
Improved Waterlogging Tolerance in Roots of Cucumber Plants after Inoculation with Arbuscular Mycorrhizal Fungi

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mycorrhizal Fungal Preparations
2.2. Plant Arrangement and Design
2.3. Determination of Plant Biomass and Root Mycorrhizal Infestation Rate
2.4. Determination of Five Osmolytes in Roots
2.5. Relative Expression of CsPIPs and CsHsp70 Genes in Roots
2.6. Data Analysis
3. Results
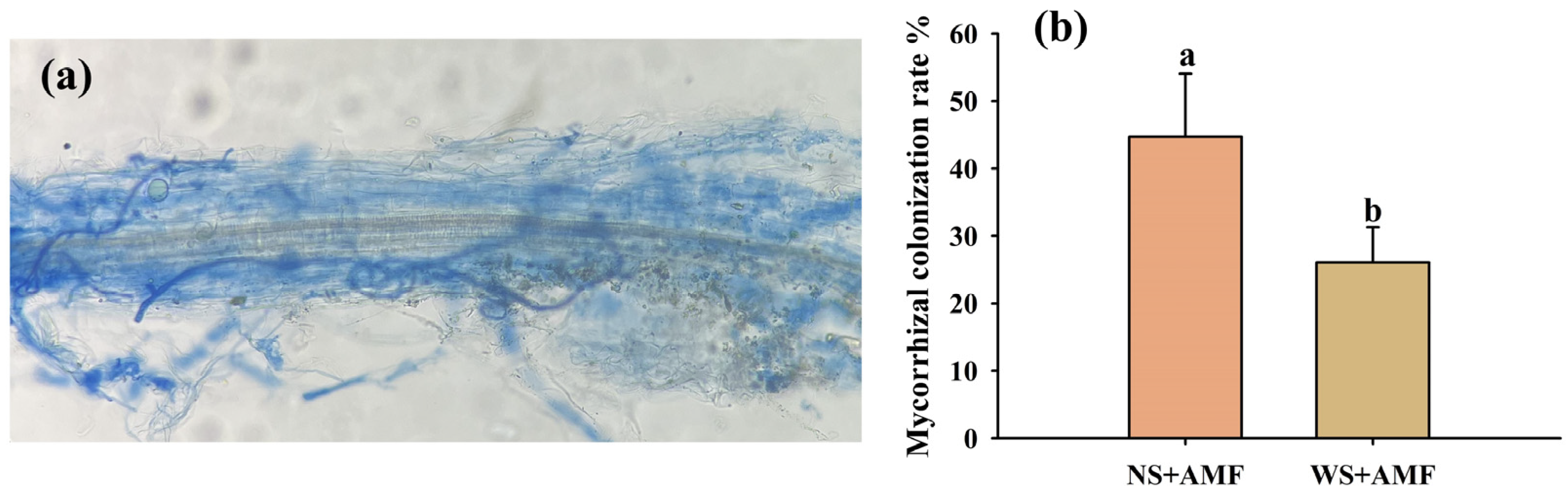
3.1. Response of Root AMF Infection and Biomass Production to Waterlogging Stress
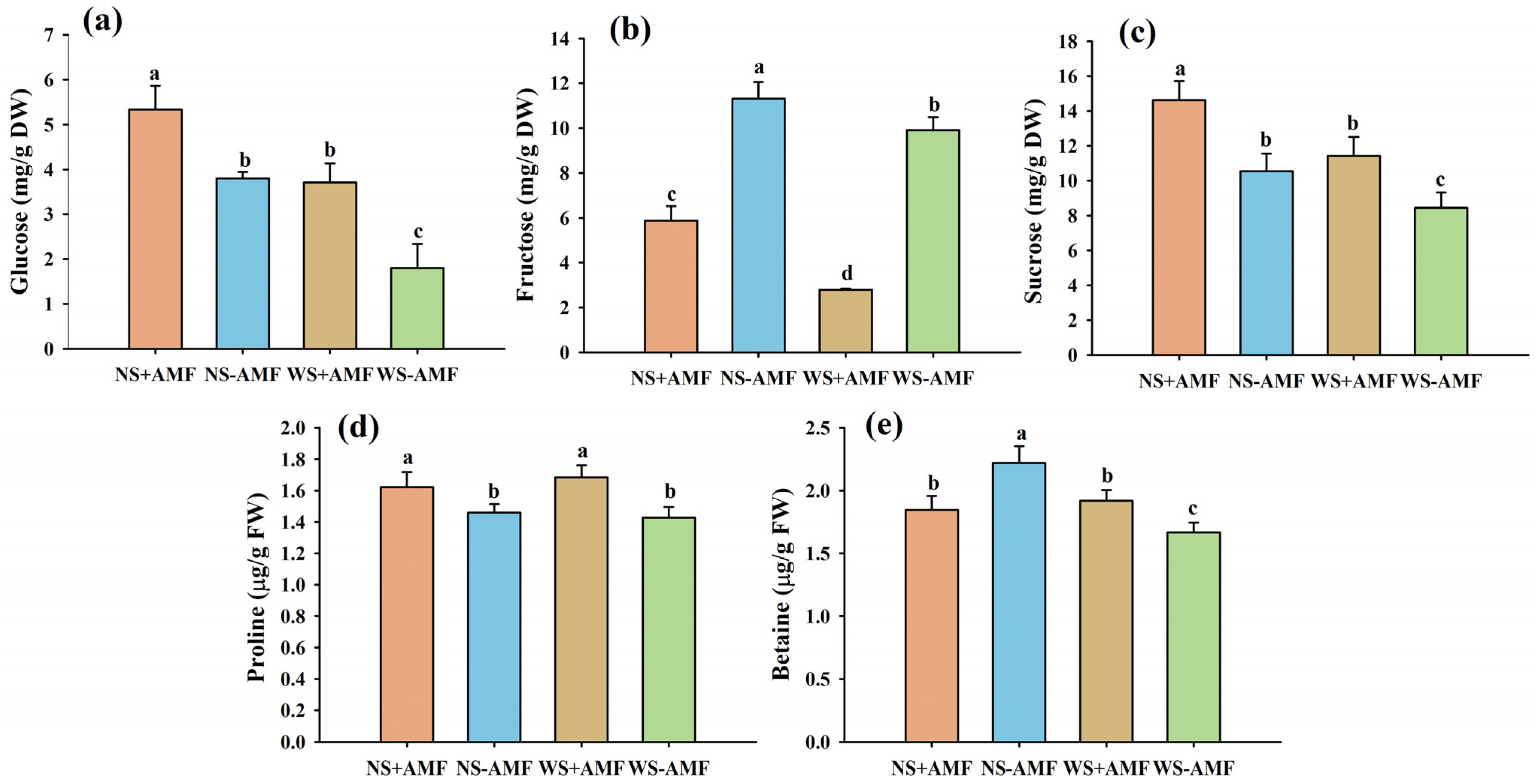
3.2. Response of Osmolytes in Roots to Waterlogging Stress
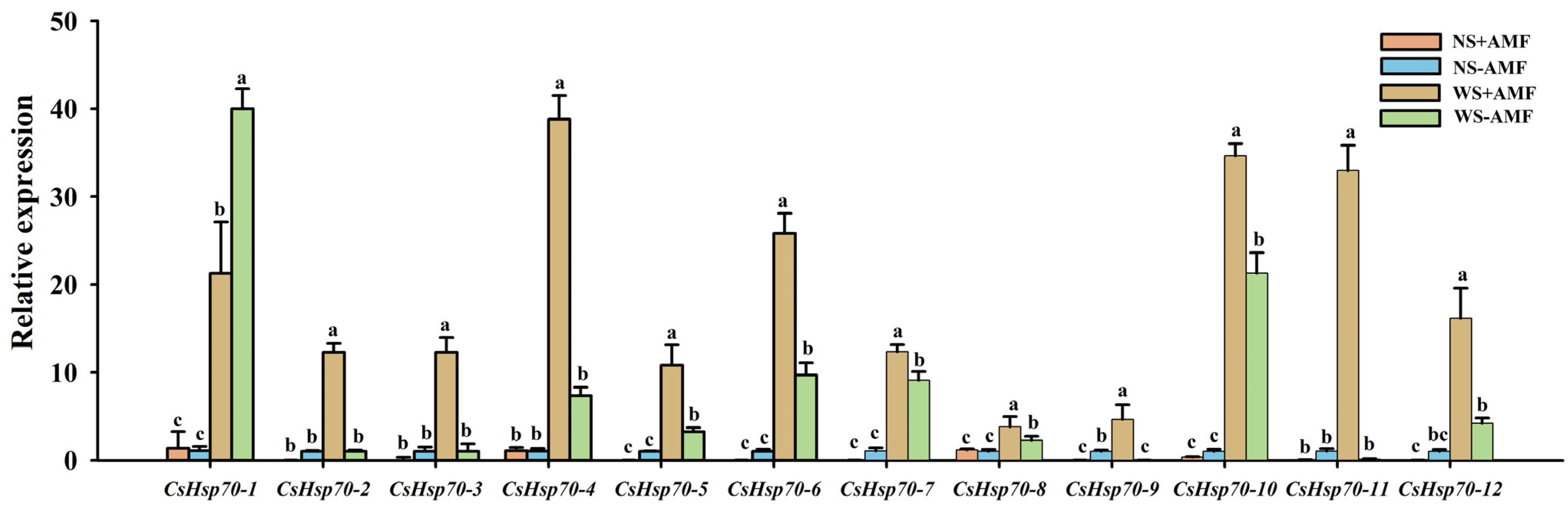
3.3. Response of CsHsp70s Gene Expression Levels in Roots to Waterlogging Stress
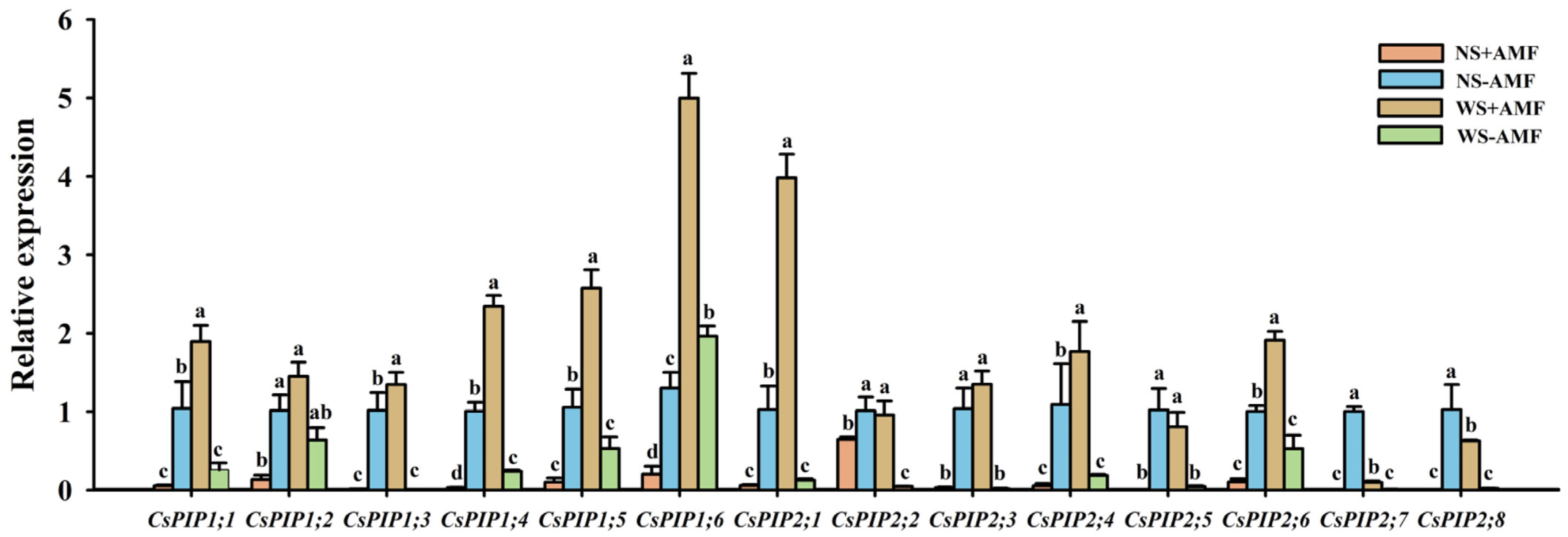
3.4. Response of CsPIP Gene Expression Levels in Roots to Waterlogging Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qi, X.H.; Xu, X.W.; Lin, X.J.; Zhang, W.J.; Chen, X.H. Identification of differentially expressed genes in cucumber (Cucumis sativus L.) root under waterlogging stress by digital gene expression profile. Genomics 2012, 99, 160–168. [Google Scholar] [CrossRef]
- Qi, X.H.; Chen, R.F.; Xu, Q.; Chen, X.H. Preliminary analysis of cucumber submergence tolerance at seedling stage. China Vegetables 2011, 4, 23–28. [Google Scholar] [CrossRef]
- Bennett, A.E.; Groten, K. The costs and benefits of plant–arbuscular mycorrhizal fungal interactions. Annu. Rev. Plant Biol. 2022, 73, 649–672. [Google Scholar] [CrossRef]
- Wang, Y.; Zou, Y.N.; Shu, B.; Wu, Q.S. Deciphering molecular mechanisms regarding enhanced drought tolerance in plants by arbuscular mycorrhizal fungi. Sci. Hortic. 2023, 308, 111591. [Google Scholar] [CrossRef]
- Yang, H.; Koide, R.T.; Zhang, Q. Short-term waterlogging increases arbuscular mycorrhizal fungal species richness and shifts community composition. Plant Soil 2016, 404, 373–384. [Google Scholar] [CrossRef]
- Tuheteru, F.D.; Kusmana, C.; Mansur, I.; Iskandar, I. Response of lonkida (Nauclea orientalis L.) towards mycorrhizal inoculum in waterlogged condition. Biotropia 2015, 22, 61–71. [Google Scholar] [CrossRef]
- Orchard, S.; Standish, R.J.; Nicol, D.; Gupta, V.V.S.R.; Ryan, M.H. The response of fine root endophyte (Glomus tenue) to waterlogging is dependent on host plant species and soil type. Plant Soil. 2016, 403, 305–315. [Google Scholar] [CrossRef]
- Matsumura, A.; Horii, S.; Ishii, T. Observation of arbuscular mycorrhizal network system between trifoliate orange and some grasses under waterlogged conditions. Acta. Hortic. 2008, 773, 69–75. [Google Scholar] [CrossRef]
- Tuo, X.Q.; Li, S.; Wu, Q.S.; Zou, Y.N. Alleviation of waterlogged stress in peach seedlings inoculated with Funneliformis mosseae: Changes in chlorophyll and proline metabolism. Sci. Hortic. 2015, 197, 130–134. [Google Scholar] [CrossRef]
- Liang, S.M.; Hashem, A.; Abd-Allah, E.F.; Wu, Q.S. Root-associated symbiotic fungi enhance waterlogging tolerance of peach seedlings by increasing flavonoids and activities and gene expression of antioxidant enzymes. Chem. Biol. Technol. Agric. 2023, 10, 124. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Huang, Y.M. The arbuscular mycorrhizal fungus Diversispora spurca ameliorates effects of waterlogging on growth, root system architecture and antioxidant enzyme activities of citrus seedlings. Fungal Ecol. 2013, 6, 37–43. [Google Scholar] [CrossRef]
- Ortas, I. Effect of mycorrhiza application on plant growth and nutrient uptake in cucumber production under field conditions. Span. J. Agric. Res. 2010, 8, 116–122. [Google Scholar] [CrossRef]
- Haghighi, M.; Mohammadnia, S.; Attai, Z.; Pessarakli, M. Effects of mycorrhiza inoculation on cucumber growth irrigated with saline water. J. Plant Nutr. Soil Sci. 2017, 40, 128–137. [Google Scholar] [CrossRef]
- Tian, X.; Liu, X.Q.; Liu, X.R.; Li, Q.S.; Abd-Allah, E.F.; Wu, Q.S. Mycorrhizal cucumber with Diversispora versiformis has active heat stress tolerance by up-regulating expression of both CsHsp70s and CsPIPs genes. Sci. Hortic. 2023, 319, 112494. [Google Scholar] [CrossRef]
- Zhang, M.Q.; Wang, Y.S. Seven species of VA mycorrhizal fungi from northern China. Acta Mycol. Sin. 1991, 10, 13–21. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Br. Mycol. Soc. Trans. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Wu, Q.S.; Srivastava, A.K.; Li, Y. Effect of mycorrhizal symbiosis on growth behavior and carbohdyrate metabolism of trifoliate orange under different substrate P levels. J. Plant Growth Regul. 2015, 34, 495–508. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water–stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Liu, X.R.; Rong, Z.Y.; Tian, X.; Hashem, A.; Abd_Allah, E.F.; Zou, Y.N.; Wu, Q.S. Mycorrhizal fungal effects on plant growth, osmolytes, and CsHsp70s and CsPIPs expression in leaves of cucumber under a short-term heat stress. Plants 2023, 12, 2917. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cheng, X.F.; Wu, H.H.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhizal response strategies of trifoliate orange under well-watered, salt stress, and waterlogging stress by regulating leaf aquaporin expression. Plant Physiol. Biochem. 2021, 162, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Šibanc, N.; Clark, D.R.; Helgason, T.; Dumbrell, A.J.; Maček, I. Extreme environments simplify reassembly of communities of arbuscular mycorrhizal fungi. Msystems 2024, 9, e01331-23. [Google Scholar] [CrossRef]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Molina, S.; Zamarreño, A.M.; García-Mina, J.M.; Aroca, R. The symbiosis with the arbuscular mycorrhizal fungus Rhizophagus irregularis drives root water transport in flooded tomato plants. Plant Cell Physiol. 2014, 55, 1017–1029. [Google Scholar] [CrossRef]
- Di Miceli, G.; Vultaggio, L.; Sabatino, L.; De Pasquale, C.; La Bella, S.; Consentino, B.B. Synergistic Effect of a plant-derived protein hydrolysate and arbuscular mycorrhizal fungi on eggplant grown in open fields: A two-year study. Horticulturae 2023, 9, 592. [Google Scholar] [CrossRef]
- Ahmed, S.; Nawata, E.; Hosokawa, M.; Domae, Y.; Sakuratani, T. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging. Plant Sci. 2002, 163, 117–123. [Google Scholar] [CrossRef]
- Loewe, A.; Einig, W.; Shi, L.; Dizengremel, P.; Hampp, R. Mycorrhiza formation and elevated CO2 both increase the capacity for sucrose synthesis in source leaves of spruce and aspen. New Phytol. 2000, 145, 565–574. [Google Scholar] [CrossRef]
- Gupta, S.; Thokchom, S.D.; Kapoor, R. Arbuscular mycorrhiza improves photosynthesis and restores alteration in sugar metabolism in Triticum aestivum L. grown in arsenic contaminated soil. Front. Plant Sci. 2021, 12, 640379. [Google Scholar] [CrossRef]
- Pfeffer, P.E.; Douds, J.D.D.; Bécard, G.; Shachar-Hill, Y. Carbon uptake and the metabolism and transport of lipids in an arbuscular mycorrhiza. J. Plant Physiol. 1999, 120, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Baher, N. Role of arbuscular mycorrhizal symbiosis in proline biosynthesis and metabolism of Cicer arietinum L. (Chickpea) genotypes under salt stress. J. Plant Growth Regul. 2013, 32, 767–778. [Google Scholar] [CrossRef]
- Abbaspour, H.; Pour, F.S.; Abdel-Wahhab, M.A. Arbuscular mycorrhizal symbiosis regulates the physiological responses, ion distribution and relevant gene expression to trigger salt stress tolerance in pistachio. Physiol. Mol. Biol. Plants 2021, 27, 1765–1778. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Landi, S.; Capasso, G.; Ben, A.F.E.; Jallouli, S.; Ayadi, S.; Trifa, Y.; Esposito, S. Different roles of heat shock proteins (70 kDa) during abiotic stresses in barley (Hordeum vulgare) genotypes. Plants 2019, 8, 248. [Google Scholar] [CrossRef]
- Liu, M.Y.; Sun, J.; Wang, K.Y.; Liu, D.; Li, Z.Y.; Zhang, J. Spermidine enhances waterlogging tolerance via regulation of antioxidant defence, heat shock protein expression and plasma membrane H+-ATPase activity in Zea mays. J. Agron. Crop Sci. 2014, 200, 199–211. [Google Scholar] [CrossRef]
- Liang, S.M.; Zou, Y.N.; Shu, B.; Wu, Q.S. Arbuscular mycorrhizal fungi and endophytic fungi differentially modulate polyamines or proline of peach in response to soil flooding. Pedosphere 2024, 34, 1–13. [Google Scholar] [CrossRef]
- Singh, R.K.; Deshmukh, R.; Muthamilarasan, M.; Rani, R.; Prasad, M. Versatile roles of aquaporin in physiological processes and stress tolerance in plants. Plant Physiol. Biochem. 2020, 149, 178–189. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Aroca, R. Evaluation of the possible participation of drought-induced genes in the enhanced tolerance of arbuscular mycorrhizal plants to water deficit. In Mycorrhiza; Varma, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 185–205. [Google Scholar] [CrossRef]
- Liu, T.; Zhen, L.; Chen, H.; Ming, T.; Hao, Q.Z. Effect of Rhizophagus irregularis on osmotic adjustment, antioxidation and aquaporin PIP genes expression of Populus × canadensis ‘Neva’ under drought stress. Acta. Physiol. Plant. 2016, 38, 191–199. [Google Scholar] [CrossRef]
- Ricardo, A.; Alberto, B.; Moira, S.; José, A.P.; Custodia, C.; Gabriela, A.; Juan, M.R.L. Expression analysis of the first arbuscular mycorrhizal fungi aquaporin described reveals concerted gene expression between salt stressed and nonstressed mycelium. Mol. Plant Microbe Interact. 2009, 22, 1169–1178. [Google Scholar] [CrossRef]
- Li, T.; Hu, Y.J.; Hao, Z.P.; Li, H.; Wang, Y.S.; Chen, B.D. First cloning and characterization of two functional aquaporin genes from an arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2013, 197, 617–630. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, N.; Liu, Z.; Tian, X.; Wang, D.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.-S.; Zou, Y.-N. Improved Waterlogging Tolerance in Roots of Cucumber Plants after Inoculation with Arbuscular Mycorrhizal Fungi. Horticulturae 2024, 10, 478. https://doi.org/10.3390/horticulturae10050478
Xiang N, Liu Z, Tian X, Wang D, Hashem A, Abd_Allah EF, Wu Q-S, Zou Y-N. Improved Waterlogging Tolerance in Roots of Cucumber Plants after Inoculation with Arbuscular Mycorrhizal Fungi. Horticulturae. 2024; 10(5):478. https://doi.org/10.3390/horticulturae10050478
Chicago/Turabian StyleXiang, Nan, Zhen Liu, Xiao Tian, Dan Wang, Abeer Hashem, Elsayed Fathi Abd_Allah, Qiang-Sheng Wu, and Ying-Ning Zou. 2024. "Improved Waterlogging Tolerance in Roots of Cucumber Plants after Inoculation with Arbuscular Mycorrhizal Fungi" Horticulturae 10, no. 5: 478. https://doi.org/10.3390/horticulturae10050478