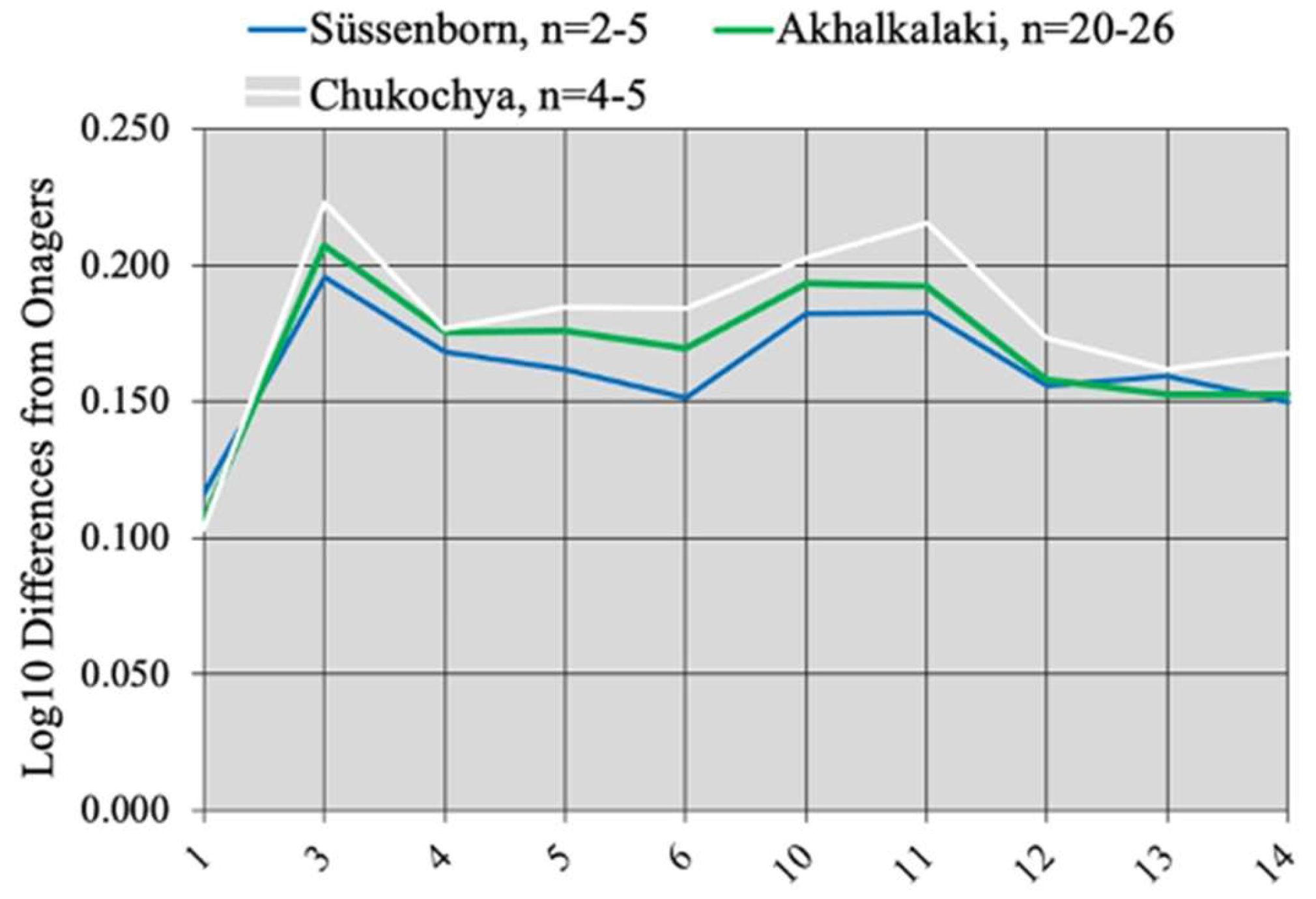
The metapodials of the three species are about the same size; the MC from Süssenborn and Akhalkalaki differ from those of E. verae by a smaller distal articular width (Figure 77).
The relative lengths of MCs, MTs, and first anterior and posterior phalanges are similar, but the width of the third anterior phalanx is notably smaller (average 93.3 mm, and 97 mm maximum at Akhalkalaki instead of 111 mm in E. verae). If the classical interpretations of plicated enamel and the width of third phalanges are correct, the Equid of Akhalkalaki lived in less humid conditions than those of Süssenborn and Chukochya.
4.8. Pseudohydruntines and E. altidens
There were many Equid slender taxa with small protocones on the upper cheek teeth and deep ectoflexids on the lower molars. Their similarities may result from genetic proximity or from parallel evolution. One of them is E. hydruntinus, which is now known to belong to Hemiones [32,33]; I refer to others as ‘Pseudohydruntines’.
Although E. hydruntinus and Pseudohydruntines share slenderness and small size of the protocones they differ by several features as illustrated in Figure 78.
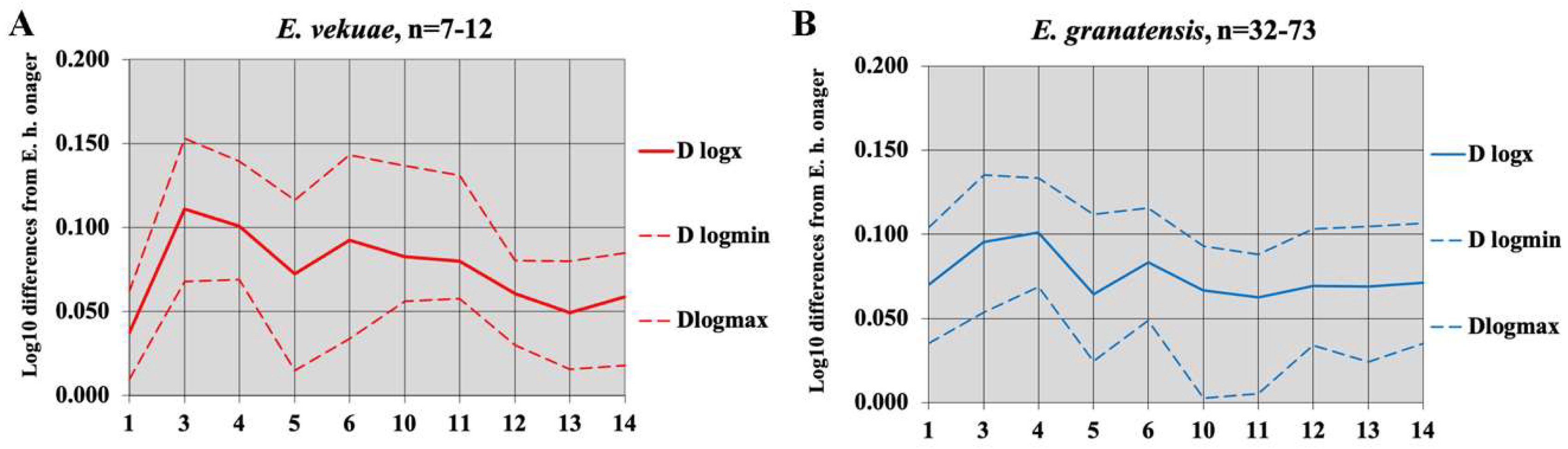
Pseudohydruntines are larger than Hydruntines, their protocones are relatively shorter, and the distal articular breadth and mid-diaphysis depth of their MTs are relatively larger. E. tabeti from Aïn Hanech (Algeria) plots with E. hydruntinus (very surprisingly since it is an African and much older species).
Because of particular dental morphologies, I have previously believed that several Pseudohydruntines discussed below belonged in the subgenus Sussemionus. I prefer now to keep them apart.
Pseudohydruntines are commonly referred to E. altidens, a referral I find unjustified, as will explain below.
4.8.1. The Problem with ‘Hippotigris’ altidens Reichenau, 1915
Fossils described as Equus altidens come from the Süssenborn quarry deposits that could cover 100 Ka between 700 and 600 Ka [100,101].
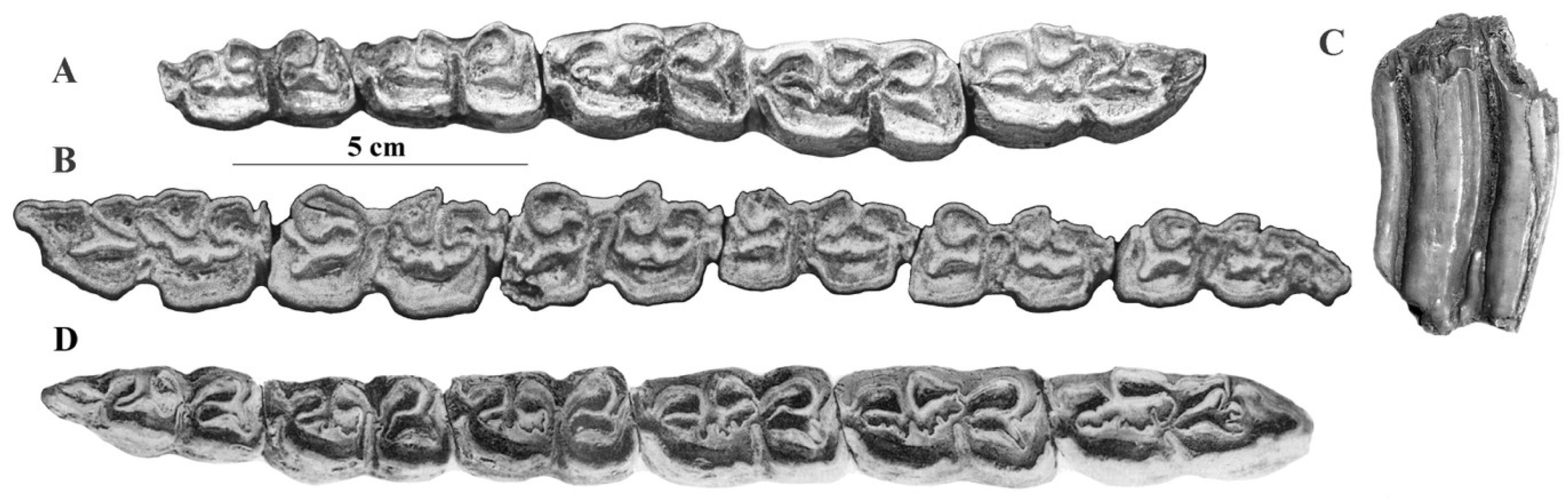
Description
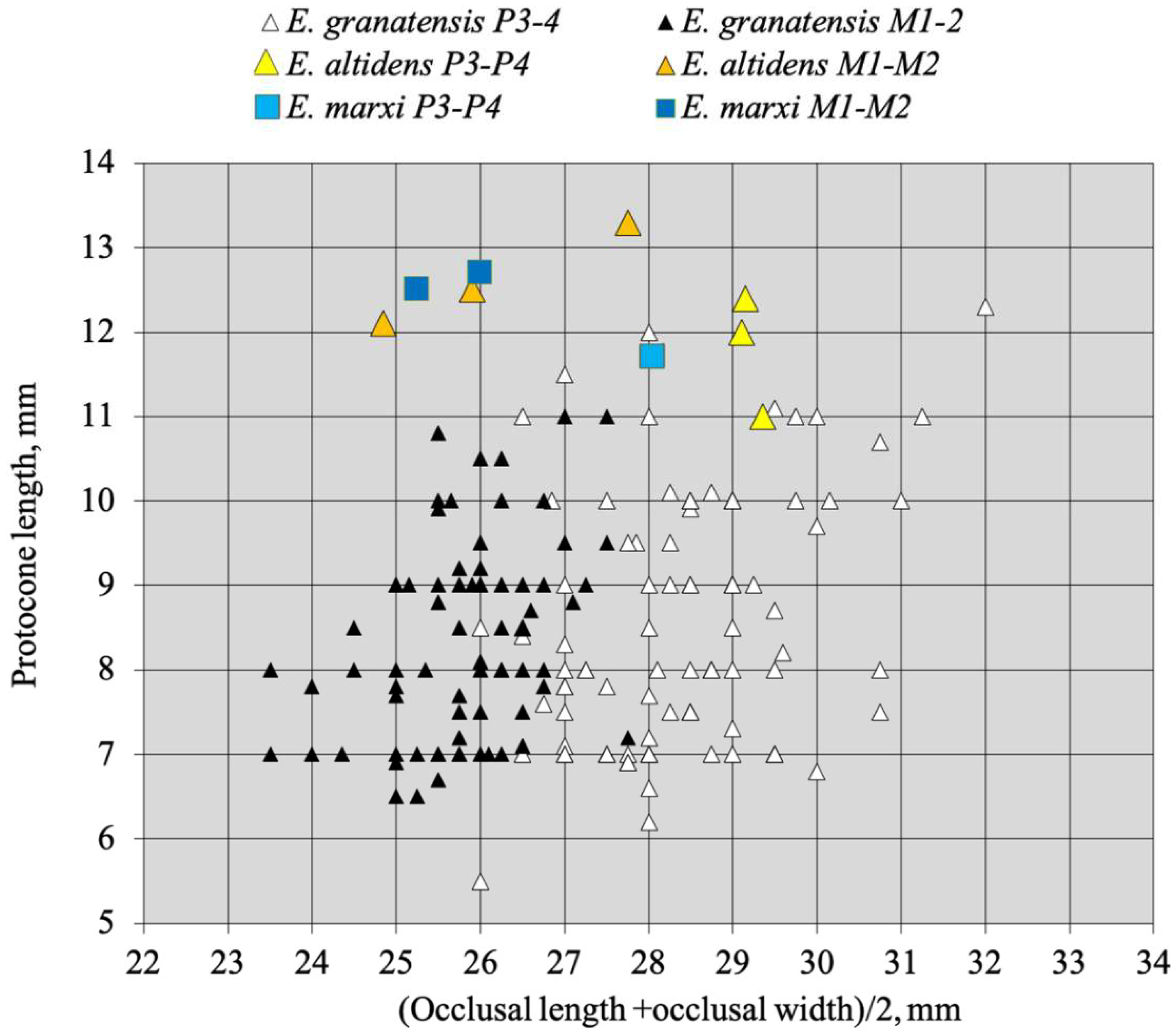
According to Reichenau’s text and illustrations [105], E. altidens is defined uniquely by the height of its lower cheek teeth and its stenonine double knot—placing it inside ‘Hippotigris’. All the upper cheek teeth referred to by Reichenau to Hippotigris altidens (Figure 79 and Figures S5–S14) have rather long protocones, so there is no reason to refer to E. altidens the fossils of Venta Micena and Pirro, which are somewhat older, have remarkably small protocones (Figure 80), and may even belong to Plesippus according to Alberdi and Palombo, ([106], p. 159).
On a biochronological basis, Pirro Nord is dated between 1.3 and 1.6 Ma [106]. According to Arvicolid evolution trends ([107], Table S1), its age is estimated at 1.2 to 1.5 Ma. The age of Venta Micena was estimated at 1.095 Ma, according to the racemization of amino acids [108] but an Early Pleistocene age based on paleomagnetic studies and ESR datations is now commonly accepted and the site is ‘usually considered as biochronologically contemporaneous with the archeological locality of Pirro Nord, Italy’ [109]. According to Arvicolid evolutionary trends ([107], Table S1), its age is estimated at 1.4 to 1.8 Ma.
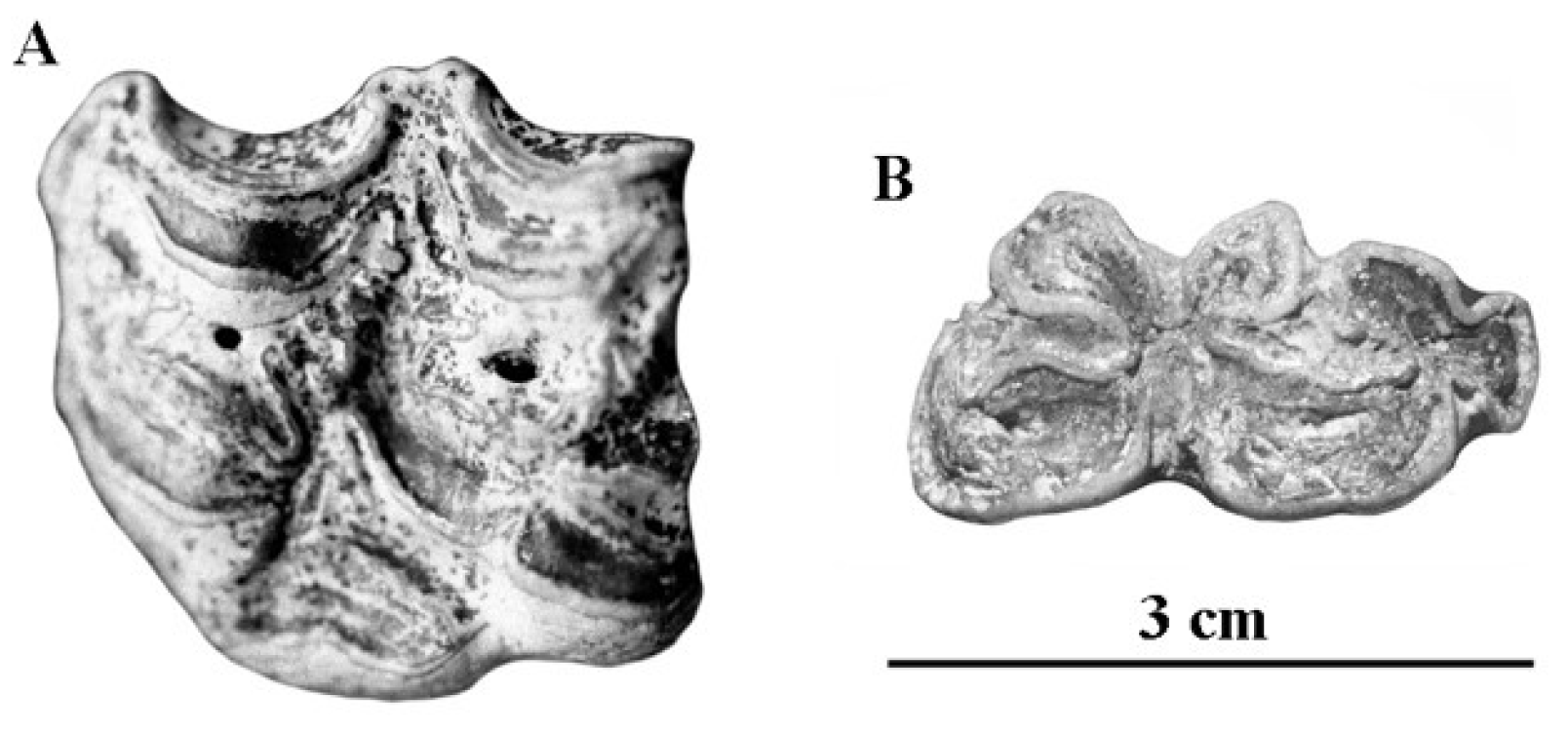
The lower cheek teeth referred by Reichenau to Hippotigris altidens have banal enamel patterns (Figure 77G–I). The series figured on Plate V-3 (Figure S6) should be referred to E. marxi, another of Reichenau’s species to be discussed later.
See the Supplementary Materials for a detailed discussion and a complete set of figures (Figures S5–S14) illustrating the syntype of Reichenau [105] for E. altidens.
There is no reason to suppose that the small fragmentary and badly preserved metapodials found at Süssenborn belong to E. altidens.
4.8.2. E. granatensis Alberdi and Bustos, 1985
No skull has been found, so a belonging to an Allohippus or a Plesippus was considered possible because the upper cheek teeth have small protocones. Its age is estimated at 1.4 to 1.8 Ma [107].
Description
The upper cheek teeth have indeed very small protocones (Figure 80 and Figure 81). The enamel is moderately plicated, the plis caballin is small, and the post-protoconal grooves are shallow.
On the lower cheek teeth, several characteristics distinguish E. granatensis from Allohippus stenonis and point to affinities with Sussemiones. Such are the occurrences of isolated stylids (Figure 82A), plis protostylid (Figure 82B,C), elongated metaconids (Figure 82B), and deep ectoflexids on the lower cheek teeth, in particular on some premolars (Figure 82D).
The metapodials are slender and deep in the diaphysis. Additional data and photographs may be found on vera-eisenmann.com (accessed on 21 June 2011).
The MCs are very slender and deep in the diaphysis and at the proximal end (Table S5).
4.8.3. E. aff. granatensis, Pirro, Italy
The small Equid from Pirro is one of the Early Middle Pleistocene ‘hydruntinus-like’ taxa, with very small protocones on the uppers (Figure 83A), stenonid patterns of lower cheek teeth (Figure 83B) and slender limb bones that have been referred to E. altidens. Important revision and comparisons [111] showed that it differs from E. granatensis by several features (to which I add the lack of stylids on the lower cheek teeth). In consequence, the Equus of Pirro was referred to E altdens altidens while the one of Venta Micena was referred to E. altdens granatensis. I have explained in the text above and in the Supplementary Materials why the use of the name E. altidens (a badly understood Middle Pleistocene species of Germany) is not appropriate and propose to refer to the Pirro Equus as E. aff. granatensis.
The MCs are deep in the diaphysis and at the proximal end but less slender than in E. granatensis (Table S5).
According to [106,107] Pirro Nord is ca. 1.2 to 1.6 Ma old. The fossils from Pirro Nord and Cava Pirro as well as from the probably slightly older Selvella were described first by De Giuli [112,113] and later by Alberdi and Palombo [38,111], who considered that they belonged to Plesippus. The discovery of a cranium would be necessary to confirm or infirm their opinion.
4.8.4. E. wuesti Musil, 2001
E. wuesti was described from Untermassfeld, Meiningen, Germany [114]. The fossiliferous sands of Untermassfeld are positioned around the onset of the Jaramillo polarity subzone, with an age of approximately 1.07 Ma BP [115]. More material was found and described recently [116] and referred to the subgenus Sussemionus because of the resemblances between its upper cheek teeth and those of E. (Sussemionus) granatensis from Venta Micena (Figure 81). However, neither the upper nor lower cheek teeth of E. wuesti have the usual Sussemione characteristics: no strange pli caballin and no elongated metaconid. I prefer now to consider this species as another of the Pseudohydruntines.
Description
Specimen IQW 2010/31 393 (Mei. 30555) is a subadult fragmentary maxillary from which the P3 and P4 were sectioned in order to better show the enamel pattern; the M1 is a little worn. The protocones are short, the postprotoconal grooves are not very deep, the plis caballin are normally developed, and the fossettes are moderately plicated (Figure 84B,C). Another P3, IQW 1999/26 560 (Mei. 26089), although very worn, shows the same aspect (Figure 84A).
IQW 1998/26 401 (Mei. 25930) is a fragmentary tooth row (p3–m2); the metastylid is pointed on the p3, rounded in the other teeth, and the vestibular groove is deep on the molars; the pattern is stenonine (Figure 85).
The metacarpals are slender, deep in the diaphysis and at the proximal end, but larger than in other Pseudohydruntines (Table S5). See [117] for a complete description.
Discussion
Among the Süsssenborn Equid fossils, there are many teeth that may be referred to E. cf. wuesti (Figure 86). The upper cheek teeth are small, with simple enamel, small plis caballin, and short protocones. The molars are very much alike to the premolars; size is variable, and thus the distinction between them is difficult. Reichenau referred one of them—probably IQW S 4609—to E. stenonis ([105], Plate VII-14).
The lower cheek teeth have symmetric double knots with thick and simple enamel, rounded metaconids, and nearly rounded metastylids. The ectoflexids are deep on the molars.
More photographs may be found in the Supplementary Figures S15 and S16.
It is probable that the fragmentary and badly preserved small metapodials referred previously to E. altidens actually belong to E. aff. wuesti. Several MC III and a lower molar from the lower levels of Mosbach probably belonged to the same form.
4.8.5. E. hipparionoides Vekua, 1962
Fossils of this species were found in association with much larger fossils similar to E. suessenbornensis at Akhalkalaki, Georgia [104].
Description
The protocones are exceedingly small; on the P4, the pli caballin has the shape of a club (Figure 87A). The p3 to m2 have plis protostylid; an isolated ectostylid is present on the m2 (Figure 87B).
The metacarpals are very slender and deep in the diaphysis. Additional data may be found on vera-eisenmann.com (accessed on 31 August 2009).
4.8.6. E. ovodovi Eisenmann and Vasiliev, 2011
The Equids from the Proskuriakova cave, SW Siberia are dated at 45–50 Ka BP. They have been first tentatively referred to E. cf. hydruntinus, and it was in that frame that samples have been subjected to biomolecular analyses in order to be compared to E. hydruntinus from other geographical areas. A subsequent osteological, more detailed study [117] of the fossils showed that they did not belong to E. hydruntinus, nor to any other extant or recently extinct species, thus confirming the results of the biomolecular analyses [22] which had found that this species belongs to a unique basal lineage of Old World Equids.
Description
The variable size and morphology are probably the result of the mixing of different levels.
The enamel of the upper cheek teeth is simple, the protocone lengths are average, the plis caballin are small, and the post-protoconal grooves are shallow. On the lower cheek teeth, the shape of the double knot and the pli protostylid on the p2 suggest an affinity with Sussemionus. So does the deep ectoflexid on the m2 from the Mousterian of Tsopi, Georgia, which may belong to the same species (Figure 88).
Metacarpals are deep in the diaphysis and at the proximal end but less slender than in other Pseudohydruntines (Table S5). A more complete description is given in [117].
Discussion
Unlike the lower cheek teeth of E. ovodovi, which have Sussemione characteristics, the upper ones do not. I think that it was wrong to refer them to Sussemionus, but the similarities with the latter should be kept in mind as well as the belonging of E. ovodovi to a unique basal lineage of Old World Equids.
4.8.7. Dmanisi
Preliminary Observations
In 1995, when I worked in Tbilisi, the used stratigraphy mentioned six Layers, the deepest—Layer VI—overlaying basalt. In Layer II were found fossils of Archidiskodon, Paleotragus, and of a large Equid. Layers IV and V were held sometimes difficult to distinguish one from the other.
Lordkipanidze et al. [118] described the stratigraphy of Block 2, above the 1.85 Ma Masavera Basalt. It is now established that there are two major stratigraphic Units: Stratum A, with a normal polarity, conformably overlies the basalt. Stratum B, with a reverse polarity, overlies Stratum A.
Ferring et al., [119] described the stratigraphy of a Unit 5 (M5) that yielded artifacts but no identifiable bones. Overlying Stratum A, Stratum B is ca. 1.77 Ma old. According to Ferring (personal communication), it is sometimes possible to correlate Layers of the old scheme with the new one inside Block 2, and the great majority of fossils from the old strata V and IV are from Stratum B1.
As the identifying numbers of the fossils are complicated, I asked for explanations. The following were given to me by Maia Bukhsianidze: “The format Dm5/154.3/4.A4.5 is the field number format, where Dm stands for Dmanisi, 5/154 stands for the square (this is a coordinate on the squares’ net of the entire excavation area and denoting the meters along the SN and EW axis), 3/4 for the sub square. In this case, this specimen was in both 3rd and 4th sub squares. A4—is the layer. 5—is the specimen number in the square”.
Even after setting aside the material from Stratum A, Layer II, and some specimens from Stratum B [MCs with exceptionally large distal and/or proximal epiphyses (Dm 9/151.3.Bj 61, Dm5/155.1.BC.10 (S20,21,22), exceptionally wide first phalanx Dm//153.Bj.61 ([120], SOM Table S1), upper cheek teeth with unusually large protocones LMC 353 (Table S6)] the fossils from Dmanisi are not homogeneous. It would be tempting to ascribe the differences in size and shape to sexual dimorphism but for the evidence that in extant Equus limb bones, the sexual dimorphism is almost absent, at least on metapodials and tali (https://vera-eisenmann.com/sexual-dimorphism-in-equus (accessed on 25 December 2011)). Possibly, it was more marked two million years ago.
Bernor et al. [120] recognized and described two Equids at Dmanisi: Equus stenonis, mostly from Stratum A, and E altidens, mostly from Stratum B. The well-preserved cranium described below was referred to the latter.
Equus vekuae n. sp.
Locality. Dmanisi, Georgia.
Age. Early Pleistocene.
Origin of the name. In honor of Abesalom Vekua who studied the site of Dmanisi [121].
Type specimen. Cranium Dm53/59.3.B1gl.192 housed in the Janashia Museum of Georgia.
Referred material. Fragmentary crania DMA 66 and 353. Maxillary Dm5/151.A2.39. Upper cheek teeth: P3 or P4, DMA 1186; M3, DMA 1089; P2–M3, DMA 353; P2–M3, DMA 66; P2–M3, Dm5/154.3/4.A4/5; P2–M3, Dm5/151.A2.39.
Mandible D4691. Lower cheek teeth: DMA 1127, P2–M1; DMA 115, P2–M3; DMA 117, P3–M3; DMA 4691; DMA 1086, P3 or P4. Metacarpals: DMA 59, 61, 62, 261, 529, 637, 717. Metatarsals: DMA 335, 709, 733, 1038, 1118, 1234, 1183, 1350, 1550, Dm733, no number. First phalanges: D1559, D1773, D1777, D2599, D5723, D5801, D5844, Dm53/59.3B1.14, Dm53/59. B1g1.190, Dm64/64.1.B12.198, Dm69/62.2B1p.143, Dm70/32.1A4.7, DmM215.A2.107, DmM215.A3.95, D5141, D5193, D523, D5366, D710, Dm8/151.A4.26, DMA 19, DMA 129, DMA 201, DMA 259, DMA 523, DMA 536, D4752. Tali: D1388, D2115, DM65/64.2B1Y61, Dm70/34.4.2A4.52, D1215, Q1850, D2249, D2649, D484, Dm6/153.1.A4.18, Dm6/153.1.AY.23, Dm64/64.1.B1z.152, Dm64/64.1BAY.23, Dm70/34.2.58.
Description and Discussion.
Crania
The cranium Dm53/59.3. B1gl.192 (Figure 89), of a middle-aged female, was found in Stratum B1. It is relatively narrow (Basilar length: 476 mm, frontal breadth: 198.4 mm) with a long and narrow muzzle (121.6 mm, 62 mm). The distances between the Staphylion and the Hormion and between the Hormion and the Basion are subequal (107.6 mm and 108 mm). The Cheek length is slightly shorter than the length of the naso-incisival notch (154 mm and 160 mm). The supra-occipital crest is long and narrow (47.4 mm).
The contemporary fragmentary cranium DMA 66 (Figure 90) was found inside Layer V. It looks like a slightly larger version of Dm53/59.3.B1gl.192.
Bernor et al. ([120], p. 10) referred both Dm53/59.3.B1gl.192 and GER 8 (Allohippus mygdoniensis from Gerakarou, Greece) to Equus altidens. The cranial proportions show that the crania differ too much to make this synonymy acceptable: Allohippus mygdoniensis (and Allohippus senezensis as well) has a longer palate (measurement 2-5) and vomerine length (measurement 3) and a somewhat deeper naso-incisival notch (measurement 31), all features characteristic of Allohippus (Figure 91).
After having looked at all the data at my disposition, the only (and slight) cranial resemblances (Figure 92) I have been able to find are with the IVCM 2673 cranium from Anza Borrego, California, ca. 2 Ma, referred to Plesippus [122] but actually belonging to Equus ([28], Figure 3).
The cranium from Anza Borrego shares with the specimen from Dmanisi the relative basi-cranial lengths (measurements 3 and 4) and to some extent the palate-to-muzzle length relation (2-5 and 5). However, it is much larger and narrower (measurement 13) and has a relatively longer cheek (measurements 31 and 32).
Upper cheek teeth
When complete, the upper series have a well-developed dP1. Most teeth have deep post-protoconal grooves and tiny plis caballin. DMA 243-244-245, 1037, and 1156 (Figure 93D,E) differ by their shallow post-protoconal grooves. The metric data are in ([120], SOM Table S1) and Table S6.
The protocones are very small, as small as in E. granatensis and much smaller than in E. altidens or E. marxi (Figure 94).
Lower cheek teeth
In the moderately worn teeth, the enamel is plicated. A pli protostylid is present on the p2 of DMA 1127 (Figure 95A). Plis caballinid are small or absent. Ectoflexids are deep in the molars and occasionally in premolars (Figure 95H). Unlike the upper cheek teeth, the lowers differ in size while they may have nearly identical enamel patterns like DMA 1127 and DMA 117: elongated metaconids, plicated enamel, and shallow ectoflexids on the p3 and p4 (Figure 95A,D). The metric data are in ([120], SOM Table S1) and Table S8.
Metacarpals
Although all originate, as much as it is possible to know, from Stratum B or its equivalent Layers IV-V, size and robustness are variable (S20,21,22). Some MCs (Figure 96A) tend to be more robust and deeper at the proximal end (measurement 6) but on the whole, they resemble Pseudohydruntines (Table S5). The data are in Table S9.
Metatarsals
Some MT are larger, but their proportions are similar to the smaller ones (Figure 97B, Figure S23). The data are in Table S10.
First Phalanges
Size and proportions vary also in the collection from Layers IV–V and Stratum B: DMA 201 is more robust than DMA 129, and DMA 647 is smaller (Figure 98). The data may be found in Table S11.
Differential Diagnosis
- -
- From all Allohippus crania, the cranium of E. vekuae differs by the longer distance between the Basion and the Hormion (measurement 4) and a shorter naso-incisival notch (measurement 32). From Allohippus mygdoniensis and Allohippus senezensis in particular (Figure 91), it differs also by shorter choanae (measurement 9). From Allohippus upper cheek teeth, those of E. vekuae differ by deeper post-protoconal grooves. From Allohippus lower cheek teeth, those of E. vekuae differ by the elongated metaconids and the occurrence of a pli protostylid on the p2.
- -
- From E. (suessemionus) granatensis—the best documented of all Equids usually referred to as ‘E. altidens’, E. vekuae differs by the lack of isolated stylids and MC robustness (Figure 99).
Data in Tables S7 and S9.
To sum up:
Although the size and proportions of the teeth and limb bones are very variable, on the whole, E. vekuae was of average size and moderately robust. As in other Pseudohydruntines, the MCs were deep in the diaphysis (Table S5). The cranium is narrow. The upper cheek teeth have short protocones, deep post-protoconal grooves, tiny plis caballin, and the dP1 are well developed. The lower cheek teeth have a plicated enamel, elongated metaconids, deep and pointed linguaflexids, and small plis caballinid; the ectoflexids are deep in the molars and occasionally in premolars; occasionally, a pli protostylid is present on the p2.
Two points must be stressed: the type cranium of E. vekuae does belong to a true Equus and it is the oldest representant of the genus in the Old World.
4.9. Incertae Sedis
The problem of Equus altidens was discussed above.
4.9.1. E. marxi Reichenau, 1915
This ‘nova et bona species’ described by Reichenau from Süssenborn was based on three upper and three lower associated cheek teeth fossilized in a different way from the rest and originating from the uppermost gravel layers of the quarry (Figure 100).
E. marxi is much better defined than E. altidens. The upper cheek teeth are similar to extant Hemiones by their enamel pattern: they have deep post-protoconal grooves and no plis caballin. The lower cheek teeth differ from Hemiones: the premolars have elongated and nearly bilobated metaconids like some extant Wild Asses (Figure 100E) and one premolar of E. (Sussemionus) granatensis (Figure 82B). Reichenau ([105], p. 25) noted that Hippotigris marxi is closely related to the smaller Hippotigris altidens by the characteristic shape of the inner valley (post-protoconal groove) of the upper cheek teeth. However, the shape of the double knot, in particular the elongated metaconids, is quite different from E. altidens.
Pictures of all the teeth mentioned and figured by Reichenau as belonging to E. marxi are in Figures S17–S19. Kahlke [123] gave as a Holotype the right P4–M2, IQW H.G. Süss. 1365 + 1366 + 1367 and as a Paratype the left p3–m1, IQW H.G. Süss. 1368–1370.
4.9.2. E. tabeti Arambourg, 1970
In the Lower Pleistocene of Africa, the only well-documented species is the Algerian E. tabeti [124]. It originates from Aïn Hanech strata which overlay Aïn Boucherit where another species, E. numidicus, was described by Pomel in 1897.
E. tabeti may be related to E. numidicus, a somewhat larger and less slender North African species found in more ancient levels. Due to the proximity of Aïn Boucherit in older levels, some fossils from either site were found inside the levels of the other.
Aïn Hanech may not be older than Layer L of the Thomas Quarry, the age of which is estimated at 1.0 to 1.5 Ma [125], or as old as 1.72 to 1.77 Ma [126].
Description
There is no skull. The species is well characterized by very gracile limb bones [127]. Its cheek teeth are small (Figure 101 and Figure 102A), with short protocones on the uppers [52]. The P2–M1 Han 1229 look like a smaller version of E. numidicus M1–M3 (Figure 102C); the upper Han 770 (Figure 99) are not very different from E. melkiensis.
The lower cheek teeth have stenonine double knots (Figure 102B) and usually deep vestibular grooves on the lower molars [53].
The metatarsals are as slender as in Hemiones; they do not resemble Asses at all (Figure 103B). On the other hand, the metacarpals, also very slender, have the deep diaphyses and proximal ends displayed by E. melkiensis (Figure 103A).
Concluding remarks
Given all the uncertainties noted above, the status of E. (or Allohippus?) tabeti remains obscure.
Fossils resembling E. tabeti and/or E. numidicus occur in member G of the Shungura Formation (Omo Valley, Ethiopia), in the Koobi Fora Formation (East Turkana, Kenya), in Bed I of Olduvai (Tanzania), and at Ubeidiyeh (Israel).
4.9.3. E. nalaikhaensis Kuznetsova and Zhegallo, 1996 [128]
This species was described by [129] on the basis of a nearly complete cranium PIN 3747-500 of a very old male (Figure 104).
Description
Cranium
The cranium is large, (545 mm for the Basilar length), narrow, and has a long muzzle and a rounded supra-occipital crest.
The Simpson’s diagram shows resemblances with E. apolloniensis (Figure 105), but the craniofacial angle is much larger in E. nalaikhaensis.
At least two other species are represented at Nalaikha [130] with different morphologies of cheek teeth (https://vera-eisenmann.com/nalaikha (accessed on 23 October 2021)) and metapodials. However, the upper cheek teeth referred to E. nalaikhaensis (Figure 106) have short protocones, shallow post-protoconal grooves, and small plis caballin, and the M3 bears a hypoconal islet; all these are characteristics found in Asses. The associated fauna suggests Nalaikha was no younger than 0.9 Ma.
4.9.4. Equus sp. from Konso, Ethiopia
A well-preserved cranium belonging to a male, KGA 8-374, was found at Konso, Ethiopia [131].
The chronological order of the deposits inside the Konso Formation, Southern Ethiopia was established by Gen Suwa et al. [132,133]. Inside levels ca. 1.3 Ma old was found a complete cranium of Equus belonging to an adult male. The cranium KGA 8- 374 is about the size of E. (Dolichohippus) grevyi (Figure 107) with a Basilar length of 529 mm.
The Simpson’s diagrams (Figure 108) show that the cranium from Konso is like a much larger E. (Asinus) africanus. It has a wide supra-occipital crest (lambdoid crest), a wide muzzle breadth at the interalveolar borders, and a shallow face. It differs from E. (Asinus) africanus by its longer muzzle and its small cranial height behind the orbits.
The upper cheek teeth (Figure 109) resemble some of the levels 23–24 and 36 of the contemporary Ubeidiyeh II, Israel [134].
5. Remarks on the Recurrence of Some Morphological Patterns
Among the Equid upper cheek teeth patterns, one of the easiest to recognize is that of E. (Asinus) melkiensis. This ‘melkiensis pattern’ (Figure 17B–D, Figure 18, Figure 19, Figure 20, Figure 21, Figure 26, Figure 27 and Figure 34) is mostly found at southern localities: in Morocco, Algeria, Tunisia, Yemen, Israel, Italy, Greece, Tadjikistan, and Uzbekistan. However, it also occurs at Süssenborn (Figure 73A).
Among the lower cheek teeth patterns, four are remarkable. One is the occurrence of the ‘bridge’ between the metaconid and metastyld noted by Groves [54] in E. (Asinus) africanus somaliensis (Figure 14B), present also in E. (Asinus) melkiensis (Figure 17E), E. ovodovi (Figure 88A), and some extant hemiones ([29], Figure 4-3). Another is the elongated metaconid with a wide base present in E. (Asinus) melkiensis (Figure 17F), and in E. marxi (Figure 100D,E). The third is the occasionally extreme depth of vestibular valleys that occurs in E. (Sussemionus) verae (Figure 72B), E. (Sussemionus) sussenbornensis (Figure 74B), and perhaps in E. ovodovi (Figure 88D). The fourth one is the occurrence of plis prtostylid on the p2 in E. Dolichohippus grevyi (Figure 43B), E. (Sussemionus) aff. sussenbornensis (Figure 76D), E. granatensis (Figure 82C), E. ovodovi (Figure 88C), and E. vekuae (Figure 95A).
Unlike morphological resemblances resulting from adaptations to similar environmental conditions, the occurrence of the same patterns in the species cited above must reflect some genetic proximities. Some may have been as close as between two sister taxa, other than the result of occasional interbreeding. The same mechanisms may explain the occurrence of Asinine features in the crania of E. (Sussemionus) coliemensis, E. nalaikhaensis, E. apolloniensis, and Equus sp. from Konso and the resemblances between the internal organs of Mountain Zebras and Asses. However, how closely these species are related is a question difficult or even impossible to answer.
6. Conclusions
According to reliable paleontological data, the following points are clear.
- Age of modern Equus subgeneraThe oldest fossils reliably related to extant African species are not older than 1 Ma:
- -
- The age of the fossil cranium of E. (Dolichohippus) grevyi from the Kapthurin Formation falls between 547 and 396 ka [13]; after recording and discussing the “proposed fossil occurrences of E. grevyi”, it is conclusively argued that most of them are not characteristic enough to be referred to that taxon, and certainly not those older than ca. 500 ka.
- -
- -
- -
- -
Approximately the same age may be supposed also for Hemiones and Horses, though E. (Sussemionus) may have appeared earlier [28].
- 2.
- Age of genus Equus
The first appearance of the genus Equus [7] is attested in the New World by a cranium found at Anza Borrego ca. 2 Ma old [122] and in the Old World by a cranium found at Dmanisi ca. 1.7 Ma old [118,119]. The latter is somewhat smaller but rather similar in proportions (Figure 74).
- 3.
- In between 1 Ma–2 Ma Equid species
In between the well-ascertained ‘modern’ Equus (younger than 1 Ma) and the first Old World true Equus (ca. 1.77 Ma), there are many slender forms with small protocones that are difficult to classify.
They were found, in particular, at: Nalaïkha (Mongolia), Dmanisi and Akhalkalaki (Georgia), Ubeidiyeh (Israel), Aïn Hanech (Algeria), Pirro (Italy), Venta Micena (Spain), and Untermassfeld and Süssenborn (Germany). While cranial proportions bring evidence that E. nalaikhaensis and E. vekuae certainly belonged to Equus there may be some doubt about others.
The species from Dmanisi, Pirro, Venta Micena, and Süssenborn are usually referred to E. altidens—a badly defined Equid from Süssenborn. Under the informal name of Pseudohydruntines, I group E. vekuae from Dmanisi, E. granatensis from Venta Micena, E. aff. granatensis from Pirro, E. hipparionoides from Akhalalkalaki, and E. wuesti from Untermassfeld. I believe that they belong to the ‘unique basal lineage of Old World Equids’ discovered by molecular biology [22] and that they are phyletically close to the subgenus Sussemionus.
Given our present state of knowledge of Plesippus and Allohippus and given the intricate morphological similarities inside Equus, I doubt a satisfying phylogenetical tree can be obtained even with the most sophisticated methods. However, any effort in that direction should include Sussemiones and all extant Equus, take into account the findings of molecular biology, and maybe even consider the surprising resemblances between modern Asses and Mountain Zebras [21,23].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/quat5030038/s1, Figure S1. Profiles of Equid crania. A: E. (Equus) caballus, MS 96872; B: Plesippus shoshonensis, LACM 210-17047; C: Allohippus vireti, QSV 240; Figure S2. Schematic occlusal view of an Equus cranium. 1: basilar length. 2: overall palatal lengh. 2-5: palatal length sensu stricto. 3: vomerine length. 4: post-vomerine length. 5. muzzle length. Measurements defined in Eisenmann 1986 [17]; Figure S3. Occlusal views of upper cheek teeth of E. cf. suessenbornensis. A: IQW S 1964/1182, left P3 or P4; B: IQW Süss. 7189, left P3 or P4; C: IQW Süss. 5617, left P4 or M1; D: IQW Süss. 7222, left P3 or P4; E: IQW Süss. 9230, left P3 or P4; F: IQW Süss. 6584, left P4 or M1; G: IQW Süss. 9264, right P3 or P4; H: IQW Süss. 9260, right P3 or P4; Figure S4. Occlusal views of lower cheek teeth of E. cf. suessenbornensis. A: IQW Süss. 6448, right P3 or P4; B: IQW Süss. 6222, left P3 or P4; C: IQW Süss. 6566, left P4 or M1; D: IQW Süss. 7204, left M1 or M2; Figures S5 to S14 collect the pictures and captions published by Reichenau [105] for ‘Hippotigris altidens’ and photographs and accession numbers of the teeth when they could be traced in the Institut von Quartär Paläontologie collections in Weimar [135]; Figure S15. Occlusal views of upper cheek teeth of E. cf. wuesti. All the accession numbers should be prefixed by IQW S; Figure S16. Occlusal views of lower cheek teeth of E. cf. wuesti. All the accession numbers should be prefixed by IQW; Figure S17. Pictures and captions published by Reichenau [105] for Hippotigris marxi and photographs and accession numbers of the teeth when they could be traced in the Institut von Quartär Paläontologie collections in Weimar; Figure S18. Occlusal views of upper cheek teeth of E. marxi. All the accession numbers should be prefixed by IQW S. IQW S 3225 to S HA: Premolars. IQW S 228 to 7297: Molars; Figure S19. Occlusal views of lower cheek teeth of E. marxi. All the accession numbers should be prefixed by IQW S. IQW S 851 to 14461: Premolars. IQW S 886 to no number: Molars; Figure S20. Simpson’s diagrams of Dmanisi slender metacarpals. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle; Figure S21. Simpson’s diagrams of Dmanisi Dm70/34.3A4.45 and Dm8/151.1/A4.26 (probably juvenile metacarpals. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle; Figure S22. Simpson’s diagrams of Dmanisi metacarpals from Layers II, III, and Stratum B. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle; Figure S23. Simpson’s diagrams of Dmanisi metatarsals from Layers IV-V, and Stratum A. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle; Table S1 Taxonomy Excel; Table S2 Typology E. hydruntinus; Table S3 Crania from Apollonia; Table S4 Typology Pseudhydruntines; Table S5 MC of pseudhydruntines; Table S6 Data and Scatter diagram of Dmanissi Strata A and B upper cheek teeth; Table S7 CraLog Dmanissi; Table S8 Data and Scatter diagram of Dmanissi lower cheek teeth; Table S9 MC Dmanisi; Table S10 MT Dmanissi; Table S11 First phalanges Dmanisi Data.
Funding
This research received no external funding.
Acknowledgments
I cannot thank in detail all the colleagues and curators of collections who helped me to gather the data I used. I am most grateful to M.-T. Alberdi, N. Aouadi, M. Bukhsianidze, A. Burke, O. Cirilli, R. Ferring, D. Geraads, A. Gkeme, L. Kolska, G. Koufos, G. Mangano, M. Mashkour, L Maul, H. Nakaya, M.-R. Palombo, Y. Sam, S. Sen and V. Ttov who all helped by answering questions and sending reprints and/or photographs. I thank also Eric Delson for his advice and help in matters of taxonomy, bibliography, and acronyms for Museums. The outsider view of my son led me to rewrite some paragraphs in the Introduction and Conclusion. Last but not least many thanks are due to the Reviewers and Editors of this work.
Conflicts of Interest
The author declares no conflict of interest.
References
- Churcher, C.S.; Richardson, M.L. Equidae. In Evolution of African Mammals; Maglio, V.J., Basil, H., Cooke, S., Eds.; Harvard University Press: Cambridge, MA, USA, 1978. [Google Scholar]
- MacFadden, B.J. Fossil Horses. In Systematics, Paleobiology, and Evolution of the Family Equidae; Cambridge University Press: Cambridge, UK, 1994; p. 369. [Google Scholar]
- MacFadden, B.J. Fossil Horses. In Systematics, Paleobiology, and Evolution of the Family Equidae, 2nd ed.; Cambridge University Press: Cambridge, UK, 1999; p. 369. [Google Scholar]
- Barron-Ortiz, C.I.; Avilla, L.S.; Jass, C.N.; Bravo-Cuevas, V.M.; Machado, H.; Mothé, D. What is Equus? Reconciling Taxonomy and Phylogenetic Analyses. Front. Ecol. Evol. 2019, 7, 343. [Google Scholar] [CrossRef]
- Cirilli, O.; Pandolfi, L.; Rook, L.; Bernor, R.L. Evolution of OLd World Equus and origin of the zebra-ass clade. Sci. Rep. 2021, 11, 10156. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, O.; Saarinen, J.; Pandolfi, L.; Rook, L.; Bernor, R.L. An updated review on Equus stenonis (Mammalia, Perissodactyla): New implications for European early Pleistocene Equus taxonomy and paleoecology, and remarks on the Old World Equus evolution. Quat. Sci. Rev. 2021, 269, 107155. [Google Scholar] [CrossRef]
- Eisenmann, V.; Baylac, M. Extant and fossil Equus (Mammalia, Perissodactyla) skulls: A morphometric definition of the genus Equus. Zologica Scr. 2000, 29, 89–100. [Google Scholar] [CrossRef]
- Samson, P. Les Equidés fossiles de Roumanie. Geol. Romana 1975, 14, 165–352. [Google Scholar]
- Forsten, A.; Eisenmann, V. Equus (Plesippus) simplicidens (Cope), not Dolichohippus. Mammalia 1995, 59, 85–89. [Google Scholar] [CrossRef]
- Eisenmann, V.; Deng, T. Equus qingyangensis (Equidae, Perissodactyla) of the Lower Pleistocene of Bajiazui, China: Evidence for the North American origin of an Old World lineage distinct from E. stenonis. Quaternaire 2005, 2, 113–122. [Google Scholar]
- Eisenmann, V.; Delson, E. The Senèze Equids. In Senèze: Life in Central France Two Million Years Ago. Paleontology, Geochronology, Stratigraphy and Taphonomy; Delson, E., Faure, M., Guérin, C., Eds.; Springer: Cham, Switzerland.
- Bernor, R.L.; Cirilli, O.; Jukar, A.M.; Potts, R.; Buskianidze, M.; Rook, L. Evolution of Early Equus in Italy, Georgia, the Indian Subcontinent, East Africa, and the Origins of African Zebras. Front. Ecol. Evol. 2019, 7, 65. [Google Scholar] [CrossRef]
- O’Brien, K.; Tryon, C.A.; Blegen, N.; Kimeu, B.; Rowan, J.; Faith, T.J. First appearance of Grévy’s zebra (Equus grevyi), from the Middle Pleistocene Kapthurin Formation, Kenya, sheds light on the evolution and ecology of large zebras. Quat. Sci. Rev. 2021, 256, 106835. [Google Scholar] [CrossRef]
- Gkeme, A.G.; Koufos, G.D.; Kostopoulos, D.S. Reconsidering the Equids from the Early Pleistocene fauna of Apollonia 1 (Mygdonian Basin, Greece). Quaternary 2021, 4, 12. [Google Scholar] [CrossRef]
- Wang, S.-Q.; Deng, T. Some evolutionary trends of Equus eisenmannae (Mammalia, Perissodactyla) in the stratigraphic sequence of Longdan, China, in comparison to modern Equus. J. Vertebr. Paleontol. 2011, 31, 1356–1365. [Google Scholar] [CrossRef]
- Eisenmann, V.; Mashkour, M. Data base for Teeth and Limb Bones of Modern Hemiones. In Fiches d’Ostéologie Animale Pour l’Archéologie; Série B: Mammifères; Centre de Recherches Archéologiques du CNRS, APDCA: Paris, France, 2000; Volume 9, p. 46. [Google Scholar]
- Eisenmann, V. Comparative osteology of modern and fossil Horses, Halfasses and Asses. In Equids in the Ancient World; Measow, R.H., Uerpmann, H.P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A: Wiesbaden, Germany, 1986; pp. 67–116. [Google Scholar]
- Simpson, G.G. Large pleistocene felines of North America. Am. Mus. Novit. 1941, 1136, 1–27. [Google Scholar]
- Bourdelle, E. Chevaux, Anes et Zèbres. Bull. Société Natl. D’acclimatation Fr. 1944, 91, 49–71. [Google Scholar]
- Bourdelle, E. Notes anatomiques et considérations zoologiques sur les Zèbres. Actes Congrès Natl. Sociétés Savantes 1934, 67, 99–112. [Google Scholar]
- Bourdelle, E. Quelques caractères anatomiques du Zèbre de Hartmann (E. zebra hartmannae Matschie). Bull. Mus. Natn. Hist. Nat. 1936, 8, 473–477. [Google Scholar]
- Orlando, L.; Metcalf, J.L.; Alberdi, M.T.; Telles-Antunes, M.; Bonjean, D.; Otte, M.; Martin, F.; Eisenmann, V.; Mashkour, M.; Morello, F.; et al. Revising the recent evolutionary history of equids using ancient DNA. Proc. Natl. Acad. Sci. USA 2009, 106, 21754–21759. [Google Scholar] [CrossRef] [Green Version]
- Eisenmann, V.; Turlot, J.C. Sur la taxinomie du genre Equus (Equidés). Les Cahiers de l’Analyse des Données 1978, 3, 179–201. [Google Scholar]
- Groves, C.P.; Grubb, P. Ungulate Taxonomy; The Johns Hopkins University Press: Baltimore, MD, USA, 2011; p. 309. [Google Scholar]
- Jónsson, H.; Schubert, M.; Seguin-Orlando, A.; Ginolhac, A.; Petersen, L.; Fumagalli, M.; Albrechtsen, A.; Petersen, B.; Korneliussen, T.S.; Vilstrup, J.T.; et al. Speciation with gene flow in equids despite extensive chromosomal plasticity. Proc. Natl. Acad. Sci. USA 2014, 111, 18655–18660. [Google Scholar] [CrossRef]
- Bennett, E.A.; Champlot, S.; Peter, J.; Arbuckle, B.; Guimaraes, S.; Pruvost, M.; Bar-David, S.; Davis, S.; Gautier, M.; Kaczensky, P.; et al. Taming the Late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA. Proc. Natl. Acad. Sci. USA 2009, 106, 21754–21759, Erratum in PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Groves, C.P.; Willoughby, D.P. Studies on the taxonomy and phylogeny of the genus Equus. Mammalia 1981, 45, 321–354. [Google Scholar] [CrossRef]
- Eisenmann, V. Pliocene and Pleistocene Equids: Paleontology versus Molecular Biology. In Late Neogene and Quaternary Biodiversity and Evolution, Regional Developments and Interregional Correlations. Proceedings Volume of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar), Weimar, Germany, 20–25 April 2004; Kahlke, R.-D., Maul, L.-C., Mazza, P., Eds.; Courier Forschungsinstitut Senckenberg: Frankfurt, Germany, 2006; p. 256. [Google Scholar]
- Eisenmann, V.; Howe, J.; Pichardo, M. Old World Hemiones and New World slender species (Mammalia, Equidae). Palaeovertebrata 2008, 36, 159–233. [Google Scholar] [CrossRef]
- Vilstrup, J.T.; Seguin-Orlando, A.; Stiller, M.; Ginolhac, A.; Raghavan, M.; Nielsen, S.C.A.; Weintstock, J.; Froese, D.; Vasiliev, S.K.; Ovodov, N.D.; et al. Mitochondrial phylogenomics of modern and ancient. PLoS ONE 2013, 8, e55950. [Google Scholar] [CrossRef]
- Eisenmann, V. Etude des cornets des dents incisives inférieures des Equus actuels et fossiles. Palaeontogr. Ital. 1979, 71, 55–75. [Google Scholar]
- Orlando, L.; Mashkour, M.; Burke, A.; Douady, C.J.; Eisenmann, V. Geographic distribution of an extinct equid (E. hydruntinus, Mammalia, Equidae) revealed by morphological and genetical analyses of fossils. Mol. Ecol. 2006, 15, 2083–2093. [Google Scholar] [CrossRef]
- Burke, A.; Eisenmann, V.; Ambler, G. The systematic position of Equus hydruntinus, an extinct species of Pleistocene equid. Quat. Res. 2003, 59, 459–469. [Google Scholar] [CrossRef]
- Eisenmann, V.; Mashkour, M. The small equids of Binagady (Azerbaidjan) and Qazvin (Iran): E. hemionus binagadensis nov. subsp., and E. hydruntinus. Géobios 1999, 32, 105–122. [Google Scholar] [CrossRef]
- Bonifay, M.-F. Equus hydruntinus Regalia minor n.ssp. from the caves of Lunel-Viel (Hérault, France). In Equids In the Ancient World; Meadow, R.H., Uerpmann, H.-P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients, Reihe A (Naturwissenschaften), Nr. 19/2; Dr Ludwig Reichert Verlag: Wiesbaden, Germany, 1991; Volume 2. [Google Scholar]
- van Asperen, E.N.; Stefaniak, K.; Proskurnyak, L.; Ridush, B. Equids from Emine-Bair-Khosar cave (Crimea, Ukraine): Co-occurrence of the stenonid Equus hydruntinus and the caballoid E. ferus latipes based on skull and postcranial remains. Palaeontol. Electron. 2012, 15, 28. [Google Scholar] [CrossRef]
- Boulbes, N.; van Asperen, E.N. Biostratigraphy and Palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, M.T.; Palombo, M.R. The Early Pleistocene Equidae from Pirro Nord (Apricena, Southern Italy). Paleontographica, Abt. A: Paleozoolgy Stratigr. 2013, 298, 147–167. [Google Scholar] [CrossRef]
- Tsoukala, E. Contribution to the study of the Pleistocene fauna of large mammals (Carnivora, Perissodactyla, Artiodactyla) from Petralona Cave (Chalkidiki, N. Greece). Preliminary report. C. R. Acad. Sci. 1991, 312, 331–336. [Google Scholar]
- Stehlin, H.G.; Graziosi, P. Ricerche sugli Asinidi fossili d’Europa. Mémoires Société Paléontologique Suisse 1935, 56, 1–73. [Google Scholar]
- Antunes, M.T. The Zebro (Equidae) and its extinction in Portugal, with an Appendix on the noun zebro and the modern “zebra”. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; Volume 17, pp. 211–236. [Google Scholar]
- Blanc, G.A. Sulla esistenza di “Equus (Asinus) hydruntinus” Regalia nel Pleistocene del Nord Africa. Bolletino Soc. Geol. Ital. 1956, 75, 3–14. [Google Scholar]
- Bibikova, V.I.; Vereshchagin, N.K.; Garutt, V.E.; Iourv, K.B. Novye materialy po chetvertichnoj faune Zabaijkalia. Mater. Arkheologiii SSSR 1953, 39, 463–475. [Google Scholar]
- Vogt, T.; Erbajeva, H. Premières preuves de conditions périglaciaires au Pléistocène inférieur en Transbaïkalie (Sibérie, Russie). C. R. Acad. Sci. Paris 1995, 320, 861–866. [Google Scholar]
- Foronova, I. Late quaternary equids (genus Equus) of South-western and South-central Siberia. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; Volume 2, pp. 20–30. [Google Scholar]
- Boule, M.; de Chardin, T.P. Paléontologie, Equus hemionus. In Le paléolithique de la Chine; Archives de l’institut de Paléontologie Humaine; Masson: Paris, France, 1928; Volume 4, pp. 36–46. [Google Scholar]
- Zhou, X.; Sun, Y.; Wang, Z.; Wang, H. Dalian Gulongshan Yizhi Yanjiu (Gulongshan Cave Site, A Upper Paleolithic site at Dalian City); Beijing Kexue Jishu Chubanshe (Beijing Scientific and Technical Publishing House): Beijing, China, 1990; p. 94. [Google Scholar]
- Sam, Y. African origins of modern asses as seen from paleontology and DNA: What about the Atlas wild ass? Geobios 2020, 58, 73–84. [Google Scholar] [CrossRef]
- Ducos, P. The Equid from Tell Muraibit, Syria. In Equids in the Ancient World; Meadow, R.H., Uerpmann, H.P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A.: Wiesbaden, Germany, 1986; pp. 237–245. [Google Scholar]
- Uerpmann, H.-P. Equus africanus in Arabia. In Equids in the Ancient World, Vol. II; Meadow, R.H., Uerpmann, H.-P., Eds.; Beihefte zum Tübinger Atlas des Vorderen Orients; Reihe A (Naturwissenschaften); Dr. Ludwig Reichert Verlag: Wiesbaden, Germany, 1991; pp. 12–33. [Google Scholar]
- Astre, G. Petit Asinien Pléistocène du Lauraguais. Bull. Société D’histoire Nat. Toulouse 1948, 83, 118–128. [Google Scholar]
- Eisenmann, V. Les Chevaux (Equus Sensu Lato) Fossiles et Actuels: Crânes et Dents Jugales Supérieures. Cahiers de Paléontologie; Editions du CNRS: Paris, France, 1980; 186p. [Google Scholar]
- Eisenmann, V. Etude des dents jugales inférieures des Equus actuels et fossiles. Palaeovertebrata 1981, 10, 3–4. [Google Scholar]
- Groves, C.P. Taxonomy. In Sull’asino Selvatico Africano; Groves, C.P., Ziccardi, F., Toschi, A., Eds.; Supplemento alle Ricerche di Zoologia Applicata alla Caccia; Universita di Bologna: Bologna, Italy, 1966; Volume 5, pp. 2–11. [Google Scholar]
- Thomas, P. Recherches stratigraphiques et paléontologiques sur quelques formations d’eau douce de l’Algérie. Mémoires Société Géologique Fr. 1884, 3, 1–51. [Google Scholar]
- Boule, M. Observations sur quelques équidés fossiles. Bull. Société Géologique Fr. 1900, 27, 531–542. [Google Scholar]
- Bagtache, B.; Hadjouis, D.; Eisenmann, V. Présence d’un Equus caballin (E. algericus n. sp.) et d’une autre espèce nouvelle d’Equus (E. melkiensis n. sp.) dans l’Atérien des Allobroges, Algérie. C. R. Ac. Sc. Paris 1984, 298, 609–612. [Google Scholar]
- Ginsburg, L.; Hilly, J.; Taquet, P. Une faune würmienne dans un remplissage de fente du massif du Filfila (littoral nord-constantinois, Algérie). C. R. Somm. Séances Soc. Géol. Fr. 1968, 5, 157. [Google Scholar]
- Souville, G. Atlas Préhistorique de l’Algérie, Feuille N°5P; Jourdan, A., Ed.; Gouvernement Général de l’Algérie: Alger, Algérie, 1958. [Google Scholar]
- Ennouchi, E. La faune des limons rouges de Bou-Knadel (Maroc). In Actes du IVème Congrès International du Quaternaire: 3–4; Août—Septembre: Rome, Italy, 1953. [Google Scholar]
- Arambourg, C. Mammifères fossiles du Maroc. Mémoires Soc. Sci. Nat. Maroc 1938, 46, 1–74. [Google Scholar]
- Tchernov, E. An attempt to synchronize the faunal changes with the radiometricdates and the cultural chronology in Southwest Asia. In Archaeozoology of the Near East III; ARC Publicaties: Groningen, The Netherlands, 1998; Volume 18, pp. 7–44. [Google Scholar]
- Marom, N.; Lazagabaster, I.A.; Horwitz, L.K. The late middle pleistocene mammalian fauna of Oumm Qatafa cave, Judean desert: Taphonomy and paleoenvironment. J. Quat. Sci. 2022, 37, 612–638. [Google Scholar] [CrossRef]
- Dodonov, A.; Zhegallo Penkov, V.A.; Maria Sotnikova, M. Stratigrafia mestonakhozhdenij pozdnepliotsenovykh pozvonochnykh Iuzhnogo Tadjikistana. Izvestia Akademii Nauk SSSR Serija Geologicheskaja 1991, 5, 12–20. (In Russian) [Google Scholar]
- Sotnikova, M.V.; Baigusheva, V.S.; Titov, V.V. Carnivores of the Khapry faunal assemblage and their stratigraphic implications. Stratigr. Geol. Correl. 2002, 10, 375–390. [Google Scholar]
- Gromova, V. Istorija Loshadej (roda Equus) v Starom Svete. Chast’ 1. Obzor i Opisanie Form; Trudy Paleontological Institute Akademi Nauk SSSR: Moscow, Russia, 1949; Volume 17, p. 373. (In Russian) [Google Scholar]
- Hadjouis, D. La faune mammaliennne d’Algérie, un renouvellement constant. Préhistoire Algérienne. Dossiers Archéol. 2003, 282, 42–53. [Google Scholar]
- Aouadi, N.; Bkhairi, A.; Belhouchhet, L.; Khedaer-El-Asmi, R. Les sites préhistoriques de l’Aïn Metherchem (Foussana, Kasserine) revisités. Bull. Mus. Anthropol. préhist. Monaco 2014, 54, 137–151. [Google Scholar]
- Van Couvering, J.A.; Delson, E. African land mammal ages. J. Vertebr. Paleontol. 2020, 40, e1803340. [Google Scholar] [CrossRef]
- Azzaroli, A. On a Late Pleistocene Ass from Tuscany; with notes on the History of Asses. Palaeontographia Italica 1979, 71, 27–47. [Google Scholar]
- Azzaroli, A. Pleistocene and living horses of the Old World. Palaeontogr. Ital. 1966, 61, 1–15. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, Macedonia, Greece. Paleontol. Evol. 1997, 30–31, 49–76. [Google Scholar]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Eisenmann, V. Le protostylide: Valeur systématique et signification phylétique chez les espèces actuelles et fossiles du genre Equus (Perissodactyla, Mammalia). Z. Säugetierkunde 1976, 41, 349–365. [Google Scholar]
- Eisenmann, V. Equus capensis (Mammalia, Perissodactyla) from Elandsfontein. Palaeontol. Afr. 2000, 36, 91–96. [Google Scholar]
- Geraads, D.; Hublin, J.J.; Jaeger, J.J.; Tong, T.; Sen, S.; Toureau, P. The Pleistocene hominid site of Ternifine, Algeria: New results on the environment, age, and human industries. Quat. Res. 1986, 25, 380–386. [Google Scholar] [CrossRef]
- Potts, R.; Behrensmeyer, A.K.; Tyler Faith, J.; Tryon, C.A.; Brooks, A.S.; Yellen, J.E. Environmental dynamics during the onset of the Middle Stone Age in eastern Africa. Science 2018, 360, 86–90. [Google Scholar] [CrossRef]
- Churcher, C.S.; Hooijer, D.A. The Olduvai zebra (Equus oldowayensis) from the Later Omo Beds, Ethiopia. Zool. Meded. Rijksmus. Nat. Hist. Leiden 1980, 55, 265–280. [Google Scholar]
- Eisenmann, V.; De Giuli, C. Caractères distinctifs entre vrais Zèbres (Equus zebra) et Zèbres de Chapman (Equus burchelli antiquorum) d’après l’étude de 60 têtes osseuses. Mammalia 1974, 38, 509–543. [Google Scholar] [CrossRef]
- Lundholm, B. Equus zebra greadheadi n. subsp., a new South African fossil zebra. Ann. Transvaal Mus. 1952, 22, 25–27. [Google Scholar]
- Lazarev, P.A. Antropogenovye Loshadi Iakutii; Nauka: Moscow, Russia, 1980; p. 190. (In Russian) [Google Scholar]
- Kuzmina, I.E. Late Pleistocene horse of the Ussurian region. Trudy Zool. Inst. Akad. Nauk SSSR. 1990, 212, 49–59. (In Russian) [Google Scholar]
- Cardoso, J.L. Les grands Mammifères du Pléistocène supérieur du Portugal. Essai de synthèse. Géobios 1996, 29, 235–250. [Google Scholar] [CrossRef]
- Eisenmann, V.; Crégut-Bonnoure, E.; Moigne, A.-M. Equus mosbachensis et les grands Chevaux de la Caune de l’Arago et de Lunel-Viel: Crâniologie comparée. Bull. Mus. Nat. Hist. Nat. 1985, 7, 157–173. [Google Scholar]
- Eisenmann, V. Proportions squelettiques de Chevaux quaternaires et actuels. Géobios 1991, 13, 25–32. [Google Scholar] [CrossRef]
- Madriaga de la Campa, B. Estudio zootecnico de las pinturas rupestres en la region cantabrica. Zephyrus 1963, XIV, 29–45. [Google Scholar]
- Eisenmann, V.; David, F. Evolution de la taille des Chevaux d’Arcy-sur-Cure et de quelques autres Chevaux quaternaires. In L’Aurignacien de la Grotte du Renne; XXXIVème Supplément à Gallia Préhistoire; CNRS: Paris, France, 2002; pp. 97–102. [Google Scholar]
- Turbanti, L. Equus Caballus nel Pleistocene delle Puglie; Università degli Studi di Firenze: Firenze, Italy, 1982; Volume 78. [Google Scholar]
- Eisenmann, V. Discriminating Equus skulls: The Franck’s Index and the new Palatal Index. 9th ICAZ Conference, Durham 2002. In Equids in Time and Space; Mashkour, M., Ed.; Oxbow Books: Oxford, UK, 2006; pp. 172–182. [Google Scholar]
- Eisenmann, V. Origins, dispersals, and migrations of Equus (Mammalia, Perissodactyla). Cour. Forsch. Senckenberg 1992, 153, 161–170. [Google Scholar]
- Vangenheim, E.A.; Zazhigin, V.S. Obzor faunisticheskikh kompleksov i faun territorii SSSR. In Chetvertichnaya Sistema, 1; Shantser, E.V., Ed.; Nedra: Moscow, Russia, 1982; pp. 267–279. (In Russian) [Google Scholar]
- Forsten, A.; Ziegler, R. The horses (Mammalia, Equidae) from the early Wuermian of Villa Seckendorff, Stuttgart-Bad Cannstadt, Germany. Stuttg. Beiträge Nat. Ser. B 1995, 224, 1–22. [Google Scholar]
- Nehring, A. Fossile Pferde aus Deutschen Diluvial-Ablagerungen und ihre Beziehungen zu den lebenden Pferden: Ein Beitrag zur Geschichte des Hauspferdes. Landwirtsch. Jahrb. 1884, 13, 81–160. [Google Scholar]
- Philippe, M. La faune würmienne du gisement paléontologique de Siréjol à Gignac (Lot). Bull. Société Sci. Hist. Archéologique Corrèze 1975, 97, 1–9. [Google Scholar] [CrossRef]
- Cardoso, J.L.; Eisenmann, V. Equus caballus antunesi, nouvelle sous-espèce quaternaire du Portugal. Palaeovertebrata 1989, 19, 47–72. [Google Scholar]
- Eisenmann, V. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia). C. R. Biol. 2010, 333, 235–240. [Google Scholar] [CrossRef]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing Equid Body Mass Estimate Equations on Modern Zebras—With Implications to Understanding the Relationship of Body Size, Diet, and Habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9, 622412. [Google Scholar] [CrossRef]
- Sher, A.V. Mlekopitaiushchie i Stratigrafia Pleistotsena Krainego Severo-Vostoka SSSR i Severnoi Ameriki; Nauka: Moscow, Russia, 1971; p. 310. (In Russsian) [Google Scholar]
- Sher, A.V. Olyorian land mammal age of Northeastern Siberia. Palaeontogr. Ital. 1987, 74, 97–112. [Google Scholar]
- Wagner, G.A.; Maul, L.C.; Löscher, M.; Schreiber, H.D. Mauer—The type site of Homo heidelbergensis: Palaeoenvironment and age. Quat. Sci. Rev. 2011, 30, 1464–1473. [Google Scholar] [CrossRef]
- Maul, L.C.; Parfitt, S.A. Micromammals from the 1995 Mammoth Excavation at West Runton, Norfolk, UK: Morphometric data, biostratigraphy and taxonomic reappraisal. Quat. Int. 2010, 228, 91–115. [Google Scholar] [CrossRef]
- Musil, R. Die Equidenreste aus dem Pleistozän von Süssenborn bei Weimar. Paläontol. Abhandl. A 1969, 3–4, 617–666. [Google Scholar]
- Wüst, E. Untersuchungen über das Pliozän und das älteste Pleistozän Thüringens Nördlich vom Thüringer Walde und Westlich von der Saale. Abhandl. Natur. Gesellschaft Halle. 1901, XXIII, 566. [Google Scholar]
- Vekua, A.K. The Lower Pleistocene Mammalian Fauna of Akhalkalaki (Southern Georgia, USSR). Palaeontogr. Ital. 1986, 74, 63–96. [Google Scholar]
- Von Reichenau, W. Beiträge zur näheren Kenntnis fossiler Pferde aus deutschen Pleistozän, insbesondere über die Entwicklung und die Abkaustadien des Gebisses vom Hochterrassenpferd (Equus mosbachensis v.R.). Abh. Grossherz.-Hess. Geol. Landesanst. Darmstadt 1915, 7, 155. [Google Scholar]
- Alberdi, M.T.; Palombo, M.R. The late Early to early Middle Pleistocene stenonoid horses from Italy. Quat. Int. 2013, 288, 25–44. [Google Scholar] [CrossRef]
- Arzarello, M.; Peretto, C.; Moncel, M.-H. The Pirro Nord site (Apricena, Fg, Southern Italy) in the context of the first European peopling: Convergences and divergences. Quat. Int. 2015, 389, 255–263. [Google Scholar] [CrossRef]
- Maul, L.; Masini, F.; Parfitt, S.A.; Rekovetz, L.; Savorelli, A. Evolutionary trends in arvicolids and the endemic murid Mikrotia—New data and a critical overview. Quat. Sci. Rev. 2014, 96, 240–258. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Torres, T.; Ramon, J.; Delgado, A.; Llamas, J.; Soler, V.; Delgado, J. Numerical dating algorithm of amino acid racemization ratios from continental ostracodes. Application to the Guadix-Baza Basin (southern Spain). Qyaternary Sci. Rev. 2004, 23, 717–730. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Ruiz Bustos, A. Descripcion y significado biostratigrafico y climatico del Equus e Hippopotamus, en el yacimiento de Venta Micena (Granada). Estud. Geol. 1985, 41, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Duval, M.; Falguères, C.; Jean-Jacques Bahain, J.-J. Age of the oldest hominin sttelements in Spain: Contribution of the combined U-series/ESR dating method applied tyo fossil teeth. Quat. Geochronol. 2012, 10, 412–417. [Google Scholar] [CrossRef]
- De Giuli, C.; Masini, F.; Torre, D. The latest villafranchian faunas in Italy: The Pirro Nord fauna (Apricena, Gargano). Palaeontogr. Ital. 1987, 74, 51–62. [Google Scholar]
- De Giuli, C. Late villafranchian faunas of Italy: The Selvella local fauna in the southern Chiana valley—Umbria. Palaeontogr. Ital. 1987, 74, 11–50. [Google Scholar]
- Musil, R. Die Equiden-Reste aus dem Unterpleistozän von Untermaßfeld. In Das Pleistozän von Untermassfeld bei Meiningen (Thüringen); Kahlke, R.-D., Ed.; Teil 2; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2001; Volume 40, pp. 557–587. [Google Scholar]
- Kahlke, R.D.; Linnermann, U.; Gärtner, A. New results on the origin and geological history of the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thüringen, Germany); Kahlke, R.-D., Ed.; Part 4; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; Volume 40, pp. 1079–1104. [Google Scholar]
- Eisenmann, V.; Boulbes, N. New results on equids from the Early Pleistocene site of Untermassfeld. In The Pleistocene of Untermassfeld Near Meiningen (Thüringen, Germany); Kahlke, R.-D., Ed.; Part 4; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2020; Volume 40, pp. 1295–1322. [Google Scholar]
- Eisenmann, V.; Vasiliev, S. Unexpected finding of a new Equus species (Mammalia, Perissodactyla) belonging to a supposedly extinct subgenus in late Pleistocene deposits of Khakassia (southwestern Siberia). Geodiversitas 2011, 33, 519–530. [Google Scholar] [CrossRef]
- Lordkipanidze, D.; Jashashvili, T.; Vekua, A.; Ponce de Leon, M.S.; Zollikofer, C.P.E.; Rightmire, G.P.; Pontzer, H.; Ferring, R.; Oms, O.; Tappen, M.; et al. Postcranial evidence from early Homo from Dmanisi, Georgia. Nature 2007, 449, 305–310. [Google Scholar] [CrossRef]
- Ferring, R.; Oms, O.; Agusti, J.; Berna, F.; Nioradze, M.; Shelia, T.; Tappen, M.; Vekua, A.; Zhvania, D.; Lordkipanidze, D. Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. Proc. Natl. Acad. Sci. USA 2011, 108, 10432–10436. [Google Scholar] [CrossRef]
- Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D. The Dmanisi Equus: Systematics, biogeography, and paleoecology. J. Hum. Evol. 2021, 158, 1–14. [Google Scholar] [CrossRef]
- Vekua, A.K. Die Wirbeltierfauna des Villafranchium von Dmanisi und ihre biostratigraphische Bedeutung. Jahrb. Des Römisch-Ger. Zent. Mainz 1995, 42, 180. [Google Scholar]
- Downs, T.; Miller, G.J. Late Cenozoic Equids from Anza-Borrego Desert of Clifornia. Contrib. Sci. Nat. Hist. Mus. Los Angeles Cty. 1994, 440, 90. [Google Scholar]
- Arambourg, C. Les Vertébrés du Pléistocène de l’Afrique du Nord. Arch. Mus. Nat. Hist. Nat. 1970, 10, 1–127. [Google Scholar]
- Geraads, D.; Raynal, J.-P.; Eisenmann, V. The earliest human occupation of North Africa: A reply to Sahnouni et al. (2002). J. Hum. Evol. 2004, 46, 751–761. [Google Scholar] [CrossRef]
- Duval, M.; Sahnoini, M.; Parés, J.M.; van der Made, J.; Abdessadok, S.; Harichane, Z.; Chebeb, R.C.; Boulaghraif, K.; Pérez-Gonzales, A. The Plio-Pleistocene sequence of Oued Boucherit (Algeria): A unique chronologically—Constrained archaeological and palaeontological record in North Africa. Quat. Sci. Rev. 2021, 271, 1–23. [Google Scholar] [CrossRef]
- Eisenmann, V. Les métapodes d’Equus sensu lato (Mammalia, Perissodactyla). Géobios 1979, 12, 863–886. [Google Scholar] [CrossRef]
- Aouadi, N. Les Chevaux Fossiles Nord-Africains. Nouvelle Analyse, Nouvelle Phylogénie; Ministère de la Culture, de la Jeuness et des Loisirs: République Tunisienne, Africa, 2004; pp. 5–16. [Google Scholar]
- Kuznetsova, T.V.; Zhegallo, V.I. Taksonomicheskoie Raznoobrazie Ekvid Mestonakhojdenia Nalaikha (Mongolia); Trudy Mejdunarodnogo soveshchania ’Sostoianie Teriofauny v Rossii i Blijnem Zarubejie’ 1-3 fevralia 1995; Nauka: Moscow, Russia, 1996; pp. 48–53. (In Rusian) [Google Scholar]
- Eisenmann, V.; Kuznetsova, T. Early Pleistocene equids (Mammalia, Perissodactyla) of Nalaikha, Mongolia, and the emergence of modern Equus Linnaeus 1758. Geodiversitas 2004, 26, 535–561. [Google Scholar]
- Nakaya, H.; Suwa, G.; Ode, S.; Asfaw, B.; Beyene, Y. A near-complete skull of Equus (Equidae, Perissodactyla, Mammalia) from the Early Pleistocene Konso Formation, Southern Ethiopia. In Proceedings of the Program and Abstracts 73rd Annual Meeting, Society of Vertebrate Paleontology, Los Angeles, CA, USA, 2 November 2013. [Google Scholar]
- Suwa, G.; Nakaya, H.; Asfaw, B.; Saegusa, H.; Amzaye, A.; Kono, R.T.; Yonas Beyene, Y.; Katoh, S. Plio-Pleistocene terrestrial mammal assemblage from Konso, Southern Ethiopia. J. Vertebr. Paleontol. 2003, 23, 901–916. [Google Scholar] [CrossRef]
- Suwa, G.; Asfaw, B.; Katoh, S.; Nakaya, H.; Beyene, Y. Early Pleistocene fauna and paleoenvironments at Konso, Ethiopia. In African Paleoecology and Human Evolution; Reynolds, S., Bobe, R., Eds.; Cambridge University Press: Cambridge, MA, USA, 2022; pp. 269–277. [Google Scholar]
- Bar-Yosef, O.; Belmaker, M. Archeology of Human Evolution. In Quaternary of the Levant, Environments, Climate Change, and Humans; Cambridge Uniersity Press: Cambridge, MA, USA, 2017; pp. 179–186. [Google Scholar]
- Kahlke, R.D. Typencatalog zu den Sammlungen des Instituts für Quartaärpaläontologie Weimar. Abh. Ber. Mus. Not. Gotha. 1984, 12, 57–63. [Google Scholar]
- Schwartz, E. Über diluviale Pferde der Equus caballus-Gruppe. Jahrbuch Preussischen Geologischen Landesanstalt 1928, 48, 429–476. [Google Scholar]
Figure 1.
(A) Affinities between subgenera of extant Equus according to Bourdelle [19]. (B) Eisenmann and Turlot first factorial plane of a multifactorial analysis of correspondence of extant Equus crania [23], adapted from: Eisenmann, V. Pliocene and Pleistocene Equids: Paleontology versus Molecular Biology [28]. In Late Neogene and Quaternary biodiversity and evolution, Regional developments and interregional correlations, Eds. Ralph-Dietrich Kahlke, Lutz-Christian Maul, and Paul Mazza; Proceedings volume of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar), 20–25 April 2004. Courier Forschungsinstitut Senckenberg, Germany, 2006, 256.
Figure 1.
(A) Affinities between subgenera of extant Equus according to Bourdelle [19]. (B) Eisenmann and Turlot first factorial plane of a multifactorial analysis of correspondence of extant Equus crania [23], adapted from: Eisenmann, V. Pliocene and Pleistocene Equids: Paleontology versus Molecular Biology [28]. In Late Neogene and Quaternary biodiversity and evolution, Regional developments and interregional correlations, Eds. Ralph-Dietrich Kahlke, Lutz-Christian Maul, and Paul Mazza; Proceedings volume of the 18th International Senckenberg Conference (VI International Palaeontological Colloquium in Weimar), 20–25 April 2004. Courier Forschungsinstitut Senckenberg, Germany, 2006, 256.

Figure 3.
E. (Hemionus) hemionus hemionus cranium ZIN 27193, Ventral view.

Figure 4.
(A) E. (Hemionus) hemionus khur DDP3, P2–m3, occlusal view. (B) E. (Hemionus) hemionus kulan MGU 74785, p2–m3, occlusal view.
Figure 4.
(A) E. (Hemionus) hemionus khur DDP3, P2–m3, occlusal view. (B) E. (Hemionus) hemionus kulan MGU 74785, p2–m3, occlusal view.

Figure 16.
E. (Asinus) melkiensis, Les Allobroges, Algeria. (A,B) IPH Allo. 61-11314, type MC proximal and dorsal views; (C) IPH Allo. 61-13, first anterior phalanx, dorsal view; (D) IPH Allo. 61-1834, paratype MT dorsal view.
Figure 16.
E. (Asinus) melkiensis, Les Allobroges, Algeria. (A,B) IPH Allo. 61-11314, type MC proximal and dorsal views; (C) IPH Allo. 61-13, first anterior phalanx, dorsal view; (D) IPH Allo. 61-1834, paratype MT dorsal view.

Figure 17.
E. (Asinus) melkiensis. Upper and lower cheek teeth from Les Allbroges, Algeria, occlusal views. (A) IPH Allo. 61-109, P2, (B) IPH Allo. 61-2704, P3 o P4; (C) IPH Allo. 61-2855, M1; (D) IPH Allo. 61-775, P3 o P4; (E) IPH Allo. 61-196, m1 or m2; (F) IPH Allo. 61-1969, paratype paratype m1 or m2.
Figure 17.
E. (Asinus) melkiensis. Upper and lower cheek teeth from Les Allbroges, Algeria, occlusal views. (A) IPH Allo. 61-109, P2, (B) IPH Allo. 61-2704, P3 o P4; (C) IPH Allo. 61-2855, M1; (D) IPH Allo. 61-775, P3 o P4; (E) IPH Allo. 61-196, m1 or m2; (F) IPH Allo. 61-1969, paratype paratype m1 or m2.

Figure 18.
Upper P3 or P4 of E. (Asinus) melkiensis, occlusal views. (A) Filfila, MNHN-F no number. (B) Aïn Tit Mellil, Morocco, MNHN-F Moc 129. (C) Sidi Bou Kndel, Morocco, MNHN-F Moc 1961-1-181. (D) Yemen, MNHN-F no number.
Figure 18.
Upper P3 or P4 of E. (Asinus) melkiensis, occlusal views. (A) Filfila, MNHN-F no number. (B) Aïn Tit Mellil, Morocco, MNHN-F Moc 129. (C) Sidi Bou Kndel, Morocco, MNHN-F Moc 1961-1-181. (D) Yemen, MNHN-F no number.

Figure 19.
P2–P4, M2–M3 of E. (Asinus) melkiensis, Aïn Tit Mellil, MNHN-F Moc 106. Occlusal view.

Figure 20.
P2–M3 of E. (Asinus) melkiensis, Sidi Abderahmane, Morocco, MNHN-F Moc 1957-54. Occlusal view.
Figure 20.
P2–M3 of E. (Asinus) melkiensis, Sidi Abderahmane, Morocco, MNHN-F Moc 1957-54. Occlusal view.

Figure 21.
P3 or P4 of E. (Asinus) aff. melkiensis, occlusal views. (A) Petralona, Greece, LGPUT PEC 1715. (B) Oum Qatafa, Israel, HUJ-ESE OK 5. (C) E. (Asinus) melkiensis Aïn el Guettar, Tunisia, MNHN-F GUE 1.
Figure 21.
P3 or P4 of E. (Asinus) aff. melkiensis, occlusal views. (A) Petralona, Greece, LGPUT PEC 1715. (B) Oum Qatafa, Israel, HUJ-ESE OK 5. (C) E. (Asinus) melkiensis Aïn el Guettar, Tunisia, MNHN-F GUE 1.

Figure 22.
E. (Asinus) aff melkiensis, MNHN-F #282, upper (A) and lower (B) associated cheek teeth series from Tchad, occlusal views.
Figure 22.
E. (Asinus) aff melkiensis, MNHN-F #282, upper (A) and lower (B) associated cheek teeth series from Tchad, occlusal views.

Figure 23.
p3 or p4 of E. (Asinus) aff. melkiensis, occlusal view. Aïn Tit Mellil, MNHN-F Moc 16.

Figure 24.
Simpson’s diagrams of E. (Asinus) africanus africanus and E. (Asinus) melkiensis MC. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.
Figure 24.
Simpson’s diagrams of E. (Asinus) africanus africanus and E. (Asinus) melkiensis MC. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.

Figure 25.
Lakhuti II, PIN 3848-281, occlusal view of P2–M3.

Figure 26.
E. valeriani. (A) upper molar; (B) protocone; (C,D) lower premolars; (E) lower molar; (F) m3. Modified from Gromova, V. Istorija loshadej (roda Equus) v Starom Svete. Chast’ 1. Obzor i opisanie form. Trudy paleont. Inst., Akad. Nauk SSSR, Moskva, 1949, 17 (1): 373p.
Figure 26.
E. valeriani. (A) upper molar; (B) protocone; (C,D) lower premolars; (E) lower molar; (F) m3. Modified from Gromova, V. Istorija loshadej (roda Equus) v Starom Svete. Chast’ 1. Obzor i opisanie form. Trudy paleont. Inst., Akad. Nauk SSSR, Moskva, 1949, 17 (1): 373p.

Figure 27.
E. (Asinus) asinus skull MNHL 384, (A) dorsal view, (B) Ventral view, (C) Right lateral view.
Figure 27.
E. (Asinus) asinus skull MNHL 384, (A) dorsal view, (B) Ventral view, (C) Right lateral view.

Figure 28.
Simpson’s diagrams of E. (Asinus) africanus africanus and E. (Asinus) africanus somaliensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length.
Figure 28.
Simpson’s diagrams of E. (Asinus) africanus africanus and E. (Asinus) africanus somaliensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length.

Figure 29.
Occlusal views of upper and lower cheek series of E. (Asinus) asinus, Kutch, India, DD N20. (A) P2–M3. (B) p2–m3.
Figure 29.
Occlusal views of upper and lower cheek series of E. (Asinus) asinus, Kutch, India, DD N20. (A) P2–M3. (B) p2–m3.

Figure 30.
E. (Asinus) asinus, MNHN-F Guy 118, p2–m3 from Aïn Benian, occlusal view.

Figure 31.
E. (Asinus) asinus. Lower cheek teeth from Les Allobroges, occlusal views. (A) IPH Allo. 61-2950, right inverted p3 or p4; (B) IPH Allo. no number, associated m1 and m2; (C) IPH Allo. 61-788, m2.
Figure 31.
E. (Asinus) asinus. Lower cheek teeth from Les Allobroges, occlusal views. (A) IPH Allo. 61-2950, right inverted p3 or p4; (B) IPH Allo. no number, associated m1 and m2; (C) IPH Allo. 61-788, m2.

Figure 32.
E. (Asinus) sp. Upper and lower associated cheek teeth from Aïn Metherchem, archeological survey SPM3, Tunisia, courtesy by Dr Nabiha Aouadi, occlusal views. (A) Fragment of P2, P3, P4, M1, M2, M3. (B) p2, p3, p4, m1.
Figure 32.
E. (Asinus) sp. Upper and lower associated cheek teeth from Aïn Metherchem, archeological survey SPM3, Tunisia, courtesy by Dr Nabiha Aouadi, occlusal views. (A) Fragment of P2, P3, P4, M1, M2, M3. (B) p2, p3, p4, m1.

Figure 34.
E. (Asinus) graziosii, IGF 192V, type cranium, left lateral view modified from Ghedoghedo [CC BY-SA].
Figure 34.
E. (Asinus) graziosii, IGF 192V, type cranium, left lateral view modified from Ghedoghedo [CC BY-SA].

Figure 35.
Simpson’s diagrams of E. (Asinus) E. (Asinus) asinus, and E. (Asinus) graziosii crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the inter-alveolar crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 35.
Simpson’s diagrams of E. (Asinus) E. (Asinus) asinus, and E. (Asinus) graziosii crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the inter-alveolar crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 36.
E. (Asinus) graziosii, IGF 192V, occlusal view of P2–M3.

Figure 37.
E. apolloniensis, Apollonia, Greece. APL 148 type cranium, (A) Dorsal view; (B) Right lateral view. (C) APL 872 Cranium, Ventral view. Courtesy by A.G. Gmeke.
Figure 37.
E. apolloniensis, Apollonia, Greece. APL 148 type cranium, (A) Dorsal view; (B) Right lateral view. (C) APL 872 Cranium, Ventral view. Courtesy by A.G. Gmeke.

Figure 38.
Simpson’s diagrams of E. apolloniensis and E. (Asinus) africanus crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 38.
Simpson’s diagrams of E. apolloniensis and E. (Asinus) africanus crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 39.
E. apolloniensis, Apollonia, Greece. Occlusal views of (A) P4–M1 of APL 343 kindly communicated by A. Gkeme. (B) P2–M3 of APL 148 modified from Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, Macedonia, Greece. Paleontol. I Evol. 1997, 30–31, 49–76.
Figure 39.
E. apolloniensis, Apollonia, Greece. Occlusal views of (A) P4–M1 of APL 343 kindly communicated by A. Gkeme. (B) P2–M3 of APL 148 modified from Koufos, G.D.; Kostopoulos, D.S.; Sylvestrou, I.A. Equus apolloniensis n. sp. (Mammalia, Equidae) from the latest Villafranchian locality of Apollonia, Macedonia, Greece. Paleontol. I Evol. 1997, 30–31, 49–76.

Figure 40.
E. apolloniensis, Apollonia, Greece. Occlusal views of (A) APL 103 associated p4–m3. (B) APL 171, p2–m3. Courtesy by A.G. Gkeme.
Figure 40.
E. apolloniensis, Apollonia, Greece. Occlusal views of (A) APL 103 associated p4–m3. (B) APL 171, p2–m3. Courtesy by A.G. Gkeme.

Figure 41.
E. (Dolichohippus) grevyi skull, NMUK-ZD 1923.10.20.16. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 41.
E. (Dolichohippus) grevyi skull, NMUK-ZD 1923.10.20.16. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 42.
Simpson’s diagram of E. (Dolichohippus) grevyi, E. (Quagga) burchelli. and E. (Hippotigris) zebra. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 42.
Simpson’s diagram of E. (Dolichohippus) grevyi, E. (Quagga) burchelli. and E. (Hippotigris) zebra. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 43.
E. (Dolichohippus) grevyi occlusal views of cheek teeth, (A) NMUK-ZD 1893.12.1.2, P2–M3; (B) ZIN 5237, p2–m3.
Figure 43.
E. (Dolichohippus) grevyi occlusal views of cheek teeth, (A) NMUK-ZD 1893.12.1.2, P2–M3; (B) ZIN 5237, p2–m3.

Figure 44.
Simpson’s diagrams of limb bone proportions of E. grevyi, E. burchelli, and E. zebra. Maximal lengths of Humerus (H), Femur (F), Radius (R), Tibia (T), Third metacarpal (MC, Third Metatarsal (MT), First Anterior Phalanx (PhIA), First Posterior Phalanx (Ph1P) and plantar breadth of Third Anterior Phalanx (Ph3A (4)).
Figure 44.
Simpson’s diagrams of limb bone proportions of E. grevyi, E. burchelli, and E. zebra. Maximal lengths of Humerus (H), Femur (F), Radius (R), Tibia (T), Third metacarpal (MC, Third Metatarsal (MT), First Anterior Phalanx (PhIA), First Posterior Phalanx (Ph1P) and plantar breadth of Third Anterior Phalanx (Ph3A (4)).

Figure 45.
E. (Quagga) quagga skull PH 6317. (A) Ventral view; (B) Dorsal view; (C) Lateral view.

Figure 46.
E. (Quagga) quagga PH 6317 upper (A) and lower (B) cheek teeth series, occlusal views.

Figure 47.
E. (Quagga) mauritanicus cranium MNHN-F TER 1543, Ternifine, Algeria. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 47.
E. (Quagga) mauritanicus cranium MNHN-F TER 1543, Ternifine, Algeria. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 48.
Simpson’s diagram of E. (Quagga) quagga, E. (Quagga) mauritanicus, E. (Quagga) capensis and KNM-OG 28612 crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 48.
Simpson’s diagram of E. (Quagga) quagga, E. (Quagga) mauritanicus, E. (Quagga) capensis and KNM-OG 28612 crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 49.
E. (Quagga) mauritanicus dentitions from Ternifine, Algeria. (A) MNHN-F TER 154, P3–M3. (B) TER 1383, p2–m3. Occlusal views.
Figure 49.
E. (Quagga) mauritanicus dentitions from Ternifine, Algeria. (A) MNHN-F TER 154, P3–M3. (B) TER 1383, p2–m3. Occlusal views.

Figure 50.
E. (Quagga) cranium from Olorgesailie, KNM-OG 22833. (A) Ventral view, (B) Left lateral view. Modified from [12].
Figure 50.
E. (Quagga) cranium from Olorgesailie, KNM-OG 22833. (A) Ventral view, (B) Left lateral view. Modified from [12].

Figure 51.
Occlusal view of E. (Quagga) P2–M3 from Olorgesailie cranium, KNM-OG 22833. Modified from [12].
Figure 51.
Occlusal view of E. (Quagga) P2–M3 from Olorgesailie cranium, KNM-OG 22833. Modified from [12].

Figure 52.
E. (Hippotigris) zebra hartmannae skull, AM 7691. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 52.
E. (Hippotigris) zebra hartmannae skull, AM 7691. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 53.
E. (Hippotigris) zebra hartmannae dentitions. (A) Windhoek XIII.73, P2–M3. (B) AM 7381, p2–m3. Occlusal views.
Figure 53.
E. (Hippotigris) zebra hartmannae dentitions. (A) Windhoek XIII.73, P2–M3. (B) AM 7381, p2–m3. Occlusal views.

Figure 54.
(A) Magdalenian painting of Cueva de la Pasiega, Santander, Spain [86]. (B) Ivory plaquette, Paléo-élamite 1A = Suse IV A (−2700–2300), https://collections.louvre.fr/ark:/53355/cl010179457, accessed on 13 March 2020.
Figure 54.
(A) Magdalenian painting of Cueva de la Pasiega, Santander, Spain [86]. (B) Ivory plaquette, Paléo-élamite 1A = Suse IV A (−2700–2300), https://collections.louvre.fr/ark:/53355/cl010179457, accessed on 13 March 2020.

Figure 55.
Schematic occlusal views of (A) an upper premolar and (B) a lower premolar of a horse.

Figure 56.
E. (Equus) ferus cranium ZIN 521. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 56.
E. (Equus) ferus cranium ZIN 521. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 57.
Occlusal view of upper (A) and lower (B) cheek teeth of E. (Equus) ferus ZIN 521.

Figure 58.
E. (Equus) przewalskii cranium ZIN 5212. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 58.
E. (Equus) przewalskii cranium ZIN 5212. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 59.
Occlusal views of upper (A) and lower (B) cheek teeth of E. (Equus) przewalskii ZIN 5214.
Figure 59.
Occlusal views of upper (A) and lower (B) cheek teeth of E. (Equus) przewalskii ZIN 5214.

Figure 60.
E. (Equus) ferus cf. scotti cranium SI 160-455 from Ulakhan Sular, NE Siberia. (A) Ventral view, (B) Right lateral view, (C) Dorsal view.
Figure 60.
E. (Equus) ferus cf. scotti cranium SI 160-455 from Ulakhan Sular, NE Siberia. (A) Ventral view, (B) Right lateral view, (C) Dorsal view.

Figure 61.
Occlusal view of E. (Equus) ferus cf. scotti SI 160-455 upper cheek teeth.

Figure 62.
E. (Equus) ferus chosaricus cranium MGRI 113-165. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 62.
E. (Equus) ferus chosaricus cranium MGRI 113-165. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 63.
Occlusal views of upper cheek teeth of E. (Equus) ferus chosaricus. MGRI 113-174.


Figure 65.
Occlusal view of the upper cheek teeth of E. (Equus) ferus germanicus. (A) MB Ma 24672, Remagen, Germany. (B) MNHL 100.858, Siréjol, France.
Figure 65.
Occlusal view of the upper cheek teeth of E. (Equus) ferus germanicus. (A) MB Ma 24672, Remagen, Germany. (B) MNHL 100.858, Siréjol, France.

Figure 66.
E. (Equus) ferus germanicus cranium from Siréjol, France, MNHL 100-854. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 66.
E. (Equus) ferus germanicus cranium from Siréjol, France, MNHL 100-854. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 67.
E. (Equus) ferus antunesi type cranium SGP-FONT. 1. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 67.
E. (Equus) ferus antunesi type cranium SGP-FONT. 1. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 68.
E. (Sussemionus) coliemensis type cranium IA 1741. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.
Figure 68.
E. (Sussemionus) coliemensis type cranium IA 1741. (A) Ventral view; (B) Dorsal view; (C) Right lateral view.

Figure 69.
Simpson’s diagram of E. (Asinus) africanus and E. (Sussemionus) coliemensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 69.
Simpson’s diagram of E. (Asinus) africanus and E. (Sussemionus) coliemensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 70.
Occlusal views of upper and lower cheek teeth of E. (Sussemionus) coliemensis. (A) IA 1741; (B) IA 1721 modified from Lazarev 1980, Pl. VII-7 and Pl. VIII.
Figure 70.
Occlusal views of upper and lower cheek teeth of E. (Sussemionus) coliemensis. (A) IA 1741; (B) IA 1721 modified from Lazarev 1980, Pl. VII-7 and Pl. VIII.

Figure 71.
Occlusal views of (A) type PIN 835-123 p2–m3; (B) PIN 835-112 M1.

Figure 72.
Occlusal views of (A) PIN 3100-333 upper P3–M1; (B) lower M3 and M2 of E. (Sussemionus) verae.
Figure 72.
Occlusal views of (A) PIN 3100-333 upper P3–M1; (B) lower M3 and M2 of E. (Sussemionus) verae.

Figure 73.
Occlusal views of upper cheek teeth of E. (Sussemionus) suessenbornensis. (A) lectotype IQW 1964/1177 (S 514); (B) ([104], Plate VI, Figure 9); (C) IQW Süss. 7177, section of an M1 or M2; (D) IQW Süss. 638, right P3 or P4; (E) IQW Süss. 4219, right P3 or P4.

Figure 74.
Occlusal views of lower cheek teeth of E. (Sussemionus) suessenbornensis. (A) paralectotype p2–m3 QW1964/1303 (S 9280); (B) p2–m3 IQW 1964/1302 (S 9279); (C) m3 Halle E 23; (D) p2–m3 figured by Wüst ([103], Plate VII, Figure 1).

Figure 75.
Occlusal views of (A) upper left premolar IQW S 6657 and (B) lower left molar IQW S 7204 of E. (Sussemionus) cf. suessenbornensis.
Figure 75.
Occlusal views of (A) upper left premolar IQW S 6657 and (B) lower left molar IQW S 7204 of E. (Sussemionus) cf. suessenbornensis.

Figure 76.
Occlusal views of upper and lower cheek teeth of E. (Sussemionus) aff. suessenbornensis. (A) TB Akha 1279, P2–M3; (B) TB Akha 4, m3; (C) TB Akha 4, m1 or m2; (D) no number, p2: (E) no number, p2–m3.
Figure 76.
Occlusal views of upper and lower cheek teeth of E. (Sussemionus) aff. suessenbornensis. (A) TB Akha 1279, P2–M3; (B) TB Akha 4, m3; (C) TB Akha 4, m1 or m2; (D) no number, p2: (E) no number, p2–m3.

Figure 77.
Simpson’s diagrams of robust Sussemiones MC. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.
Figure 77.
Simpson’s diagrams of robust Sussemiones MC. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.

Figure 78.
Simpson’s diagrams of metapodial and upper cheek teeth dimensions in (A) E. hydruntinus and (B) Pseudohydruntines. MC: third metacarpal. MT; third metatarsal. 1: maximal length. 11: distal articular breadth. 4: depth at mid-diaphysis. P: upper P3 and P4. M: upper M1 and M2. size: occlusal (length + width)/2. prot.: protocone length. The numbers of specimens may be found in Tables S2 and S4.
Figure 78.
Simpson’s diagrams of metapodial and upper cheek teeth dimensions in (A) E. hydruntinus and (B) Pseudohydruntines. MC: third metacarpal. MT; third metatarsal. 1: maximal length. 11: distal articular breadth. 4: depth at mid-diaphysis. P: upper P3 and P4. M: upper M1 and M2. size: occlusal (length + width)/2. prot.: protocone length. The numbers of specimens may be found in Tables S2 and S4.

Figure 79.
Occlusal views of upper and lower cheek teeth referred to Hippotigris altidens by Reichenau, 1915. (A) IQW Süss. 4435, right M3; (B) IQW Süss. 4433, right M1; (C) ([106], Plate VII-24), right M1 or M2; (D) ([106] Plate VII-18), right P3 or P4; (E) ([106], Plate VII-13), left P3 or P4 and M3 IQW Süss. 4434; (F) IQW HG Süss 1371, left dP4; (G) IQW Süss. 4437, left m1 or m2; (H) ([106], Plate VI-7), right p3 or p4; (I): ([106], Plate VI-73) right p3 or p4.
Figure 79.
Occlusal views of upper and lower cheek teeth referred to Hippotigris altidens by Reichenau, 1915. (A) IQW Süss. 4435, right M3; (B) IQW Süss. 4433, right M1; (C) ([106], Plate VII-24), right M1 or M2; (D) ([106] Plate VII-18), right P3 or P4; (E) ([106], Plate VII-13), left P3 or P4 and M3 IQW Süss. 4434; (F) IQW HG Süss 1371, left dP4; (G) IQW Süss. 4437, left m1 or m2; (H) ([106], Plate VI-7), right p3 or p4; (I): ([106], Plate VI-73) right p3 or p4.

Figure 80.
Scatter diagram of P3–P4 and M1–M2 occlusal dimensions in E. granatensis, and specimens referred to E. altidens and E. marxi by Reichenau [105].
Figure 80.
Scatter diagram of P3–P4 and M1–M2 occlusal dimensions in E. granatensis, and specimens referred to E. altidens and E. marxi by Reichenau [105].

Figure 81.
E. granatensis upper cheek teeth from Venta Micena, Spain. (A) Occlusal view of VM H11, P3; (B) section at mid-height of VM K11 82-1038, P3 or P4; (C) section at mid-height of VM 83-C3-B5-1, P4; (D) Occlusal view of VM 87-C3, P2–M1; (E) Occlusal view of VM 82-1037, M3.
Figure 81.
E. granatensis upper cheek teeth from Venta Micena, Spain. (A) Occlusal view of VM H11, P3; (B) section at mid-height of VM K11 82-1038, P3 or P4; (C) section at mid-height of VM 83-C3-B5-1, P4; (D) Occlusal view of VM 87-C3, P2–M1; (E) Occlusal view of VM 82-1037, M3.

Figure 82.
Occlusal views of E. granatensis lower cheek teeth from Venta Micena, Spain. (A) VM 84-C3-J9-16, m3; (B) VM 84-C3-B9-12, p3; (C) VM 3576, p2; (D) VM 3654, p2–m3.
Figure 82.
Occlusal views of E. granatensis lower cheek teeth from Venta Micena, Spain. (A) VM 84-C3-J9-16, m3; (B) VM 84-C3-B9-12, p3; (C) VM 3576, p2; (D) VM 3654, p2–m3.

Figure 83.
Occlusal views of the upper and lower cheek teeth of E. aff. granatensis from Pirro, Italy. (A) IGF no collection number, left P2–M3 of a fragmentary cranium, type. (B) IGF, DSTF, P77.1, left p2–m3. Courtesy by M.T. Alberdi and M.R. Palombo.
Figure 83.
Occlusal views of the upper and lower cheek teeth of E. aff. granatensis from Pirro, Italy. (A) IGF no collection number, left P2–M3 of a fragmentary cranium, type. (B) IGF, DSTF, P77.1, left p2–m3. Courtesy by M.T. Alberdi and M.R. Palombo.

Figure 84.
Occlusal view of E. wuesti upper cheek teeth. (A) IQW 1999/26 560 (Mei. 26089), P3; (B) IQW 2010/31 393 (Mei. 30555), P3–P4; (C) the same in section.
Figure 84.
Occlusal view of E. wuesti upper cheek teeth. (A) IQW 1999/26 560 (Mei. 26089), P3; (B) IQW 2010/31 393 (Mei. 30555), P3–P4; (C) the same in section.

Figure 85.
E. wuesti, IQW 1998/26 401 (Mei. 25930), p3–m2, occlusal view.

Figure 86.
Occlusal views of upper and lower cheek teeth of E. cf. wuesti. (A) IQW S 3404, left P3 or P4; (B) IQW S 12719, left M1 or M2; (C) IQW S 482, left p3 or p4; (D) IQW S 677, right m1 or m2.
Figure 86.
Occlusal views of upper and lower cheek teeth of E. cf. wuesti. (A) IQW S 3404, left P3 or P4; (B) IQW S 12719, left M1 or M2; (C) IQW S 482, left p3 or p4; (D) IQW S 677, right m1 or m2.

Figure 87.
Occlusal views of E. hipparionoides, (A) type TB Akha 100, P4–M3; (B) TB Akha 99, p2–m2.

Figure 88.
Occlusal views of E. ovodovi lower cheek teeth. (A,B) Proskuriakova Cave N° 6 and N° 4, p4. (C) Proskuriakova Cave N° 6, p2. (D) Tsopi 58-1031, m2.
Figure 88.
Occlusal views of E. ovodovi lower cheek teeth. (A,B) Proskuriakova Cave N° 6 and N° 4, p4. (C) Proskuriakova Cave N° 6, p2. (D) Tsopi 58-1031, m2.

Figure 89.
E. vekuae. Dm53/59.3.B1gl.192, (A) Ventral view, (B) Left lateral view, (C) dorsal view, modified from Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D, The Dmanisi Equus: Systematics, biogeography, and paleoecology, Volume: 158,2021, Journal of Human Evolution.
Figure 89.
E. vekuae. Dm53/59.3.B1gl.192, (A) Ventral view, (B) Left lateral view, (C) dorsal view, modified from Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D, The Dmanisi Equus: Systematics, biogeography, and paleoecology, Volume: 158,2021, Journal of Human Evolution.

Figure 90.
E. vekuae. TB DMA 66, Ventral view.

Figure 91.
Simpson’s diagrams of Dmanisi Dm53/59.3.B1gl.192, Senèze NMB Se 336 and Gerakarou GER 8 crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. See Table S7 for Data.
Figure 91.
Simpson’s diagrams of Dmanisi Dm53/59.3.B1gl.192, Senèze NMB Se 336 and Gerakarou GER 8 crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. See Table S7 for Data.

Figure 92.
Simpson’s diagrams of crania from Anza Borrego (IVCM 2673) and Dmanisi (Dm53/59.3.B1gl.192). 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. Data in Table S7.
Figure 92.
Simpson’s diagrams of crania from Anza Borrego (IVCM 2673) and Dmanisi (Dm53/59.3.B1gl.192). 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. Data in Table S7.

Figure 93.
Occlusal views of upper cheek teeth of E. vekuae. (A) P3 or P4, DMA 1186. (B) section of M3, DMA 1089. (C) P2–M3, DMA 66. (D) P2–M3, Dm53/59.3.B1gl.192. (E) P3–M1, DMA 243-244-245. (F) M1–M2, DMA 1037-1156.
Figure 93.
Occlusal views of upper cheek teeth of E. vekuae. (A) P3 or P4, DMA 1186. (B) section of M3, DMA 1089. (C) P2–M3, DMA 66. (D) P2–M3, Dm53/59.3.B1gl.192. (E) P3–M1, DMA 243-244-245. (F) M1–M2, DMA 1037-1156.

Figure 94.
Scatter diagram of Protocone length versus Upper cheek size (Occlusal length + Occlusal width)/2.
Figure 94.
Scatter diagram of Protocone length versus Upper cheek size (Occlusal length + Occlusal width)/2.

Figure 95.
Occlusal views of lower cheek teeth from Dmanisi. (A) DMA 1127, p2–m1. (B,C) DMA 115, p2-p4 and p4–m3. (D) DMA 117, p3–m3. (E) DMA 4691, p2–m3, modified from Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D, The Dmanisi Equus: Systematics, biogeography, and paleoecology, Volume: 158,2021, Journal of Human Evolution. (F,G) DMA 1086, p3 or p4, occlusal view and section. (H) DMA 65, p2-p4. (I) DMA 1209, m1–m3.
Figure 95.
Occlusal views of lower cheek teeth from Dmanisi. (A) DMA 1127, p2–m1. (B,C) DMA 115, p2-p4 and p4–m3. (D) DMA 117, p3–m3. (E) DMA 4691, p2–m3, modified from Bernor, R.L.; Cirilli, O.; Buskianidze, M.; Lordkipanidze, D, The Dmanisi Equus: Systematics, biogeography, and paleoecology, Volume: 158,2021, Journal of Human Evolution. (F,G) DMA 1086, p3 or p4, occlusal view and section. (H) DMA 65, p2-p4. (I) DMA 1209, m1–m3.

Figure 96.
Simpson’s diagrams of Dmanisi metapodials. (A) Metacarpals. (B) Metatarsals. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.
Figure 96.
Simpson’s diagrams of Dmanisi metapodials. (A) Metacarpals. (B) Metatarsals. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.

Figure 97.
Simpson’s diagrams of M7 of (A) larger; (B) smaller. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 13 NY: Least depth of the lateral condyle. 14: Greatest depth of the medial condyle.
Figure 97.
Simpson’s diagrams of M7 of (A) larger; (B) smaller. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 13 NY: Least depth of the lateral condyle. 14: Greatest depth of the medial condyle.

Figure 98.
First phalanges of E. vekuae. (A–E) dorsal views of anterior phalanges DMA 19, 268 and of posterior phalanges DMA 129, 201 and 647. (F–J) plantar views of the same.
Figure 98.
First phalanges of E. vekuae. (A–E) dorsal views of anterior phalanges DMA 19, 268 and of posterior phalanges DMA 129, 201 and 647. (F–J) plantar views of the same.

Figure 99.
Simpson’s diagrams of MC of (A) E. vekuae; (B) E. granatensis. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.
Figure 99.
Simpson’s diagrams of MC of (A) E. vekuae; (B) E. granatensis. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle.

Figure 100.
Occlusal views of E. marxi types. (A) IQW HG Süss. 1365, left P3 or P4; (B) IQW HG Süss. 1366 left M1 or M2; (C) IQW HG Süss. 1367 right M1 or M2; (D) IQW HG Süss. 1368, left p3; (E) IQW HG Süss. 1368, left p4; (F) IQW HG Süss. 1370, left m1 or m2.
Figure 100.
Occlusal views of E. marxi types. (A) IQW HG Süss. 1365, left P3 or P4; (B) IQW HG Süss. 1366 left M1 or M2; (C) IQW HG Süss. 1367 right M1 or M2; (D) IQW HG Süss. 1368, left p3; (E) IQW HG Süss. 1368, left p4; (F) IQW HG Süss. 1370, left m1 or m2.

Figure 101.
Ventral view of the palate MNHN-F Han 770 type of E. tabeti, Aïn Hanech, Algeria.

Figure 102.
Occlusal views of (A) P2–M1 MNHN-F Han 1229; (B) p2–m3 MNHN-F Han 530 (courtesy by N. Aouadi, [127] of E. tabeti, Aïn Hanech, Algeria (C) M1–M3 MNHN-F Boc 46-48 of E. numidicus from Aïn Boucherit, Algeria.
Figure 102.
Occlusal views of (A) P2–M1 MNHN-F Han 1229; (B) p2–m3 MNHN-F Han 530 (courtesy by N. Aouadi, [127] of E. tabeti, Aïn Hanech, Algeria (C) M1–M3 MNHN-F Boc 46-48 of E. numidicus from Aïn Boucherit, Algeria.

Figure 103.
Simpson’s diagrams of (A) MC and (B) MT of E. tabeti, E. africanus, and E. melkiensis. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.
Figure 103.
Simpson’s diagrams of (A) MC and (B) MT of E. tabeti, E. africanus, and E. melkiensis. 1: Maximal length. 3: Breadth at the middle of the diaphysis. 4: Depth of the diaphysis at the same level. 5: Proximal breadth. 6: Proximal depth. 10: Distal supra-articular breadth. 11: Distal articular breadth. 12: Depth of the sagittal crest. 13: Least depth of the medial condyle. 14: Greatest depth of the medial condyle. n: number of specimens.

Figure 104.
E. nalaikhaensis, PIN 3747-500 type cranium, (A) Dorsal view; (B) Right lateral view. (C) Ventral view.
Figure 104.
E. nalaikhaensis, PIN 3747-500 type cranium, (A) Dorsal view; (B) Right lateral view. (C) Ventral view.

Figure 105.
Simpson’s diagrams of E. apolloniensis and E. nalaikhaensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 105.
Simpson’s diagrams of E. apolloniensis and E. nalaikhaensis crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 106.
Occlusal views of the right associated P2–M3 (PIN 3747-522, 600, 358, 402, 362, 395) from Nalaikha.
Figure 106.
Occlusal views of the right associated P2–M3 (PIN 3747-522, 600, 358, 402, 362, 395) from Nalaikha.

Figure 107.
Kenyan Gregory Konso fossils, KGA 8-374. Cranium, (A) Ventral view; (B) Left lateral view. (C) Dorsal view. Photographs kindly communicated to me in 2003 by Prof. H. Nakaya.
Figure 107.
Kenyan Gregory Konso fossils, KGA 8-374. Cranium, (A) Ventral view; (B) Left lateral view. (C) Dorsal view. Photographs kindly communicated to me in 2003 by Prof. H. Nakaya.

Figure 108.
Simpson’s diagrams of KGA 8-374 and E. (Asinus) africanus crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.
Figure 108.
Simpson’s diagrams of KGA 8-374 and E. (Asinus) africanus crania. 16: Breadth of the supra-occipital crest. 23: Anterior ocular line. 3: Vomerine length. 4: Post-vomerine length. 2-5: Palatal length sensu stricto. 5: Muzzle length. 17: Muzzle breadth at the posterior borders of I3. 17bis: Least muzzle breadth (between the crests). 13: Frontal breadth. 10: Greatest choanal breadth. 25: Facial height in front of P2. 28: Cranial height. 9: Length of choanae. 20: Height of the external auditive meatus. 31: Length of the naso-incisival notch. 32: Cheek length. n: number of measurements.

Figure 109.
Occlusal views of upper cheek teeth from Konso and Ubeidiyeh. (A) KGA 8-374; (B) HUJ-ESE U II, 23-24 (195), P2; (C): HUJ-ESE U II, 36 (203), P3 or P4.
Figure 109.
Occlusal views of upper cheek teeth from Konso and Ubeidiyeh. (A) KGA 8-374; (B) HUJ-ESE U II, 23-24 (195), P2; (C): HUJ-ESE U II, 36 (203), P3 or P4.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Eisenmann, V. Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms. Quaternary 2022, 5, 38. https://doi.org/10.3390/quat5030038
AMA Style
Eisenmann V. Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms. Quaternary. 2022; 5(3):38. https://doi.org/10.3390/quat5030038
Chicago/Turabian StyleEisenmann, Vera. 2022. "Old World Fossil Equus (Perissodactyla, Mammalia), Extant Wild Relatives and Incertae Sedis Forms" Quaternary 5, no. 3: 38. https://doi.org/10.3390/quat5030038