
A Comparative Study of Yeasts for Rosa roxburghii Wine Fermentation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals and Yeast Strains
2.1.2. Preparation of Cili Juice
2.1.3. Preparation of Cili Wine Using Different Yeast Strains
2.2. Methods
2.2.1. Sulfite Tolerance and Flocculation
2.2.2. Hydrogen Sulfide Production Capacity
2.2.3. Fermentation Vigor and Growth Curve Determination
2.2.4. Chemical Analysis of Resultant Wines
2.2.5. Volatile Components of Resultant Wines
2.3. Suitability of Yeast Strains for Cili Winemaking
2.4. Statistical Analysis
3. Results
3.1. Traits of Yeast Strains for Winemaking
3.1.1. Tolerance of Total SO2
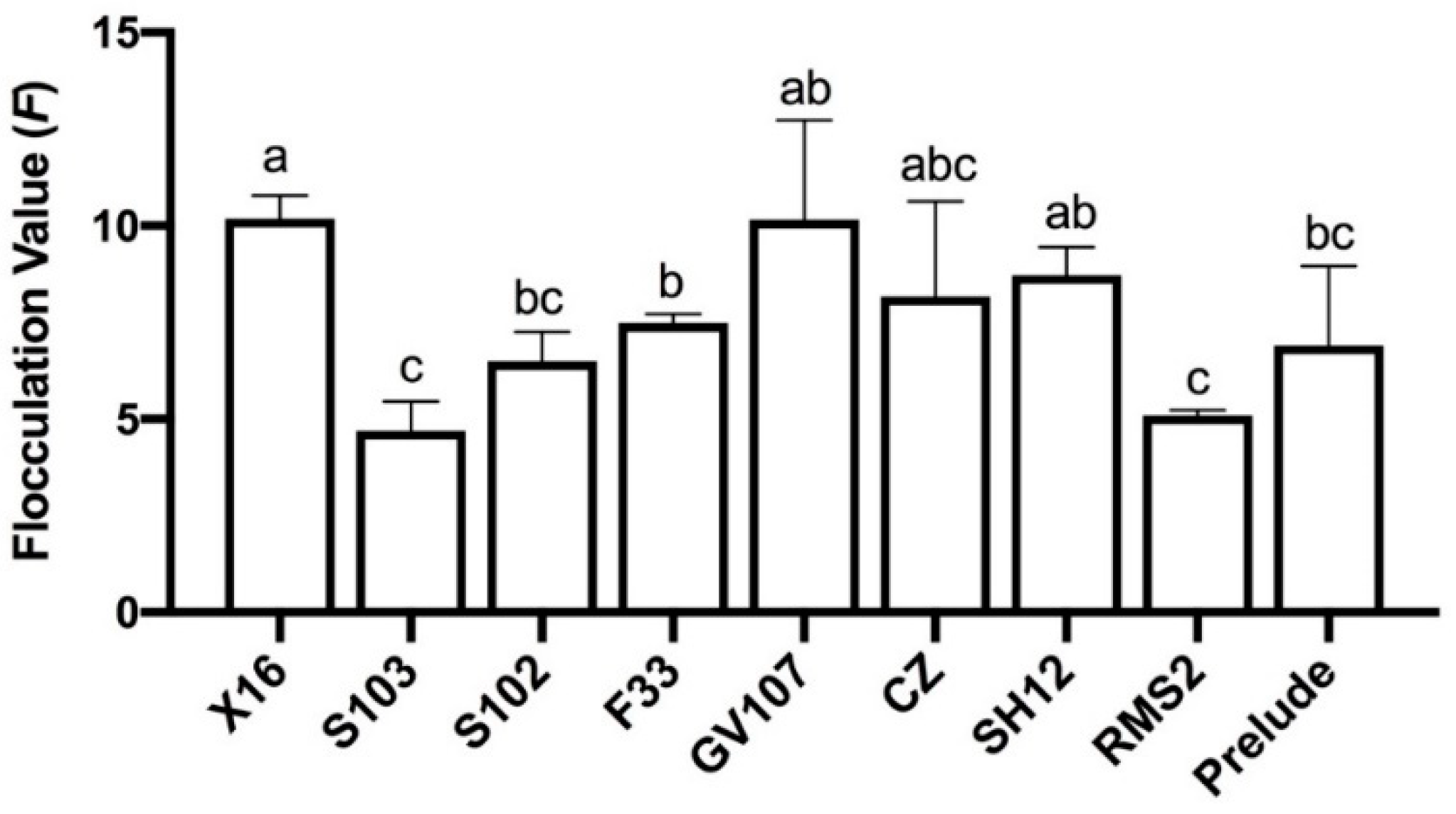
3.1.2. Flocculation
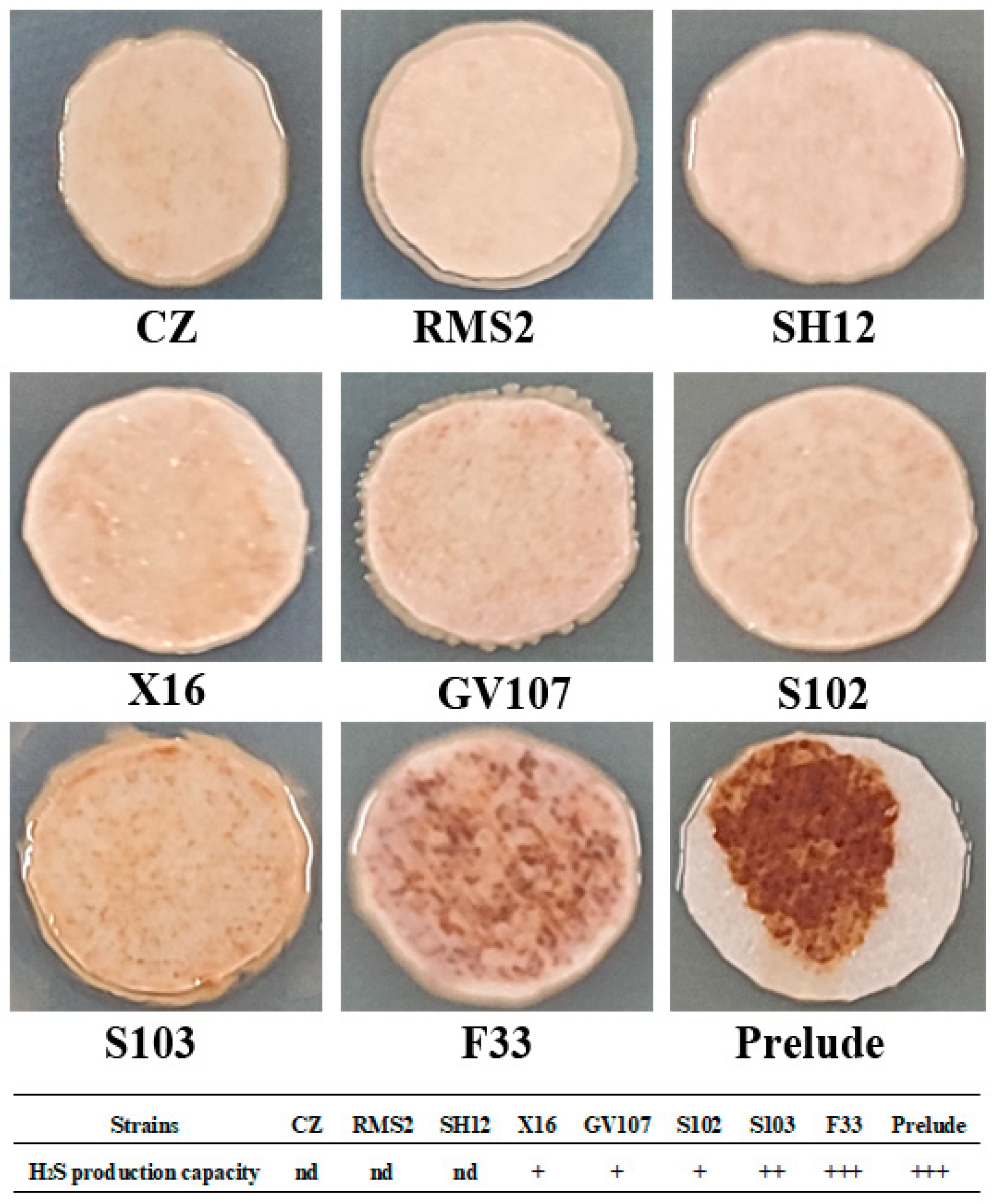
3.1.3. H2S Production Capacity
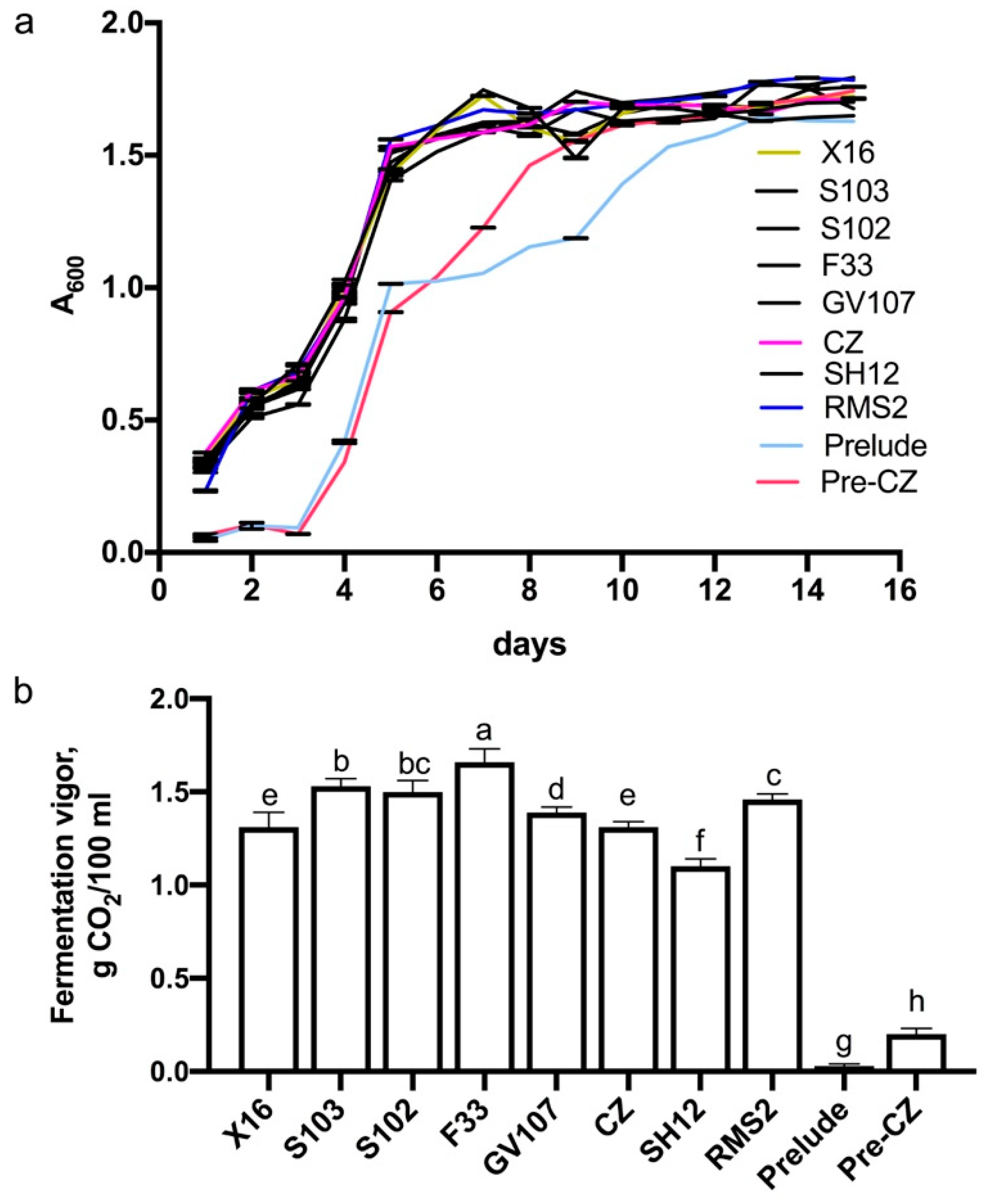
3.1.4. Growth Curve and Fermentation Vigor
3.2. Wine Analysis
3.2.1. Chemical Components
3.2.2. Headspace Volatile Compounds of Cili Wines
Higher Alcohols
Acids
Esters
Aldehydes
Volatile Phenols
Other Volatile Compounds
3.2.3. Aroma-Contributing Compounds of Cili Wine
3.3. Scoring of Oenological Properties of Yeast Strains for Cili Fermentation and the Resultant Wine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| X16-Saccharomyces cerevisiae | X16 (Laffort) |
| F33-S. cerevisiae | Actiflore F33, (Laffort) |
| RMS2-S. cerevisiae | Actiflore RMS2 (Laffort) |
| SH12-S. cerevisiae | SafOEno SH 12 (Fermentis) |
| GV107-S. cerevisiae | SafOEno GV107 (Fermentis) |
| S102-S. cerevisiae | SafOEno BC S102 (Fermentis) |
| CZ-S. cerevisiae | CZ strain (Guizhou Institute of Technology) |
| S103-S. cerevisiae var. bayanus | SafOEno BC S103 (Fermentis) |
| Prelude-Torulaspora delbrueckii | Viniflora Prelude (Chr. Hansen) |
| Pre-CZ | sequential inoculation with Prelude followed by CZ |
References
- Board, E. Flora of China; Science Press: Beijing, China, 2003; Volume 25. [Google Scholar]
- Wang, L.T.; Lv, M.J.; An, J.Y.; Fan, X.H.; Dong, M.Z.; Zhang, S.D.; Wang, J.D.; Wang, Y.Q.; Cai, Z.H.; Fu, Y.J. Botanical characteristics, phytochemistry and related biological activities of Rosa roxburghii Tratt fruit, and its potential use in functional foods: A review. Food Funct. 2021, 12, 1432–1451. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Fang, W.; Wang, Z.; Chen, Y. Physicochemical, biological properties, and flavour profile of Rosa roxburghii Tratt, Pyracantha fortuneana, and Rosa laevigata Michx fruits: A comprehensive review. Food Chem. 2022, 366, 130509. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Z.; Li, Y.F.; Yu, Z.H.; Liu, X.H.; Hardie, W.J.; Huang, M.Z. Screening and characterization of β-glucosidase production strains from Rosa roxburghii Tratt. Int. J. Food Eng. 2020, 17, 1–9. [Google Scholar] [CrossRef]
- Yu, Z.H.; Yue, L.R.; Wang, Q.; Wu, J.S.; Zhong, X.X.; Liu, X.Z.; Huang, M.Z. Identification and brewing potential of a Saccharomyces cerevisiae strain from Rosa roxburghii. F&FI. 2022, 48, 70–75. (In Chinese) [Google Scholar]
- Liu, X.Z.; Li, Y.F.; Yu, Z.H.; Hardie, W.J.; Huang, M.Z. Biodiversity of non-Saccharomyces yeasts during natural fermentation of Rosa roxburghii. Acta Microbiol. Sin. 2020, 60, 1696–1708. [Google Scholar]
- Zhao, H.B.; Li, H.; Tian, Y.; Yu, Z.H.; Liu, X.H.; Huang, M.Z.; Liu, X.Z. Isolation, identification, physiological characteristics and mixed fermentation of a non-Saccharomyces cerevisiae from Rosa roxburghii. Sci. Technol. Food Ind. 2020, 41, 114–120. (In Chinese) [Google Scholar]
- Liu, X.Z.; Zhao, H.B.; Li, Y.F.; Yu, Z.H.; Liu, X.H.; Huang, M.Z. Identification and oenological properties analysis of a strain of Hanseniaspora uvarum from Rosa roxburghii. F&FI. 2020, 46, 97–104. (In Chinese) [Google Scholar]
- Liu, X.Z.; Li, Y.F.; Zhao, H.B.; Yu, Z.H.; Huang, M.Z. Oenological property analysis of selected Hanseniaspora uvarum isolated from Rosa roxburghii Tratt. Int. J. Food Eng. 2021, 17, 445–454. [Google Scholar] [CrossRef]
- Fernandes, T.; Silva-Sousa, F.; Pereira, F.; Rito, T.; Soares, P.; Franco-Duarte, R.; Sousa, M.J. Bio-technological importance of Torulaspora delbrueckii: From the obscurity to the spotlight. J. Fungi 2021, 7, 712. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Ianieva, O.; Podgorsky, V. Enological potential of non-Saccharomyces yeast strains of enological and brewery origin from Ukrainian collection of microorganisms. Mycology 2020, 12, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, M.; Valazquez, R. The yeast Torulaspora delbrueckii: An interesting but difficult-to-use tool for winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Tondinia, F.; Langa, T.; Chena, L.; Herderich, M.; Jiranek, V. Linking gene expression and oenological traits: Comparison between Torulaspora delbrueckii and Saccharomyces cerevisiae strains. Int. J. Food Microbiol. 2019, 294, 42–49. [Google Scholar] [CrossRef]
- Vejarano, R.; Gil-Calderon, A. Commercially available non-Saccharomyces yeasts for winemaking: Current market, advantages over Saccharomyces, biocompatibility, and safety. Fermentation 2021, 7, 171. [Google Scholar] [CrossRef]
- Boudreau, T.F.T.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Free amino nitrogen concentration correlates to total yeast assimilable nitrogen concentration in apple juice. Food Sci. Nutr. 2018, 6, 119–123. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Investigating the biochemical and fermentation attributes of Lachancea species and strains: Deciphering the potential contribution to wine chemical composition. Int. J. Food Microbiol. 2019, 290, 273–287. [Google Scholar] [CrossRef]
- Vigentini, I.; Barrera Cardenas, S.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of native yeast strains for in-bottle fermentation to face the uniformity in sparkling wine production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef]
- OIV. Compendium of International Methods of Wine and Must Analysis; OIV Publication: Paris, France, 2019; Volume 1. [Google Scholar]
- Zhao, T.M.; Ma, C.; Zhu, G.F. Chemical composition and biological activities of essential oils from the leaves, stems, and roots of Kadsura coccinea. Molecules 2021, 26, 6259. [Google Scholar] [CrossRef]
- Huang, M.Z.; Li, T.T.; Hardie, W.J.; Tang, W.W.; Li, X. Comparative characterization and sensory significance of volatile compounds in Rosa roxburghii Tratt fruit from five geographic locations in Guizhou, China. Flavour Fragr. J. 2022, 37, 163–180. [Google Scholar] [CrossRef]
- Moyano, L.; Zea, L.; Moreno, J.; Medina, M. Analytical study of aromatic series in sherry wines subjected to biological aging. J. Agric. Food Chem. 2002, 50, 7356–7361. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Ardanuy, M.; Lopez, R.; Cacho, J.F. Relationship between flavor dilution values and odor unit values in hydroalcoholic solutions: Role of volatility and a practical rule for its estimation. J. Agric. Food Chem. 1998, 46, 4341–4346. [Google Scholar] [CrossRef]
- Giri, A.; Osako, K.; Ohshima, T. Identification and characterization of headspace volatiles of fish miso, a Japanese fish meat based fermenented paste, with special emphasis on effect of fish species and meat washing. Food Chem. 2010, 120, 621–631. [Google Scholar] [CrossRef]
- Fan, W.L.; Xu, Y. Liquor Flavor Chemistry; China Light Industry Press Ltd.: Beijing, China, 2020. (In Chinese) [Google Scholar]
- Moreno, J.A.; Zea, L.; Moyano, L.; Medina, M. Aroma compounds as markers of the changes in sherry wines subjected to biological ageing. Food Control 2005, 16, 333–338. [Google Scholar] [CrossRef]
- Chaves, M.; Zea, L.; Moyano, L.; Medina, M. Changes in color and odorant compounds during oxidative aging of Pedro Ximinez sweet wines. J. Agric. Food Chem. 2007, 55, 3592–3598. [Google Scholar] [CrossRef]
- Escudero, A.; Gogorza, B.; Melús, M.A.; Ortin, N.; Cacjo, J.; Ferreira, V. Characterization of the aroma of a wine from Maccabeo. Key role played by compounds with low odor activity values. J. Agric. Food Chem. 2004, 52, 3516–3524. [Google Scholar] [CrossRef]
- Brennand, C.P.; Ha, J.K.; Lindsay, R.C. Aroma properties and thresholds of some branched-chain and other minor volatile fatty acids occurring in milkfat and meat lipids. J. Sens. Stud. 1989, 4, 105–120. [Google Scholar] [CrossRef]
- Salo, P.; Nykänen, L.; Suomalainen, H. Odor thresholds and relative intensities of volatile aroma components in an artificial beverage imitating whisky. J. Food Sci. 1972, 37, 394–398. [Google Scholar] [CrossRef]
- Moyano, L.; Zea, L.; Moreno, J.A.; Medina, M. Evaluation of the active odorants in Amontillado sherry wines during the aging process. J. Agric. Food Chem. 2010, 58, 6900–6904. [Google Scholar] [CrossRef]
- King, B.M. Sensory evaluation of binary mixtures made from E and Z isomers of 3-hexen-1-ol or E and Z isomers of 3-hexenyl-1-acetate. In Progress in Flavour Research; Adda, J., Ed.; Elsevier Science Publishers: Amsterdam, The Netherland, 1984; pp. 67–80. [Google Scholar]
- Dietz, F.; Traud, J. Geruchsund Geschmacks-Schwellen-Konzentrationen von Phenolkörpern. GWF Wasser Abwasser. 1978, 119, 318–325. [Google Scholar]
- Gemert, L.J.V. Odour Thresholds-Compilations of Odour Threshold Values in Air, Water and Other Media; Oliemans Punter & Partners BV: Amsterdam, The Netherland, 2011. [Google Scholar]
- Zea, L.; Moyano, L.; Moreno, J.; Cortes, B.; Medina, M. Discrimination of the aroma fraction of Sherry wines obtained by oxidative and biological ageing. Food Chem. 2001, 75, 79–84. [Google Scholar] [CrossRef]
- Boulton, R.B.; Singleton, V.L.; Bisson, L.F.; Kunkee, R.E. Principles and Practices of Winemaking; Springer: New York, NY, USA, 2010. [Google Scholar]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Guillamón, J.M.; Mas, A.; Rozès, N. Effects of fermentation temperature and Saccharomyces species on the cell fatty acid composition and presence of volatile compounds in wine. Int. J. Food Microbiol. 2003, 85, 127–136. [Google Scholar] [CrossRef]
- Nicolini, G.; Moser, S.; Roman, T.; Mazzi, E. Effect of juice turbidity on fermentative volatile compounds in white wines. Vitis 2011, 50, 131–135. [Google Scholar]
- Dufour, J.P.; Malcorps, P.H.; Silcock, P. Control of ester synthesis during brewery fermentation. In Brewing Yeast Fermentation Performance, Blackwell Science; Wiley: Hoboken, NJ, USA, 2003; Volume 8, pp. 213–233. [Google Scholar]
- Stipanuk, M.H. Biochemical, Physiological, and Molecular Aspects of Human Nutrition; Elsevier/Saunders: St. Louis, MO, USA, 2013. [Google Scholar]
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Waterhouse, A.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2016; p. 184. [Google Scholar]
- Viegas, C.A.; Rosa, M.F.; Sa-Correia, I.; Novais, J.M. Inhibition of yeast growth by octanoic and decanoic acids produced during ethanolic fermentation. Appl. Environ. Microbiol. 1989, 55, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Fugelsang, K.C.; Edwards, C.G. Wine Microbiology Practical Applications and Procedures; Springer: New York, NY, USA, 2017. [Google Scholar]
- Zhou, Z.; Xu, Y.X.; Hu, H.; Cheng, C.; Pan, S.Y. Comparative application of headspace-solid phase micro-extraction and simultaneous distillation extraction for GC-MS analysis of volatile components in Chestnut Rose (Rosa roxburghii Tratt.) juice. Food Sci. 2011, 32, 279–282. [Google Scholar]
- Linhares, M.F.D.; Alves Filho, E.G.; Silva, L.M.A.; Fonteles, T.V.; Wurlitzer, N.J.; de Brito, E.S.; Fernandes, F.A.N.; Rodrigues, S. Thermal and non-thermal processing effect on açai juice composition. Food Res. Int. 2020, 136, 109506. [Google Scholar] [CrossRef]
- Garcia-Rios, E.; Guillamon, J.M. Sulfur dioxide resistance in Saccharomyces cerevisiae: Beyond SSU1. Microb. Cell. 2019, 6, 527–530. [Google Scholar] [CrossRef]
- An, H.M.; Liu, M.; Yang, M.; Fan, W.G. Analysis of main organic acid compositions in Rosa roxburghii Tratt. Sci. Agric. Sin. 2011, 44, 2049–2100. [Google Scholar]
- Summerson, V.; Gonzalez Viejo, C.; Pang, A.; Torrico, D.D.; Fuentes, S. Assessment of volatile aromatic compounds in smoke tainted Cabernet Sauvignon wines using a low-cost e-nose and machine learning modelling. Molecules 2021, 26, 5108. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.W.; Wang, R.L.; Xiao, Z.B.; Sun, X.X.; Wang, P.P.; Zhu, J.C.; Cao, X.Y. Characterization of volatile compounds of Rosa roxburghii Tratt by gas chromatography-olfactometry, quantitative measurements, odor activity value, and aroma intensity. Molecules 2021, 26, 6202. [Google Scholar] [CrossRef]
- Cordente, A.G.; Nandorfy, D.E.; Solomon, M.; Schulkin, A.; Kolouchova, R.; Francis, I.L.; Schmidt, S.A. Aromatic higher alcohols in wine: Implication on aroma and palate attributes during Chardonnay aging. Molecules 2021, 26, 4979. [Google Scholar] [CrossRef]
- Wu, L.; Wen, Y.D.; Chen, W.Y.; Yan, T.S.; Tian, X.F.; Zhou, S.S. Simultaneously deleting ADH2 and THI3 genes of Saccharomyces cerevisiae for reducing the yield of acetaldehyde and fusel alcohols. FEMS Microbiol. Lett. 2021, 368, fnab094. [Google Scholar] [CrossRef]
- Liu, S.; Laaksonen, O.; Kortesniemi, M.; Kalpio, M.; Yang, B. Chemical composition of bilberry wine fermented with non-Saccharomyces yeasts (Torulaspora delbrueckii and Schiz-osaccharomyces pombe) and Saccharomyces cerevisiae in pure, sequential and mixed fermentations. Food Chem. 2018, 266, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Renault, P.; Coulon, J.; Revel, G.; Barbe, J.-C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Capece, A.; Comitini, F.; Canonica, L.; Siesto, G.; Romano, P. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | |
|---|---|---|
| Saccharomyces cerevisiae | X16 | Laffort, France |
| F33 | ||
| RMS2 | ||
| SafOEno SH 12 | Fermentis, France | |
| SafOEno GV107 | ||
| SafOEno CK S102 | ||
| CZ * | Guizhou Institute of Technology | |
| S. cerevisiae var. bayanus | SafOEno BC S103 | Fermentis, France |
| Torulaspora delbrueckii | Viniflora Prelude | Chr. Hansen, Denmark |
| Parameter | Target Value | Method |
|---|---|---|
| Juice density (Brix) | 24 | Add sucrose |
| YAN * (mg/L) | ~140 | Add Thiazote® (Laffort) |
| pH | 3.6 | Add tartaric acid/Potassium bicarbonate |
| Dilution ratio) | 2 | Add water |
| Suspended solids (NTU) | <100 or 1–2% | Centrifugation |
| Items | Range | ||
|---|---|---|---|
| Yeast Traits for Winemaking | |||
| SO2 tolerance (Relative biomass concentration %) | 60–70 | 71–80 | >80 |
| Score | 1 | 2 | 3 |
| Flocculation (F value) | 71–100 | 31–70 | 0–30 |
| Score | 1 | 2 | 3 |
| H2S production capacity | ≥3 | 1–2 | 0 |
| Score | 1 | 2 | 3 |
| Growth curve (day) | >8–14 | >6–7 | >0–5 |
| Score | 1 | 2 | 3 |
| Fermentation vigor (g CO2/100 mL) | 0–0.60 | 0.61–1.20 | 1.21–1.80 |
| Score | 1 | 2 | 3 |
| Wine Attributes | |||
| pH | 2.0–3.0 | 3.1–4.0 | 4.1–5.0 |
| Score | 1 | 3 | 1 |
| Volatile acidity (g/L) | 0–0.6 | 0.61–1.2 | >1.2 |
| Score | 3 | 2 | 0 |
| Titratable acidity (g/L) | 0–6 | 7–12 | 13–18 |
| Score | 3 | 2 | 1 |
| Residual sugar (g/L) | ≤4 | 4.1–12 | ≥12.1 |
| Score | 3 | 2 | 1 |
| Alcohol (% vol) | 7.0–9.0 | 9.1–11.0 | 11.1–13.0 |
| Score | 1 | 2 | 3 |
| Ascorbic acid (g/L) | 1–2 | 3–4 | 5–6 |
| Score | 1 | 2 | 3 |
| Volatile Components Higher Alcohols (mg/L) | 100–130 | 131–160 | >160 |
| Score | 3 | 2 | 1 |
| Acids (mg/L) | 20–35 | 36–45 | >45 |
| Score | 3 | 2 | 1 |
| Esters (mg/L) | 50–100 | 100–200 | >200 |
| Score | 2 | 3 | 1 |
| Aldehydes (mg/L) | >1 | 0.1-1.0 | <0.1 |
| Score | 3 | 2 | 1 |
| Phenols (mg/L) | 0.1–5 | 5.1–10 | >10 |
| Score | 1 | 2 | 3 |
| Group | Strains | Control * | Relative Biomass Concentration (%) | |||
|---|---|---|---|---|---|---|
| Total SO2 mg/L | ||||||
| 100 | 200 | 300 | 400 | |||
| Saccharomyces cerevisiae | X16 | 1.45 ± 0.01 | 85.69 ± 1.46bc | 56.59 ± 0.71a | 52.94 ± 0.42c | 41.38 ± 0.38b |
| S102 | 1.47 ± 0.01 | 78.70 ± 3.47d | 56.28 ± 0.60a | 52.90 ± 0.92c | 40.85 ± 0.49b | |
| F33 | 1.42 ± 0.02 | 66.79 ± 19.39e | 62.62 ± 18.51c | 59.10 ± 0.26c | 43.43 ± 1.71b | |
| GV107 | 1.41 ± 0.01 | 91.71 ± 0.86abc | 59.04 ± 0.67a | 54.28 ± 0.85c | 41.79 ± 1.29b | |
| CZ | 1.47 ± 0.02 | 90.64 ± 4.01a | 57.70 ± 1.21a | 52.95 ± 0.49c | 41.67 ± 0.90b | |
| RMS2 | 1.43 ± 0.01 | 85.91 ± 0.75cd | 83.83 ± 0.34b | 76.85 ± 1.19a | 42.86 ± 0.37b | |
| SH12 | 1.47 ± 0.02 | 89.16 ± 1.29ab | 56.08 ± 1.31a | 52.84 ± 0.64c | 41.51 ± 1.19b | |
| S. cerevisiae var. bayanus | S103 | 1.49 ± 0.01 | 87.89 ± 1.03ab | 51.19 ± 2.74a | 50.70 ± 0.73c | 40.91 ± 0.36b |
| Torulaspora | Prelude | 1.06 ± 0.01 | 77.50 ± 1.37d | 73.28 ± 1.21b | 64.86 ± 1.42b | 60.75 ± 1.83a |
| Wine | pH | Volatile Acidity g/L | Titratable Acidity g/L | Residual Sugar g//L | Alcohol Concentration %vol | Ascorbic Acid g/L |
|---|---|---|---|---|---|---|
| X16 | 3.65 ± 0.02bc | 0.56 ± 0.08bc | 11.03 ± 0.37 | 82.32 ± 9.26bcd | 10.50 ± 0.55ab | 5.63 ± 0.13ab |
| S103 | 3.64 ± 0.01c | 0.61 ± 0.04bc | 11.46 ± 0.09 | 74.56 ± 1.32d | 10.50 ± 0.55ab | 5.45 ± 0.22ab |
| S102 | 3.65 ± 0.01bc | 0.71 ± 0.10a | 11.54 ± 0.65 | 85.75 ± 1.66c | 9.08 ± 1.02b | 5.36 ± 0.10b |
| F33 | 3.65 ± 0.03bc | 0.34 ± 0.05d | 10.70 ± 0.55 | 75.57 ± 3.71d | 9.43 ± 0.48b | 5.35 ± 0.29ab |
| GV107 | 3.71 ± 0.01a | 0.65 ± 0.06ab | 11.37 ±0.33 | 65.97 ± 6.13de | 10.00 ± 0.02b | 5.60 ± 0.13ab |
| CZ | 3.65 ± 0.04bc | 0.63 ± 0.04b | 11.68 ± 0.31 | 81.51 ± 5.09cd | 9.95 ± 0.61b | 5.55 ± 0.49ab |
| SH12 | 3.66 ± 0.01b | 0.55 ± 0.06c | 11.52 ± 0.25 | 85.78 ± 5.46bcd | 9.13 ± 0.29b | 5.58 ± 0.41ab |
| RMS2 | 3.68 ± 0.02ab | 0.52 ± 0.06c | 11.28 ± 0.14 | 57.47 ± 2.71e | 11.55 ± 0.50a | 5.37 ± 0.09b |
| Prelude | 3.64 ± 0.08abc | 0.56 ± 0.07bc | 11.32 ± 0.72 | 107.45 ± 48.28a | 7.58 ± 3.22c | 5.68 ± 0.14a |
| Pre-CZ | 3.66 ± 0.01b | 0.56 ± 0.08bc | 11.20 ± 0.15 | 91.82 ± 2.60b | 8.78 ± 0.47bc | 5.57 ± 0.30ab |
| Yeast Strain | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | Compounds | CAS | Odor Quality | Odor Threshold (mg/L) | X16 | S103 | S102 | F33 | GV107 | SH12 | RMS2 | CZ | Prelude | Pre-CZ |
| OAV | ||||||||||||||
| Higher alcohols | ||||||||||||||
| 1 | 1-Propanol | 71-23-8 | Fusel, alcoholic | 314 [24] | <1 | nd | <1 | nd | nd | nd | <1 | <1 | nd | <1 |
| 2 | 2-Methyl-1-propanol | 78-83-1 | fusel whiskey | 82 [24] | <1 | 1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| 3 | 1-Penten-3-ol | 616-25-1 | Green, fruity | 0.4 [23] | nd | nd | nd | nd | nd | nd | nd | nd | 28 | nd |
| 4 | 1-Butanol | 71-36-3 | Fusel oil sweet balsam whiskey | 160 [24] | nd | nd | nd | <1 | <1 | nd | nd | nd | nd | nd |
| 5 | 3-Methyl-1-butanol | 123-51-3 | Alcoholic, fruity | 40 [25] | 3 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 6 | 1-Pentanol | 71-41-0 | Fermented, bready, fusel | 0.1502 [26] | 2 | 2 | 2 | 1 | 2 | 3 | 2 | 1 | 2 | 2 |
| 7 | Hexyl alcohol | 111-27-3 | Pungent, fruity, alcoholic | 8 [25] | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| 8 | (E)-3-Hexen-1-ol | 928-97-2 | Green | 1 [24] | nd | nd | nd | nd | <1 | nd | nd | nd | nd | <1 |
| 9 | 3-Ethoxy-1-propanol | 111-35-3 | Fruity | 0.1 [24] | nd | nd | nd | nd | nd | nd | nd | nd | 2 | nd |
| 10 | Leaf alcohol | 928-96-1 | Green, grassy | 1 [24] | 27 | 26 | 26 | 17 | 28 | 25 | 26 | 24 | 22 | 27 |
| 11 | (R, R)-2,3-Butanediol | 24347-58-8 | Buttery, Creamy, Fruity | 0.0951 * [26] | 26 | 17 | 21 | 32 | 31 | 21 | 30 | 20 | 27 | 22 |
| 12 | 2-Nonanol | 628-99-9 | Waxy, green, Creamy | 0.075 [27] | nd | nd | nd | nd | nd | 7 | nd | nd | nd | nd |
| 13 | 1-Octanol | 111-87-5 | Waxy, green, fatty | 10 [24] | <1 | nd | nd | nd | nd | nd | nd | <1 | <1 | <1 |
| 14 | Furfuryl alcohol | 98-00-0 | Musty, sweet | 15 [28,29] | nd | nd | nd | nd | nd | <1 | nd | nd | nd | nd |
| 15 | Phenethyl alcohol | 60-12-8 | Sweet, floral | 14 [25] | nd | <1 | 1 | 2 | 1 | <1 | 1 | <1 | 1 | <1 |
| 16 | Decyl alcohol | 112-30-1 | Fatty, sweet | 0.5 [24] | nd | nd | nd | nd | nd | nd | nd | nd | nd | 2 |
| Acids | ||||||||||||||
| 17 | Acetic acid | 64-19-7 | Vinegar | 300 [30] | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| 18 | Octanoic acid | 124-07-2 | Vegetable, cheesy | 10 [25] | 2 | 2 | 2 | 3 | 2 | 2 | 2 | 3 | 4 | 4 |
| 19 | Decanoic acid | 334-48-5 | Sour, fatty | 0.5 [24] | 2 | 2 | 5 | 34 | 5 | 6 | 4 | 3 | 15 | 8 |
| 20 | 9-Decenoic acid | 14436-32-9 | Waxy, green, fatty | 4.3 [31] | <1 | nd | <1 | nd | nd | <1 | <1 | <1 | 1 | <1 |
| Esters | ||||||||||||||
| 21 | Ethyl acetate | 141-78-6 | Fruity, sweet | 15 [24] | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | 2 | 1 |
| 22 | Isobutyl acetate | 110-19-0 | Sweet, fruity | 3.4 [32] | <1 | nd | nd | <1 | nd | nd | nd | <1 | <1 | <1 |
| 23 | Ethyl propionate | 105-37-3 | Sweet, fruity | 2.1 [24] | nd | nd | nd | nd | nd | nd | nd | nd | <1 | nd |
| 24 | n-Propyl acetate | 109-60-4 | Fusel, sweet, fruity | 65 [28] | nd | nd | nd | nd | nd | nd | nd | nd | <1 | nd |
| 25 | Ethyl butyrate | 105-54-4 | Sweet, fruity | 0.6 [24] | 2 | 3 | 3 | 2 | 3 | 4 | 3 | 3 | 13 | 4 |
| 26 | Isoamyl acetate | 123-92-2 | Sweet, fruity, banana | 0.03 [33] | 804 | 1071 | 626 | 582 | 643 | 614 | 502 | 860 | 2603 | 1300 |
| 27 | Ethyl hexanoate | 123-66-0 | Sweet, fruity, pineapple | 0.014 [25] | 1704 | 1748 | 2729 | 2597 | 2950 | 3118 | 3652 | 3303 | 4993 | 4575 |
| 28 | Hexyl acetate | 142-92-7 | Fruity, green | 1 [29] | 3 | 3 | 3 | 2 | 3 | 4 | 3 | 6 | 10 | 8 |
| 29 | (E)-3-Hexenyl acetate | 3681-82-1 | Sharp fruity, green | 0.87 [34] | 31 | 31 | 24 | 18 | 24 | 27 | 23 | 34 | 58 | 52 |
| 30 | Ethyl octanoate | 106-32-1 | Sweet, musty, fruity | 0.6 [24] | 125 | 95 | 218 | 266 | 210 | 205 | 348 | 253 | 343 | 312 |
| 31 | Ethyl decanoate | 110-38-3 | Sweet, fruity, apple | 0.2 [25] | 25 | 24 | 89 | 329 | 68 | 85 | 126 | 89 | 173 | 97 |
| 32 | 3-methylbutyl octanoate | 2035-99-6 | Sweet, fruity, green | 0.125 [30] | 7 | 5 | 3 | 5 | 4 | 5 | 6 | 6 | 15 | nd |
| 33 | Phenethyl acetate | 103-45-7 | Sweet, honey, floral rosy | 0.25 [33] | 15 | 13 | 19 | 28 | 10 | 7 | 13 | 11 | 32 | 17 |
| 34 | Isoamyl decanoate | 2306-91-4 | Waxy, banana fruity | >5.0 [32] | nd | nd | nd | <1 | nd | nd | <1 | <1 | <1 | nd |
| 35 | Ethyl myristate | 124-06-1 | Sweet, waxy | 0.5 [24] | nd | <1 | 3 | 2 | 1 | 1 | 1 | 2 | 2 | nd |
| 36 | Ethyl laurate | 106-33-2 | Sweet, waxy, floral nuance | 0.5 [24] | nd | nd | 23 | 42 | 18 | nd | 22 | 26 | 42 | 38 |
| 37 | Ethyl palmitate | 628-97-7 | Waxy, fruity, creamy | 1 [24] | nd | nd | nd | <1 | nd | nd | nd | nd | nd | nd |
| Aldehydes | ||||||||||||||
| 38 | Acetaldehyde | 75-07-0 | Pungent, fresh, fruity, musty | 110 [24] | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 | <1 |
| Phenols | ||||||||||||||
| 39 | Butylated hydroxytoluene | 128-37-0 | Mild phenolic camphor | 1 * [35] | <1 | <1 | 2 | <1 | nd | <1 | 18 | 12 | 1 | 20 |
| 40 | Naphthalene | 91-20-3 | Pungent dry tarry | 0.006 * [36] | nd | nd | nd | nd | nd | nd | nd | nd | 13 | nd |
| 41 | Methyl eugenol | 93-15-2 | Spicy, musty, vegetative | 10 [37] | nd | nd | nd | nd | nd | nd | nd | nd | <1 | nd |
| 42 | 2-Ethoxynaphthalene | 93-18-5 | Powder, floral | 0.1 [24] | nd | nd | nd | 7 | nd | nd | nd | nd | 4 | 11 |
| Yeast Attributes | Wine Attributes | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Standard Parameters | Headspace Volatiles | ||||||||||||||||
| Yeast Strains | Sulfite Tolerance | Flocculation | H2S Production Capacity | Growth Curve | Fermentation Vigor | pH | Volatile Acidity | Titratable Acidity | Residual Sugar | Ethanol | Ascorbic Acid | Higher Alcohols | Acids | Esters | Aldehydes | Phenols | Total Score |
| X16 | 3 | 3 | 2 | 3 | 3 | 3 | 3 | 2 | 1 | 2 | 3 | 2 | 3 | 3 | 3 | 1 | 40 |
| S103 | 3 | 3 | 2 | 3 | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 1 | 3 | 3 | 3 | 1 | 38 |
| S102 | 2 | 3 | 2 | 3 | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 2 | 3 | 1 | 3 | 1 | 36 |
| F33 | 1 | 3 | 1 | 3 | 3 | 3 | 3 | 2 | 1 | 2 | 3 | 3 | 1 | 1 | 3 | 1 | 34 |
| GV107 | 3 | 3 | 2 | 3 | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 2 | 3 | 1 | 3 | 1 | 37 |
| CZ | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 3 | 2 | 1 | 3 | 3 | 40 |
| SH12 | 3 | 3 | 3 | 3 | 2 | 3 | 3 | 2 | 1 | 2 | 3 | 1 | 3 | 1 | 3 | 1 | 37 |
| RMS2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 1 | 3 | 3 | 2 | 3 | 1 | 2 | 3 | 41 |
| Prelude | 2 | 3 | 1 | 1 | 1 | 3 | 3 | 2 | 1 | 1 | 3 | 3 | 1 | 1 | 2 | 1 | 29 |
| Pre-CZ | - | - | - | 2 | 1 | 3 | 3 | 2 | 1 | 1 | 3 | 2 | 1 | 1 | 3 | 3 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.-H.; Huang, G.-D.; Huang, X.-Y.; Pu, J.-H.; Wu, J.-S.; Yue, L.-R.; Hardie, W.J.; Liu, X.-Z.; Huang, M.-Z. A Comparative Study of Yeasts for Rosa roxburghii Wine Fermentation. Fermentation 2022, 8, 311. https://doi.org/10.3390/fermentation8070311
Yu Z-H, Huang G-D, Huang X-Y, Pu J-H, Wu J-S, Yue L-R, Hardie WJ, Liu X-Z, Huang M-Z. A Comparative Study of Yeasts for Rosa roxburghii Wine Fermentation. Fermentation. 2022; 8(7):311. https://doi.org/10.3390/fermentation8070311
Chicago/Turabian StyleYu, Zhi-Hai, Gui-Dan Huang, Xiao-Yan Huang, Jiang-Hua Pu, Jia-Sheng Wu, Li-Rong Yue, William James Hardie, Xiao-Zhu Liu, and Ming-Zheng Huang. 2022. "A Comparative Study of Yeasts for Rosa roxburghii Wine Fermentation" Fermentation 8, no. 7: 311. https://doi.org/10.3390/fermentation8070311