
Trypanorhynch Assemblages Indicate Ecological and Phylogenetical Attributes of Their Elasmobranch Final Hosts



1
Aquaculture and Sea-Ranching, Agricultural and Environmental Sciences, University of Rostock, Justus-von-Liebig Weg 6, 18059 Rostock, Germany
2
Marine Fisheries Department, Faculty of Fisheries and Marine Sciences, Bogor Agricultural University (IPB), Jl. Raya Darmaga Kampus IPB Darmaga, 16680 Bogor, Indonesia
3
GEOMAR, Helmholtz Centre for Ocean Research Kiel, Düsternbrooker Weg 20, 24105 Kiel, Germany
*
Author to whom correspondence should be addressed.
Fishes 2017, 2(2), 8; https://doi.org/10.3390/fishes2020008
Submission received: 7 February 2017
/
Revised: 16 May 2017
/
Accepted: 12 June 2017
/
Published: 17 June 2017
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:This study explores non-metric multidimensional scaling (nMDS) as a tool for investigating parasites as indicators of the elasmobranch biology. An attractive feature of nMDS is its ability to allow assemblage-level parasite data to be simultaneously applied to questions of host biology. This method was examined using the tapeworm order Trypanorhyncha Diesing, 1863, which is known to be transmitted among their hosts through the marine food web (via predation), can unambiguously be identified in the intermediate and final hosts, and has the potential as an indicator of the host feeding biology. Our analyses focused on trypanorhynch assemblages in elasmobranchs as definitive hosts. The relationships between trypanorhynch assemblages and the depth, feeding ecology, habitat, and phylogeny for all sharks were complex, but we found that depth distribution, diet composition and habitat type were the major influencing factors. Several species of sharks showed different characters than known from their descriptions that could be attributed to the change of shark behavior or the trypanorhynch host path. The relationship between the trypanorhynch assemblage and factors for carcharhiniform species alone was more robust than for all sharks. In the carcharhiniform analysis, the relationship between habitat type and trypanorhynch assemblage was most remarkable. Overlapping host ecology was evident even in phylogenetically-distant related hosts.
1. Introduction
Elasmobranchs are difficult to observe in nature and information on rare or less-frequently caught species is scarce. This especially concerns aspects of their ecology, main habitat, depth range, and most important prey items. Their prey is rapidly digested and affiliation, e.g., to mainly fish or invertebrate-feeding activities, is difficult. Fish parasites have been widely used as biological indicators for the host ecology, including their zoogeographical distribution, migration, and feeding behavior [1]. However, the methods applied for the elasmobranchs are, so far, limited due to often restricted and unpredictable catches and less availability of specimens to study.
The cestode order Trypanorhyncha Diesing, 1863, is considered to be a basal tapeworm group of elasmobranchs. Its members can be characterized by a scolex with a tentacular apparatus that can be used for classification [2,3,4,5]. The most recent revision of the trypanorhynchs subdivided the order into five different superfamilies and 15 families [6]. The first large-scale molecular phylogenetic analyses of this group included trypanorhynch species belonging to all superfamilies and 12 of the 15 recognized families [7,8]. The analysis of complete simple sequence repeats DNA (ssrDNA) and partial long sequence repeat DNA (lsrDNA) (D1–D3) revealed high resolution and the presence of a basal split, representing the shark- and ray-inhabiting lineages as new suborders. These results gave, in part, support for the trypanorhynch classification by Palm [6] as well as more insight into the intrafamily phylogenetic relationships within the Trypanorhyncha. The host-parasite-checklist of these cestodes indicated that some of the superfamilies and families seem to have host preferences [6], while the patterns of others can be explained by host switches, even between rajiform and selachiform elasmobranchs [7].
The ~300 described species of the order Trypanorhyncha so far exhibit the following general life-cycle pattern [6,9,10,11]. The first intermediate host becomes infected when it consumes an oncosphere or free-swimming coracidium larva. The first parasitic larval stage is a procercoid; the limited available data suggest that this stage parasitizes copepods. Second intermediate hosts include a wide array of marine animals, but invertebrates and teleosts figure particularly prominent at this level. Unlike most groups of tapeworms, the trypanorhynch final larval stage, though having a different morphology (plerocercoid, plerocercus, merocercoid), is generally identifiable to species because it exhibits the adult hook pattern, a pattern on which much of the taxonomy of the group is based [4,5,6]. Some trypanorhynchs use a paratenic host following the final intermediate host; this host serves to bridge food-web gaps. As adults, trypanorhynchs parasitize the spiral intestine of sharks and rays (elasmobranchs). Fortunately, it is common for individual elasmobranch species to host multiple trypanorhynch species and, thus, assemblage-level data are readily available for many elasmobranchs that serve as definitive hosts.
Our study uses non-metric multidimensional scaling (nMDS) as a tool for investigating parasites as indicators for selected ecological attributes and the phylogeny of their elasmobranch final hosts. A particularly novel and attractive feature of nMDS is the possibility to allow assemblage-level parasite data to be simultaneously applied to questions of host biology. The method was examined using trypanorhynch tapeworm assemblages, which have obvious potential as indicators of host feeding biology in the elasmobranch. The complex life-cycles of these parasites, each of which involves a minimum of three different hosts and three distinct life-cycle stages that vary in their degree of host specificity [11] also suggest trypanorhynchs as appropriate indicators of other aspects of their host biology.
Among elasmobranchs, sharks (Selachii) were specifically chosen because they vary substantially in diet and habitat, which allows a wide spectrum of aspects of host biology to be explored. Within the Selachii, more detailed analyses were conducted on sharks of the order Carcharhiniformes (ground sharks). Comprehensive trypanorhynch data were available for this shark order, and provided a unique opportunity to perform nMDS analyses on a more precise taxonomic level. Factors concerning the shark biology included (1) taxonomy (order for the Selachii analyses, or family for the Carcharhiniform analyses); (2) habitat (benthic vs. pelagic); (3) depth distribution (above or below 200 m depth); and (4) diet (primarily invertebrates vs. primarily teleosts). Trypanorhynch assemblage dependence on these four factors was assessed by superimposing the different variables into the two-dimensional ordination plots resulting from nMDS analysis of the hierarchical agglomerative classification of shark species generated from their trypanorhynch assemblages. Trypanorhynch assemblages were considered to be appropriate as indicators of the biological factors that were found to be significantly correlated with the agglomerative classification.
2. Results
2.1. All Sharks
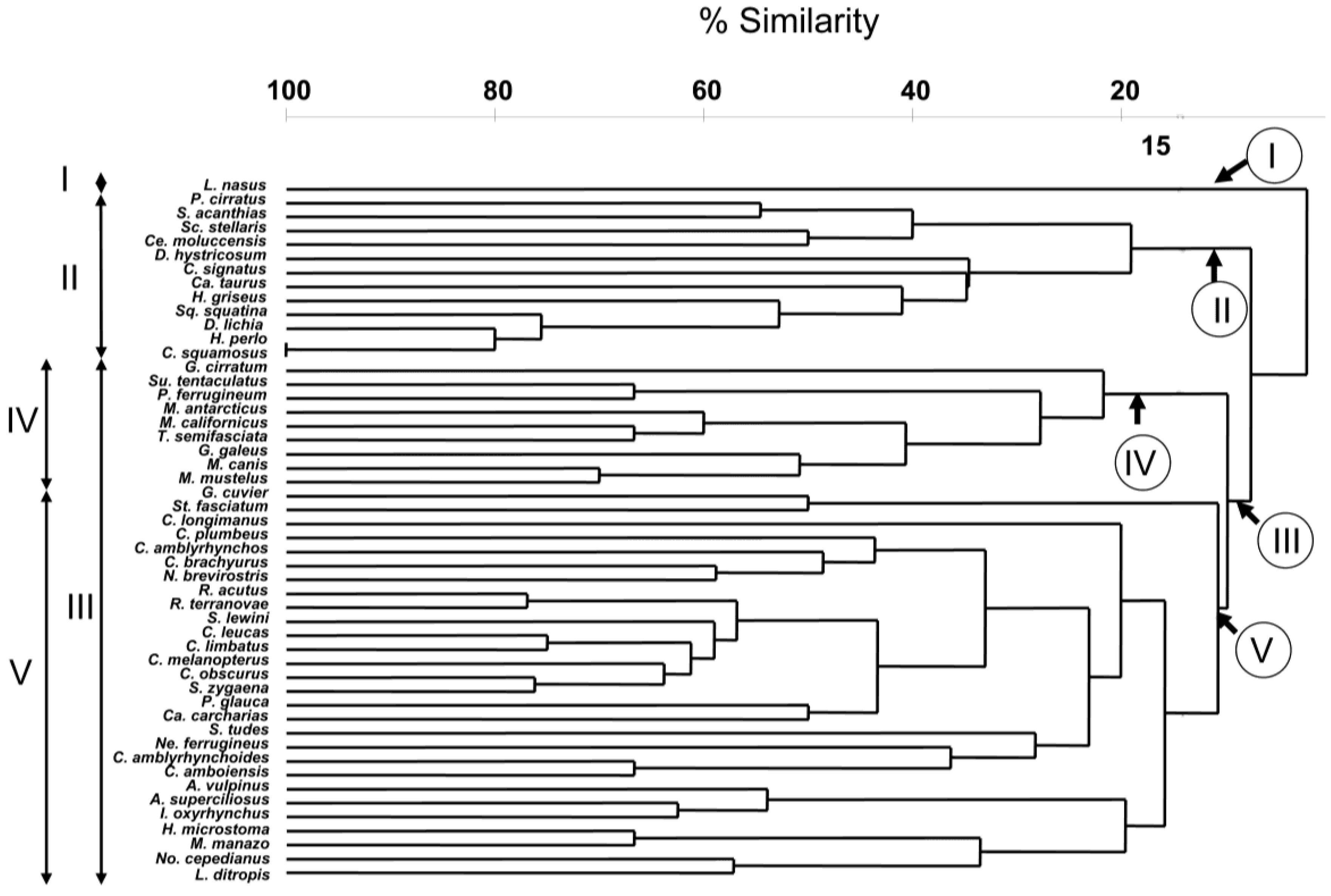
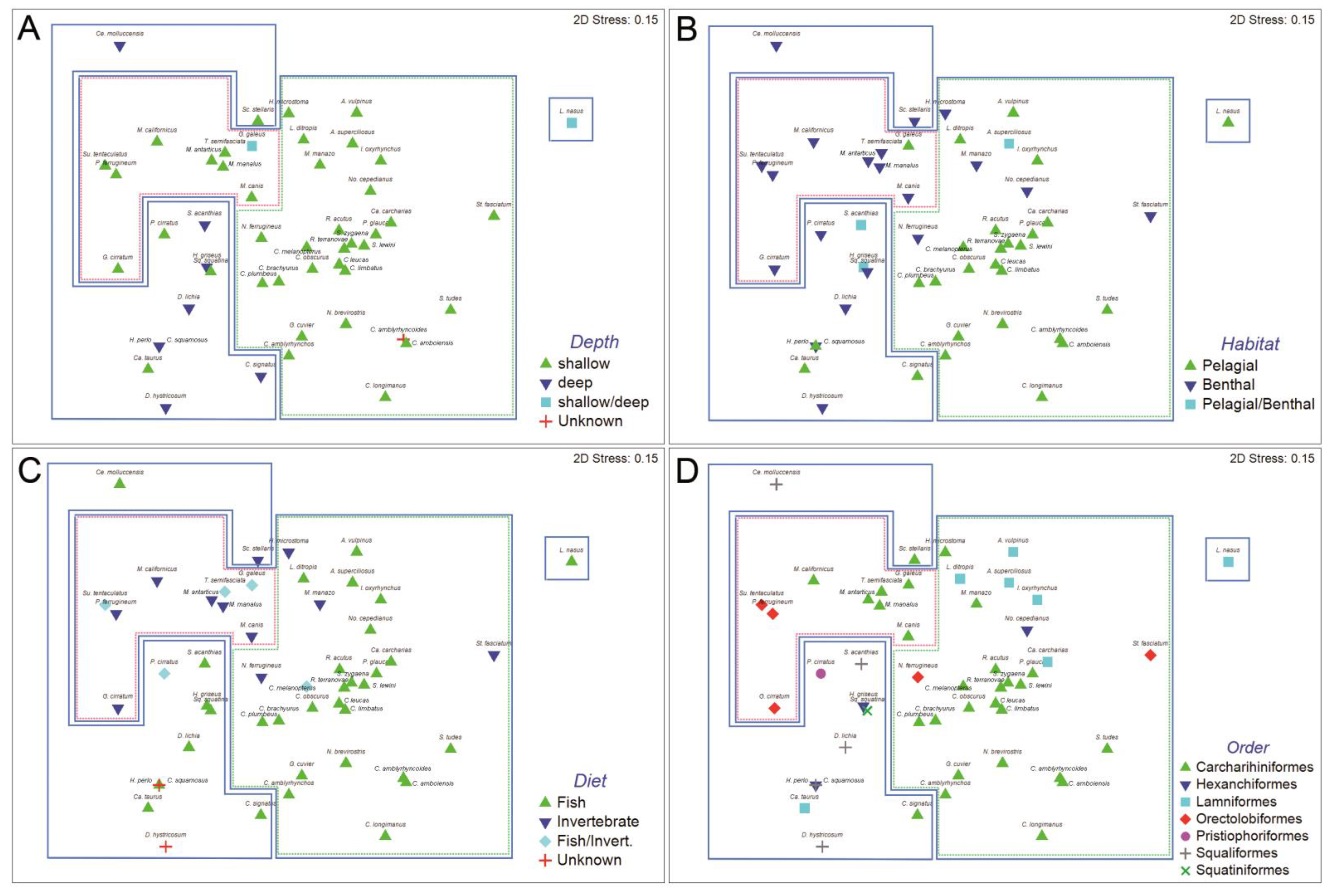
The result of the hierarchical, agglomerative classification (CLUSTER) of trypanorhynch assemblages of 50 species of Selachii revealed five major clusters of shark species at an arbitrary dissimilarity level of 78% (Figure 1). The two-dimensional ordination plots resulting from nMDS of this agglomerative classification, with sharks coded for taxonomic order, diet, habitat, and depth distribution are shown in Figure 2. The analysis of similarities (ANOSIM) revealed significant differences in trypanorhynch assemblage composition between sharks that primarily consume invertebrates versus those that primarily consume teleosts (global R = 0.279; p < 0.01), between those from benthic and pelagic habitats (global R = 0.275; p < 0.01), and between sharks occurring in deep versus shallow water (global R = 0.327; p < 0.01), thus supporting the significance of the major clusters resulting from the classification and ordination methods. With respect to taxonomy, the ANOSIM showed significant differences among shark orders (global R = 0.381; p < 0.01), suggesting a relationship between trypanorhynch genera and shark orders. In ANOSIM pairwise comparison, the trypanorhych assemblages of the Carcharhiniformes were significantly different from those of the Squaliformes (global R = 0.546; p < 0.01), the Orectolobiformes (global R = 0.466; p < 0.1), the Lamniformes (global R = 0.312; p < 0.01), and the Hexanchiformes (global R = 0.275; p < 0.05). In addition, the trypanorhynch assemblages of the Lamniformes were significantly different from those of the Orectolobiformes (global R = 0.432; p < 0.01) and Squaliformes (global R = 0.355; p < 0.05), and the assemblages of the Orectolobiforms were significantly different from those of the Squaliformes (global R = 0.620; p < 0.01). No significant differences were found between the trypanorhynch assemblages of any of the other 16 pairs of shark orders.
The following similarity percentage (SIMPER) analysis indicated which trypanorhynch genera contributed most to the differences between the studied shark species (Table S2). The average similarity was 22.29% for the pelagic sharks, with the genera Nybelinia Poche, 1926, Otobothrium Linton, 1891, and Dasyrhynchus Pintner, 1928, accounting for a cumulative contribution of 54.30%. Including Heteronybelinia Palm, 1999, Callitetrarhynchus Pintner, 1931, Tentacularia Bosc, 1797, Grillotia Guiart, 1927, and Floriceps Cuvier 1817, the trypanorhynch genera reached a cumulative contribution of 91.51%. The benthic sharks which had an average similarity of 15.64% were characterised by five genera, Dollfusiella Campbell and Beveridge, 1994, Nybelinia, Grillotia, Lacistorhynchus Pintner, 1913, and Gilquinia Guiart 1927, which accounted for a cumulative contribution of 92.84%. The genera Nybelinia (12.48%), Otobothrium (8.26%), Dasyrhynchus (6.85%), Heteronybelinia (6.44%), and Callitetrarhynchus (5.07%) for the pelagic sharks and Dollfusiella (7.81%) and Grillotia (7.93%) for the benthic sharks contributed over 50% to the total dissimilarity. Other characteristic trypanorhynch genera of the benthic sharks were Gilquinia, Prochristianella Dollfus, 1946, Diesingium Pintner, 1929, Deanicola Beveridge, 1990 and Pseudolacistorhynchus Palm, 1995. The parasite genera whose species numbers significantly differed between pelagic and benthic sharks were Tentacularia (U = 171.0, p = 0.003), Nybelinia (U = 177.5, p = 0.025), Heteronybelinia (U = 193.0, p = 0.029), Pseudogrillotia Dollfus, 1969 (U = 216.0, p = 0.033), Dasyrhynchus (U = 144.0, p = 0.000), Lacistorhynchus (U = 195.0, p = 0.013), Pseudolacistorhynchus (U = 232.0, p = 0.034), Callitetrarhynchus (U = 180.0, p = 0.013), Floriceps (U = 180.0, p = 0.005), Poecilancistrum Dollfus, 1929 (U = 207.0, p = 0.020), Otobothrium (U = 155.0, p = 0.002), Tetrarhynchobothrium Diesing, 1854 (U = 217.5, p = 0.012) and Dollfusiella (U = 156.5, p = 0.001).
The average similarity and genus composition of trypanorhynchs for fish vs. invertebrate feeding sharks is compiled in Table S3. Fish-feeding sharks had a 19.38% average similarity, with the trypanorhynch genera Nybelinia (30.43%), Grillotia (14.98%), Heteronybelinia (10.53%), Otobothrium (9.89%), Dasyrhynchus (9.72%), Callitetrarhynchus (6.96%), Tentacularia (4.57%) and Floriceps (3.29%) having a contribution of 90.37%. Invertebrate-feeding sharks had an average similarity of 18.76%, and were characterized by the genera Dollfusiella (40.53%), Nybelinia (30.13%), Lacistorhynchus (8.27%), Prochristianella (4.29%), Diesingium, Trigonolobium Dollfus, 1929 and Tetrarhynchobothrium (together 9.66%). The decreasing contribution to the total similarity, up to 100%, is given in Table S3. The parasite genera whose species numbers significantly differed between fish and invertebrate-feeding sharks were Tentacularia (U = 126.5, p = 0.025), Heteronybelinia (U = 124.0, p = 0.036), Kotorella Euzet and Radujkovic, 1989 (U = 160.0, p = 0.044), Dasyrhynchus (U = 110.0, p = 0.009), Grillotia (U = 129.0, p = 0.048), Paragrillotia Dollfus, 1969 (U = 160.0, p = 0.044), Lacistorhynchus (U = 123.0, p = 0.007), Pseudolacistorhynchus (U = 144.0, p = 0.007), Diesingium (U = 128.0, p = 0.001), Floriceps (U = 132.0, p = 0.035), Otobothrium (U = 123.0, p = 0.034), Tetrarhynchobothrium (U = 128.0, p = 0.001), Zygorhynchus Beveridge & Campbell, 1988 (U = 160.0, p = 0.044), Eutetrarhynchus Pintner, 1913 (U = 149.5, p = 0.047), Dollfusiella (U = 74.5, p = 0.000), Trigonolobium (U = 144.0, p = 0.007), Trimacracanthus Beveridge and Campbell, 1987 (U = 160, p = 0.044) and Prochristianella (U = 133.0, p = 0.008).
The average similarity and genus composition of trypanorhynchs for fish vs. invertebrate feeding sharks is compiled in Table S3. Fish-feeding sharks had a 19.38% average similarity, with the trypanorhynch genera Nybelinia (30.43%), Grillotia (14.98%), Heteronybelinia (10.53%), Otobothrium (9.89%), Dasyrhynchus (9.72%), Callitetrarhynchus (6.96%), Tentacularia (4.57%) and Floriceps (3.29%) having a contribution of 90.37%. Invertebrate-feeding sharks had an average similarity of 18.76%, and were characterized by the genera Dollfusiella (40.53%), Nybelinia (30.13%), Lacistorhynchus (8.27%), Prochristianella (4.29%), Diesingium, Trigonolobium Dollfus, 1929 and Tetrarhynchobothrium (together 9.66%). The decreasing contribution to the total similarity, up to 100%, is given in Table S3. The parasite genera whose species numbers significantly differed between fish and invertebrate-feeding sharks were Tentacularia (U = 126.5, p = 0.025), Heteronybelinia (U = 124.0, p = 0.036), Kotorella Euzet and Radujkovic, 1989 (U = 160.0, p = 0.044), Dasyrhynchus (U = 110.0, p = 0.009), Grillotia (U = 129.0, p = 0.048), Paragrillotia Dollfus, 1969 (U = 160.0, p = 0.044), Lacistorhynchus (U = 123.0, p = 0.007), Pseudolacistorhynchus (U = 144.0, p = 0.007), Diesingium (U = 128.0, p = 0.001), Floriceps (U = 132.0, p = 0.035), Otobothrium (U = 123.0, p = 0.034), Tetrarhynchobothrium (U = 128.0, p = 0.001), Zygorhynchus Beveridge & Campbell, 1988 (U = 160.0, p = 0.044), Eutetrarhynchus Pintner, 1913 (U = 149.5, p = 0.047), Dollfusiella (U = 74.5, p = 0.000), Trigonolobium (U = 144.0, p = 0.007), Trimacracanthus Beveridge and Campbell, 1987 (U = 160, p = 0.044) and Prochristianella (U = 133.0, p = 0.008).
Only three trypanorhynch genera, mainly Grillotia with a 94.25% total similarity contribution, Gilquinia (4.24%), and Nybelinia (1.51%) characterised the deep water sharks (Table S4). The other trypanorhynch genera were characteristic of shallow water shark species occurring above 200 m depth. Nybelinia (39.30%), Otobothrium (10.10%) and Dollfusiella (9.60%) contributed most, followed by Dasyrhynchus (6.23%), Callitetrarhynchus (6.15%), Heteronybelinia (5.07%), Grillotia (3.99%), and Tentacularia, Mixonybelinia Palm, 1999, and Lacistorhynchus (together 10.20%) (Table S4). The parasite genera whose species numbers significantly differed between shallow and deep water sharks were Nybelinia (U = 85.0, p = 0.016), Chimaerarhynchus Beveridge and Campbell, 1989 (U = 136.5, p = 0.014), Dasyrhynchus (U = 108.0, p = 0.038), Deanicola (U = 136.5, p = 0.014), Grillotia (U = 34.5, p = 0.000), Callitetrarhynchus (U = 104.0, p = 0.030), Dollfusiella (U = 108.0, p = 0.037), and Otobothrium (U = 100.0, p = 0.025).
Similarity percentage analysis further revealed the importance of the different trypanorhynch genera for the characterisation of the shark orders (Table S5). The Carcharhiniformes were characterized by Nybelinia (33.03%), Otobothrium (13.43%), Dasyrhynchus (10.14%), Callitetrarhynchus (9.98%), Heteronybelinia (8.83%), Dollfusiella (4.90%), Tentacularia (4.67%), Floriceps (4.39%), and Lacistorhynchus (2.88%). The genera Grillotia and Nybelinia contributed 73.33% and 26.67% to the similarity of the Hexanchiformes, while the Lamniforms were characterised by Nybelinia (35.68%), Sphyriocephalus Pintner, 1913 (20.43%), Mixonybelinia (19.68%), Gymnorhynchus Rudolphi, 1819 (11.09%), and Molicola Dollfus 1935 (9.24%). The genera Dollfusiella (83.56%) and Mixonybelinia (9.86%) contributed most to the similarity of the Orectolobiformes, and Grillotia (86.11%) and Gilquinia (13.89%) to that of the Squaliformes (Table S5).
The genera Grillotia (15.18%), Nybelinia (11.69%), Gilquinia (9.24%), Otobothrium (7.92%), and Dasyrhynchus (6.19%) contributed over 50% to the total dissimilarity of Carcharhiniformes and Squaliformes, and Dollfusiella (12.66%), and Pseudolacistorhynchus (4.78%) were important to separate the Orectolobiformes from the Carcharhiniformes. The genera Nybelinia (13.36%), Sphyriocephalus (9.15%), Otobothrium (7.98%), Mixonybelinia (6.44%), Dasyrhynchus (6.32%), and Gymnorhynchus (5.45%) contributed over 50% to the dissimilarity of the Carcharhiniformes and the Lamniformes, while the Lamniformes differed from the Orectolobiformes by Dollfusiella (15.4%), Nybelinia (14.18%), Sphyriocephalus (10.78%), Mixonybelinia (8.21%), and Pseudolacistorhynchus (5.25%), and differed from the Squaliformes by Grillotia (18.81%), Nybelinia (14.46%), Sphyriocephalus (12.40%), and Gilquinia (11.00%). The genera Grillotia (25.87%), Nybelinia (12.26%), and Otobothrium (8.04%) contributed over 50% to the dissimilarity of the Carcharhiniformes and the Hexanchiformes. Other orectolobiform characteristic genera contributed less than 5% (Hornelliella Yamaguti, 1954, Molicola, Hepatoxylon Bosc, 1811). Only four genera, Grillotia (20.90%), Dollfusiella (16.76%), Gilquinia (11.88%) and Deanicola (6.73%) contributed 58.02% to the dissimilarity between the Orectolobiformes and the Squaliformes. The parasite genera where the species numbers significantly differed in seven orders of shark were Nybelinia (H = 11.0, p = 0.044), Mixonybelinia (H = 13.8, p = 0.016), Sphyriocephalus (H = 15.9, p = 0.007), Gilquinia (H = 13.2, p = 0.020), Molicola (H = 13.3, p = 0.019), Gymnorhynchus (H = 13.5, p = 0.017), Grillotia (H =19.7, p = 0.002), Pseudolacistorhynchus (H = 18.4, p = 0.002), Callitetrarhynchus (H = 11.2, p = 0.004), Otobothrium (H = 11.1, p = 0.043), and Dollfusiella (H = 13.5, p = 0.018).
2.2. Carcharhiniformes
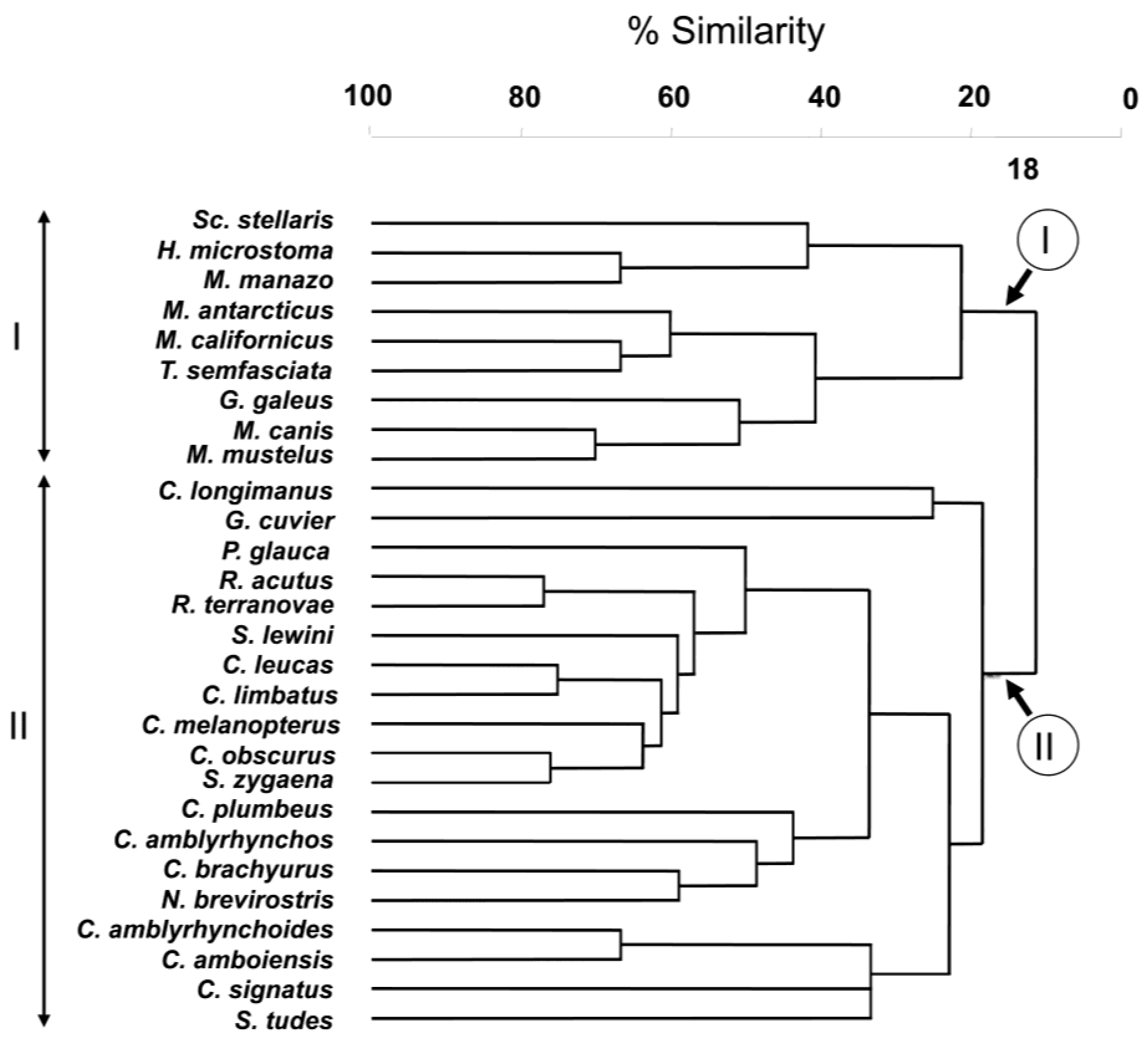
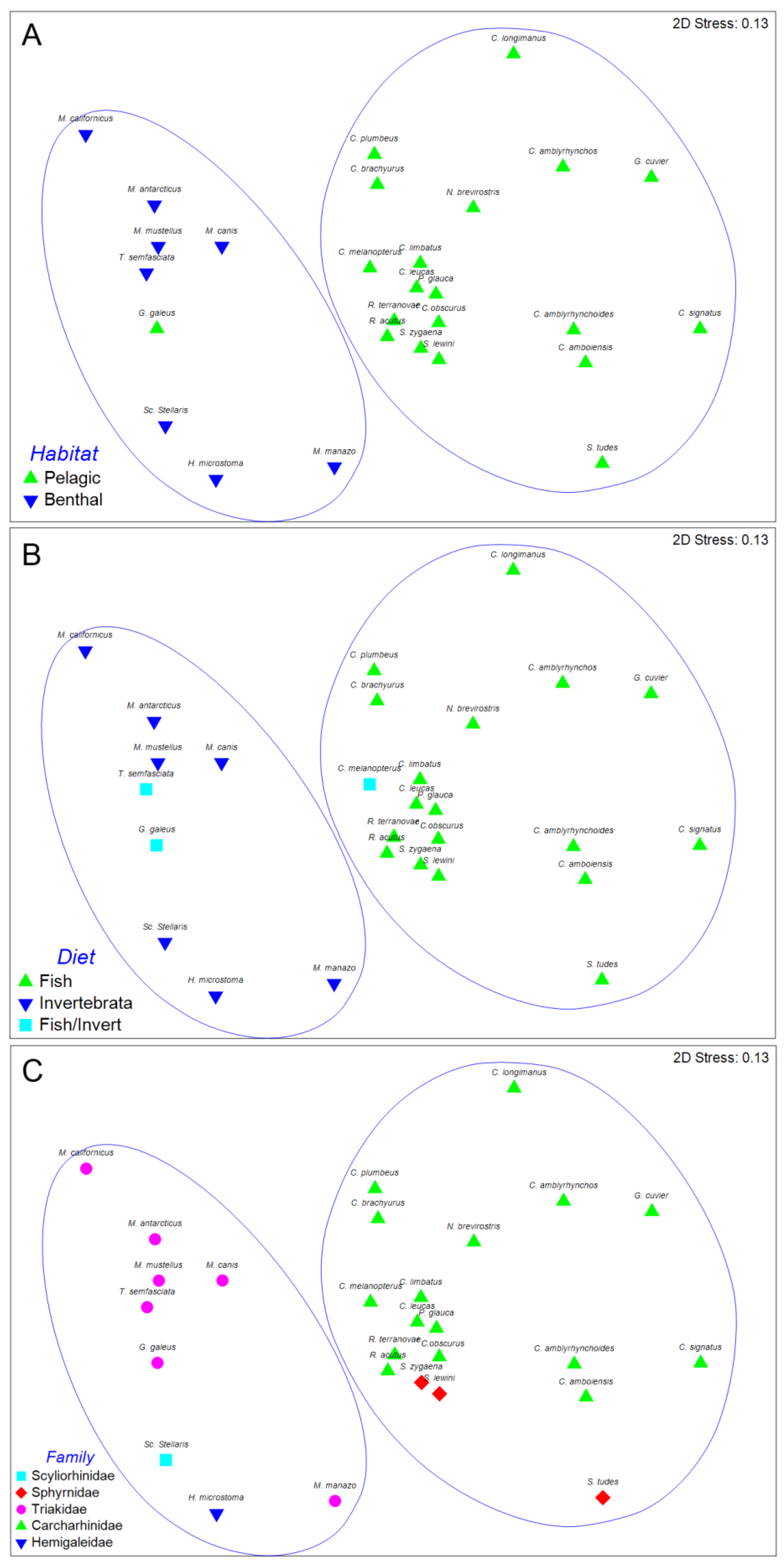
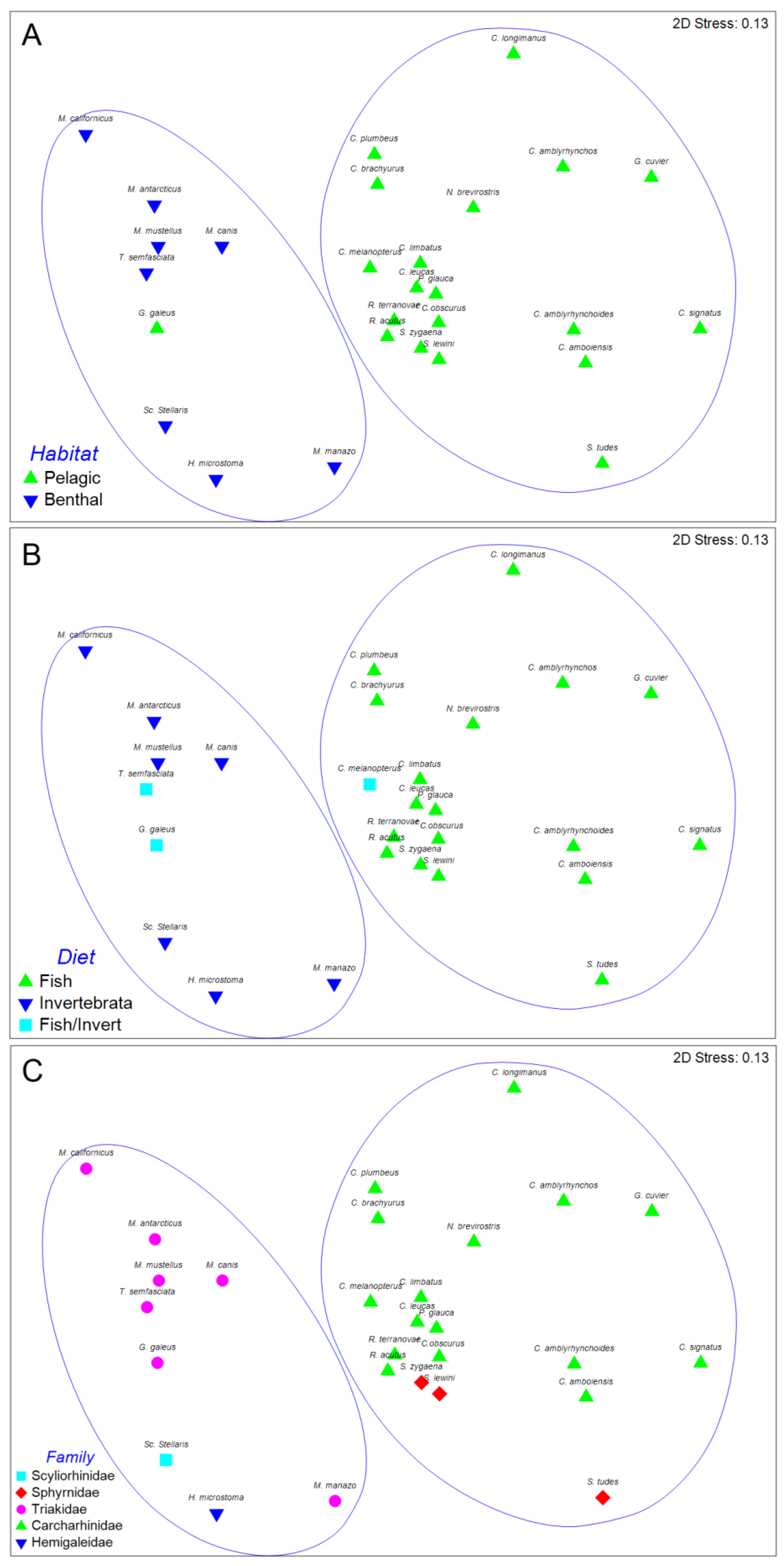
The result of the hierarchical, agglomerative classification (CLUSTER) of trypanorhynch assemblages of Carcharhiniformes, the order with the most comprehensive trypanorhynch dataset in Palm [6], is shown in Figure 3. Two major clusters of sharks at an arbitrary dissimilarity level of 82% were recovered (I and II). The nMDS plots of the classification, with sharks coded for family, diet, and habitat are shown in Figure 4. Analysis of similarities revealed significant differences in trypanorhynch assemblage composition between sharks that primarily consume invertebrates versus those that primarily consume teleosts (global R = 0.683; p < 0.01), and also between those from benthic and pelagic habitats (global R = 0.590; p < 0.01). Regarding taxonomy, however, ANOSIM pairwise comparisons, detected significant differences on trypanorhynch assemblages only between Carcharhinidae and Triakidae (global R = 0.678; p < 0.01) and between Sphyrnidae and Triakidae (global R = 0.748; p < 0.01). No significant differences were found between the trypanorhynch assemblages of any of the other eight pairs of carcharhiniform families.
The results of the SIMPER analyses to explore the contributions of individual trypanorhynch genera to the degree of similarity observed among the conditions for the factors environment and diet are shown in Tables S6 and S7, respectively. The average similarity was 31.16% for pelagic carcharhiniforms, with the genera Nybelinia, Otobothrium and Dasyrhynchus having a cumulative contribution of 52.53%. Together with Heteronybelinia, Callitetrarhynchus, Tentacularia, and Floriceps, these trypanorhynch genera reached a cumulative contribution of 92.37%. Benthic carcharhiniforms that had an average similarity of 33.88% were characterized by four genera, Dollfusiella, Lacistorhynchus, Nybelinia, and Diesingium, having a cumulative contribution of 91.22%. Parasite genera whose species numbers significantly differed between pelagic and benthic carcharhiniforms were Tentacularia (U = 39.0, p = 0.031 one-tailed), Heteronybelinia (U = 33.0, p = 0.017), Pintneriella Yamaguti 1934 (U = 55.0, p = 0.028), Dasyrhynchus (U = 30.0, p = 0.012), Lacistorhynchus (U = 16.5, p = 0.000), Diesingium (U = 35.5, p = 0.002), Floriceps (U = 39.0, p = 0.031), Otobothrium (U = 37.5, p = 0.040), Tetrarhynchobothrium (U = 44.0, p = 0.002), Eutetrarhynchus (U = 47.0, p = 0.024), Dollfusiella (U = 19.0, p = 0.000), and Trimacracanthus (U = 55.0, p = 0.028).
Carcharhiniforms with different diet preferences, either fish or invertebrates, had an average similarity of 32.44 for fish and 29.47 for invertebrates. Similarly to the SIMPER analysis of benthic versus pelagic carcharhiniforms, Nybelinia, Otobothrium, and Dasyrhynchus reached a cumulative contribution close to 50% (48.95%) for preliminary fish eating sharks (Table S7). Together with Heteronybelinia, Callitetrarhynchus, Tentacularia, and Floriceps, the cumulative contribution was 91.43%. Invertebrate-feeding carcharhiniforms were characterized by the genus Nybelinia, that had a contribution of 50.24%, and together with Dollfusiella, Lacistorhynchus, Diesingium, and Trigonolobium reached a cumulative contribution of 91.92%. The decreasing contribution to the total similarity, up to 100%, is given in Table S7. The parasite genera whose species numbers were significantly different between fish and invertebrate feeders were Tentacularia (U = 35.0, p = 0.018), Heteronybelinia (U = 24.5, p = 0.004), Gilquinia (U = 45, p = 0.010), Dasyrhynchus (U = 24.5, p = 0.005), Lacistorhynchus (U = 27.0, p = 0.000), Diesingium (U = 36.0, p = 0.002), Callitetrarhynchus (U = 32.5, p = 0.019), Floriceps (U = 35, p = 0.018), Poecilancistrum (U = 42.0, p = 0.043), Otobothrium (U = 30.5, p = 0.016), Tetrarhynchobothrium (U = 45.0, p = 0.010), Eutetrarhynchus (U = 45.0, p = 0.010), Dollfusiella (U = 33.5, p = 0.008), and Trigonolobium (U = 45.0, p = 0.010).
The trypanorhynch composition within the shark family Carcharhinidae is characterised by the genera Dasyrhynchus (20.05% contribution), Nybelinia (14.52%), Callitetrarhynchus (13.90%), Otobothrium (12.61%), Floriceps (9.87%), Heteronybelinia (9.70%), Tentacularia (8.19%), and Poecilancistrum (3.96%) (Table S8). The Triakidae differed significantly from the Carcharhinidae and Sphyrnidae, being characterized by the genera Lacistorhynchus (31.15% contribution), Nybelinia (31.15%), Dollfusiella (23.20%), and Diesingium (9.10%). The Sphyrnidae harbored the genera Otobothrium (38.72%), Heteronybelinia (31.20%), and Nybelinia (22.56%). Only two genera, Nybelinia and Trigonolobium, were found in the Hemigaleidae, and Nybelinia and Gilquinia in the Scyliorhinidae. The parasite genera whose species numbers significantly differed in five families of carcharhiniforms were Heteronybelinia (H = 10.4, p = 0.017), Gilquinia (H = 12.6, p = 0.006), Dasyrhynchus (H = 10.5, p = 0.016), Pseudogrillotia (H = 8.3, p = 0.040), Lacistorhynchus (H = 21.9, p = 0.000), Diesingium (H = 13.4, p = 0.004), Floriceps (H = 9.6, p = 0.024), Otobothrium (H = 8.5, p = 0.037), Dollfusiella (H = 8.8, p = 0.033), and Trigonolobium (H = 14.5, p = 0.003).
3. Discussion
3.1. All Sharks
Our results demonstrate that depth distribution, diet composition and habitat type (benthic or pelagic) were the major influencing factors responsible for the composition of trypanorhynch assemblages and the groupings (cluster) of the sharks. Depth distribution affected the formation of all shark clusters (Figure 2): Cluster I included a single shallow/deep water species, Cluster II was dominated by deep water species, and Cluster III by shallow water species. Clusters IV and V were generated within Cluster III, and were dominated by invertebrate or teleost feeders, respectively. Cluster IV and Cluster V were also influenced by habitat type (benthic or pelagic), respectively. Moreover, Cluster II was dominated by piscivorous sharks. In general, no obvious pattern of the shark orders was found in the analysis. For instance, the most represented order Carcharhiniformes occurred in Clusters II and III (IV and V), and Lamniformes were found in Clusters I, II, and III (V).
Host-parasite associations are essential to gain information on the ecology of the hosts. The four shallow water shark species, Squatina squatina, Pristiophorus cirratus, Carcharias taurus, and Scyliorhinus stellaris, became an exception in Cluster II, because they were included in the deep water species group. That was probably due to the occurrence of Nybelinia, Grillotia, and Gilquinia that generally characterized the deep water species group, but which were also found in these four exception species. Thus, we can conclude that these four species which had been coded as shallow water types [12,13] might be collated to shallow/deep water shark species. Additionally, two shark species (Deania hystricosa and Centrophorus squamosus) that were part of Cluster II had been coded as unknown diet species. Grillotia that was found in both species characterized them as belonging into the fish feeder shark group. In Cluster III, Carcharhinus amblyrhynchoides was recognized as an unknown depth species. The trypanorhynchs Heteronybelinia, Callitetrarhynchus and Dasyrhynchus were found in this shark. Based on SIMPER analysis and Mann-Whitney U test, Callitetrarhynchus and Dasyrhynchus are characteristic trypanorhynchs of shallow water sharks. Hence, we consider C. amblyrhynchoides as a shallow water species. Three exception species were found in Cluster IV that was dominated by benthic and invertebratefeeding sharks, Galeorhinus galeus, Triakis semifasciata and Sutorectus tentaculatus. This exception leads us to suspect that the habitat type of G. galeus needs additional scrutiny; although Compagno [13] indicates this to be a pelagic species. Its trypanorhynch fauna suggests that it may be more benthic in its habits than currently recognized. From further analysis of the trypanorhynch assemblages we suggest that G. galeus, T. semifasciata, and S. tentaculatus prefer invertebrates over fish, although that had been coded as fish/invertebrate feeders.
In the last cluster V that was dominated by pelagic and piscivorous sharks, we found seven exception species. These were Hymenolepis microstoma, Mustelus manazo, Notorynchus cepedianus, Stegostoma fasciatum, Nebrius ferrugineus, Alopias superciliosus, and Carcharhinus melanopterus. C. melanopterus that had been coded as fish/invertebrate feeder seems to prefer more fish over invertebrates based on its trypanorhynch fauna. In A. superciliosus that was pelagic/benthic realm, we found Nybelinia, Mixonybelinia and Sphyriocephalus. Nybelinia, and Mixonybelinia characterized both groups, pelagic and benthic environment, while Sphyriocephalus characterized the pelagic environment only. Hence, we conclude that A. superciliosus, where adult species of Sphyriocephalus can be found, prefers the pelagic habitat. We also noted that adult Hepatoxylon was found in No. cepedianus [6] which suggests this shark to live in a pelagic/benthic habitat, although coded as benthic [12]. Hepatoxylon is a sphyriocephalid trypanorhynch occurring in oceanic but also deep water hosts. Although Sphyriocephalus characterises pelagic species, the species number of Sphyriocephalus was not significant different between pelagic and benthic sharks. We found parasite genera that characterised the pelagic and benthic group in M. manazo, S. fasciatum, and N. ferrugineus. The parasite genera found in these three sharks also characterized fish and invertebrate feeder. We suspect the three shark species were changing from invertebrate feeder and benthic habitat to be fish/invertebrate feeder and occurring in the pelagic/benthic habitat.
The last exception species in cluster V was H. microstoma. Based on its trypanorhynch fauna, H. microstoma was a benthic and invertebrate feeder. It became part of the group that was dominated by pelagic and fish feeder species. A plausible reason for this was the genus Nybelinia that was found in H. microstoma. Nybelinia was found in pelagic and benthic habitat species, as well as fish and invertebrate feeder species. The species numbers of Nybelinia in pelagic species were significantly higher than the numbers in benthic species. However, this was not significantly different between fish and invertebrate feeders. Another genus found in H. microstoma was Trigonolobium (one species). The species numbers of Trigonolobium in invertebrate feeders was significantly higher than the numbers in fish feeders, but it was not significantly different between pelagic and benthic sharks. We conclude that based on H. microstoma, habitat was a more dominant factor than diet in generating the Clusters IV and V within Cluster III.
Several shark species, based on the trypanorhynch fauna, were differently characterized than known from the literature [12,13,14,15]. An obvious reason for this is the poor knowledge on many shark species. Another explanation can be that the shark species can changed their behavior due to environmental changes (e.g., availability of fish, temperature increase, less production). Sharks might change their diet with regard to their size and prey diversity [16,17]. It has been stated that sharks are one of the most affected species concerning habitat loss or habitat change caused by climate change [18], which could explain their habitat changes. Another factor causing difficult affiliation of the sharks is that the knowledge on the trypanorhynch fauna is also far from complete, with new host records and new species descriptions added year by year. In general, trypanorhynchs have low (second) intermediate host and high final host specificity [11]. Although many trypanorhynchs have higher final host specificity, when they change the intermediate host (e.g., from invertebrate to fish or from fish to other fish that has a different habitat) they have a high possibility to reach further final host. This leads to regular host switches that have been recognized in trypanorhynch phylogeny. When this situation happens, then the trypanorhynch fauna makes it more difficult to characterize the shark species and results in conflicting evidence.
3.2. Carcharhiniformes
Among the factors investigated in the carcharhiniform analysis, the relationship between habitat and trypanorhynch assemblage was most remarkable. In fact, with the exception of G. galeus, the two major clusters of sharks recovered from the CLUSTER analysis of trypanorhynch assemblage data exactly corresponded to habitat occupied by the sharks. However, G. galeus coded as a pelagic shark was identified as a benthic shark (see Discussion above).
nMDS suggests that the trypanorhynch fauna has substantial potential to indicate the habit type of their elasmobranch final hosts. However, it is interesting to explore this in more depth by considering the individual genera comprising those assemblages. The trypanorhynch fauna of pelagic carcharhiniform sharks is primarily characterised by species of the genera Otobothrium, Dasyrhynchus, Heteronybelinia, Tentacularia, and Floriceps. In contrast, the trypanorhynch fauna of benthic carcharhiniform sharks is primarily characterized by species of Dollfusiella, Lacistorhynchus, Diesingium, and Eutetrarhynchus. All nine of these genera can be considered to be useful discriminatory taxa of environmental tendencies as the former five genera are abundant in pelagic sharks and scarce in benthic sharks while the latter four genera are scarce in pelagic sharks and abundant in benthic species. It should be noted that Nybelinia is abundant in sharks from both pelagic and benthic environments, as it occurs likewise in sharks and rays and, thus, it must be carefully used in combination with other taxa as a discriminatory taxon assigning the elasmobranch to a habit type. Thus a combination of tentaculariids, lacistorhynchids and otobothriids characterizes the Charcharhiniformes. According to Palm [6] these trypanorhynch taxa genera represent at least two different life-cycles. The intermediate hosts of tentaculariids, such as Tentacularia and Nybelinia, are pelagic or oceanic teleosts as well as squids as paratenic hosts. These trypanorhynchs develop robust plerocercoids in the intermediate host, and are characterised by low host specificity [11]. Lacistorhynchoids and otobothroids develop plerocerci with blastocysts, and the intermediate hosts are often pelagic fish. For example, the otobothriid Otobothrium penetrans mainly infests belonids as second intermediate hosts that are most common in the upper layer (10 m) of the water column [19,20]. Eutetrarhynchoids and gymnorhynchoids are generally missing within the pelagic carcharhiniforms. The former have life cycles that involve invertebrates as intermediate hosts, and are most commonly found on rajiform elasmobranchs, often in the benthic environment. The latter develop merocercoids and live in pelagic hosts.
The relationship between diet and trypanorhynch assemblage was similarly notable. Of the 28 species of carcharhiniform sharks included in the analysis, the diets of all but four species were coincident with the two major clusters of sharks recovered from the CLUSTER analysis of trypanorhynch assemblage data (Figure 4). Remarkably, these exceptions were the three species (C. melanopterus, T. semifasciata, and G. galeus) coded as consuming both fish and invertebrates based on the diet data available in Compagno [13]. These results suggest that fish play a larger role in the diet of C. melanopterus, than do invertebrates. They further suggest that the diets of T. semifasciatea and G. galeus emphasize invertebrates over fish.
At the individual genus level, Otobothrium, Dasyrhynchus, Heteronybelinia, Callitetrarhynchus, Tentacularia, and Floriceps appear to be excellent indicators of a preference for fish as they are particularly abundant in piscivorous sharks and rare to non-existent in sharks that consume invertebrates. In contrast, Dollfusiella, Lacistorhynchus, Diesingium, and Trigonolobium appear to be particularly significant in invertebrate feeders, and rare in piscivorous shark species. According to food preferences, the SIMPER analysis showed a similar picture. This is in line with the typical life cycle of the trypanorhynchs (see above). According to the trypanorhynch fauna, the studied carcharhiniforms of group 2, Carcharhinidae, and Sphyrnidae, obviously, are mainly pelagic fish feeders.
Carcharhiniforms that were separately clustered from the pelagic fish feeders were the Triakidae, Scyliorhinidae and Hemigaleidae. The benthic carcharhiniforms of group 1 are characterized by the trypanorhynch genera Dollfusiella, Lacistorhynchus, Nybelinia and Diesingium, while the invertebrate feeders were characterized by Dollfusiella, Lacistorhynchus, Diesingium, and Trigonolobium. The life cycle of the eutetrarhynchoid Dollfusiella is known to involve benthic invertebrates such as shrimps and crabs as second intermediate host, while the second intermediate host of Trigonolobium is yet unknown [6]. However, our results give a first indication that Trigonolobium might also involve marine invertebrates within its life cycle. The importance of Lacistorhynchus and Diesingium to distinguish this group of carcharhiniforms gives conflicting evidence. Both genera are typical trypanorhynchs from triakid sharks, with the plerocercus stage of the former often infecting a range of, mainly demersal or bottom-dwelling fish species. Thus, Lacistorhynchus and also the lacistorhynchid genus Diesingium might indicate the benthic environment of triakid carcharhiniform sharks, however, they cannot be utilized to analyze the feeding ecology of their hosts.
The two major clusters of sharks recovered from CLUSTER analysis were consistent with the carcharhiniform family. In no instance were species of the same family found in both clusters. In addition, the ANOSIM test showed that the trypanorhynch assemblage of the Carcharhinidae significantly differed from that of the Triakidae. The lack of significance between the Carcharhinidae and the Scyliorhinidae and the Hemigaleidae is likely to be the result of the lack of power of this test caused by the analyzed dataset, as the analysis included only a single species in each of the latter families. However, an additional level of resolution emerges if the phylogenetic relationships among carcharhiniform families will be considered. Based on the relatively robust, albeit preliminary molecular study Vélez-Zuazo and Agnarsson [21] concluded that hammerhead sharks (Sphyrnidae) were grouped with whaler sharks (Carcharhinidae) in phylogenetic relationship. The largest cluster (Cluster II) resulting from the agglomerative classification consists solely of members of this clade of sharks. In addition, ANOSIM failed to detect a significant difference between the assemblages of the Sphyrnidae and the Carcharhinidae. This test would be interesting to explore further with additional sphyrnid species because it included 16 species of carcharhinid sharks, but only three of the nine species of Sphyrnidae.
4. Materials and Methods
To study the host-parasite relationships between the Trypanorhyncha and their elasmobranch hosts, trypanorhynch data were analyzed from ca. 4000 host/parasite records compiled by Palm [6]. Only those shark species for which at least two adult trypanorhynch records had been available, were included in the analyses (See Table S1). Based on this minimum criterion, the trypanorhynch data consisted of 307 host/parasite records for a total of 50 shark species representing seven of the eight selachian orders (Carcharhiniformes, Lamniformes, Hexanchiformes, Orectilobiformes, Pristiophoriformes, Squaliformes, and Squatiniformes). These data, however, conspicuously emphasized carcharhiniform sharks, with 207 of the host/parasite records belonging to 28 species of this most speciose shark order.
A matrix was generated by scoring each shark species according to the total number of species in each trypanorhynch genus reported from any of the shark species. The absence of a trypanorhynch genus from a shark species was scored 0 in the matrix. A resemblance matrix among shark species was generated from this matrix of trypanorhynch assemblages choosing the Bray-Curtis measure [22,23]. All further multivariate data analyses were performed using the PRIMER software (release 6, Primer-E Ltd. 6.1.11, Meadow View, Plymouth, UK; Clarke and Warwick 2001) [24]. Sharks were classified by hierarchical agglomerative cluster analysis of this resemblance matrix using the group-average linking method CLUSTER [23]. The results of these analyses were then ordinated using nMDS two-dimensional ordination plots of shark species which were generated from trypanorhynch assemblage data for each of the factors under consideration. These plots were produced from both the full matrix of data for the selachian analyses, and also from the more restricted matrix consisting exclusively of carcharhiniform shark data. In each case, the conditions of the factor under consideration were superimposed on the ordination plot for that factor by coding each shark species with a symbol representing the condition it exhibited. For each factor, ANOSIM [25,26] was performed to test significant differences among conditions of the factor as reflected by trypanorhynch assemblage. In instances in which significant differences were seen, SIMPER analyses [27] were performed to explore which trypanorhynch genera contributed most to these differences. In these analyses trypanorhynch genera were ordered according to decreasing contribution (%) to the total similarity observed. Genera accounting for high levels of similarity were determined to be good candidates as individual indicators.
For the analyses, each shark species was categorized for habitat (benthic or pelagic) and diet (primarily invertebrates and primarily teleosts). For the analysis of the Selachii, sharks were also coded for depth of distribution (below 200 m as deepwater, 200 m or less as shallow water species). In the carcharhiniform analyses taxonomy was assessed at the level of family; in the Selachii analyses taxonomy was assessed at the level of order? Sharks were categorized for these factors based on information published in the work of Compagno [12,13,14,15], because this provides detailed data for all of the target species. Shark taxonomic position at order and family levels was also based on Compagno [14]. Sharks were categorised for diet, habitat, and depth distribution for the Hexanchiformes, Squaliformes, and Squatiniformes [11], for the Carcharhiniformes [12], and for the Lamniformes, Orectilobiformes, and Pristiophoriformes [14]. Categorization by habitat type was fairly straightforward as most shark species were either benthic or pelagic. Coding for depth was similarly unambiguous with the recorded depths of most sharks being either above, or below 200 m. Categorisation of sharks according to diet, however, was somewhat more ambiguous. While many sharks had a clear preference for either invertebrate or teleost prey, other species consumed both types of prey with some regularity. In such instances, sharks were coded as exhibiting both conditions of this factor. For the purpose of this study, two mutually exclusive conditions for this factor were chosen, based on the final intermediate hosts utilized by trypanorhynchs (i.e., invertebrates vs. teleosts).
Kruskal-Wallis test and Mann-Whitney U test were performed to compare the number of parasite species from each genus in each shark group [28,29]. The Kruskal-Wallis test was employed for more than two categories of sharks (e.g., selachian order; Carcharhiniformes, Lamniformes, Hexanchiformes, Orectilobiformes, Pristiophoriformes, Squaliformes, and Squatiniformes), and Mann-Whitney U test was employed for two categories of sharks (e.g., habitat; benthic and pelagic).
5. Conclusions
This study demonstrates that nMDS of the elasmobranch parasitic Trypanorhyncha Diesing, 1863, is a useful tool for investigating parasites as indicators of the host biology in marine ecosystems. Transmission through the marine food web (via predation) and an unambiguous identification in the final (sharks and rays) and intermediate (teleosts, other marine invertebrates) hosts allow conclusions to be made on the feeding biology of the host. The relationship between trypanorhynch assemblage and investigated factors for all sharks was complex, but we found that depth distribution, diet, and habitat type were the major influencing factors. The relationships within carcharhiniform species were more robust, especially if the factor habitat is considered. Overlapping host ecology was evident even in phylogenetically-distant hosts, demonstrating host switches and low host specificity in some of the trypanorhynch taxa. Considering that the underlying dataset is based on a host-parasite list from 2004 [6] and more species descriptions and host records are added continuously nMDS proves to be a powerful tool to attribute aspects of shark biology (phylogeny, habitat, depth distribution, diet) to rare and difficult-to-observe elasmobranch species in future.
Supplementary Materials
The following are available online at www.mdpi.com/2410-3888/2/2/8/s1, Table S1: List of shark species analysed in the recent study; Table S2: SIMPER analysis results for similarities in each habitat of all sharks; Table S3: SIMPER analysis results for similarities in each diet of all sharks; Table S4: SIMPER analysis results for similarities in each depth of all sharks; Table S5: SIMPER analysis results for similarities in each order of all sharks; Table S6: SIMPER analysis results for similarities in each habitat of carcharhiniform; Table S7: SIMPER analysis results for similarities in diet of carcharhiniform; Table S8: SIMPER analysis results for similarities in each family of carcharhiniform.
Acknowledgments
This paper was presented and discussed on the 9th International Symposium on Fish Parasites, 31 August–4 September 2015, in Valencia, Spain. The analyses were conducted together with Janine N. Caira during the stay of HWP at the University Connecticut, Ecology and Evolutionary Biology. We would like to thank critical reviewers for improving the manuscript. The study was financially supported by the German Research Council (DFG PA 664/4-1).
Author Contributions
H.W.P. collected the data and together with U.P. designed and performed the analyses. I.Y. contributed figures and supported the SIMPER analyses and statistical tests. The paper was jointly written among authors.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study, in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Palm, H.W. Fish parasites as biological indicators in a changing world: Can we monitor environmental impact and climate change? In Progress in Parasitology; Mehlhorn, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 223–250. [Google Scholar]
- Vaullegeard, A. Recherches sur le Tétrarhynques. Mém. Soc. Linn. Normandie 1899, 2, 187–376. [Google Scholar]
- Southwell, T. A monograph on cestodes of the order Trypanorhyncha from Ceylon and India, part 1. Ceylon J. Sci. Ser. B 1929, 15, 169–317. [Google Scholar]
- Dollfus, R.P. Études Critiques sur les Tétrarhynques du Museum de Paris; Éditions du Muséum: Paris, France, 1942; Volume 19, pp. 1–466. [Google Scholar]
- Campbell, R.A.; Beveridge, I. Order Trypanorhyncha diesing, 1863. In Keys to the Cestode Parasites of Vertebrates; Khalil, L.F., Jones, A., Bray, R.A., Eds.; CAB International: Wallingford, UK, 1994; pp. 51–148. [Google Scholar]
- Palm, H.W. The Trypanorhyncha Diesing, 1863; PKSPL-IPB Press: Bogor, Indonesia, 2004. [Google Scholar]
- Palm, H.W.; Waeschenbach, A.; Olson, P.D.; Littlewood, T.J. Molecular phylogeny and evolution of the Trypanorhyncha Diesing, 1863 (Platyhelminthes: Cestoda). Mol. Phylogenet. Evol. 2009, 52, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Olson, P.D.; Caira, J.N.; Jensen, K.; Overstreet, R.M.; Palm, H.W.; Beveridge, I. Evolution of the trypanorhynch tapeworms: Parasite phylogeny supports independent lineages of sharks and rays. Int. J. Parasitol. 2010, 40, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, I.; Campbell, R.A. Three new genera of trypanorhynch cestodes from Australian elasmobranch fishes. Syst. Parasitol. 2005, 60, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, I.; Justine, J.L. Gilquiniid cestodes (Trypanorhyncha) from elasmobranch fishes off New Caledonia with descriptions of two new genera and a new species. Syst. Parasitol. 2006, 65, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Palm, H.W.; Caira, J. Host specificity of adult versus larval cestodes of the order Trypanorhyncha (Cestoda). Int. J. Parasitol. 2008, 38, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date: Part 1. Hexanchiformes to Lamniformes; Food and Agricultural Organization: Rome, Italy, 1984. [Google Scholar]
- Compagno, L.J.V. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date: Part 2.Carcharhiniformes; Food and Agricultural Organization: Rome, Italy, 1984. [Google Scholar]
- Compagno, L.J.V. Checklist of living elasmobranchs. In Sharks, Skates, and Rays: The Biology of Elasmobranch Fishes; Hamlett, W.C., Ed.; Johns Hopkins University Press: Baltimore, MD, USA, 1999; pp. 471–498. [Google Scholar]
- Compagno, L.J.V. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date: Bullhead, Mackerel and Carpet Sharks (Heterodontifofrmes, Lamniformes, and Orectolobiformes); Food and Agricultural Organization: Rome, Italy, 2001. [Google Scholar]
- Elbert, D.A. Ontogenetic changes in the diet of the sevengill shark (Notorynchus cepedianus). Mar. Freshw. Res. 2002, 53, 517–523. [Google Scholar] [CrossRef]
- McElroy, W.D.; Wetherbee, B.M.; Mostello, C.S.; Lowe, C.G.; Crow, G.L.; Wass, R.C. Food habits and ontogenetic changes in the diet of the sandbar shark, Carcharhinus plumbeus, in Hawaii. Environ. Biol. Fishes 2006, 76, 81–92. [Google Scholar] [CrossRef]
- Hazen, E.L.; Jorgensen, S.; Rykaczewski, R.R.; Bograd, S.J.; Foley, D.G.; Jonsen, I.D.; Shaffer, S.A.; Dunne, J.P.; Costa, D.P.; Crowder, L.B.; et al. Predicted habitat shifts of Pacific top predators in a changing climate. Nat. Clim. Chang. 2012, 3, 234–238. [Google Scholar] [CrossRef]
- Jakob, E.; Palm, H.W. Parasites of commercially important fish species from the southern Java coast, Indonesia, including the distribution pattern of trypanorhynch cestodes. Verh. Ges. Ichthyol. Bd. 2006, 5, 165–191. [Google Scholar]
- Petersen, F.; Palm, H.W.; Möller, H.; Cuzi, M.A. Flesh parasites of fish from central Philippine waters. Dis. Aquat. Org. 1993, 15, 81–86. [Google Scholar] [CrossRef]
- Vélez-Zuazo, X.; Agnarsson, I. Shark tales: A molecular species-level phylogeny of sharks (Selachimorpha, Chondrichthyes). Mol. Phylogenet. Evol. 2011, 58, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Field, J.G.; Clarke, K.R.; Warwick, R.M. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser. 1982, 8, 37–52. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in marine communities: An approach to statistical analysis and interpretation, PRIMER-E, 2nd ed.Plymouth, 2001; 172p. [Google Scholar]
- Chapmand, M.G.; Underwood, A.J. Ecological patterns in multivariate assemblages: Information and interpretation of negative values in ANOSIM tests. Mar. Ecol. Prog. Ser. 1999, 180, 257–265. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. A comparison of some methods for analysing changes in benthic community structure. J. Mar. Biol. Assoc. UK 1991, 71, 225–244. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Kruskal, J.B.; Wish, M. Multidimensional scaling. In Sage University Paper series on Quantitative Applications in the Social Sciences; Sage Publications: Beverly Hills, CA, USA; London, UK, 1978. [Google Scholar]
Figure 1.
Cluster analysis of all sharks according to Trypanorhyncha assemblages. Roman numerals I–V indicate the five major clusters.
Figure 1.
Cluster analysis of all sharks according to Trypanorhyncha assemblages. Roman numerals I–V indicate the five major clusters.

Figure 2.
Multidimensional scaling plot for all shark species based on Bray-Curtis similarity and grouped by depth (A), habitat (B), diet (C), and order (D). Clusters I–III marked in blue, cluster IV in red, and cluster V in green.
Figure 2.
Multidimensional scaling plot for all shark species based on Bray-Curtis similarity and grouped by depth (A), habitat (B), diet (C), and order (D). Clusters I–III marked in blue, cluster IV in red, and cluster V in green.

Figure 3.
Cluster analysis of carcharhiniform sharks according to Trypanorhyncha assemblages. Roman letters I–II indicate the two major clusters.
Figure 3.
Cluster analysis of carcharhiniform sharks according to Trypanorhyncha assemblages. Roman letters I–II indicate the two major clusters.
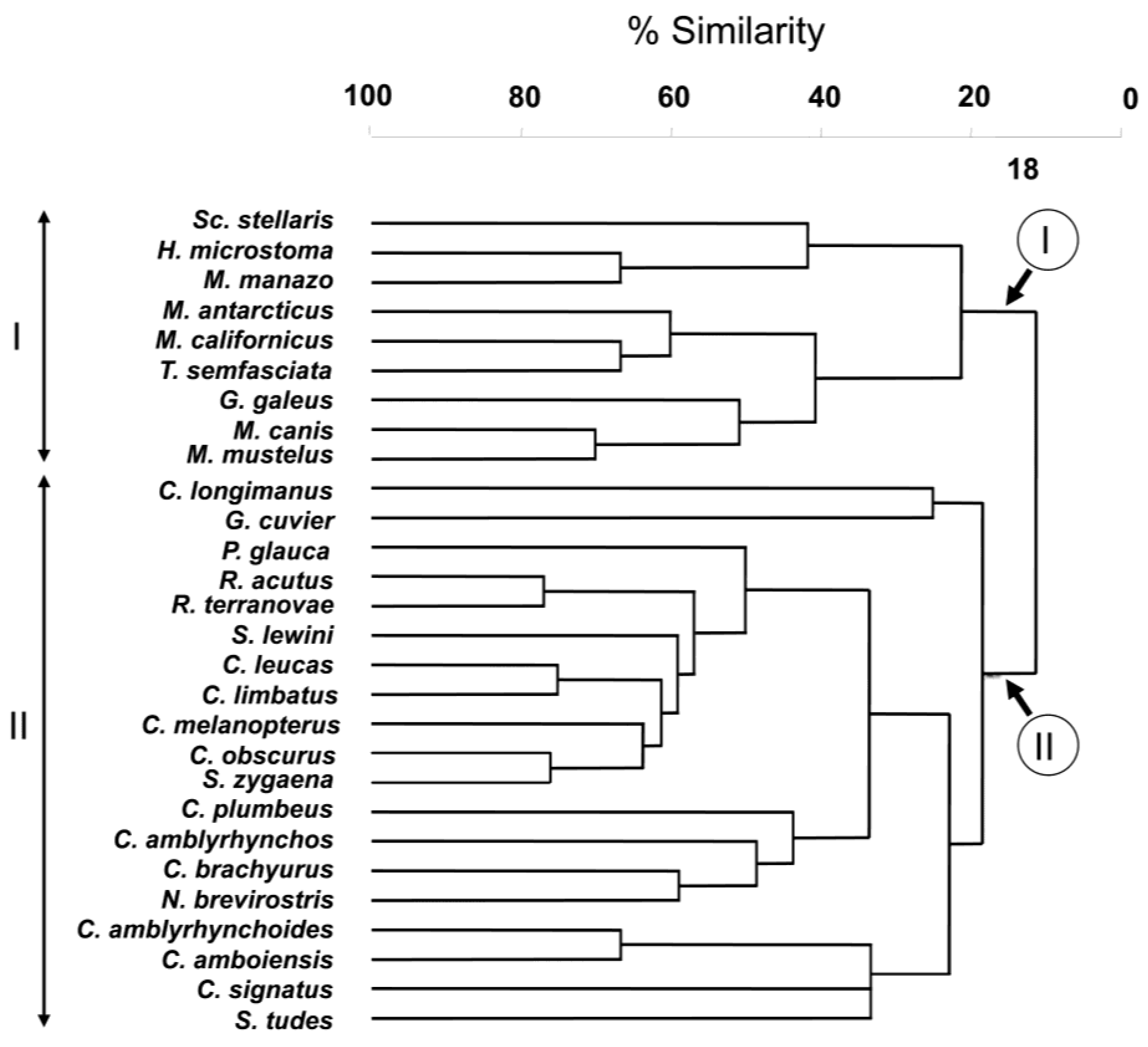
Figure 4.
Multidimensional scaling plot for carcharhiniform species based on Bray-Curtis similarity and grouped by habitat (A), diet (B), and family (C). Clusters I and II marked in blue.
Figure 4.
Multidimensional scaling plot for carcharhiniform species based on Bray-Curtis similarity and grouped by habitat (A), diet (B), and family (C). Clusters I and II marked in blue.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Palm, H.W.; Yulianto, I.; Piatkowski, U. Trypanorhynch Assemblages Indicate Ecological and Phylogenetical Attributes of Their Elasmobranch Final Hosts. Fishes 2017, 2, 8. https://doi.org/10.3390/fishes2020008
AMA Style
Palm HW, Yulianto I, Piatkowski U. Trypanorhynch Assemblages Indicate Ecological and Phylogenetical Attributes of Their Elasmobranch Final Hosts. Fishes. 2017; 2(2):8. https://doi.org/10.3390/fishes2020008
Chicago/Turabian StylePalm, Harry W., Irfan Yulianto, and Uwe Piatkowski. 2017. "Trypanorhynch Assemblages Indicate Ecological and Phylogenetical Attributes of Their Elasmobranch Final Hosts" Fishes 2, no. 2: 8. https://doi.org/10.3390/fishes2020008