Yeast Flocculation—Sedimentation and Flotation †

The International Centre for Brewing and Distilling, Heriot-Watt University, Riccarton, Edinburgh EH144AS, UK
†
Some of the information included in this review paper was obtained (with the permission of the publisher) from: Stewart, G.G. Harvesting and cropping yeast: Flocculation and centrifugation. Brewing and Distilling Yeasts; Springer International Publishing, Cham, Switzerland, 2017; pp. 259–308.
Fermentation 2018, 4(2), 28; https://doi.org/10.3390/fermentation4020028
Submission received: 8 March 2018
/
Revised: 9 April 2018
/
Accepted: 10 April 2018
/
Published: 16 April 2018
(This article belongs to the Special Issue Brewing & Distilling)
Abstract
:Unlike most fermentation alcohol beverage production processes, brewers recycle their yeast. This is achieved by employing a yeast culture’s: flocculation, adhesion, sedimentation, flotation, and cropping characteristics. As a consequence of yeast recycling, the quality of the cropped yeast culture’s characteristics is critical. However, the other major function of brewer’s yeast is to metabolise wort into ethanol, carbon dioxide, glycerol, and other fermentation products, many of which contribute to beer’s overall flavour characteristics. This review will only focus on brewer’s yeast flocculation characteristics.
1. Introduction
The objectives of brewer’s wort fermentations are to consistently metabolise wort constituents into ethanol, carbon dioxide, glycerol, and other fermentation products, many of which contribute to beer flavour in order to produce it with satisfactory quality, drinkability, and stability. Another important fermentation objective is to produce yeast crops that can be harvested, stored, and subsequently re-pitched into a later brew [1]. During brewing, overall yeast performance is controlled by a plethora of factors that include (in no order of priority):
- The yeast strains employed and their condition during pitching and throughout the wort fermentation cycle [2];
- the concentration and category of assimilable nitrogen [3];
- the spectrum of wort sugars [4];
- fermentation temperature [2];
- yeast pitching (inoculation) rate [1];
- concentration of inorganic ions [5];
- yeast tolerance to stress factors such as: heat, osmotic pressure, temperature, ethanol, mechanical tension, desiccation, etc. [6];
- wort gravity [7];
- wort dissolved oxygen concentration at pitching and during fermentation [8];
- flocculation, adhesion, sedimentation, and cropping characteristics [9];
- culture characteristics during storage between fermentations and its subsequent re-pitching into wort [10].
Many of the above factors will be discussed in other parts of this monograph. They influence yeast performance either individually or in combination with other factors. Together, they permit definition of the requirements of an acceptable brewer’s yeast strain [10]: “in order to achieve beer of high quality, it is axiomatic that not only must the yeast be effective in removing the required nutrients from the growth/fermentation medium (the malt extract—the wort), able to tolerate the prevailing environmental conditions (for example, ethanol toxicity) and impart the desired flavour and stability to the beer, but the microorganisms themselves at the end of fermentation must be effectively removed from the wort by flocculation (Figure 1), sedimentation, centrifugation (Figure 2), and/or filtration after they have fulfilled their metabolic role.”
It is also worthy of note that brewing is the only major alcoholic beverage process that recycles its yeast [2]. It is, therefore, important to jealously protect the quality of the cropped yeast because it will be used to pitch subsequent wort fermentations and will therefore have a profound effect on the quality of the beer resulting from subsequent wort fermentations.
This review initially examines brewer’s yeast flocculation, including both nature and nurture aspects [11]. It subsequently analyses other aspects of yeast’s sedimentation and cropping characteristics during brewing. In addition, yeast flocculation and adhesion, as they apply to other industrial processes [3,12] and medical aspects [13], are briefly discussed.
The flocculation properties of brewer’s yeast strains are of considerable importance because the number of yeast cells suspended in wort during both primary and/or secondary fermentations affects wort fermentation velocity, beer flavour, maturation, cell crop formation, and filtration [14]. A number of hypotheses have been proposed to explain the mechanisms of flocculation in Saccharomyces cerevisiae [9]. The prevalent hypothesis is the lectin-like theory of flocculation [15]. According to this model, a specific protein (lectin) present only in flocculent yeast cells is secreted into the outer extremities of the cell wall and the N-terminal part of this protein binds two mannose residues present in the cell walls of neighbouring flocculent and non-flocculent yeast cells; details of this follow [16] (Figure 3). In this process, calcium ions are necessary for the activation of the lectins [17].
2. Yeast Flocculation
Flocculation is a complex phenomenon that is affected by genetic [16], physiological [9], and environmental factors—the nature-nurture effects [11]. Within the two brewer’s yeast species (Saccharomyces cerevisiae (ale) and Saccharomyces pastorianus (lager)) [17,18] there are both flocculent and non-flocculent strains (Figure 1). These characteristics are illustrated by the concentration of yeast in suspension during the course of a static wort fermentation [1] (Figure 1) and overall wort sugar uptake properties [19].
The reasons for, and advantages of, yeast flocculation (particularly, but not only, during the brewing process) have been extensively studied and debated. Flocculation may enhance the survival of yeast cells during adverse (for example, starvation) conditions [20]. Flocculation is an important characteristic in an environment with limited nutrients because the death and autolysis of the cells inside flocs can provide further nutrients to cells in the surrounding environment [21]. It has been shown that some highly flocculent cultures lost their flocculation characteristic during the early stages of growth, in the presence of nutrients, and recovered it towards the end of the exponential growth phase coinciding with nutrient depletion [22].
Individual strains of brewer’s yeast (both ale and lager) differ considerably in their flocculating characteristics. At one extreme, there are highly non-flocculent strains, often referred to as powdery strains [23]. At the other extreme, there are very flocculent strains (Figure 4A). The latter strains tend to separate early out of suspension from fermenting wort, which can result in an under-attenuated, sweeter, and less fully fermented beer. Beers of this nature, because of the presence of fermentable sugars (usually maltotriose and maltose), can be liable to biological instability. In contrast, poorly flocculent (non-flocculent or powdery) yeasts (Figure 4B) produce a dry yeast, fully fermented, more biologically stable beer in which clarification is slow, leading to filtration difficulties and the possible acquisition of yeasty off-flavours.
The disadvantages presented by the two flocculation types of brewer’s yeast strains are particularly relevant to traditional fermentation systems where the fermentation process is dependent upon the yeast sedimentation characteristics. Contemporary brewing technology has largely reversed this situation, as yeast sedimentation characteristics are now applied to fermenter design. The efficiency, economy, and speed of batch fermentations have improved with the use of cylindro-conical fermentation vessels [24] (Figure 5), which are often (but not always) employed in tandem with flocculent cultures; details of this are discussed later. There is no doubt that differences in the flocculation characteristics of various yeast cultures are primarily manifestations of the culture’s cell wall structure, as also discussed later.
Flocculation requires the presence of cell surface protein and mannan receptors (Figure 3). If these are not available or are masked, blocked, inhibited, or denatured, flocculation cannot occur. The onset of yeast flocculation is an aspect of the subject where there is great commercial interest but about which relatively little is known. As previously discussed, the ideal brewing culture remains in suspension as fermenting single cells or very small flocs until the end of fermentation when wort sugars (sucrose, glucose, fructose, maltose and maltotriose) are depleted by the yeast and the vicinal diketones (VDKs) (unwanted diacetyl—butterscotch flavour) are managed appropriately [25]; only then does the culture flocculate out of suspension. What signals the onset of flocculation or relief from its inhibition? This is still an unanswered question that is currently being studied by a number of research laboratories worldwide [9,10,14,25,26,27].
Brewer’s yeast flocculation is an off-cost process which does not require significant energy input. However, cooling at the end of fermentation does facilitate yeast separation and this normally requires some energy. It is considered that flocculation increases process efficiency and reduces the energy consumption associated with cell separation with, for example, the use of a centrifuge—details of this follow. Indeed, yeast flocculation in brewing is an example of Lean Manufacturing [7].
3. Measurement of Yeast Flocculation
Although the requirement for yeast flocculation during brewer’s wort fermentation is a much-studied phenomenon, there is dire need for some degree of flocculation test standardisation. There are a plethora of flocculation tests and many laboratories working in this field appear to employ their own favourite method. As a consequence, it is difficult to translate results from one laboratory to another. The methods used to measure flocculation can be roughly divided into three groupings, as listed below.
3.1. Sedimentation Methods (For Example, the Helm Sedimentation Test)
In this test, the yeast culture is removed from the growth medium and the cells are washed a number of times with deionised water containing calcium ions (usually 80 mg/mL calcium chloride) at pH 4.0 and, depending on the scale of the test, the suspension is placed in a test tube (10-mL scale) or a measuring cylinder (100-mL scale) (Figure 6A shows results of the Helm’s Sedimentation Test [28] for co-flocculation—details to follow).
3.2. Direct Observation of Floc Formation in the Growth/Fermentation Medium
During this method, small inocula of the yeast strain’s being studied are seeded into 20-mL screw capped glass bottles each containing 15 mL of fermentation media. After three days of incubation (usually at 25 °C), the flocculation characteristics of the cultures are determined by the ratio of the flocs subsequent to the yeast sediment being brought back into suspension by shaking the bottles. This method permits routine flocculation determinations of a large number of cultures and has been employed extensively for segregation studies on flocculation during FLO gene mapping studies [30]—details to follow. An alternative measurement of flocculation is to microscopically examine the flocs and determine the percentage of cells in a number of flocs compared to un-flocculated cells.
3.3. Static Fermentation Methods
In this method, the concentration of yeast in suspension is determined during the course of static fermentation (Figure 1). The first two methods for measuring yeast flocculence can be viewed as artificial tests for flocculation due to the fact that they are conducted under in vitro conditions in relation to the brewing process. The latter method is an in vivo style test because it is carried out under conditions more closely akin to the static fermentation conditions encountered in a typical brewery [31] (Figure 6B).
4. Cell Wall Characteristics
Yeast cell surface morphology differences between flocculent and non-flocculent brewer’s yeast cultures (another yeast species) have been extensively examined [32]. Although details of yeast cell wall structure will be considered later, it is worth discussing here that the wall consists of an inner layer composed predominantly of β-glucan and chitin together with an outer fibrilar layer on flocculent cells (Figure 7) [33], consisting primarily of α-mannan (highly glycosylated) associated with mannoproteins [34] (Figure 8).
In addition to cell-cell adhesion in the formation of flocs (Figure 3), some yeast cells (both brewing and distilling strains) possess the capacity to adhere to abiotic (non-living) surfaces, cells and, tissues, resulting in biofilm formation [12] (Figure 9). The cell wall functions as a means for yeast cells (and other microorganisms) to interact with their environment. It has already been discussed that one of the most critical functions of some cell surfaces is their ability to adhere to other cells (floc formation) and surfaces. Adhesion prevents cells from being washed away when they are present in a nourishing environment, while it also allows them to form biofilms that protect the cells from hazardous and stressful conditions [35]. In addition to an industrial environment, pathogenic yeasts (for example, Candida albicans) exploit their capacity to adhere to abiotic surfaces such as plastic prosthetics in order to gain access to the bloodstream and the internal organs of patients [36]. It is believed that adhesion of C. albicans cells to mucosal surfaces involves lectin-like interaction between the protein portion of mannoproteins located in the fibrils on the cell surface and glycoside receptors on epithelial cells [13] (Figure 10).
The word floc derives from the Latin word floccus, which means a tuft of wool, while cells that are not able to form flocs are known as non-flocculent or powdery cells [37]. S. cerevisiae can be found aggregated in different ways not to be confused with flocculation, such as sexual aggregation [38], co-flocculation—addressed later in this paper [29] (Figure 6A and Figure 11), biofilm formation [12] (Figure 9), and chain (pseudohyphae) formation, where daughter cells do not separate physically from their mother cells [39] (Figure 12).
5. Sexual Aggregation
Sexual aggregation between haploid strains of S. cerevisiae involves complementary mating types (MATα and MATa) which can occur after an exchange of pheromones (an agent secreted by an organism that produced a change in the sexual behaviour of a related organism)—a and α, respectively. This induces the appearance of complementary molecules (proteins) on the surface of the cells which facilitates the fusion of haploid cells—mating a cells with α cells [42]. Co-flocculation [29], mutual aggregation [43], mutual agglutination [44], and mutual flocculation [40] all describe a heterotypic aggregation process (while single-strain flocculation is homotypic) between two microbial strains which are usually (not always) yeast [45] (Figure 6A and Figure 11). When two yeast strains (one being non-flocculent and the other weakly flocculent) are mixed together, in the presence of Ca2+ ions at pH 4.0–4.5, flocs form, which rapidly settle out of suspension or rise to the surface of a wort fermentation [46] (Figure 6A). Details of co-flocculation can be found in Section 8.
It has already been discussed that chain formation (also called pseudohyphal growth) occurs due to failure of a young daughter yeast cell to separate from the mother cell. This results in an aggregate, usually composed of 30–50 cells [12] (Figure 12). These cell aggregates are physically linked to each other and, consequently, following dispersion of the cells (for example, physical disruption, sonication, or enzyme treatment), cells will not be able to re-aggregate prior to a subsequent growth cycle. Chain formation in brewing yeast strains is restricted to ale yeast species (S. cerevisiae); lager yeast strains (S. pastorianus) do not usually exhibit chain formation characteristics [47]. Various effects on yeast flocculation and aggregation are discussed in greater detail later.
6. Yeast Cell Wall Structure and Flocculation
It has already been stated that flocculation is a cell surface characteristic [32]. Some (not all) heat-treated flocculent cells retain their ability to flocculate [26], as well as isolated in vitro cell walls prepared from a flocculent culture [48]. The yeast cell wall carries a negative charge due to the ionisation of carboxyl and phosphodiester groups in cell wall proteins and phosphomannans, respectively. The repulsion of charges of the same ionisation sign prevents cells from getting sufficiently close and acts as an effective barrier to cell aggregation [49]. Also, a positive correlation between cell surface hydrophobicity (CSH) and flocculation has been reported [50]. CSH is partially responsible for triggering flocculation in brewing yeast strains [51]. Based on these results, an increase in yeast surface hydrophobicity has been described when dominant flocculation genes (FLO genes) (details to follow) are expressed in the structure of yeast cell walls [27].
It has been proposed that lectins (carbohydrate-binding proteins) are macromolecules that are specific for sugar moieties [52]. They are considered to be present on flocculent cell surfaces. Specific lectin-like proteins interact with carbohydrate residues for α-mannans (receptors) on neighbouring cells (Figure 7). Also, calcium ions enable lectins to achieve their active conformation [53] (details of calcium effects on yeast flocculation are discussed later). Although flocculation lectins are only present on the surface of flocculent cells, the receptors are present in both flocculent and non-flocculent cells since, as already discussed, and the outer layers of the S. cerevisiae cell wall are in part composed of mannan (Figure 8). Analysis of the inhibitory action of sugars and the use of mannan synthesis mutants and concanavalin A (a lectin originally extracted from the jack-bean) have shown that the wall specifically consists of glycoproteins with mainly α-d-mannosyl and α-d-glucosyl groups [54]. This supports the theory that flocculation receptors are most likely nonreducing residues [55]. Besides specific lectin sugar interactions, other non-specific interactions such as hydrogen bonds and hydrophobic interactions can occur.
7. Factors Affecting Yeast Flocculation
Several factors can affect yeast flocculation [56]. The first factor is the genetic background of a strain. Flocculin proteins are encoded by members of the FLO group of genes [23,56]. The genetic background (nature effects) with regard to FLO genes varies greatly amongst various types of brewer’s yeast (ale or lager) and other strains. Strains contain a variety of FLO gene combinations [57], resulting in a spectrum of flocculation characteristics [58]. The current status with respect to S. cerevisiae (and S. pastorianus) FLO genes are discussed in Section 9.
Second, flocculation is affected by the physiological environment (nurture effects) [59]. This includes the pH, availability of appropriate metal ions, and nutrients during the growth phase [60]. The pH will influence the cell surface charge which will have an effect on the flocculation phenotype. Changes in pH may also modify the ionisation of functional groups in flocculin proteins, which will modify their conformation [21]. The environment is sensed by cells leading to the expression of FLO, their translation to Flo proteins, and their location into the cell wall, which is influenced by a number of environmental factors [27].
Thirdly, the physical environment affects flocculation. The hydrodynamic (relevant to liquids in motion) conditions must be favourable and promote collision rates between cells. However, agitation must not be sufficiently violent to cause flocs to disperse. In addition, there must be sufficient cells in suspension in order to result in cell collisions to form flocs [61]. Factors that increase cell walls’ hydrophobic character (cell surface hydrophobicity) and factors that decrease the repulsive negative electrostatic charges on cell walls (cell surface change) result in stronger flocculation, because they facilitate cell-cell contact [62].
Flocculation, as such, is not absolutely necessary for brewer’s yeast cells to sediment out of wort at the end of primary fermentation. This is because the size and density of yeast cells can overcome the Brownian motion that maintains cells in suspension [26]. However, under these circumstances, the sedimentation rate is slow, which is dependent on particle size. Smaller particles settle more slowly than larger particles of similar density, because they are more retarded by friction (overall viscosity). Older (larger) yeast cells settle faster than younger, smaller cells. Nevertheless, the sedimentation of single cells is too slow to be of practical importance during wort fermentation, where flocculation is necessary to achieve the sedimentation of most yeast cultures during the latter stages of a typical fermentation cycle.
Flocculating yeast cultures contain some cells that are not entrapped in flocs which do not necessarily have the ability to flocculate [63]. Single cells are set free from flocs and, at the same time, new cells are entrapped. Loosening of cells occurs from flocs, because of hydrodynamic forces, resulting in a dynamic equilibrium [64]. Dilution favours an increased proportion of free cells. An appropriate concentration of yeast cells is required for effective flocculation and some cells often remain in suspension even following the bulk sedimentation of yeast flocs. As with many aspects of brewer’s yeast metabolism, yeast flocculation is primarily strain-dependent.
During the later stages of fermentation, the following conditions are favourable for culture sedimentation:
- The carbon dioxide production rate is slow;
- wort attenuation is approaching completion—most of the fermentable sugars in wort have been removed by the yeast culture, including glucose, fructose, sucrose, maltose, and finally maltotriose [4];
- flocculation ability is high but not too high;
- yeast concentration in suspension is maximal [61];
There are several factors that influence the rate of floc sedimentation out of the wort:
- The way that yeast cells are packed into flocs;
- the floc size, shape, and density;
- higher gravity worts, following fermentation, results in “green” (immature) beers with a higher viscosity and density. Both these factors will retard yeast sedimentation and lead to increased osmotic pressure and ethanol prior to dilution to the fermented wort’s sales gravity and alcohol concentration.
The floc size often decreases as settling proceeds because the concentration of yeast cells in the suspension continues to decrease and smaller flocs form which will slowly settle out of the suspension.
8. Co-Flocculation
Co-flocculation, also known as mutual aggregation or mutual flocculation, is a heterotypic aggregation process (while flocculation is homotypic) amongst two separate yeast strains [67]. One strain is non-flocculent and the other strain is weakly flocculent. When these strains are mixed together in the presence of Ca2+ ions, flocs form and the culture rapidly settles out of suspension [29]. Heterotypic yeast non-sexual flocculation was first described by Eddy and Rudin [68] with a number of ale strains. The focus of our research group on co-flocculation began with the top-cropping Labatt ale yeast culture. In the 1960s and 1970s, consumption of pale ales was popular in Canada; in 1970 it represented 60% of the beer consumed in Ontario and 80% in Quebec [69]. However, similar to the situation in Britain, the ale yeast cultures employed (unlike lager strains) were (and many still are) largely uncharacterised. Also, no Labatt employee (or Labatt pensioner) could inform us much about the history of their ale culture. This ale culture possessed classical top-cropping properties. It also exhibited intermittent premature flocculation characteristics, resulting in under-fermented worts containing residual sugars (mainly maltotriose). This was a problem in Canada because the alcohol specification was a legal regulation (Federal Health Protection Board Regulations) and the beer’s alcohol composition was (and still is) specified on the label of the bottle, can, or keg with a ±0.2% (v/v) alcohol specification permitted. This problem was exacerbated during high gravity brewing trials [69] of ales. Consequently, it was important to enumerate the number of strains in this ale culture and characterise them.
A suitable method to examine yeast culture’s strain composition in the 1960s and 1970s was the giant colony morphology method initially developed by Gilliland [70] and Richards [71]. The Richard’s method involved inoculating the yeast culture in question onto wort solid media and examining the colonial morphology that developed after incubating under humid conditions for at least three weeks at 18 °C. It was found that gelatin, as the solidifying matrix, tends to enhance the distinctive features of the colonial morphology to a greater extent than agar and that wort, instead of a synthetic or defined medium, gave distinctive and reproducible results (Figure 13). Also, lager yeast strains do not exhibit distinctive colonial morphologies on wort gelatin media (Figure 13).
Analysis of the Canadian brewery’s top cropping ale yeast culture’s strain composition showed that two morphologically different colony types were present (Figure 13A,B) [72]. On isolation, both colony types proved to be stable respiratory sufficient individual yeast strains of the species S. cerevisiae and they were coded LAB A/69 and LAB B/69, with the former strain being ~75% of the ale culture and the latter comprising ~25% of the culture. A production-scale fermentation trial with ale strain LAB A/69 was conducted in a 200-hL (20,000 L) open wood fermenter with a 12 °Plato wort at 21 °C. Although the fermentation was under-pitched (under-inoculated), it proceeded rapidly, and all of the wort’s fermentable sugars were metabolised in less than 96 h. It was then that problems began. The expected top-crop failed to develop on the fermentation’s surface (which occurred in the original two strain ale mixed culture), and most of the yeast culture remained in suspension. As the brewery in question (at that time) did not possess a centrifuge, it was not possible to collect the yeast for reuse in a subsequent fermentation and the fermented wort (all 200 hL of it) had to be discarded into the sewer with the cost charged to the brewery’s effluent budget.
Following the above traumatic brewing-scale trial, a laboratory-scale characterisation of both ale yeast strains isolated from the Labatt ale cultures was conducted. When the two isolated strains were cultured alone in wort using 2-L glass cylinders, both were non-flocculent during all phases of growth. However, when cultured together in wort in a 1:1 ratio, the culture was flocculent in the later stages of fermentation and sedimented out of suspension [72]; a top crop also formed (Figure 6B). This type of behaviour, where two yeast strains are non-flocculent alone but flocculent when mixed together [69], has been termed co-flocculation. As mentioned above, co-flocculation has also been termed mutual aggregation and mutual flocculation [44]. When stationary phase cells of the two strains were mixed together in a 1:1 ratio in a solution of calcium ions at pH 4.5 [29], flocs immediately began to appear and a very flocculent culture resulted that sedimented out of suspension (Figure 6A). This is the Helm’s Sedimentation Test [28].
Protein-denaturants (for example, urea and guanidine) and several sugars (mannose, glucose, fructose, and galactose) cause the reversible inhibition of co-flocculation in the presence of Ca2+ ions. Also, the effect of cell treatment with proteolytic enzymes (trypsin and chymotrypsin) and chemical modification of the cell surface protein and carbohydrate components suggests that co-flocculation results from an interaction between surface protein cellular components of one of the strains and surface carbohydrate components of the other strain [43].
To date, co-flocculation has only been observed with ale strains [29]. There are no reports (including extensive studies in the Labatt research laboratories) of co-flocculation between non-flocculent lager yeast strains. Another type of co-flocculation reaction that has been described occurs when an ale yeast strain has the ability to aggregate and co-sediment with contaminating bacteria such as Hafnia protea [73], Lactobacillus brevis [45], Pediococcus sp., or Lactobacillus fermentum [40] (Figure 11). This Lactobacillus fermentum strain was isolated from a fuel alcohol molasses fermentation in Brazil and its co-flocculation characteristics were studied in Canada.
The two-strain composition of the Labatt co-flocculent production ale culture was deemed to be undesirable, particularly because of its tendency for premature flocculation and the consequent wort under-attenuation that occurred. This was particularly the case with high gravity worts [7] (>14 °Plato). This often resulted in the failure to comply with the beer’s alcohol specification (this was prior to the introduction of high gravity brewing procedures on a production basis, which exacerbated the problem). Production trials with the LAB A/69 ale strain were conducted in both regular (sales) (12 °Plato) and higher gravity (16 °Plato) worts. This strain proved to be capable of successfully fermenting both wort gravities but, because of its non-flocculent property, centrifugation was required in order to harvest the culture for yeast removal, beer clarification, and yeast collection for reuse. This strain has been employed for ale production by Labatt with high gravity worts for the past 40 years and was one of the reasons for the introduction of centrifuges into Labatt plants (further details on the use of centrifuges and their effect on brewer’s yeast strains are given later in this paper).
Mortier and Soares [67] studied co-flocculation from a different perspective. They viewed co-flocculation as a process to separate non-flocculent yeast cells from a fermentation medium. The possibility of this process being employed for the cell separation of different yeast species was assessed. The fission yeast Schizosaccharomyces pombe was used as a control, since these cells are unable to be aggregated by flocculent cells of S. cerevisiae, due to a lack of compatible receptors with S. cerevisiae flocculation lectins [56]. However, the yeast Kluyveromyces marxianus can exhibit co-flocculation, and consequently separate this culture from suspension by settling using flocculent cells of S. cerevisiae to enhance it. The different degrees of co-flocculation amongst non-flocculent strains were the consequence of the different composition and structure of the yeast cell wall, particularly subtle differences in its detailed mannan architecture.
Recent studies by Rossouw et al. [74] examined the relevance of different Flo proteins in mediating differential interspecies aggregation in a “natural” yeast ecosystem in the wine industry. This publication reports on co-flocculation behaviour in mixed cultures of S. cerevisiae and non-Saccharomyces yeast cultures. It is suggested that adhesion phenotypes and, in particular, Flo proteins, may play roles in their system dynamics beyond the roles previously assigned to these proteins. Further elucidation of the molecular mechanisms interpinning different co-flocculation yeast behaviours will be of fundamental importance to understand the role of direct cell-cell interactions between different species in a shared environment.
Cell adhesion phenotypes are complex, and are influenced not only by genetic factors which determine the composition and properties of the cell wall, but also by a number of environmental parameters which impart adhesion behaviour. The observations, under controlled laboratory conditions, therefore presents a simplified view of interspecies co-flocculation as discussed in this review. The data provides, for the first time, genetic insights into the phenomenon of co-flocculation in S. cerevisiae.
Future studies should include additional yeast species and strains under various experimental conditions in order to comprehensively investigate the yeast co-flocculation concept. In addition, the relevance of yeast species adhesion phenotypes to microbial interactions in natural ecosystems requires further detailed investigation [75,76].
9. Genetic Control of Yeast Flocculation
Genetic studies on yeast flocculation began over 70 years ago. The first publication on this subject was by Pomper and Burkholder [77], who reported crossing a haploid culture that possessed a “dispersed” character (non-flocculent) with a haploid culture that possessed a non-dispersed character (flocculent). The “dispersed” character was reported to be dominant over the non-dispersed character. In the early 1950s, Gilliland (working in the Guinness laboratory in Dublin) and Thorne (working in the Alfred Jorgensen laboratory in Copenhagen) independently carried out extensive studies on the genetics of yeast flocculation. Gilliland [17] studied two non-brewing strains of S. cerevisiae which differed only in their flocculation properties. It was proposed that a single gene was responsible for the flocculent phenotype. Thorne [18] confirmed Gilliland’s studies and demonstrated that yeast flocculence was an inherited characteristic and that the flocculence phenotype was dominant over the non-flocculent characteristic. These studies were also confirmed by Guo et al. [41].
It has already been discussed that genetic studies, involving brewer’s yeast strains, is fraught with difficulties because of their frequent triploid, polyploid, or aneuploid ratios [78]. It was confirmed that this yeast characteristic is controlled by dominant genes that were termed FLO genes [79]. FLO genes from S. cerevisiae resemble the EPA (Epithelial Adhesin) gene family from the opportunistic pathogenic yeast species Candida glabrata [80] and the ALS genes (agglutinin-like sequence) from Candida albicans [81]. C. glabrata encodes a family of at least 23 ETA genes, which encode cell surface proteins capable of mediating adherence to epithelial cells.
The first flocculation gene from S. cerevisiae to be studied in detail was FLO1 [79]. The early studies focussed on laboratory haploid and diploid flocculent and non-flocculent strains. The FLO1-containing haploid strain studied (coded 169) was a MATα mating type opposite to that of a non-flocculent haploid strain (coded 168-MATα). These two strains were mated using micromanipulation techniques and found to be flocculent, confirming previous findings that the flocculent character (coded by FLO1) was dominant and stable [82].
The mapping of the FLO1 gene employed classical gene mapping techniques (mating, sporulation, micromanipulation, tetrad analysis, spore germination, multiple flocculation tests, etc.) according to the methods discussed by Sherman and his colleagues [83]. Since the successful mapping of FLO1, novel genetic techniques have been developed, the principal of which began with the sequencing of the Saccharomyces genome [82,84] with the haploid yeast strain S288C. This has significantly expanded our knowledge regarding the genetic control of yeast flocculation. As a result of the reversible inhibition of flocculation by sugars, salt, a low pH environment, or protease sensitivity, two main flocculation phenotypes have been distinguished: Flo1 and NewFlo phenotypes [55]. The Flo1 phenotype includes strains in which flocculation is specifically inhibited by the monosaccharide mannose and its derivatives. The NewFlo phenotype contains the majority of brewing ale strains. In this phenotype, flocculation is reversibly inhibited by mannose, maltose, glucose, and sucrose, but not galactose. NewFlo phenotype strains are more sensitive to inhibition by cations, low pH conditioning, and digestion by trypsin or proteinase [22]. Also, phenotypes display different sensitivities to culture conditions such as temperature [9], pH [85], ions, and nutrient availability [86].
It has already been discussed that there are a number of dominant and recessive genes as well as a flocculation actuator and suppressor genes. The genes that encode flocculation lectins are called FLO genes. FLO1 is the most studied [87]. There are at least nine FLO genes—FLO1, FLO5, FLO8, FLO9, FLO10, FLO11, FLONL, FLONS, and Lg-FLO in both S. cerevisiae and S. pastorianus strains that encode flocculin proteins (Table 1). The flocculin encoded by the FLO11 gene differs from the others in that it is involved in filamentous growth (pseudomycelia), adhesion to solid surfaces, and flor formation rather than flocculation per se [88]. Another FLO gene, FLO8, encodes for a transcription factor regulating the expression of other FLO genes [89]. Many yeast strains usually possess several FLO genes [90]. For example, it has already been briefly described that the first completely sequenced haploid strain—S288C [82]—contains six FLO genes (FLO1, FLO5, FLO8, FL010, and FLO11) as well as four non-functional FLO pseudogenes [90]. The amino acid sequences of Flo5, Flo9, and Flo10 proteins are 96%, 94%, and 58% identical, respectively, to Flo10p. As expected, Flo11p is the most distantly related to other flocculins, being only 37% identical to Flo1p [27].
The DNA sequences of FLONL and FLONS are very similar to that of FLO1, but compared to this gene they have lost some of the internal tandem repeats. More tandem repeats were lost in FLONL than in FLONS. The deletion of these repeats appears to have converted the flocculation phenotype from Flo1 to the NewFlo phenotype [91]. The NewFlo phenotype includes the majority of brewing ale yeast strains. In this phenotype, flocculation is reversibly inhibited by mannose, maltose, glucose, and sucrose, but not galactose [26,55]. Moreover, the NewFlo flocculation phenotype conferred by FLONS and FLONL is also inhibited by galactose [92], whereas the usual NewFlo phenotype is insensitive to galactose. This indicates that the sugar-binding properties of the flocculins are dependent upon the number of tandem repeats present in the flocculation gene. The different flocculent phenotypes are summarised in Table 1.
10. Flocculation Gene Structure
FLO genes are very long (up to 4.6 Kbp), whereas the average gene length in the Saccharomyces genome is only 1.0 Kbp because the number of tandem repeated DNA sequences of about 100 nucleotides are repeated 10–20 times in each gene [93]. Although tandem repeated DNA sequences in FLO genes are highly dynamic components of the genome, they change more rapidly than other parts of the Saccharomyces genome [27]. They enable rearrangements both between and within flocculin genes. The longer the Flo protein, the stronger the yeast strain’s flocculation ability [94]. FLO1 has been identified as the longest FLO1 gene, with the most repeats, and it confers the strongest flocculation phenotype [95].
Also, all FLO genes except FLO11 are located close to telomeres (disposable buffers at the end of chromosomes). Consequently, they are prone to rearrangements such as deletions, duplications, and translocations. Indeed, the Lg-FLO1 gene itself represents a translocation event between FLO genes located on different chromosomes. In addition, near-telometric genes (as well as FLO11) [96] can become transcriptionally repressed by an epigenetic process known as telomeric silencing. This is caused by an alteration in chromatin structure near the telomeres, leading to the silencing of genes located in that region. This effect can be long-lasting. For example, the epigenetic state of FLO11 is inheritable for many generations. The strength of this telomeric silencing varies greatly between different telomeres in yeast. Also, significant variation has also been detected between strains. The series of proteins associated with a trithorax-related SET domain protein (COMPASS) complex is involved in telomeric silencing in yeast [97]. FLO and MAL (a gene complex that controls maltose metabolism in yeast) genes near to telomeres were found to be silenced in some yeast strains, whereas in other strains with an inactivated COMPASS complex the expression of these genes was not affected. This is consistent with the finding of significant variation in the strength of the silencing effect between different chromosome ends and in different strains. In strains where the COMPASS complex had a strong silencing effect, genetic inactivation of this complex increased the expression of FLO1, FLO5, and FLO9 genes. Compared to wild cells, these mutants displayed enhanced flocculation properties during high gravity wort fermentation. Flocculation occurred earlier and formed much larger aggregates [96]. These findings are of considerable interest concerning the regulation of flocculation.
11. The Genetic Instability of Flocculation in Brewer’s Yeast Strains
The sedimentation performance of a brewer’s yeast strain very often changes during repeated cropping and re-pitching in a brewery [98]. In principle, this could be due to irreversible or reversible genetic change [98]. Alternatively, it could be due to long-lasting physiological, perhaps epigenetic (the study of cellular or physiological phenotypic traits) effects caused by external or environmental factors that switch genes on and affect how cells read genes [99] and respond to modifications in yeast-handling and fermentation environments (for example, high gravity worts [7]). When a genetic change, conferring a non-flocculent phenotype, occurs in a yeast culture, the culture gradually becomes a mixture of flocculent and non-flocculent cells [100]. Often within a production lager yeast population exhibiting moderate flocculent characteristics, a more flocculent variant within the culture can be isolated. An example of this development occurred when a Canadian brewing company began brewing its lager beer, under contract, in breweries located in the United Kingdom. Most of the contracted UK breweries employed vertical fermenters (Figure 5). However, at that time, Canadian breweries predominantly employed horizontal tanks (as both fermentation and maturation vessels). This difference in tank geometry influenced the yeast culture’s sedimentation characteristics. In vertical fermenters, this yeast culture was too non-flocculent (powdery), with considerable yeast cells remaining in suspension at the end of fermentation (centrifuges were not available in the UK breweries at the time). It was thought that possibly this culture contained a spectrum of isolates that exhibited differing flocculation intensities. Consequently, one of the variants from this strain, with more intensive flocculation characteristics, was successfully employed in the vertical fermenters. The result was less yeast in suspension at the end of fermentation. However, care had to be taken to ensure that the flocculent variant used was not too flocculent because under-fermented wort (unfermented maltose and maltotriose) and residual undesirable flavours (for example, diacetyl and other vicinal diketones) could have been the result [69].
PCR-based methodology (a technique that amplifies a single copy or a few copies of DNA in order to generate millions of copies of a particular DNA sequence) has been employed to detect FLO5 genes [101]. All 48 single colonies isolated from the stock culture of a lager strain exhibited this PCR band. After repeated recycling of this yeast culture in a brewery, single cells were again isolated from batches of the yeast that exhibited poor sedimentation performance. In one case, most (75%) single-cell colonies failed to yield the FLO5 PCR band, indicating that a genetic mutation (loss of intact FLO5) had occurred and spread throughout the population, and it was necessary to replace the brewery yeast with a fresh culture [102]. In the other case, nearly all (>90%) of the single colonies still showed the FLO5 PCR band, indicating that the poor sedimentation behaviour did not exclusively have a genetic basis, and was due in many instances to the culture’s physiological condition. In this case, the flocculation ability gradually recovered during repeated recycling of the yeast in the brewery.
Spontaneous changes in flocculation behaviour as a result of repeated re-pitching in wort during brewery fermentations have been studied by a number of research groups [103,104] and changes in flocculation intensity [76] have been observed. Cropped ale yeast, during 30 successive fermentations, has been studied [105]. During the first seven cycles, flocculation intensity increased from 50% to 100% of the original culture. Between the ninth and 23rd cycle, flocculation remained high. Then, between the 24th and 32nd cycles, flocculation ability and cell viability diminished. Sato and colleagues [100] reported that the flocculation intensity of a lager strain decreased after serial re-pitching. A long-term study in a number of breweries showed that flocculation intensity tended to decrease whilst at the same time other fermentation parameters (wort fermentation rate, ester, and higher alcohol production, etc.) usually remained constant. However, studies in the author’s breweries with high gravity worts (16–18 °Plato) do not confirm this fact—rather, they proposed that many observations of this nature must be yeast strain-dependent [7].
Cropping methods favour the enrichment of certain cell types. Few modern lager breweries currently recycle their yeast culture more than 20 times (many breweries less than 10 times) [1]. However, as the original wort concentration (gravity) increased, in the situation that occurs with high gravity brewing (HGB), the number of yeast cycles has been significantly reduced. Typically, with a 16 °Plato wort, the number of cycles is 10 times or less [7]. The results of these studies suggest that it is more likely that a change in flocculation behaviour is due to a modification in process conditions or raw materials (for example, a malt leading to changes in wort composition also increased the adjunct (un-malted cereal levels)), especially if the change persists when freshly propagated yeast is introduced into the process.
It has been suggested [103] that the PCR-based method to detect FLO5 genes (discussed above) can be used for the early detection of non-flocculent mutants in brewery fermentations. More than 30 different production lager strains with PCR primers designed to detect FLO5 have been detected. The PCR product varies in size from 4.8 to 2.3 Kb, with more flocculent strains showing larger gene products (>4 Kb) and less flocculent strains showing smaller products (2.3 Kb in a non-flocculent strain). Cell surface hydrophobicity also decreased with the reduced size of the PCR product. Consequently, the appearance of non-flocculent mutants during a series of brewery fermentations could be tested by PCR analyses. Interestingly, PCR primers designed for Lg-FLO1, FLO1, or the regulatory gene, FL08, did not exhibit this predictive power. In strains failing to form a PCR product with the PCR primers, Southern blot hybridisation (a method to detect a specific DNA sequence in DNA samples [106]) after chromosome fingerprinting using a FLO5 fragment as a probe showed that FLO5 was missing from chromosome VIII, whereas hybridisation to chromosome I (the location of FLO1) still occurred. The sequences of the successful FLO5 PCR primers were not revealed. Consequently, we do not know whether these primers only recognise FLO5 (which is 96% identical to FLO1).
12. The Influence of Cell Surface Hydrophobicity (CSH) and Cell Surface Charge on Yeast Flocculation
There have been a number of published studies that indicate that an increase of CSH and a decrease of cell surface charge occur at the outset of flocculation [107]. CSH increases rapidly as cells pass through the exponential growth phase and reach higher and more stable levels during the stationary phase [63]. Low CSH in an exponential phase culture is due to the presence of many daughter cells, which are significantly less hydrophobic than older (more mature) cells [69]. It is considered that CSH plays an important role in maintaining the correct conformation of flocculin molecules [50], so that the flocculins located in a stationary phase cell are more active [108]. It has already been discussed that the yeast cell surface has an overall negative charge. As a result, the cell wall phosphate groups and mannoproteins [109] are greater when the pH of the fermented medium is higher [110]. The cell surface charge of brewer’s yeast strains has been shown to vary during growth [111]. However, no clear relationship between cell surface charge and flocculation onset has been observed [112]. A number of environmental factors have been observed to affect CSH. Increased CSH can be stimulated by higher ethanol concentrations [21], lower temperatures [113], and higher pitching (inoculation) rates [21]. It has been proposed that hydrophobic oxylipins located at the cell surface of flocculent cells are a cause of increased CSH [91]. However, a recent publication [114] suggested that the precise rate of 3-OH oxylipin formation is still unclear and detection methods must be combined with novel techniques to target the cell wall architecture.
Ale strains (S. cerevisiae) have been found to be more hydrophobic and less negatively charged than lager (S. pastorianus) strains [107]. As well as contributing to floc formation, the greater hydrophobicity of ale strains (not definite) explains why the flocs of this species associate with CO2 bubbles and rise to the beer surface during traditional top-cropping ale fermentations, whereas the flocs of lager strains descend to the bottom of the fermenter (bottom cropping) [1].
13. Premature Yeast Flocculation
It is worth repeating that the timing of yeast flocculation during wort fermentation is crucial for beer production with the necessary quality characteristics. Occasionally, certain malts can cause premature or heavy flocculation, leaving the wort under-fermented with sugars still in solution and preventing the achievement of the alcohol specification [115]. This phenomenon has been termed premature yeast flocculation (PYF) [116] and is detected by way of a fermentability test [117].
Most of the PYF studies have not focussed on yeast per se. They have focussed on the malt employed to produce the wort used during the fermentation studies. Nevertheless, nature-nurture interactions are critical [11]. Also, the need for future detailed PYF studies with a number of brewer’s yeast strains (both ale and lager) are emphasised later.
Depending on the local national beer regulations, often a positive result with the fermentability test does not always translate into a problem because this discrepancy is not questioned. However, it has already been discussed (Section 8) that some countries (for example, Canada) enforce a rigid beer alcohol specification (±0.2 v/v) which is listed on every bottle, can, or keg.
During the past 50 years, more than 30 papers have discussed the PYF phenomenon. In the last two decades, South African Breweries Research Group in Johannesburg devoted considerable attention to PYF, which has considerably extended our understanding of the problem [117]. It should be emphasised that PYF is distinct from typical yeast flocculation, which discussed in other sections of this paper.
What exactly is PYF [66,116]? The published literature contains a number of definitions [118]. Although the details vary, the definitions are similar. Lake and Speers [119] noted that PYF behaviour results in:
- Rapid yeast flocculation out of suspension, resulting in elevated apparent extract values relative to a “normal” malt (depending on the fermentation vessel employed);
- typically, normal fermentations exhibit parabolic yeast-in-suspension trends while PYF yeast-in-suspension curves proceed in a normal and parabolic manner to a peak and then decline in a concave fashion (Figure 14).
Two theories dominate the literature with respect to the development of PYF factors in barley and malt. PYF tends to be a sporadic phenomenon that occurs concurrently with wet and rainy seasons. In the first theory, it is believed that increased microbial loads during wet seasons lead to the production of PYF factors [120]. Barley husk is the main carrier of microorganisms [121]. The microflora consists of bacteria, wild yeasts, and filamentous fungi.
The second theory regarding PYF mechanisms is that antimicrobial peptide factors inhibit or negatively affect yeast metabolism. This would initiate flocculation earlier and/or to a greater extent than normal. It has been shown that a PYF-positive malt leads to a minor decrease in wort sugar (particularly maltose and maltotriose) metabolism by yeast [10]. However, it is unclear whether this reduction in sugar metabolism is due to insufficient yeast in suspension (due to PYF) or a direct inhibition of yeast metabolism. However, Lake and colleagues [119], who employed a miniature fermentation assay, did not detect metabolic differences between PYF and a control malt. This course of events does not preclude antimicrobials associating with yeast cells, which would lead to increased flocculation.
As would be expected, yeast strains show different susceptibility to PYF malts [119]. It is unclear whether strain susceptibility is due to differing mechanisms, categories of PYF, or whether variability is related to the differing flocculation potential of various yeast strains. Also, it is unclear whether ale yeast (S. cerevisiae) strains are less susceptible or if the effects of PYF are masked by the flocculation behaviour of hydrophobic ale yeasts.
Studies by Chinese maltsters have conducted small-scale fermentations with worts produced from seven malts. These fermentations monitored their PYF potential and showed that PYF factors were present in both the malt husk and non-husk portions. Also, it was shown that antimicrobial substances that damage yeast cells were present in the non-husk portion [118]. Further research is required on the mechanisms of PYF activity. No single theory supports or challenges any of the currently proposed mechanisms. Most publications have reported on the gross chemical ratio of PYF factors and have avoided discussing or investigating PYF mechanisms.
14. Phenotypic Effects on Flocculation
Yeast flocculation complexities cannot be overlooked. In addition to the genetic characteristics (nature) of brewing strains (FLO genes together with their suppressors and activators), a number of nurture parameters will also affect yeast flocculation [11,65].
14.1. Cations
Cations have similar roles in both ale and lager yeast flocculation. However, calcium ions are recognised as the most effective ions for the promotion of flocculation [53]. Although details of this importance have been known for a long time [46], the interaction with the yeast cell surface requires further elaboration. It is important to reiterate the importance of calcium in yeast flocculation. Calcium can be removed from the yeast cell surface by washing with deionised water, through which many flocculent cultures will be de-flocculated. A de-flocculated culture will become flocculent again when calcium ions are added (Figure 15). Some flocculent yeast strains are not de-flocculated by washing with water, but rather the cells need to be treated with a solution of a chelating agent such as EDTA—10 mM [1] followed by washing with water to remove the EDTA. This treatment de-flocculates these cultures, and the flocculation phenotype is restored upon the re-addition of calcium ions.
Employing radio-labelled Ca45, it has been shown that flocculent cultures bind more Ca2+ ions than non-flocculent cultures [60]. There was no direct correlation found between the total calcium adsorbed and the strain’s flocculation phenotype. However, there is strain-to-strain variation in calcium binding, and furthermore this variation does not correlate with flocculation and non-flocculation when one strain was compared to another. With the knowledge that many flocculent yeast cultures can be de-flocculated by washing with deionised water, it was of interest to see if the amount of calcium washed off a yeast culture could be correlated with the visible loss of floc formation.
As a result of this calcium adsorption study, an improved perspective of calcium-binding behaviour in yeast and its relationship to flocculation might be obtained to test this hypothesis. Aliquots of flocculent and non-flocculent yeast suspensions were taken and incubated with the Ca45 solution. The yeast pellet was then washed four times with deionised water and the activity of each centrifuged supernatant was determined with a scintillation counter. In addition, the amount of calcium removed with each washing was determined. The first wash did not de-flocculate the flocculent yeast cultures but did remove adhering calcium around and in the interstitial spaces between the yeast cells. This source of calcium was expected to be relatively the same percentage of the total calcium bound for each yeast culture and was in all probability not related to flocculation, since the visible observation of flocculation did not disappear during this first wash.
Subsequent washings gradually dispersed any flocculation characteristic. The sum of the calcium removed in washings 2 to 4 were expressed as a percentage of the total calcium removed during washing. When the results were expressed in these terms (Table 2) for both flocculent and non-flocculent cultures, the flocculent cultures were found to have bound 28–48% more calcium after four washings than did non-flocculent cultures. As would be expected, there is strain-to-strain variation in calcium adsorption [60]. This variation is in all likelihood a reflection of diversities in cell wall structure strain to strain. In addition, this strain-to-strain variation in calcium adsorption per se does not correlate with the flocculation phenotype when one strain is compared to another. The only meaningful measure of calcium behaviour that correlated with flocculation was the case in which calcium was washed off the cell, and this coincided with the visible loss of flocculation.
Zn2+, Mg2+, and Mn2+ have also been described as inducers of flocculation [56,122]. Second to calcium, the effect of zinc on flocculation has received the greatest attention regarding yeast flocculation and there are a number of publications considering this parameter [123,124]. In addition, a great deal of research has been conducted studying the effect of Zn2+ on enzymatic activity (particularly—but not only—alcohol dehydrogenase) and fermentation efficiency [125]. The flocculation-de-flocculation behaviour of S. cerevisiae is strongly dependent on the concentration of Zn++ ions in the fermentation medium (for example, wort) and is yeast strain-specific. However, S. pastorianus is not affected by the presence of Zn2+, which suggests another useful method for distinguishing between lager and ale flocculent yeast strains [124]. However, the zinc effect requires considerable further study [126].
Cations such as Ba2+, Sr2+, and Ph2+ competitively inhibit yeast flocculation because of the similarity of their ionic ratios to Ca2+ [43]. It is possible that these cations compete for the same “calcium site” of flocculation lectins, but are not able to induce the appropriate conformation of the lectins. At low concentrations, Na+ and K+ induce flocculation most likely because of a reduction in the electrostatic repulsive forces with the yeasts and/or they stimulate the leakage of intracellular Ca2+ [122,127].
14.2. Medium pH
Medium pH can have a profound effect on the yeast flocculation phenotype. With many laboratory and industrial strains, flocculation occurs over a wide pH range (2.5–9.0), while many brewing strains (a sub-group of the NewFlo phenotype) only flocculate within a narrow pH range (3.9–5.5) [86]. In both cases, the optimum pH value takes place between 3.0 and 5.0, according to the yeast strain being studied. Extreme pH values promote a reversible dispersion of flocs. The modification of the pH value affects the ionisation of lectin amino acids with the consequent change in its conformation [52].
Most ale strains (S. cerevisiae) do not flocculate following growth in a chemically defined medium such as yeast nitrogen base (YNB) [11]. It could be that these strains also exhibit a narrow pH range of flocculation [86]. However, peptone, certain peptides, and wort also plays an important role because ale cells grown in a peptone-containing medium or wort exhibit the flocculation phenotype. This does not only occur in the culture medium. Following cell harvesting and washing, the cells were still flocculent in an in vitro flocculation test such as the Helm’s Flocculation Test [28] (Figure 6).
14.3. Temperature
Incubation temperature at different levels can affect the expression of the yeast flocculation phenotype. The lowering of growth and fermentation temperature results in a decrease in yeast metabolism and CO2 production. Consequently, there is a reduction in turbulence which favours yeast sedimentation. During beer fermentation, the agitation (shear force—as discussed later) temperature can also affect yeast flocculation by acting on cell-cell interactions. A rise in temperature to 50–60 °C, for only a few minutes, promotes the reversible dispersion of flocs [117], probably because of denatured flocculation lectins. Indeed, the incubation of yeast strains above the optimum growth (35–37 °C) leads to a reduction [22] or impairment of yeast flocculation [35]. It is probable that heat stress (<37 °C) acts directly on mitochondrial activity [128] and indirectly on cell membrane structure, which affects lectin secretion with a consequent reduction in flocculation.
14.4. Oxygen
The influence of wort oxygenation (aeration) in the early stages of both brewing and distilling fermentation is well documented [2]. In addition, there are a number of publications that discuss why moderate aeration is beneficial for yeast flocculation [129]. However, the principal role of oxygen during the initial stages of wort fermentation does not focus on yeast flocculation. It acts as a catalyst during the synthesis of unsaturated fatty acids and sterols, whose primary role is to participate in the structure of membranes, particularly the plasma and mitochondrial membranes [128]. The membrane structure is important for the stress protection of yeast cultures during adverse environmental conditions that prevail such as high gravity wort fermentations [7].
Cell wall mannoproteins are differently expressed under aerobic and anaerobic conditions [130]. The transition from semi-aerobic to anaerobic conditions, which occurs during the course of brewing fermentations, is probably associated with the expression of genes that regulate or encode flocculation lectins. Cells pitched into oxygen-depleted wort flocculated relatively early during the wort fermentation cycle, but to a limited extent. The addition of ergosterol or Tween80 (a non-ionic surfactant and emulsifier often used in foods and cosmetics) to the same wort restored the normal flocculation behaviour of the culture. This study concluded that lack of oxygen inhibited the synthesis of ergosterol and unsaturated fatty acids (UFAs) [128,129]. This limits cell growth and results in the early onset of stationary growth phase and cell flocculation [131].
14.5. Sugars
The presence or absence of fermentable sugars is a major factor influencing flocculation by NewFlo phenotypic strains [55]. It has long been discussed that as long as glucose, maltose, or maltotriose are present in sufficient amounts, flocculation is inhibited because these sugars occupy the flocculins. This inhibits binding to the mannose residues of adjacent cells. The addition of glucose (20 g/L) rapidly dissociated the flocs of a starved ale yeast culture containing the NewFlo phenotype [26]. This is a result that might be expected because glucose blocks the mannose-binding sites of NewFlo-type flocculins. However, it has been suggested that the loss of flocculation requires energy. Soares and Durante [132] examined the effect of nutrients on the loss of flocculation and presented evidence of six aspects regarding the stimulation of flocculation loss with an ale NewFlo phenotype strain under growing conditions:
- Carbohydrate sources are nutrients that stimulate the loss of flocculation in a defined growth medium (for example, yeast nitrogen base (YNB)) [60];
- all metabolisable carbon sources (for example, glucose, fructose, galactose, maltose, and sucrose) induce the loss of flocculation in YNB, which ethanol does not—details to follow [4];
- the rate of sugar-induced flocculation appears to be associated with the rate of sugar metabolism;
- the rate of sugar-induced flocculation loss most likely requires energy and this process is blocked by ethanol;
- growth does not always trigger flocculation loss because cells grown in a medium containing ethanol remained flocculent;
- glucose-induced loss of flocculation requires de novo protein synthesis—cycloheximide addition (an inhibitor of protein synthesis) to glucose-growing cells impairs the loss of flocculation [133].
15. Adhesion and Biofilm Formation
Yeast cells (including brewing and distilling strains) possess a capacity to adhere to abiotic (non-living) surfaces, cells, and tissues [9,10]. The cell wall serves as a means for yeast cultures (and other microbes) to interact with their environment. One of the most critical functions of the cell surface is its ability to adhere to other cells (floc formation) (Figure 4) and inert surfaces (Figure 9). Adhesion prevents cells from being washed away when they are present in a nourishing environment, and allows them to form biofilms that protect the cells from hazardous and stressful conditions. In addition to industrial environments, pathogenic yeasts exploit their capacity to adhere to abiotic surfaces such as plastic prosthetics in order to gain access to the bloodstream and internal organs of patients (Figure 10) [13].
Yeast cell adhesion is of considerable economic importance for food and beverage processing companies because adherent yeasts (and other fungi) can form highly resistant biofilms. As well as cell-cell adhesion (flocculation) in brewing (and wine) yeast cultures, biofilm formation is often exploited as a convenient and cost-effective way to separate biomass from fermentation products (for example, fermented wort and must). The fermentation of biofilms appears to be an adaptive mechanism because it usually ensures access to oxygen and permits continued growth on substrates such as nonfermentable ethanol. Biofilm-adhering cells have been shown to have elevated and/or modified lipid content as well as increased surface hydrophobicity [12].
16. Centrifuges to Crop Yeast
The incentive to optimise brewing operating costs while reducing processing times is imperative for the commercial survival of the process. Breweries (and many other manufacturing industries) continuously search for ways to exploit production efficiency [134] and lean technology concepts [135]. As a consequence, the disc stack centrifuge has become a popular component of yeast process management systems in order to reduce fermentation, maturation, and clarification times as well as to control effluent treatment costs [136] (Figure 2).
The use of centrifuges in breweries was initially viewed with misgivings by many brewers [137]. However, the advantages and disadvantages are now much clearer [138].
Modern centrifuges produce gravitational forces in excess of 10,000 times atmospheric pressure, achieving solid separation in seconds with reduced equipment volume. Centrifuges have a number of applications. They can be used with brewing yeast cultures for:
- Cropping of non-flocculent yeast cultures at the end of primary fermentation;
- reducing the yeast quantity from green beer before the start of secondary fermentation/maturation;
- beer recovery from cropped yeast [138];
- removal of cold break (precipitated protein, etc.) and yeast at the end of maturation;
- separation of the hot break after wort boiling.
Yeast management and handling systems are influential in determining the physiological status of yeasts’ subsequent fermentation performance activity [138] and clarification during aging/lagering/maturation [137]. It has been documented that beer haze can result from the yeast cell wall releasing mannan as it is processed during agitation in storage and centrifugation [139]. Hydrodynamic stresses can have the potential to inflict damage to the yeast cell wall during beer production and may lead to unfilterable beer haze formation. Intermediate haze formation from yeast (there are also other sources of beer haze—a discussion of which is beyond the scope of this review [140]).
The uses of centrifuges during brewing are diverse (discussed above). This section concentrates on the effect that centrifugation has on brewer’s yeast, particularly regarding yeast cell wall damage and the resulting impact on beer quality and stability together with yeast viability and vitality. Studies were conducted at Heriot-Watt University during the first decade of this century and focused on a better understanding of the effects of passing brewing yeast cultures through a disc stack centrifuge [136] as part of yeast cropping. In order to confirm that the effects on yeast was from centrifugation, an extensive number of centrifugation cycles operating at two different G-forces (high and low) was employed.
The passage of yeast through a disc stack centrifuge exposes cells to mechanical and hydrodynamic shear stresses [138]. These stresses can result in a decrease in cell viability and vitality, reduced flocculation intensity, cell wall damage, increased yeast extracellular proteinase A (PrA) levels, hazier beers, and poorer beer foam stability [2]. In a more recent study, biological indicators of yeast physiology such as viable and damaged cells, intracellular pH, glycogen levels, and trehalose levels, as well as beer physical stability parameters including mannan residues, particle size, and beer haze have been employed to quantify the damage to yeast cells as a function of the number of cycles through a centrifuge operating at high G-force (Table 3) [141].
A disc stack centrifuge (Figure 2) is typically made up of 50 to 150 discs (Figure 16); the number of discs is determined by process requirements. The discs are truncated cones and flanged at the inner and outer diameters (Figure 16). The close proximity of the discs to one another reduces the sedimentation distance for yeast cells. Due to centrifugal forces within the centrifuge, numerous reactionary forces take place between liquid, solid, and discs with shear and beer physical instability stress occurring on the yeast cell surface (Figure 17).
It has already been discussed here that in the late 1970s and early 1980s, Labatt in Canada (along with a number of other brewing companies) began installing centrifuges in their breweries. This was to harvest ale and lager yeasts at the end of fermentation and, for environmental reasons, to defer sewer surcharges. All of a sudden in 1988, one of the Labatt breweries with a centrifuge (operating at 300 hL/h) reported that its ale fermentations were exhibiting slower and incomplete fermentations with a 16 °Plato OG wort. Closer study of the fermented wort revealed reduced wort maltose and particularly maltotriose uptake rates, with residual sugars when fermentation ceased. Consequently, the fermented wort’s alcohol specification was not achieved. In addition, diacetyl (butterscotch-like flavour) and other vicinal diketone (VDK) levels were elevated at the end of fermentation because of difficulties with VDK reabsorption by yeast at the end of primary fermentation and during maturation [1]. Also, yeast autolysis occurred, resulting in reduced foam stability due to excreted intracellular proteinase, elevated unfilterable haze consisting mainly of mannoproteins from disrupted cell walls, and autolysed yeast off-flavours [141].
The centrifuged ale yeast exhibited decreasing cell viability (determined with methylene blue staining) during repeated cycles. Also, the same cultures had a higher percentage of respiratory deficient (RD) petite mutants, determined with the triphenyl tehazolium chloride overlay method [142]. This increasing RD level and decreasing viability were due to centrifugation when the exit temperature was 30 °C [128]. When the bowl of the centrifuge was cooled and the exit temperature reduced to 20 °C, the cell viability increased, the RD level decreased, the wort fermentation characteristics returned to normal, and the beer was drinkable again [141].
Studies at Heriot Watt University with a 5 hL/h centrifuge (Figure 2) quantified the damage that occurs to brewer’s yeast strains as a function of cycles through the centrifuge operating at low and high G-forces [136]. Biological indicators of yeast physiology as a result of centrifugation include non-viable and damaged cells, intracellular pH, glycogen and trehalose levels, as well as beer physical stability parameters such as mannan residues and particle size distribution. Many of these measurements were conducted with a flow cytometer (Table 3) [143,144]. A detailed discussion of the principles of flow cytometry is beyond the scope of this paper, but it has been developed to measure yeast cell viability, damaged cells, intracellular pH (pHi), extracellular PrA levels, mannose residues, and intracellular glycogen and trehalose (Table 3). When yeast cultures are passed through a disc stack centrifuge they exhibit lower cell viabilities when compared with cells that had not been centrifuged. In addition, the effects of G-force upon yeast cells as a function of centrifugation cycles have been studied [141]. In order to establish a relationship between a centrifuge G-force and effects on the physiological state of yeast, it was necessary for cells to possess an analogous history and a similar physiological state. The yeast strain employed in this study, S. cerevisiae (an ale strain), was subjected to similar fermentation conditions. The yeast culture was exposed to a high G-force (12,000 rpm ≡ 20,000 G) and the centrifuged cells were subsequently conditioned for an additional 48 h.
Centrifugation of cultures after nine times recycling through 20 °Plato worts demonstrated that the yeast had been damaged in a number of ways (Table 3). These included a reduction in cell viability as well as glycogen and trehalose levels. Also, the hydrodynamic shear generated by the centrifuge resulted in cell surface components being released (mannoprotein) (Figure 17), together with a PrA increase, resulting in a reduction in wort hydrophobic polypeptides and poor beer foam stability [141].
Although centrifugation can exhibit negative effects, the positive effects of controlled centrifugation on beer production and effluent control cannot be overstated. However, yeast is subjected to numerous factors that individually and collectively impose stresses on yeast cells (Figure 18) [2]. The effects of environmental conditions and beer production equipment may have been underestimated (and even ignored). A more complete understanding of yeast’s biological response to interactions with cell physiology and brewing equipment is an important criterion in maintaining process efficiency and beer quality. It is worth noting that the advent of flow cytometry [143,144] and confocal imaging [145] has introduced analytical methods to yeast research that have broadened the scope of research in this area.
17. Conclusions
Brewers employ a number of methods to crop their yeast cultures at the end of a typical wort fermentation, which vary depending on whether one is dealing with traditional top-cropping, accepted lager bottom-cropping, cylindroconical fermentation systems, a non-flocculent culture where the yeast, still in suspension, is cropped with a centrifuge or a portion of the yeast, still in suspension, is blended into the fresh wort of a subsequent fermentation. Yeast quality is influenced by the manner that the yeast is cropped and centrifuges play an important part in this regard. The cell wall structure is a critical parameter involved in yeast flocculation. The wall consists of an inner layer composed predominantly of β-glucose and chitin with a fibrillar outer layer consisting primarily of α-mannan associated with mannoproteins (Figure 8). Yeast management and handling systems, including culture harvesting, are critical in determining the yeast’s physiological status.
Acknowledgments
The invaluable assistance and support of Anne Anstruther in developing this review paper is gratefully acknowledged.
Conflicts of Interest
The author declare no conflicts of interest.
References
- Stewart, G.G.; Russell, I. An Introduction to Brewing Science and Technology. Series III. Brewer’s Yeast, 2nd ed; Institute of Brewing and Distilling: London, UK, 2009. [Google Scholar]
- Stewart, G.G. Brewing and Distilling Yeasts; Springer: Cham, Switzerland, 2017; pp. 241–258. [Google Scholar]
- Lekkas, C.; Stewart, G.G.; Hill, A.; Taidi, B.; Hodgson, J. Elucidation of the role of nitrogenous wort components in wort fermentation. J. Inst. Brew. 2007, 113, 183–191. [Google Scholar] [CrossRef]
- Stewart, G.G. Studies on the uptake and metabolism of wort sugars during brewing fermentations. Tech. Quart. Master Brew. Assoc. Am. 2006, 43, 265–269. [Google Scholar] [CrossRef]
- Stewart, G.G. Biochemistry of brewing. In Biochemistry of Food; Eskin, N.A.M., Shahidi, N., Eds.; Elsevier: Oxford, UK, 2012; pp. 291–318. [Google Scholar]
- Piper, P.W.; Talreja, K.; Panaretou, B.; Moradas-Ferreira, P.; Byrne, K.; Praekelt, U.M.; Meacock, P.; Récnacq, M.; Boucherie, H. Induction of major heat-shock proteins of Saccharomyces cerevisiae, including plasma membrane Hsp30, by ethanol levels above a critical threshold. Microbiology 1994, 140, 3031–3038. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.G. Brewing Intensification; American Society for Brewing Chemists: St. Paul, MN, USA, 2014. [Google Scholar]
- Verbelen, P.J.; Saerens, S.M.; Van Mulders, S.E.; Delvaux, F.; Delvaux, F.R. The role of oxygen in yeast metabolism during high cell density brewery fermentations. Appl. Microbiol. Biotechnol. 2009, 82, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V. Flocculation in Saccharomyces cerevisiae: A review. J. Appl. Microbiol. 2010, 110, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.G.; Hill, A.E.; Russell, I. 125th anniversary review—Developments in brewing and distilling yeast strains. J. Inst. Brew. 2013, 119, 202–220. [Google Scholar] [CrossRef]
- Stewart, G.G. The concept of nature-nurture applied to brewer’s yeast and wort fermentations. Tech. Quart. Master Brew. Assoc. Am. 2015, 52, 146–151. [Google Scholar] [CrossRef]
- Mamvura, T.A.; Paterson, A.E.; Fanucchi, D. The impact of pipe geometry variations on hygiene and success of orbital welding of brewing industry equipment. J. Inst. Brew. 2017, 123, 81–97. [Google Scholar] [CrossRef]
- Douglas, L.J. Adhesion of Candida species to epithelial surfaces. Crit. Rev. Microbiol. 1987, 15, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Speers, R.A. A review of yeast flocculation. In International Brewers Symposium: Yeast Flocculation, Vitality and Viability; Master Brewers Association of the Americas: St. Paul, MN, USA, 2012. [Google Scholar]
- Miki, B.; Poon, N.H.; James, A.P.; Seligy, V.L. Possible mechanisms for flocculation interactions governed by gene FLO1 in Saccharomyces cerevisiae. J. Bacteriol. 1982, 150, 878–889. [Google Scholar] [PubMed]
- Stewart, G.G.; Russell, I. Yeast Flocculation. Brewing Science; Pollock, J.R.A., Ed.; Academic Press: London, UK, 1981; Volume 2, pp. 61–92. [Google Scholar]
- Gilliland, R.B. The flocculation characteristics of brewing yeasts during fermentation. Proc. Eur. Brew. Conv. Congr. Brighton 1951, 35–58. [Google Scholar]
- Thorne, R.S.W. Some aspects of yeast flocculation. Proc. Eur. Brew. Conv. Congr. Brighton 1951, 21–34. [Google Scholar]
- Stewart, G.G.; Russell, I.; Sills, A.M. Factors that control the utilization of wort carbohydrates by yeast. Tech. Quart. Master Brew. Assoc. Am. 1983, 20, 1–8. [Google Scholar]
- Marika, H.; Straver, J.; Kijne, J.W.; Smit, G. Cause and control of flocculation in yeast. Trends Biotechnol. 1993, 11, 228–232. [Google Scholar]
- Jin, Y.; Speers, R.A. Effect of environmental conditions on the flocculation of Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2000, 58, 108–116. [Google Scholar]
- Soares, E.V.; Teixeira, J.A.; Mota, M. Effect of cultural and nutritional conditions on the control of flocculation expression in Saccharomyces cerevisiae. Can. J. Microbiol. 1994, 40, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mulders, S.E.; Ghequire, M.; Daenen, L.; Verbelen, P.J.; Verstrepen, K.J.; Delvaux, F.R. Flocculation gene variability in industrial brewer’s yeast strains. Appl. Microbiol. Biotechnol. 2010, 88, 1321–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, L. Improvements in the fermentation and maturation of beers. J. Inst. Brew. 1930, 36, 538–554. [Google Scholar] [CrossRef]
- Krogerus, A.; Gibson, B.R. 125th anniversary review: Diacetyl and its control during brewery fermentation. J. Inst. Brew. 2013, 119, 86–97. [Google Scholar]
- Stratford, M. Yeast flocculation—A new perspective. Adv. Microb. Physiol. 1992, 33, 1–71. [Google Scholar]
- Verstrepan, K.J.; Klis, F.M. Flocculation, adhesion and biofilm formation in yeasts. Mol. Microbiol. 2006, 60, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Helm, E.; Nohr, B.; Thorne, R.S.W. The measurement of yeast flocculation and its significance in brewing. Wall. Lab. Commun. 1953, 16, 315–326. [Google Scholar]
- Stewart, G.G.; Garrison, I. Some observations on co-flocculation in Saccharomyces cerevisiae. Am. Soc. Brew. Chem. Proc. 1972, 118–131. [Google Scholar]
- Stewart, G.G.; Russell, I. The identification, characterization, and mapping of a gene for flocculation in Saccharomyces sp. Can. J. Microbiol. 1977, 23, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Boulton, C. Yeast handling. Brew. Dig. Int. 2011, 7, 7–10. [Google Scholar]
- Calleja, G.G. Microbial Aggregation; CRC Press: Boca Raton, FL, USA, 1984. [Google Scholar]
- Day, A.W.; Poon, N.H.; Stewart, G.G. Fungal fimbriae. III. The effect of flocculation in Saccharomyces. Can. J. Microbiol. 1975, 21, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Claro, F.B.; Rijsbrack, K.; Soares, E.V. Flocculation onset in Saccharomyces cerevisiae: Effect of ethanol, heat and osmotic stress. J. Appl. Microbiol. 2007, 102, 693–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashbee, R.; Bignell, E.M. (Eds.) Pathogenic Yeasts. The Yeast Handbook; Springer: Berlin, Germany, 2010. [Google Scholar]
- Stewart, G.G. MBAA Award of Merit Lecture. A love affair with yeast. Tech. Quart. Master Brew. Assoc. Am. 2010, 47, 4–11. [Google Scholar]
- Winge, O. On haplophase and diplophase in some Saccharomycetes. C. R. Trav. Lab. Carlsberg. Ser. Physiol. 1935, 21, 77–111. [Google Scholar]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth; regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar] [CrossRef]
- Zarattini, R.A.; Williams, J.W.; Ernandes, J.R.; Stewart, G.G. Bacterial-induced flocculation in selected brewing strains of Saccharomyces cerevisiae. Cerevisia Biotechnol. 1993, 18, 65–70. [Google Scholar]
- Guo, B.; Styles, C.A.; Feng, Q.; Fink, G.R. A Saccharomyces gene family involved in invasive growth, cell-cell adhesion, and mating. Proc. Nat. Acad. Sci. USA 2000, 97, 12158–12163. [Google Scholar] [CrossRef] [PubMed]
- Bilinski, C.A.; Russell, I.; Stewart, G.G. Analysis of sporulation in brewer’s yeast: Induction of tetrad formation. J. Inst. Brew. 1986, 92, 594–598. [Google Scholar] [CrossRef]
- Nishihara, H.; Toraya, T.; Fukui, S. Flocculation of cell walls of brewer’s yeast and effects of metal ions, protein-denaturants and enzyme treatments. Arch. Microbiol. 1982, 131, 112–115. [Google Scholar] [CrossRef]
- Eddy, A.A. Composite ratio of the flocculation process of top and bottom strains of Saccharomyces cerevisiae. J. Inst. Brew. 1958, 64, 143–151. [Google Scholar] [CrossRef]
- Peng, X.; Sun, J.; Iserentant, D.; Michiels, C.; Verachtert, H. Flocculation and co-flocculation of bacteria by yeasts. Appl. Microbiol. Biotechnol. 2007, 21, 31–37. [Google Scholar]
- Stratford, M. Yeast flocculation: Calcium specificity. Yeast 1992, 5, 487–496. [Google Scholar] [CrossRef]
- Stewart, G.G.; Maskell, D.L.; Speers, A. Brewing fundamentals—Fermentation. Tech. Quart. Master Brew. Assoc. Am. 2016, 53, 2–22. [Google Scholar]
- Lyons, T.P.; Hough, J.S. The role of yeast cell walls in brewing. Brew. Dig. 1970, 45, 52–60. [Google Scholar]
- Beavan, M.T.; Belki, D.; Stewart, G.G.; Rose, A.H. Changes in electrophoretic mobility and lytic enzyme activity associated with developments of flocculating ability in Saccharomyces cerevisiae. Can. J. Microbiol. 1979, 25, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Speers, R.A. Effect of cell surface hydrophobicity, charge, and zymolectin density on the flocculation of Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2001, 58, 108–116. [Google Scholar]
- Smit, G.; Straver, M.H.; Lugtenberg, B.J.; Kijne, J.W. Flocculence of Saccharomyces cerevisiae cells is induced by nutrient limitation, with cell surface hydrophobicity as a major determinant. Appl. Environ. Microbiol. 1992, 58, 3709–3714. [Google Scholar] [PubMed]
- Halina, S.; Nathan, L. Lectins; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Miki, B.L.A.; Poon, N.H.; Seligy, N.L. Repression and induction of flocculation interactions to Saccharomyces cerevisiae. J. Bacteriol. 1982, 150, 890–899. [Google Scholar] [PubMed]
- Goldstein, I.J.; Poretz, R.D. Isolation, physicochemical characterization and carbohydrate-binding specificity of lectins. In The Lectins; Liener, I.E., Goldstein, I.J., Eds.; Academic Press: Orlando, FL, USA, 1986; pp. 52–54. [Google Scholar]
- Stratford, M.; Assinder, S. Yeast flocculation: Flo1 and new Flo phenotypes and receptor structure. Yeast 1991, 7, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V.; Mota, M. Flocculation onset, growth phase, and genealogical age in Saccharomyces cerevisiae. Can. J. Microbiol. 1996, 42, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunissen, A.W.R.H.; Steensma, H.Y. Review: The dominant flocculation genes of Saccharomyces cerevisiae constitute a new subtelomeric gene family. Yeast 1995, 11, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Van Mulders, S.E.; Christianen, E.; Saerens, S.E.; Daenen, L.; Verbelen, P.J.; Willaert, R.; Verstrepen, K.J.; Delvaux, F.R. Phenotypic diversity of Flo protein family-mediated adhesion in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Siero, C.; Reboredo, N.M.; Villa, T.G. Flocculation of industrial and laboratory strains of Saccharomyces cerevisiae. J. Ind. Microbiol. 1994, 14, 461–466. [Google Scholar] [CrossRef]
- Stewart, G.G.; Russell, I.; Garrison, I.F. Some considerations of the flocculation characteristics of ale and lager yeast strains. J. Inst. Brew. 1975, 81, 248–257. [Google Scholar] [CrossRef]
- Van Hamersveld, E.H.; van der Lans, R.G.; Luyben, K.G. Quantification of brewer’s yeast flocculation in a stirred tank: Effect of physical parameters on flocculation. Biotechnol. Bioeng. 1997, 56, 190–200. [Google Scholar] [CrossRef]
- Wilcocks, K.R.; Smart, K.A. The importance of surface charge of hydrophobicity for the flocculation of chain forming brewing yeast strains and resistance of these parameters to acid washing. FEMS Microbiol. Lett. 1995, 15, 293–297. [Google Scholar] [CrossRef]
- Speers, R.A.; Wan, Y.-Q.; Stewart, R.J. Effects of fermentation parameters and cell wall properties on yeast flocculation. J. Inst. Brew. 2006, 112, 246–254. [Google Scholar] [CrossRef]
- Jarvis, P.; Jefferson, B.; Parsons, S.A. Measuring flocstructural characteristics. Environ. Sci. Biotechnol. 2005, 4, 1–18. [Google Scholar] [CrossRef]
- Stewart, G.G.; Russell, I.; Goring, T.E. Nature-nurture anomalies—Further studies in yeast flocculation. Am. Soc. Brew. Chem. Proc. 1975, 33, 137–147. [Google Scholar]
- Axcell, B.C.; van Nierop, S.; Vundla, W. Malt induced premature yeast flocculation. Tech. Quart. Master Brew. Assoc. Am. 2000, 37, 501–504. [Google Scholar]
- Mortier, A.; Soares, E.V. Separation of yeasts by addition of flocculent cells of Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 1401–1407. [Google Scholar] [CrossRef]
- Eddy, A.A.; Rudin, A.D. Part of the yeast surface apparently involved in flocculation. J. Inst. Brew. 1958, 64, 19–21. [Google Scholar] [CrossRef]
- Stewart, G.G. Seduced by yeast. J. Am. Soc. Brew. Chem. 2015, 73, 1–21. [Google Scholar]
- Gilliland, R.B. Determination of yeast viability. J. Inst. Brew. 1959, 65, 424–429. [Google Scholar] [CrossRef]
- Richards, M. The use of giant-colony morphology for the differentiation of brewing yeasts. J. Inst. Brew. 1967, 73, 162–166. [Google Scholar] [CrossRef]
- Stewart, G.G. Recent developments in the characterization of brewery yeast strains. Tech. Quart. Master Brew. Assoc. Am. 1973, 9, 183–191. [Google Scholar]
- White, F.H.; Kidney, E. The influence of yeast strain on beer spoilage bacteria. Proc. Eur. Brew. Conv. Congr. Berl.-West 1979, 801–815. [Google Scholar]
- Rossouw, D.; Bagheri, B.; Setati, E.; Bauer, F.F. Co-flocculation of yeast species, a new mechanism to govern population dynamics in microbial ecosystems. PLoS ONE 2015, 10, e0136249. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.L.; Cookson, B.T. Apoptosis, pyroptosis, and necrosis: Mechanistic description of dead and dying eukaryotic cells. Infect. Immun. 2005, 73, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.D.; Diacetis, A.N. Long term serial repitching and the genetic and phenotypic stability of brewer’s yeast. J. Inst. Brew. 2007, 113, 67–74. [Google Scholar] [CrossRef]
- Pomper, S.; Burkholder, P.R. Studies on the biochemical genetics of yeast. Proc. Natl. Acad. Sci. USA 1949, 35, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Panchal, C.J.; Russell, I.; Sills, A.M.; Stewart, G.G. Genetic manipulation of brewing and related yeast strains. Food Technol. 1984, 111, 99–106. [Google Scholar]
- Lewis, C.W.; Johnston, J.R.; Martin, P.A. The genetics of yeast flocculation. J. Inst. Brew. 1976, 82, 158–160. [Google Scholar] [CrossRef]
- Mundy, R.D.; Cormack, B. Expression of Candida glabrata adhesion following exposure to chemical preservatives. J. Infect. Dis. 2009, 199, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Green, C.B.; OH, S.H.; Zhao, X. Discovering the secrets of the Candida albicans agglutinin-like sequence (ALS) gene family—A sticky pursuit. Med. Mycol. 2008, 46, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Oliver, S.G. Life with 6000 genes. Science. 1996, 274, 546, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Sherman, F.; Fink, G.R.; Hicks, J.B. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1986. [Google Scholar]
- Meaden, P.G. DNA fingerprinting of brewer’s yeast. Ferment 1996, 9, 267–272. [Google Scholar]
- Soares, E.V.; Seynaeve, J. The use of succinic acid, as a pH buffer, expands the potentialities of utilisation of a chemically defined medium in Saccharomyces cerevisiae flocculation studies. Biotechnol. Lett. 2000, 22, 859–863. [Google Scholar] [CrossRef]
- Stratford, M. Induction of flocculation in brewing yeasts by change in pH value. FEMS Microbiol. Lett. 1996, 136, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Russell, I.; Stewart, G.G. Revised nomenclature of genes that control flocculation. J. Inst. Brew. 1980, 86, 120–121. [Google Scholar] [CrossRef]
- Bayly, J.C.; Douglas, L.M.; Pretorium, I.S.; Bauer, F.F.; Dranginis, A.M. Characteristics of Flo11-dependent flocculation in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2005, 5, 1151–1156. [Google Scholar]
- Bester, M.C.; Pretorius, I.S.; Bauer, F.F. The regulation of Saccharomyces cerevisiae FLO gene expression and Ca2+-dependent flocculation by Flo8p and Mss11p. Curr. Genet. 2006, 49, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Govender, P.; Domingo, J.L.; Bester, M.C.; Pretorius, I.S.; Bauer, F.F. Controlled expression of the dominant flocculation genes FLO1, FLO5 and FLO11 in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 6041–6052. [Google Scholar] [CrossRef] [PubMed]
- Kock, J.L.; Venter, P.; Smith, D.P.; Van Wyk, P.W.; Botes, P.J.; Coetzee, D.J.; Pohl, C.H.; Both, A.; Riedel, K.-H.; Nigam, S. A novel oxylipin-associated “ghosting” phenomenon in yeast flocculation. Antonie Van Leeuwenhoek 2000, 77, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; MacGurn, J.A.; Chu, T.; Stefan, C.J.; Emr, S.D. Arrestin-related ubiquitin-ligase adaptors regulate endocytosis and protein turnover at the cell surface. Cell 2008, 135, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Reynolds, T.B.; Fink, G.R. Origins of variation in the fungal cell surface. Nat. Rev. Microbiol. 2004, 2, 533–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halme, A.; Bumgarner, S.; Styles, C.; Fink, G.R. Genetic and epigenetic resolution of the FLO gene family generates cell-surface variation in yeast. Cell 2004, 116, 405–415. [Google Scholar] [CrossRef]
- Dunn, B.; Sherlock, G. Reconstitution of the genome origins and evolution of the hybrid lager yeast Saccharomyces pastorianus. Genome Res. 2008, 18, 1610–1623. [Google Scholar] [CrossRef] [PubMed]
- Loney, E.R.; Inglis, P.M.; Sharp, S.; Pryde, F.E.; Kent, N.A.; Mellor, J.; Louis, E.J. Repressive and non-repressive chromatin at nature telomeres in Saccharomyces cerevisiae. Epigenet. Chromat. 2009, 2, 18–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, T.; Krogan, N.J.; Dover, J.; Erdjument-Bromage, H.; Tempst, P.; Johnston, M.; Greenblatt, J.F.; Shilatifard, A. COMPASS: A complex of proteins associated with a tuthorax-related SET domain protein. Proc. Natl. Acad. Sci. USA 2001, 98, 12902–12907. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.G. The Horace Brown Medal Lecture—Forty years of brewing research. J. Inst. Brew. 2009, 115, 3–29. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Watari, J.; Shinotusuka, K. Genetic instability of flocculation of bottom-fermenting yeast. J. Am. Soc. Brew. Chem. 2001, 59, 130–134. [Google Scholar]
- Jibiki, M.; Ishibiki, T.; Yuuki, T.; Kagami, N. Application of polymerase chain reaction to determine the flocculation properties of brewer’s lager yeast. J. Am. Soc. Brew. Chem. 2001, 59, 107–110. [Google Scholar]
- Stewart, G.G. Yeast performance and management. Brewer 1996, 82, 211–215. [Google Scholar]
- Smart, K.A.; Whisker, S. Effect of serial repitching on the fermentation properties and condition of brewing yeast. J. Am. Soc. Brew. Chem. 1996, 54, 41–44. [Google Scholar]
- Teixeira, J.M.; Teixeira, J.A.; Mota, M.; Manuela, M.; Guerra, B.; Machado, M.; Cruz, J.M.; S’Almeida, A.M. The influence of cell wall composition of a brewer’s flocculent lager yeast on sedimentation during successive industrial fermentations. Proc. Eur. Brew. Conv. Congr. Lisb. 1991, 23, 241–248. [Google Scholar]
- Wightman, P.; Quain, D.E.; Meadon, P.G. Analysis of production brewing strains of yeast by DNA fingerprinting. Lett. Appl. Microbiol. 1996, 22, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Southern, E.M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
- Amory, D.E.; Rouxhet, P.G.; Dufour, J.P. Flocculence of brewery yeasts and their surface properties: Chemical composition, electrostatic charge and hydrophobicity. J. Inst. Brew. 1988, 94, 79–84. [Google Scholar] [CrossRef]
- Lange, C.; Nett, J.H.; Trumpower, B.L.; Hunte, C. Specific roles of protein-phospholipid interactions in the yeast cytochrome bc1 complex structure. EMBO J. 2001, 20, 6591–6600. [Google Scholar] [CrossRef] [PubMed]
- Lyons, T.P.; Hough, J.S. Flocculation of brewer’s yeast. J. Inst. Brew. 1970, 76, 564–571. [Google Scholar] [CrossRef]
- Boulton, C.; Quain, D. Brewing Yeast and Fermentation; Blackwell Science: Oxford, UK, 2001. [Google Scholar]
- Van Holle, A.; Machado, M.D.; Soares, E.V. Flocculation in ale brewing strains of Saccharomyces cerevisiae: Re-evaluation of the role of cell surface charge and hydrophobicity. Appl. Microbiol. Biotechnol. 2011, 93, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Dengis, P.B.; Nélissen, L.R.; Rouxhet, P.G. Mechanisms of yeast flocculation, comparison of top- and bottom-fermenting strains. Appl. Environ. Microbiol. 1995, 61, 718–728. [Google Scholar] [PubMed]
- Van Lersel, M.F.M.; Meersman, E.; Arntz, M.; Robvouts, F.M.; Abee, T. Effect of environmental conditions on flocculation and immobilisation of brewer’s yeast during production of alcohol-free beer. J. Inst. Brew. 1998, 104, 131–136. [Google Scholar]
- Potter, G.; Budge, S.M.; Speers, R.A. Flocculation, cell surface hydrophobicity and 3-OH oxylipins in the SMA strain of Saccharomyces pastorianus. J. Inst. Brew. 2015, 121, 31–37. [Google Scholar] [CrossRef]
- Ishimaru, S.; Kudo, S.; Hattan, M.; Yoshida, T.; Kataoka, J. Selection of small vessels for fermentation tests in the laboratory. Rep. Res. 1967, 10, 61–65. [Google Scholar]
- Krugger, L.; Ryder, D.S.; Alcock, C.; Murray, J.P. Malt quality: Prediction of malt fermentability. Part 1. Tech. Quart. Master Brew. Assoc. Am. 1982, 19, 45–51. [Google Scholar]
- Koizumi, H.; Ogawa, T. Rapid and sensitive method to measure premature yeast flocculation activity in malt. J. Am. Soc. Brew. Chem. 2005, 63, 147–150. [Google Scholar]
- Van Nierop, S.N.E.; Rautenbach, M.; Axcell, B.C. The impact of microorganisms on barley and malt quality—A review. J. Am. Soc. Brew. Chem. 2006, 62, 69–79. [Google Scholar]
- Lake, T.C.; Speers, R.A. A Discussion of malt-induced premature yeast flocculation. Tech. Quart. Master Brew. Assoc. Am. 2008, 4, 253–262. [Google Scholar]
- Van Nierop, S.N.E.; Cameron-Clarke, A.; Axcell, B.C. Enzymatic generation of malt factors responsible for premature yeast flocculation. J. Am. Soc. Brew. Chem. 2004, 62, 108–116. [Google Scholar]
- Armstrong, K.; Bendiak, D. PYF malt: Practical brewery observations of fermentability. Tech. Quart. Master Brew. Assoc. Am. 2007, 44, 40–46. [Google Scholar] [CrossRef]
- Stewart, G.G.; Goring, T.E. Effect of some monovalent and divalent metal ions on the flocculation of brewer’s yeast strains. J. Inst. Brew. 1976, 82, 341–342. [Google Scholar] [CrossRef]
- Russell, I.; Dowhanick, T.; Raspor, P.; Stewart, G.G. Yeast flocculation—The influence of divalent ions. Proc. Eur. Brew. Conv. Congr. Zurich 1989, 529–536. [Google Scholar]
- Raspor, P.; Russell, I.; Stewart, G.G. An update of zinc ion as an effector of flocculation in brewer’s yeast strains. J. Inst. Brew. 1990, 96, 303–305. [Google Scholar] [CrossRef]
- Rees, E.M.R.; Stewart, G.G. Strain specific response of brewer’s yeast strains to zinc concentration in conventional and high gravity worts. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Rees, E.M.R.; Stewart, G.G. The effects of divalent ions magnesium and calcium on yeast fermentation performance in conventional (12 °P) and high (20 °P) gravity worts in both static and shaking fermentations. Proc. Eur. Brew. Conv. Congr. Maastricht 1997, 461–468. [Google Scholar]
- Gouveio, C.; Soares, E.V. Pb2+ inhibits competitively flocculation of Saccharomyces cerevisiae. J. Inst. Brew. 2004, 110, 141–145. [Google Scholar] [CrossRef]
- Stewart, G.G. Yeast mitochondria—Their influence on brewer’s yeast fermentation and medical research. Tech. Quart. Master Brew. Assoc. Am. 2004, 51, 3–11. [Google Scholar]
- Lorenz, R.T.; Parks, L.W. Involvement of heme components in sterol metabolism of Saccharomyces cerevisiae. Lipids 1991, 26, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Abramova, N.; Sertil, O.; Mehta, S.; Lowry, C.V. Reciprocal regulation of anaerobic and aerobic cell wall mannoprotein gene expression in Saccharomyces cerevisiae. J. Bacteriol. 2001, 183, 2881–2887. [Google Scholar] [CrossRef] [PubMed]
- Straver, M.; Aar, P.C.V.D.; Sinit, G.; Kijne, J.W. Determinants of flocculence of brewer’s yeast during fermentation in wort. Yeast 1993, 9, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V.; Duarte, A.A. Addition of nutrients induce a fast loss of flocculation-stirred cells of Saccharomyces cerevisiae. Biotechnol. Lett. 2002, 24, 1957–1960. [Google Scholar] [CrossRef]
- Baker, D.A.; Kirsop, B.H. Flocculation in Saccharomyces cerevisiae is influenced by wort composition and by actidione. J. Inst. Brew. 1972, 78, 454–458. [Google Scholar] [CrossRef]
- Bower, J.L.; Christensen, C.M. Disruptive technologies: Catching the wave. Harv. Bus. Rev. 1995, 73, 43–47. [Google Scholar]
- Dudbridge, M. Handbook of Lean Manufacturing; Wiley: New York, NY, USA, 2011. [Google Scholar]
- Chlup, P.H.; Bernard, D.; Stewart, G.G. Disc stack centrifuge operating parameters and their impact on yeast physiology. J. Inst. Brew. 2008, 114, 45–61. [Google Scholar] [CrossRef]
- Siebert, K.J.; Stenroose, L.E.; Reid, D.S.; Grabowski, D. Filtration difficulties resulting from damage to yeast during centrifugation. Tech. Quart. Master Brew. Assoc. Am. 1987, 24, 1–8. [Google Scholar]
- Chlup, P.H.; Stewart, G.G. Centrifuges in brewing. Tech. Quart. Master Brew. Assoc. Am. 2011, 48, 48–50. [Google Scholar] [CrossRef]
- Lewis, M.J.; Poerwantaro, W.M. Release of haze material from the cell walls of agitated yeast. J. Am. Soc. Brew. Chem. 1991, 49, 43–46. [Google Scholar]
- Stewart, G.G.; Russell, I.; Anstruther, A. (Eds.) Handbook of Brewing, 3rd ed.; Taylor & Francis Group: Abingdon, UK, 2018. [Google Scholar]
- Chlup, P.H.; Bernard, D.; Stewart, G.G. The disc stack centrifuge and its impact on yeast and beer quality. J. Am. Soc. Brew. Chem. 2007, 65, 151–155. [Google Scholar]
- Ogur, M.; St. John, R.; Nagai, S. Tetrazolium overlay technique for population studies of respiratory deficiency in yeast. Science 1957, 125, 928–929. [Google Scholar] [CrossRef] [PubMed]
- Chlup, P.H.; Conery, J.; Stewart, G.G. Detection of mannan from Saccharomyces cerevisiae. J. Am. Soc. Brew. Chem. 2007, 65, 29–37. [Google Scholar]
- Chlup, P.H.; Wang, T.; Lee, E.G.; Stewart, G.G. Assessment of the physiological status of yeast during high- and low-gravity wort fermentations determined by flow cytometry. Tech. Quart. Master Brew. Assoc. Am. 2007, 44, 286–295. [Google Scholar] [CrossRef]
- Schlee, C.; Miedl, M.; Leiper, K.A.; Stewart, G.G. The potential of confocal imaging for measuring physiological changes in brewer’s yeast. J. Inst. Brew. 2006, 112, 134–147. [Google Scholar] [CrossRef]
Figure 1.
Static fermentation flocculation characteristics [1].
Figure 1.
Static fermentation flocculation characteristics [1].

Figure 2.
A disc stack centrifuge (5 hL per hour) [2].
Figure 2.
A disc stack centrifuge (5 hL per hour) [2].

Figure 3.
Lectin theory of flocculation. Protein lectins on the yeast cell surface interact with either mannose-containing and/or glucose-containing carbohydrate determinants on the cell walls of adjacent cells only in the presence of calcium [15].
Figure 3.
Lectin theory of flocculation. Protein lectins on the yeast cell surface interact with either mannose-containing and/or glucose-containing carbohydrate determinants on the cell walls of adjacent cells only in the presence of calcium [15].

Figure 4.
Flocculation characteristics of Saccharomyces cerevisiae strains. (A) Flocculent culture; (B) non-flocculent culture [10].
Figure 4.
Flocculation characteristics of Saccharomyces cerevisiae strains. (A) Flocculent culture; (B) non-flocculent culture [10].


Figure 6.
(A) Helm’s sedimentation flocculation test—co-flocculation [28,29]; (B) ale yeast co-flocculation—2-L cylinder wort fermentation test [29].

Figure 7.
Schematic diagram of the architecture yeast cell wall. Components such as mannoprotein occur as a matrix and are distributed throughout the entire wall; therefore, the layered arrangement should show zones of enrichment [15].
Figure 7.
Schematic diagram of the architecture yeast cell wall. Components such as mannoprotein occur as a matrix and are distributed throughout the entire wall; therefore, the layered arrangement should show zones of enrichment [15].
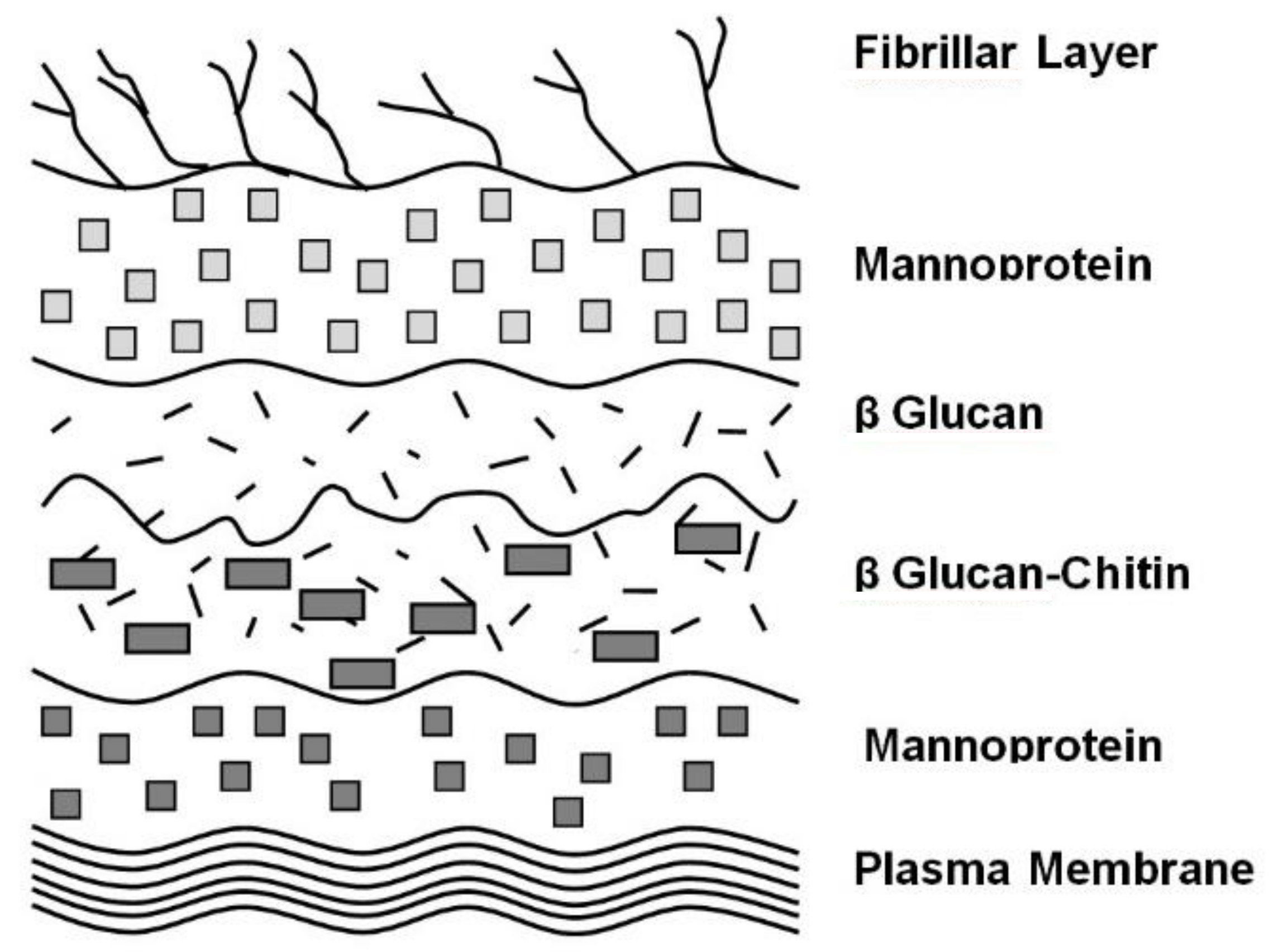
Figure 8.
Electron photomicrograph of Saccharomyces cerevisiae flocculent and non-flocculent strains shadow-cast with tungsten oxide [33].
Figure 8.
Electron photomicrograph of Saccharomyces cerevisiae flocculent and non-flocculent strains shadow-cast with tungsten oxide [33].
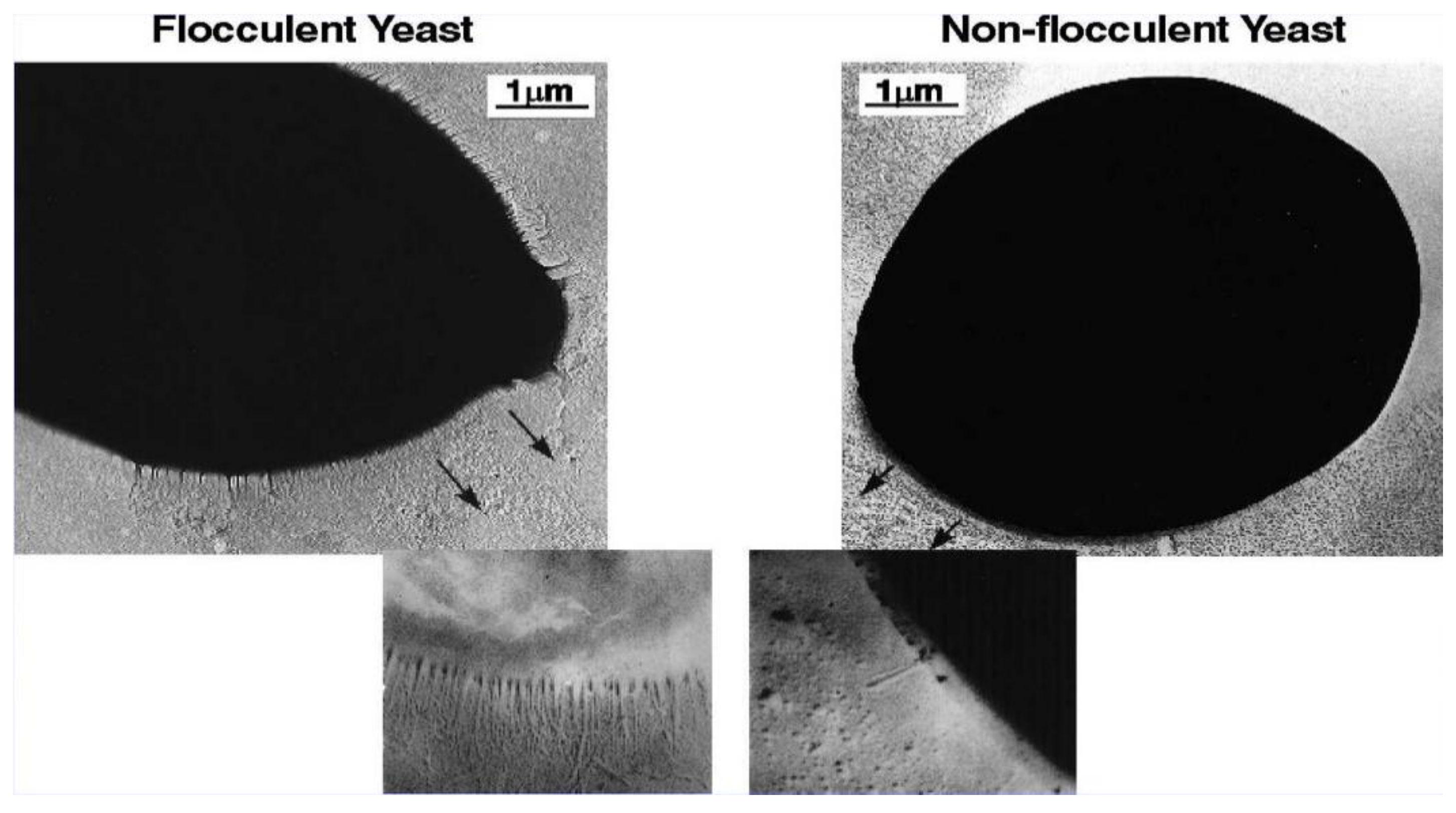
Figure 9.
Micrograph of a biofilm of microorganisms (yeast and bacteria) on the inner surface of a beer dispersing line [12].
Figure 9.
Micrograph of a biofilm of microorganisms (yeast and bacteria) on the inner surface of a beer dispersing line [12].

Figure 10.
Electron photomicrographs of adhering and non-adhering cultures of Candida albicans (photograph courtesy of L. J. Douglas) [13].
Figure 10.
Electron photomicrographs of adhering and non-adhering cultures of Candida albicans (photograph courtesy of L. J. Douglas) [13].
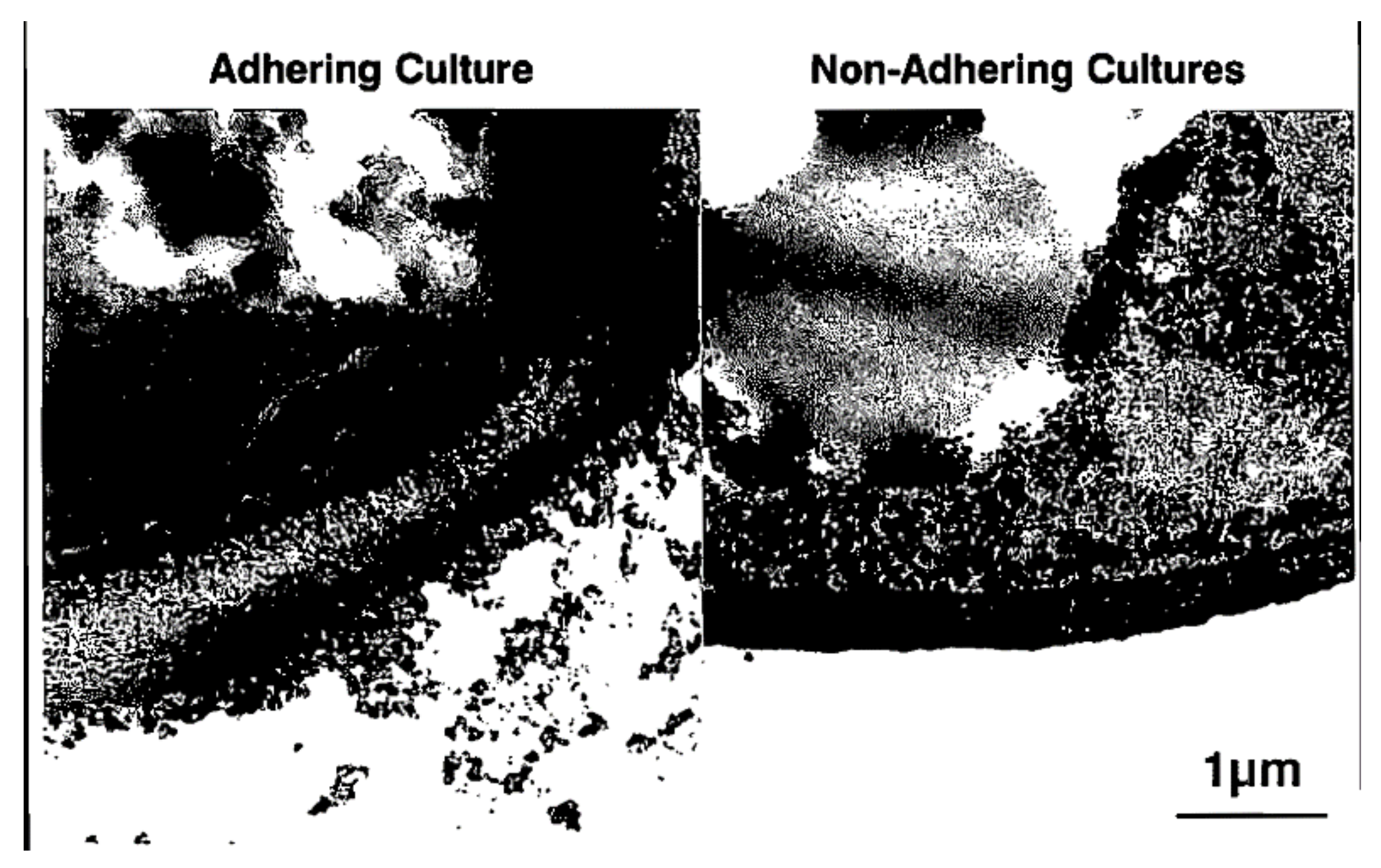
Figure 11.
Co-flocculation between an ale yeast strain and a Lactobacillus fermentum strain—bacterial-induced yeast flocculation [40].
Figure 11.
Co-flocculation between an ale yeast strain and a Lactobacillus fermentum strain—bacterial-induced yeast flocculation [40].

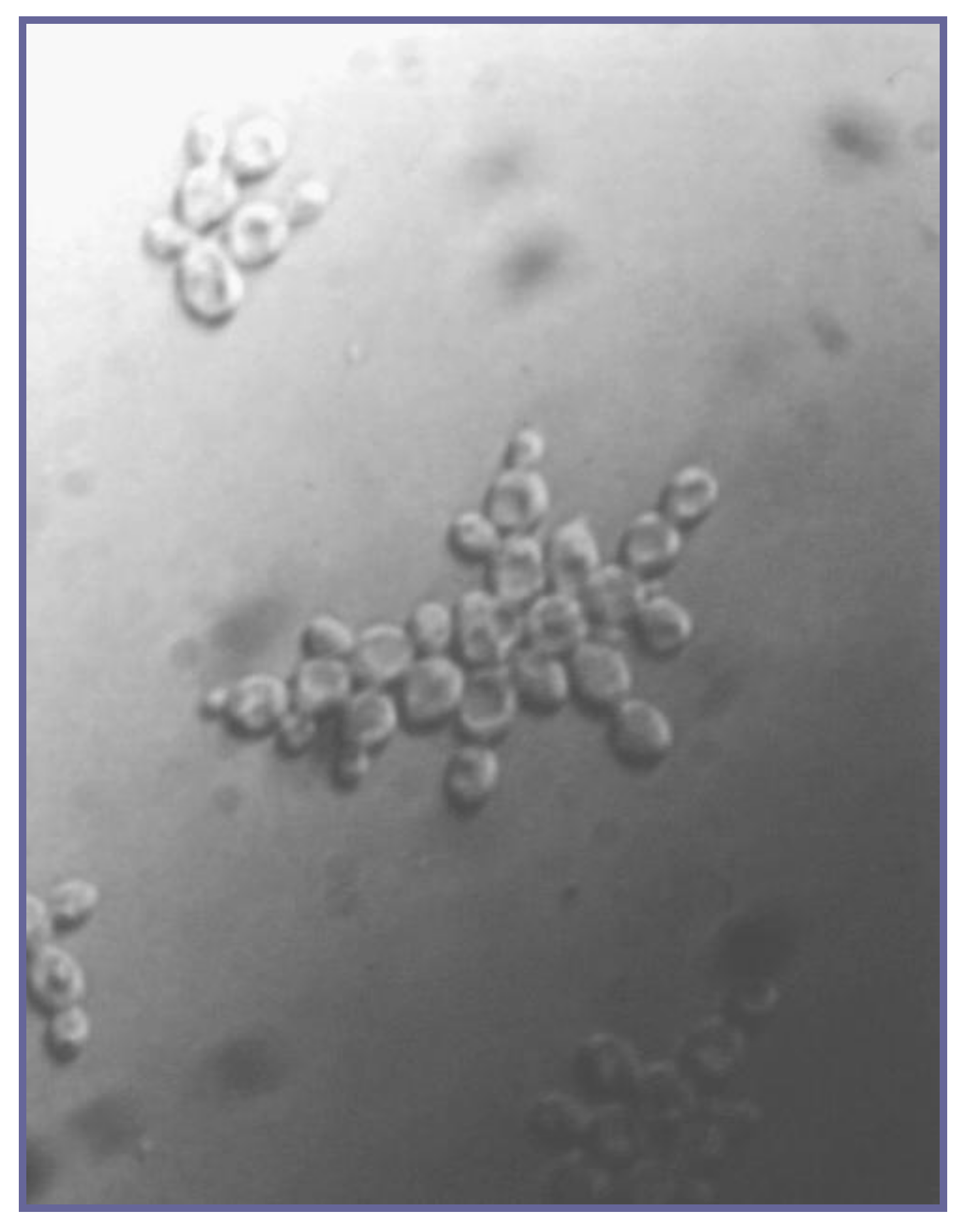
Figure 13.
Giant colony morphologies of brewer’s yeast strains (A,B) exhibit the co-flocculation phenotype [16,29,71].

Figure 14.
Test tube-sized (15 mL) fermentations of a premature yeast flocculation (PYF) and control malt yeast in suspension continuously measured [119].
Figure 14.
Test tube-sized (15 mL) fermentations of a premature yeast flocculation (PYF) and control malt yeast in suspension continuously measured [119].
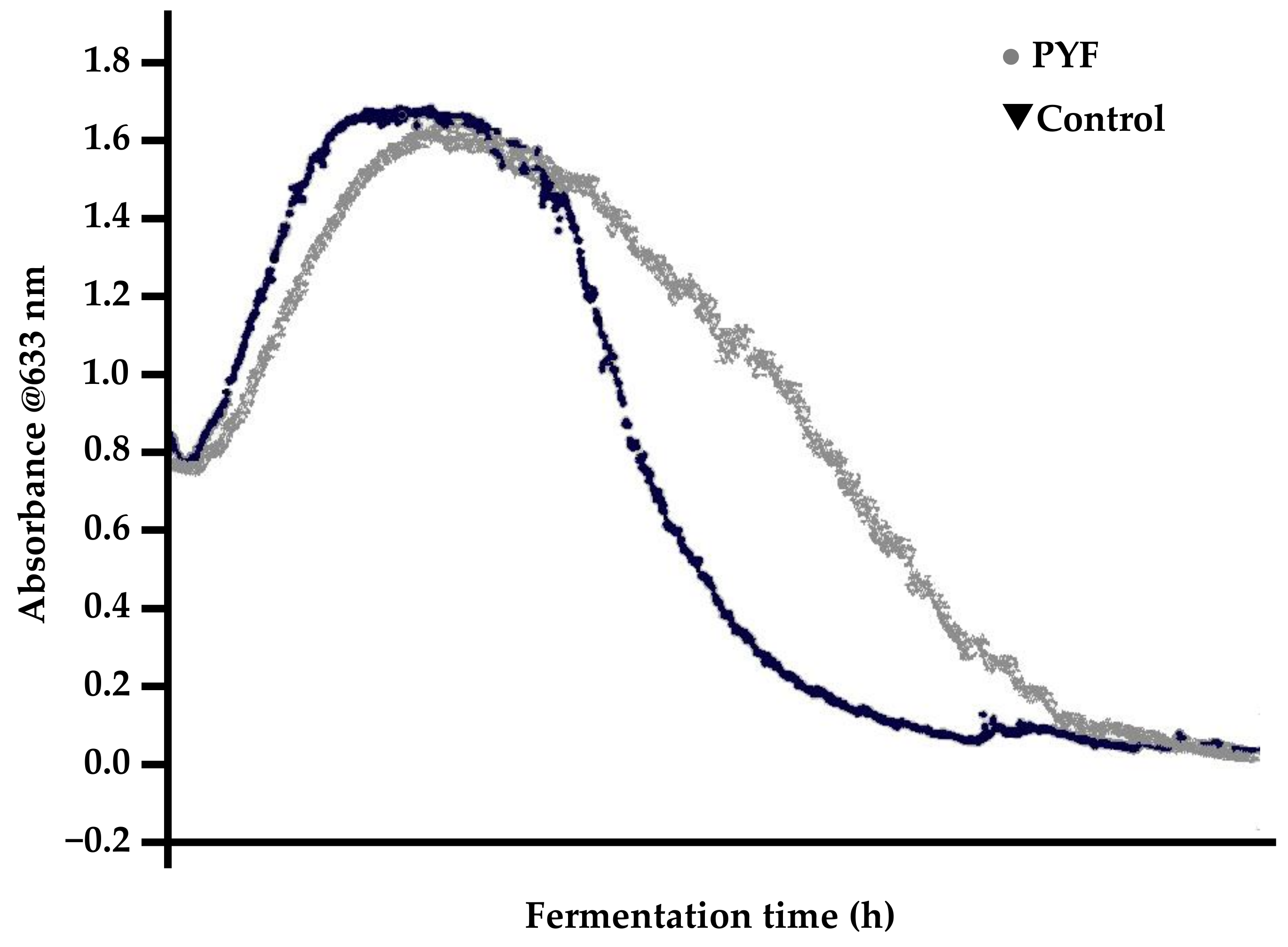
Figure 15.
De-flocculation of yeast as a result of repeated water washing and re-flocculation following the addition of calcium ions [60].
Figure 15.
De-flocculation of yeast as a result of repeated water washing and re-flocculation following the addition of calcium ions [60].
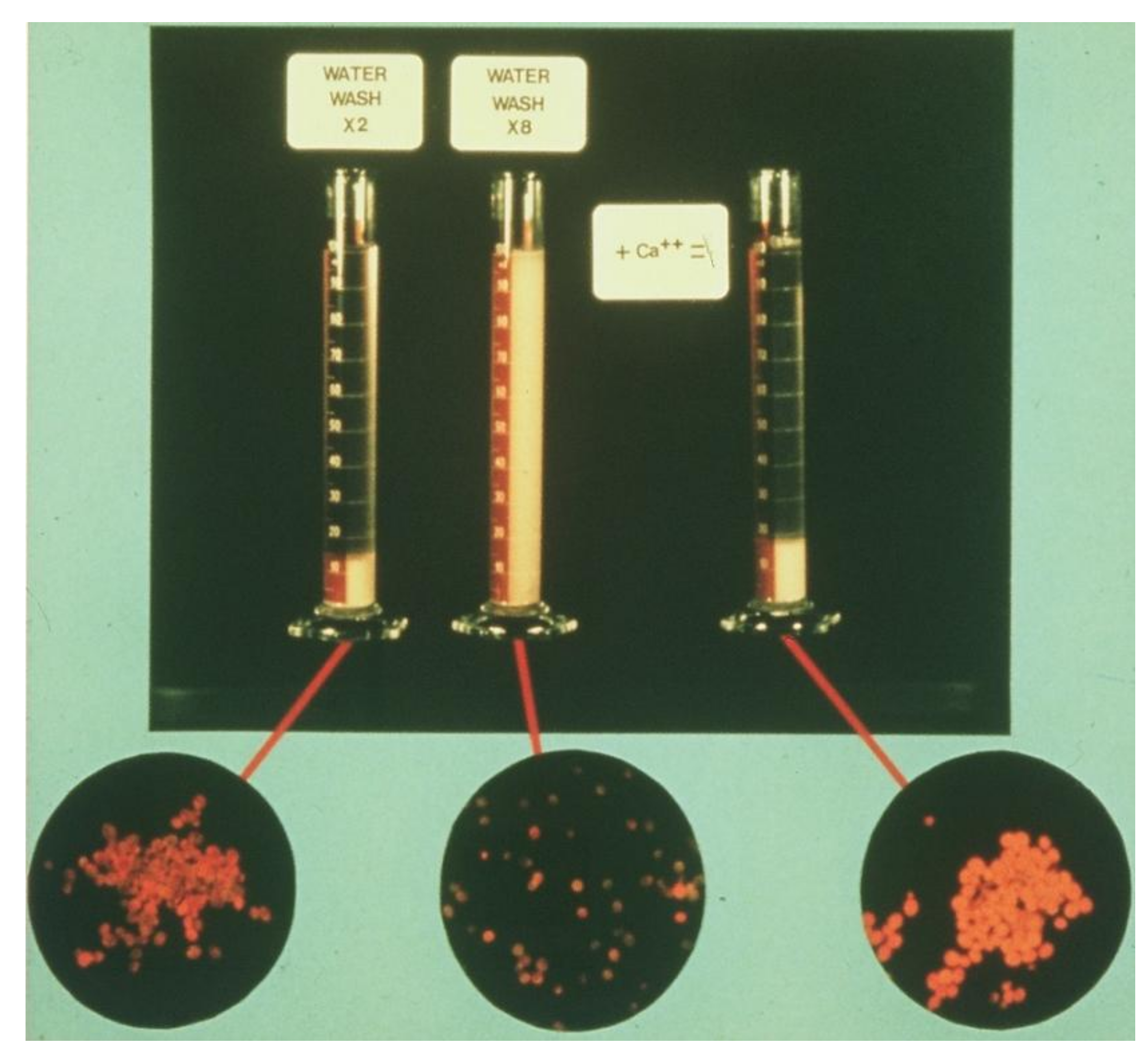
Figure 16.
The disc stack in a centrifuge and the flow pattern [138].
Figure 16.
The disc stack in a centrifuge and the flow pattern [138].

Figure 17.
Environment scanning electron microscope (ESEM) of ale yeast strain damage: (A) Cells before passage through a disc centrifuge, and (B) cells following passage through a disc centrifuge operating at high G-force [141].
Figure 17.
Environment scanning electron microscope (ESEM) of ale yeast strain damage: (A) Cells before passage through a disc centrifuge, and (B) cells following passage through a disc centrifuge operating at high G-force [141].

Figure 18.
Factors which promote yeast stress [7].
Figure 18.
Factors which promote yeast stress [7].

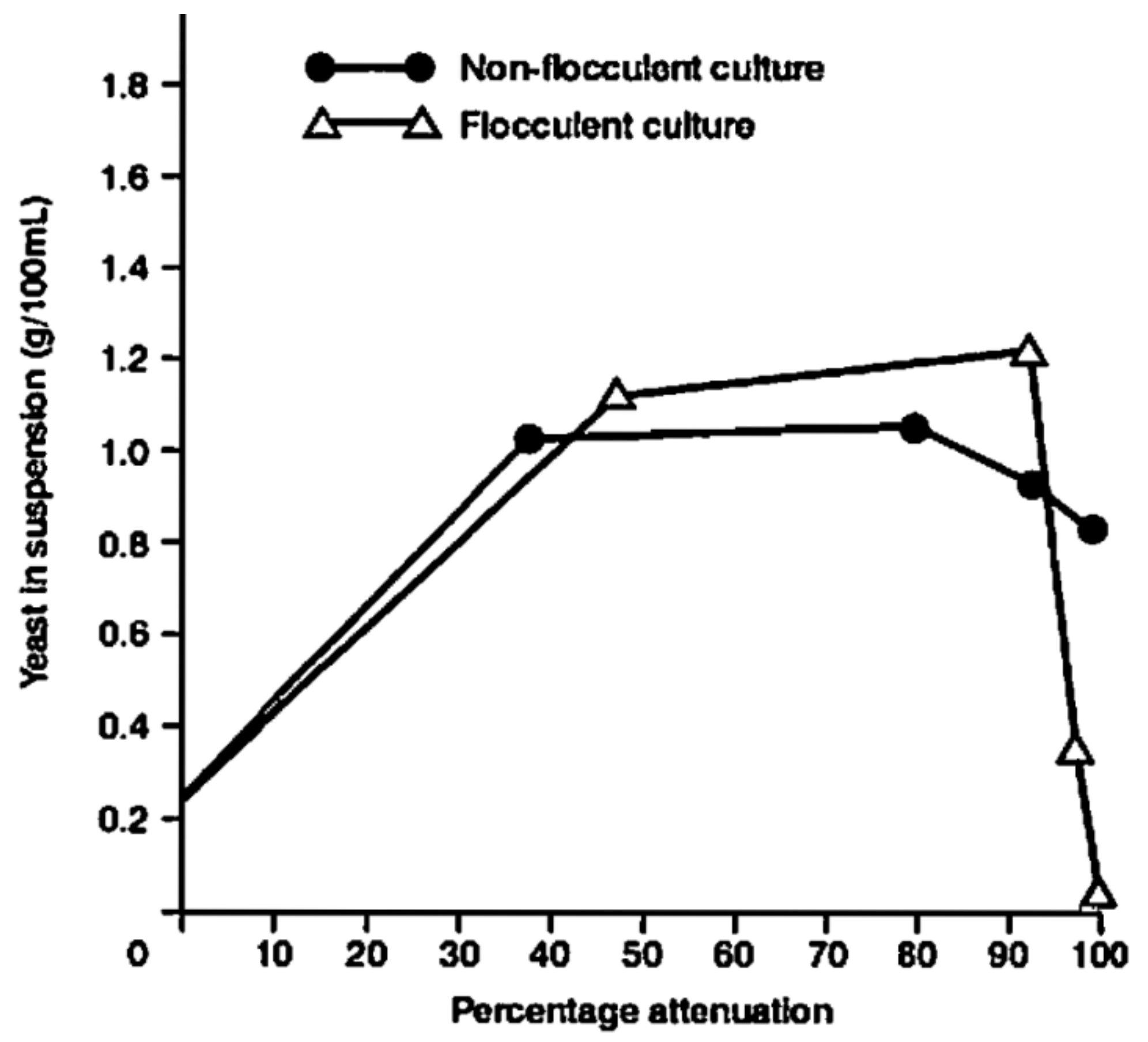{kind=link}
{kind=link}
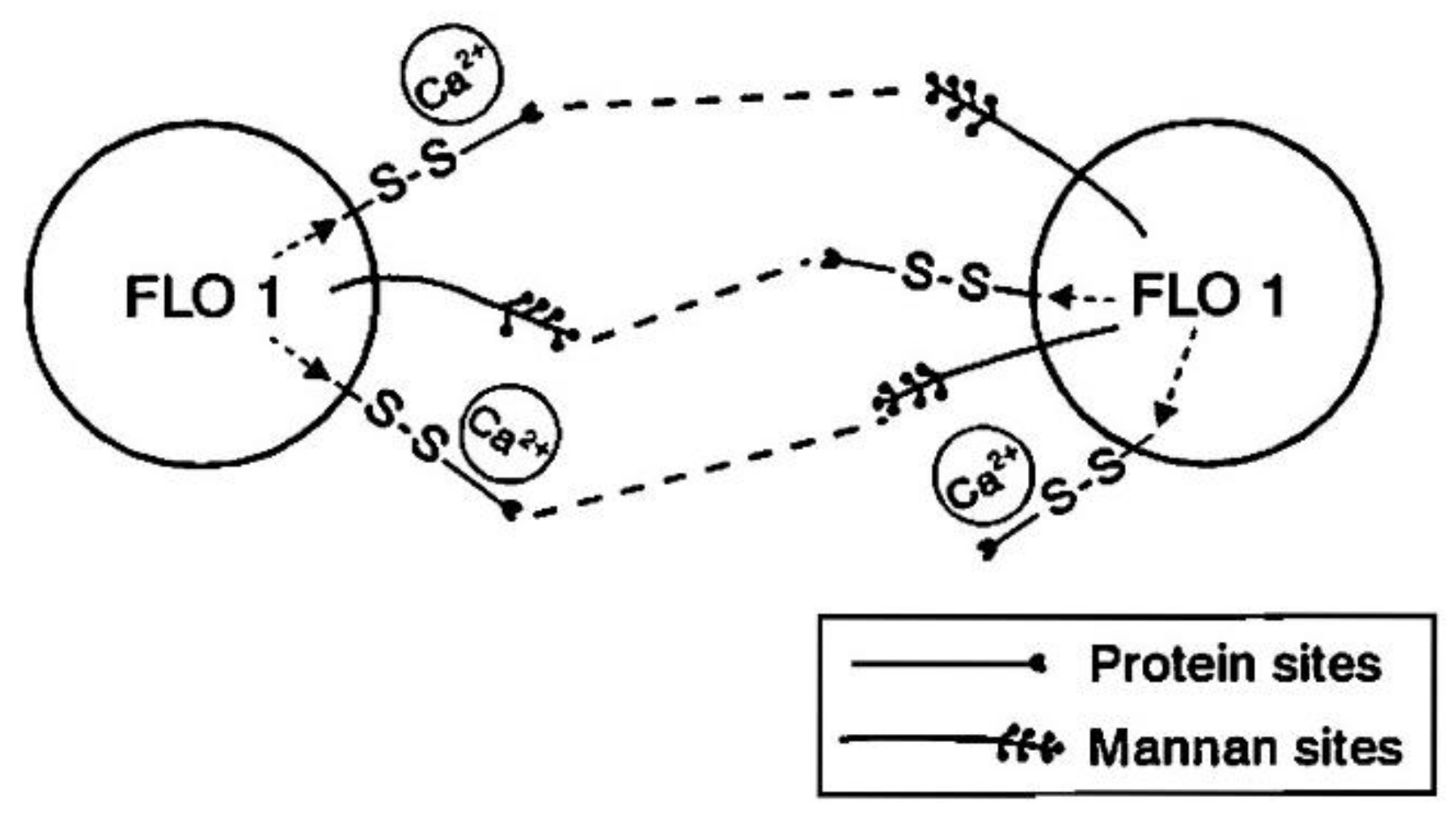{kind=link}
{kind=link}
{kind=link}
{kind=link}
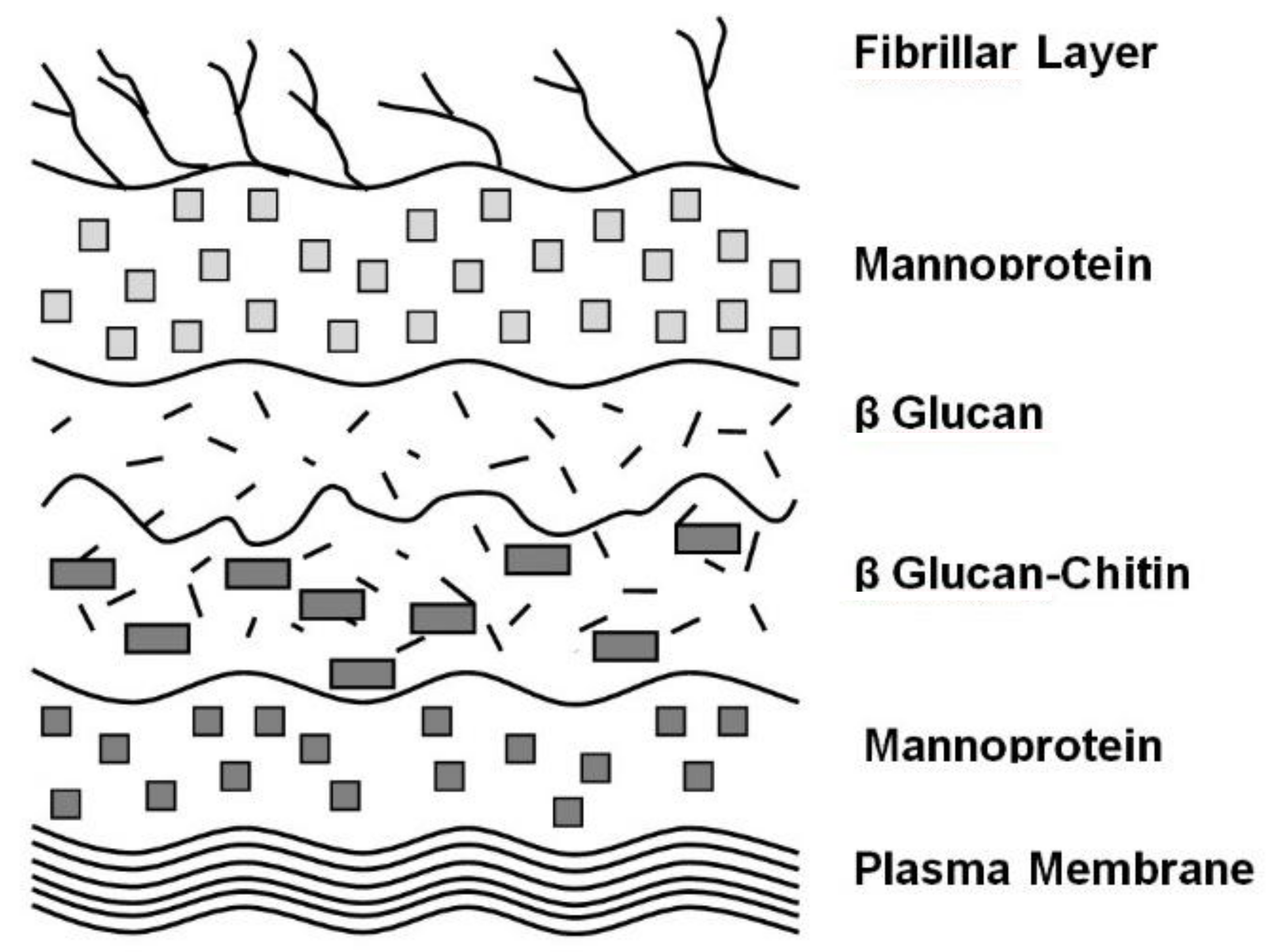{kind=link}
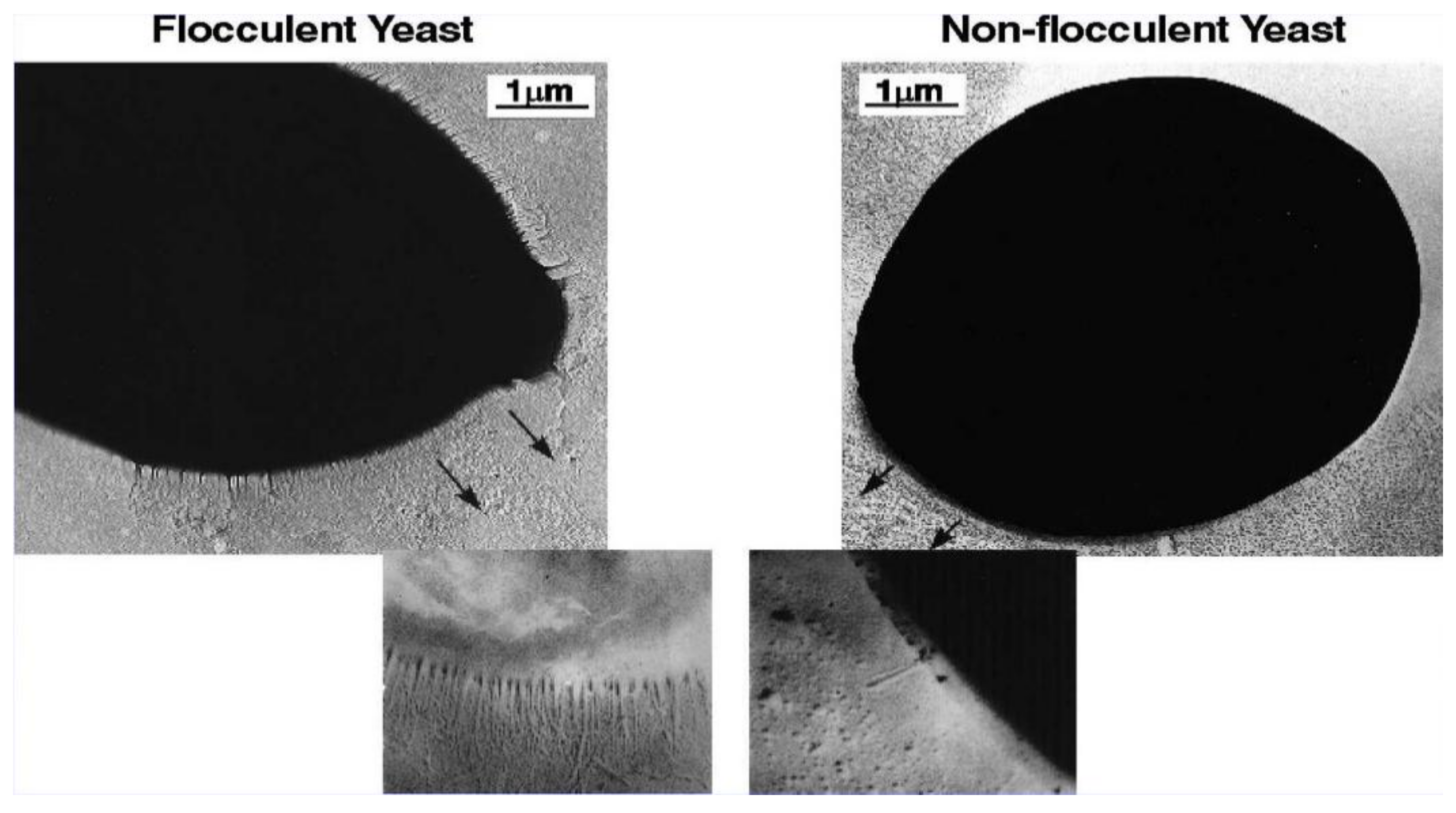{kind=link}
{kind=link}
{kind=link}
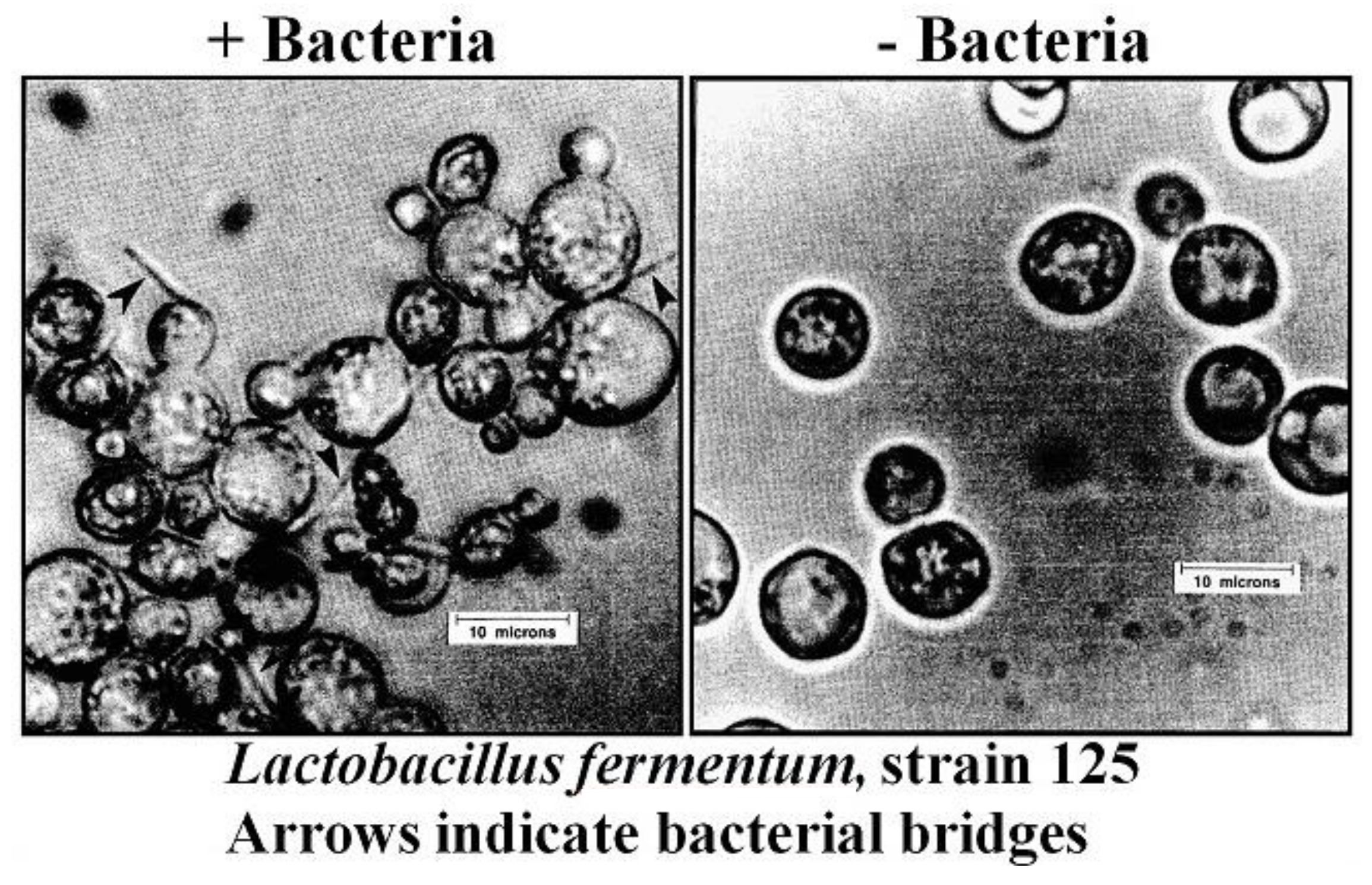{kind=link}
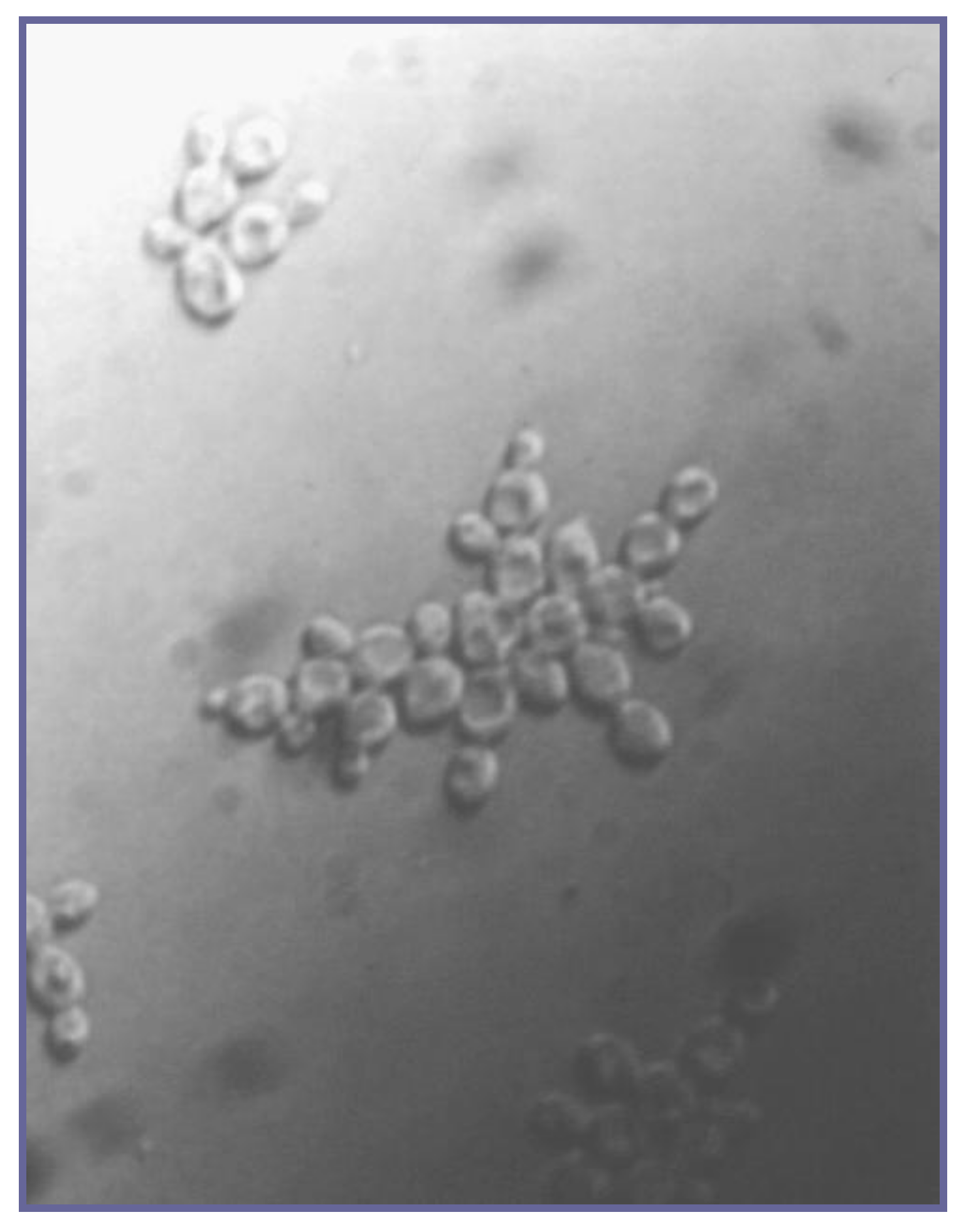{kind=link}
{kind=link}
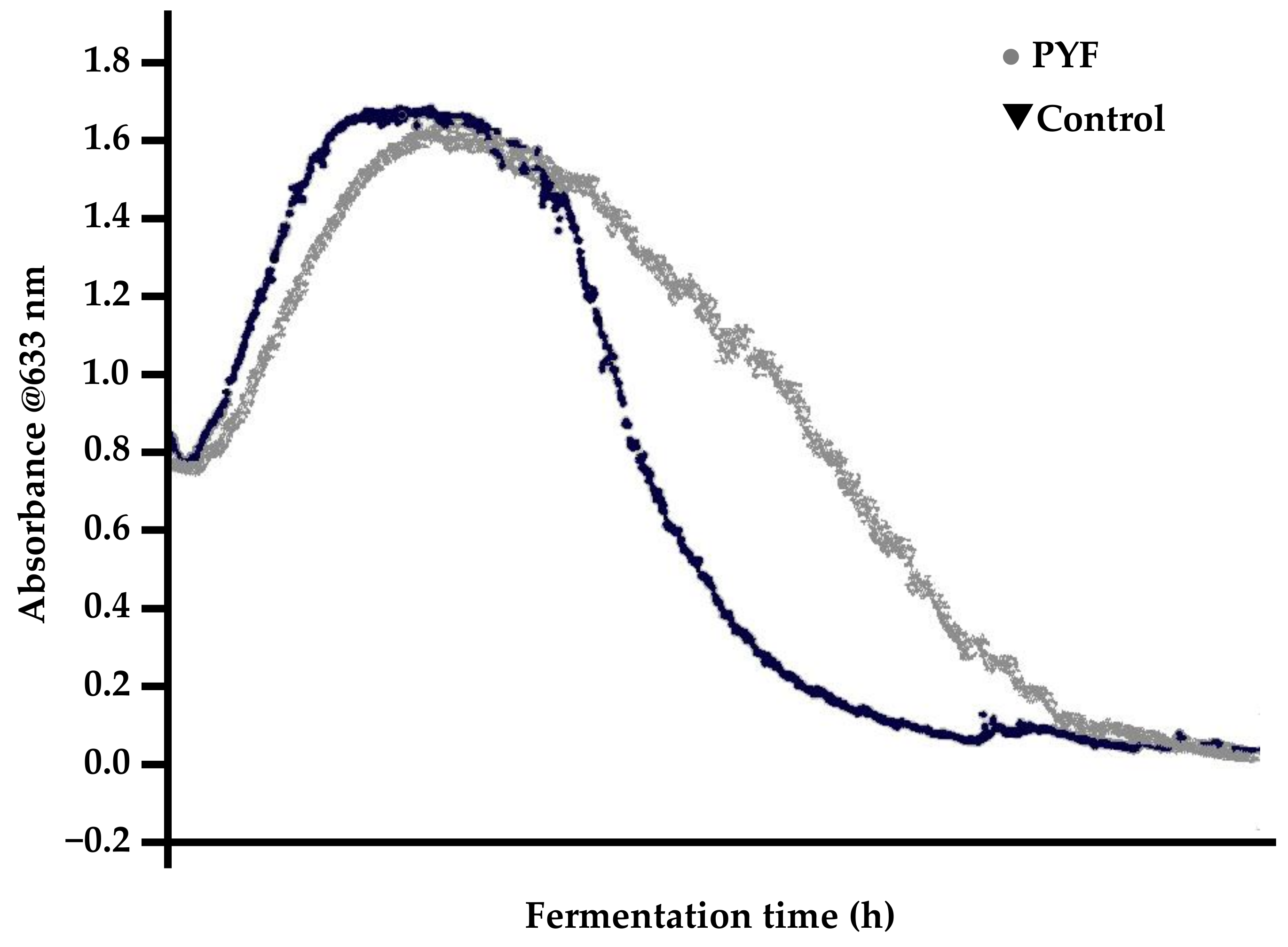{kind=link}
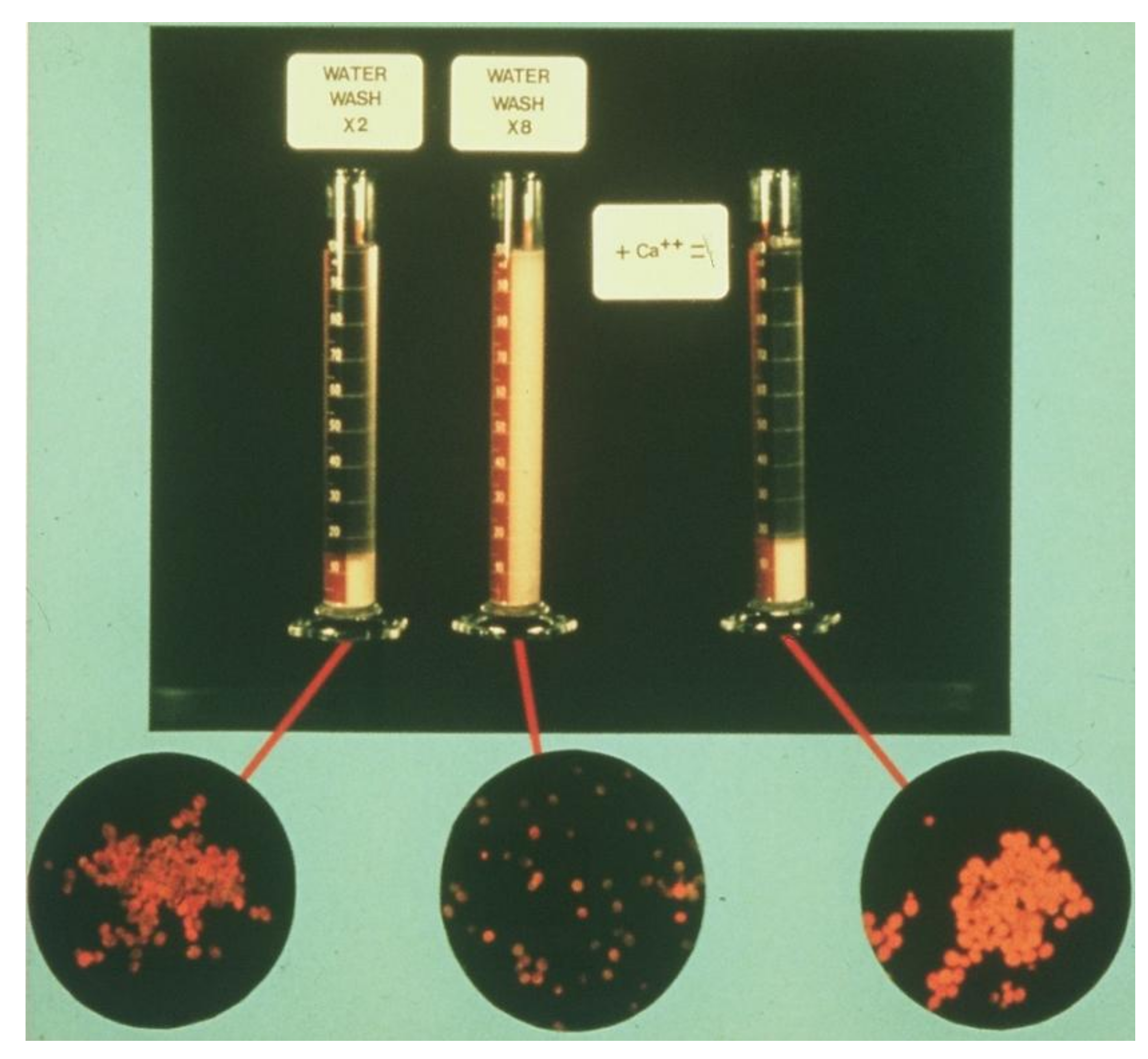{kind=link}
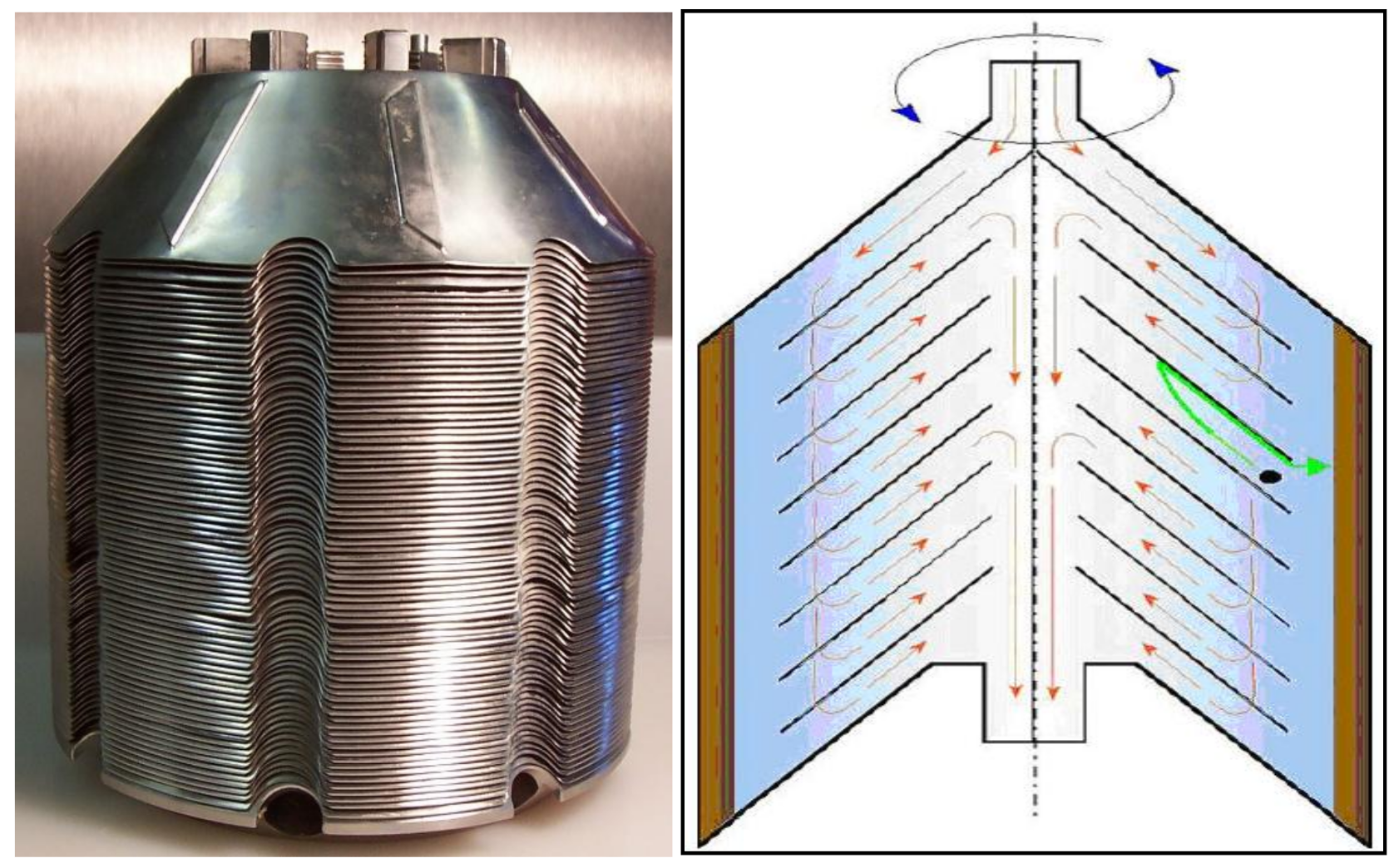{kind=link}
{kind=link}
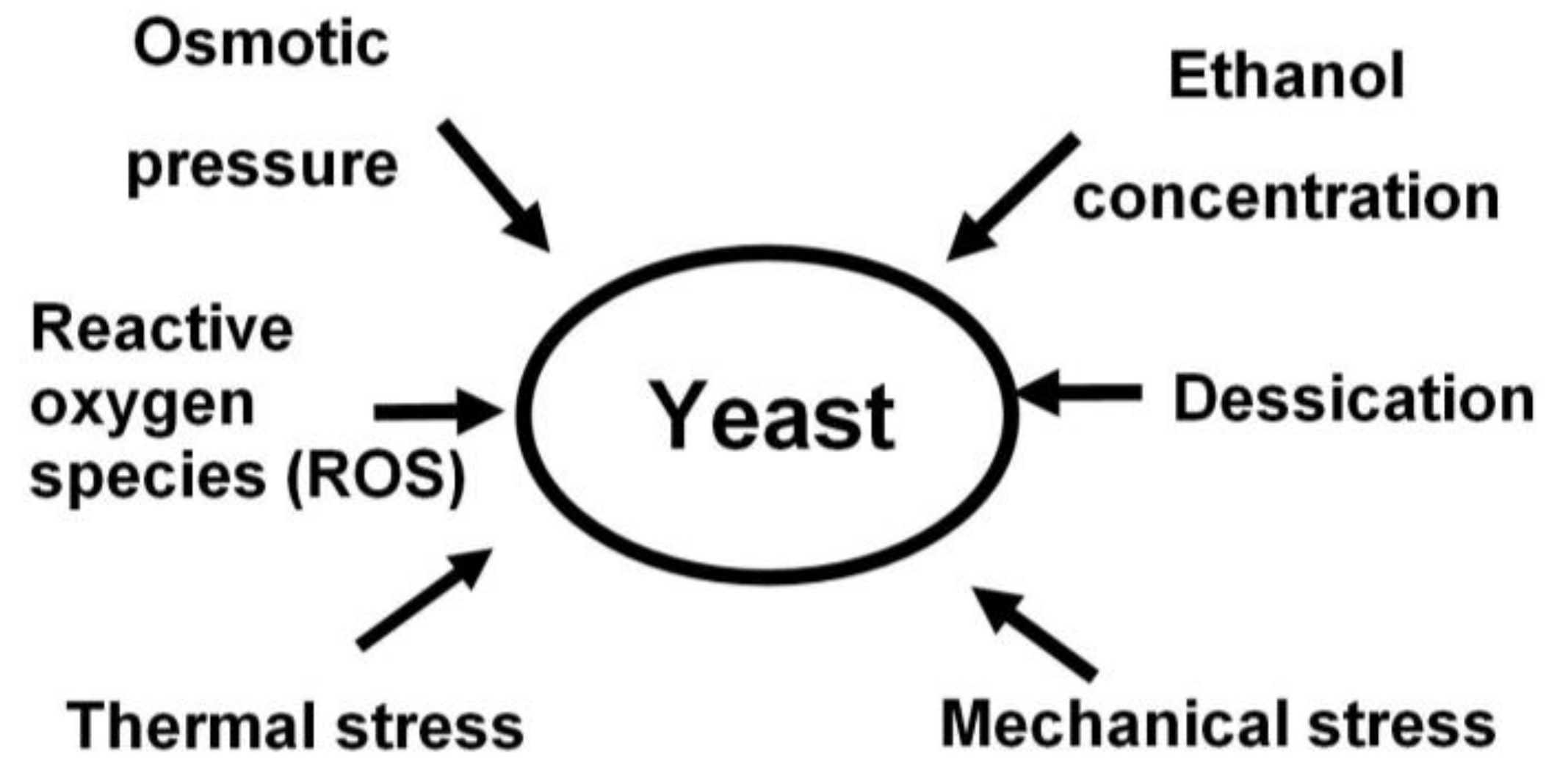{kind=link}
Table 1.
Current view of flocculation phenotypes [89].
Table 1.
Current view of flocculation phenotypes [89].
| Genes | Character | Sugars that Inhibit Flocculation |
|---|---|---|
| FLO1, FLO5, FLO9, FLO10 | Strong Flo1 phenotype | Only mannose |
| FL08 | Regulation of other FLO genes | Unknown |
| Lg-FLO1 | NewFlo phenotype | Mannose, glucose, sucrose, maltose, and maltotriose (not galactose) |
| FLO11 | Chain formation—pseudohyphae | No inhibition by sugars |
| FLONL, FLONS | Like NewFlo phenotype | Mannose, glucose, sucrose, maltose, maltotriose, and galactose |
| Not known | Mannose-insensitive (MI) flocculation (Ca-independent) | No inhibition by sugars |
Table 2.
Labelled (*) calcium removed from co-flocculent and non-flocculent cultures during de-flocculation washings [60].
Table 2.
Labelled (*) calcium removed from co-flocculent and non-flocculent cultures during de-flocculation washings [60].
| Yeast Culture | Flocculation Characteristic | Total Calcium Washed Off Yeast * | |
|---|---|---|---|
| Ale | S. cerevisiae | Non-flocculent | 18 |
| Non-flocculent | 19 | ||
| Flocculent | 30 | ||
| Flocculent | 42 | ||
| Lager | S. pastorianus | Non-flocculent | 12 |
| Non-flocculent | 14 | ||
| Flocculent | 20 | ||
| Flocculent | 22 | ||
Table 3.
Characteristics of yeast cells recycled nine times in 20 °P wort and beer before and after centrifugation at high G-force [138].
Table 3.
Characteristics of yeast cells recycled nine times in 20 °P wort and beer before and after centrifugation at high G-force [138].
| Characteristic | Before Centrifugation | After Centrifugation |
|---|---|---|
| Viability (%) | 85 | 42 |
| Extracellular pH | 4.2 | 6.0 |
| Intracellular pH | 5.8 | 5.3 |
| Damaged cells (%) | 4 | 15 |
| Glycogen (ppm) | 18 | 8 |
| Trehalose (ppm) | 22 | 6 |
| Mannan released (counts) | 400 | 1000 |
| Proteinase A (U/mL) | 3.1 | 6.2 |
| Hydrophobic polypeptides (mg/L) | 48 | 25 |
| Beer foam stability (NIBEM) | 110 | 82 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stewart, G.G. Yeast Flocculation—Sedimentation and Flotation. Fermentation 2018, 4, 28. https://doi.org/10.3390/fermentation4020028
AMA Style
Stewart GG. Yeast Flocculation—Sedimentation and Flotation. Fermentation. 2018; 4(2):28. https://doi.org/10.3390/fermentation4020028
Chicago/Turabian StyleStewart, Graham G. 2018. "Yeast Flocculation—Sedimentation and Flotation" Fermentation 4, no. 2: 28. https://doi.org/10.3390/fermentation4020028
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.