
Use of Nutritional Requirements for Brettanomyces bruxellensis to Limit Infections in Wine

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Nutritional Requirements
2.3. Biotin Removal
2.4. Statistics
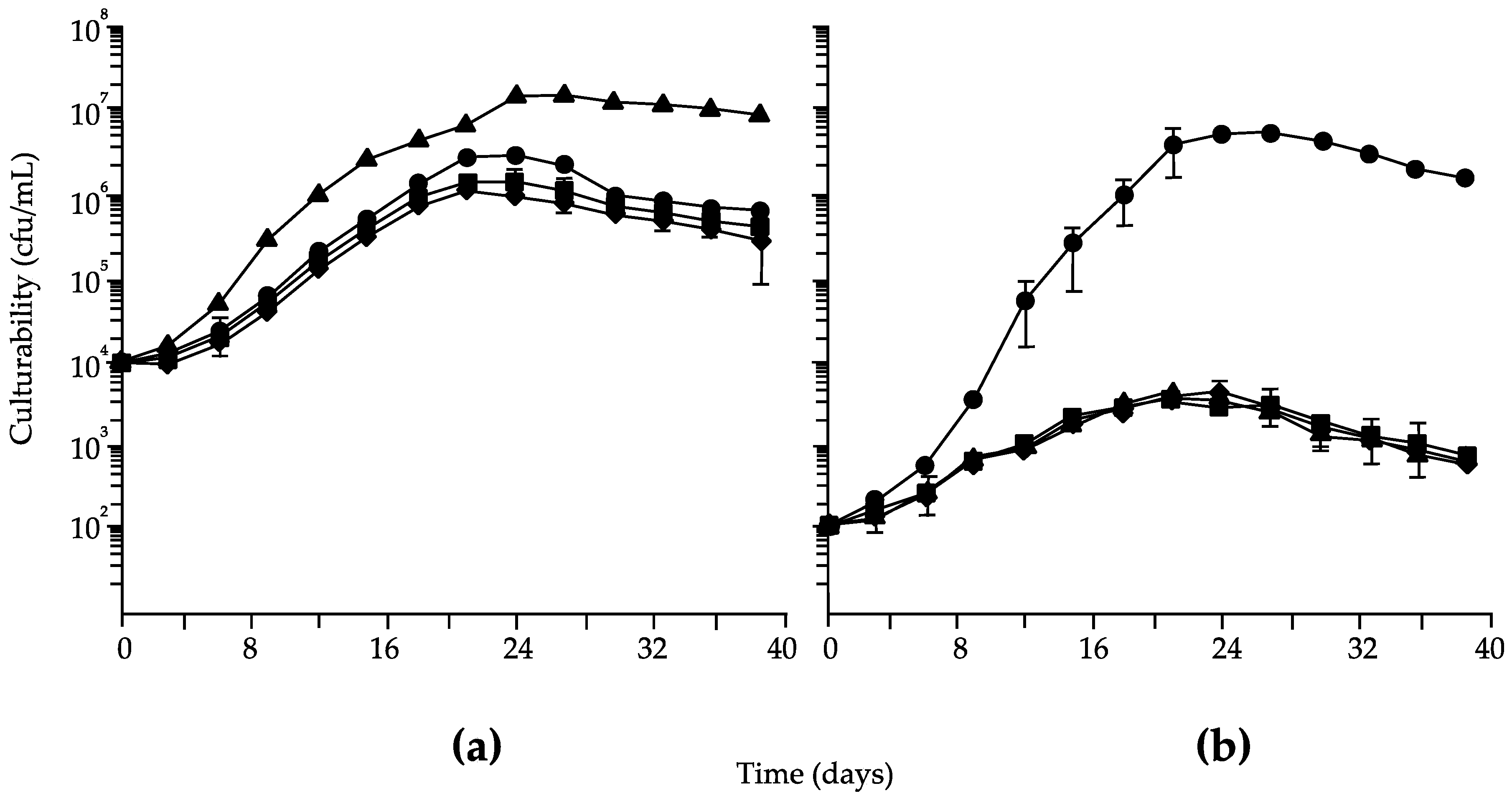
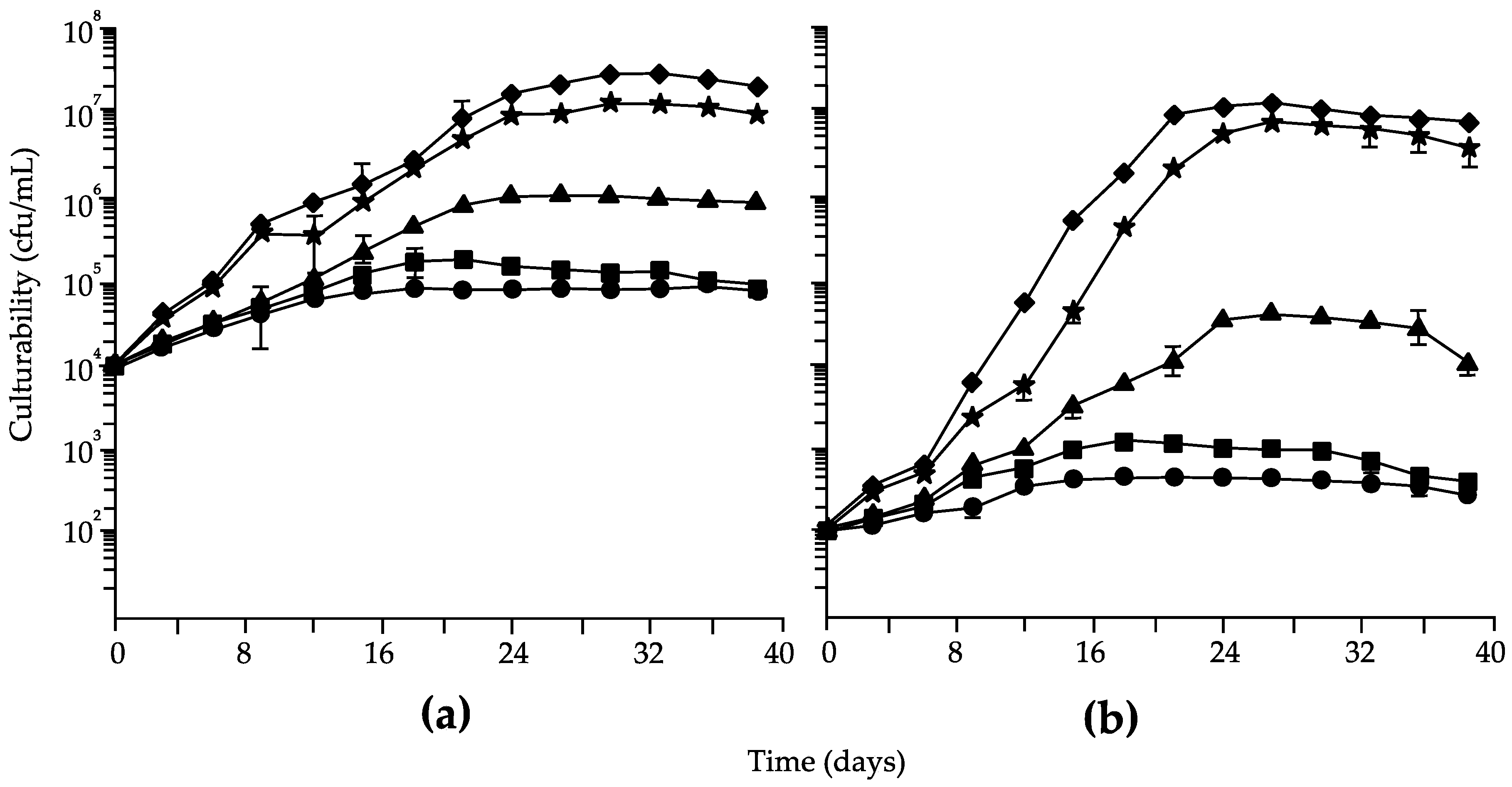
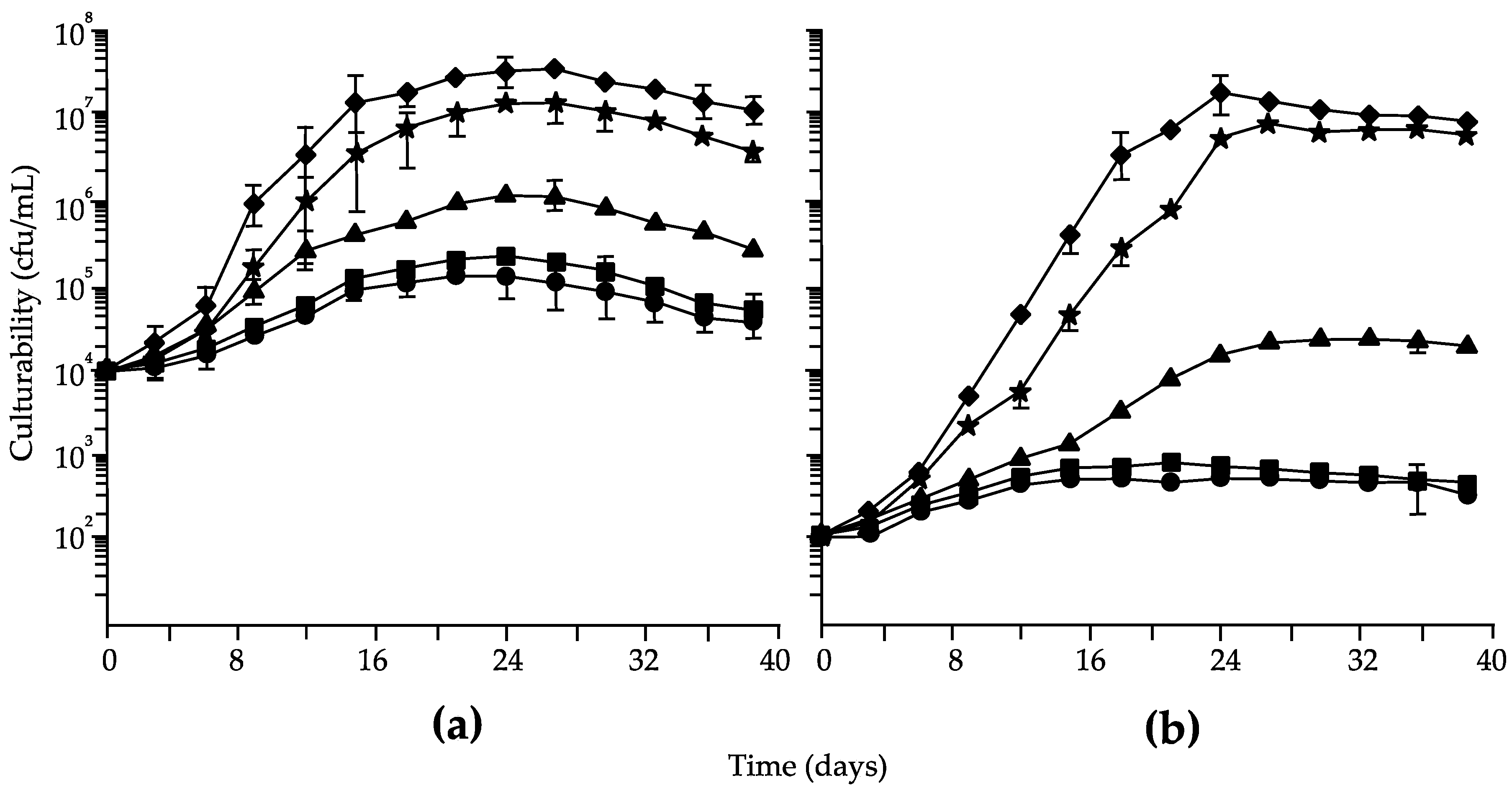
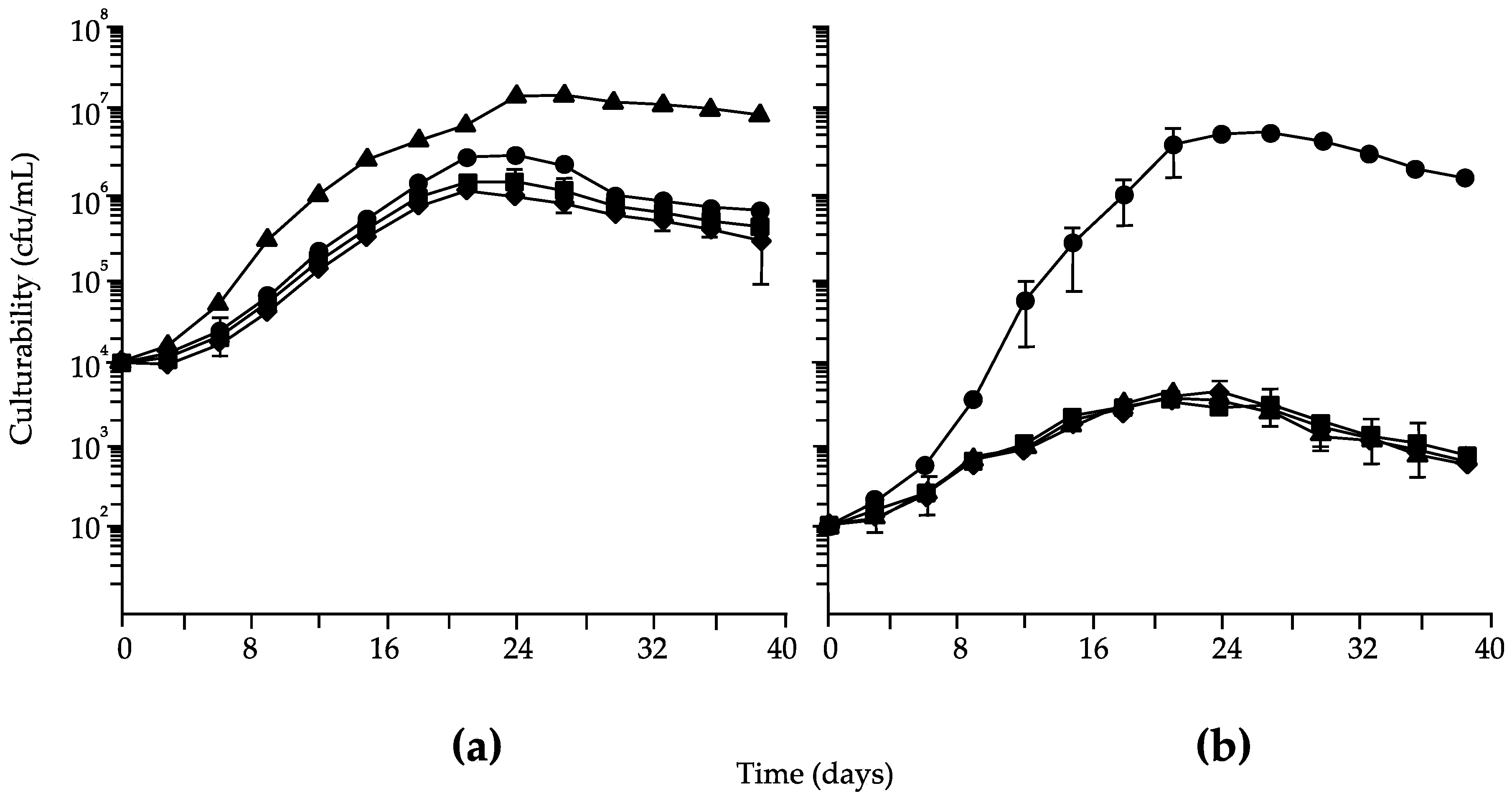
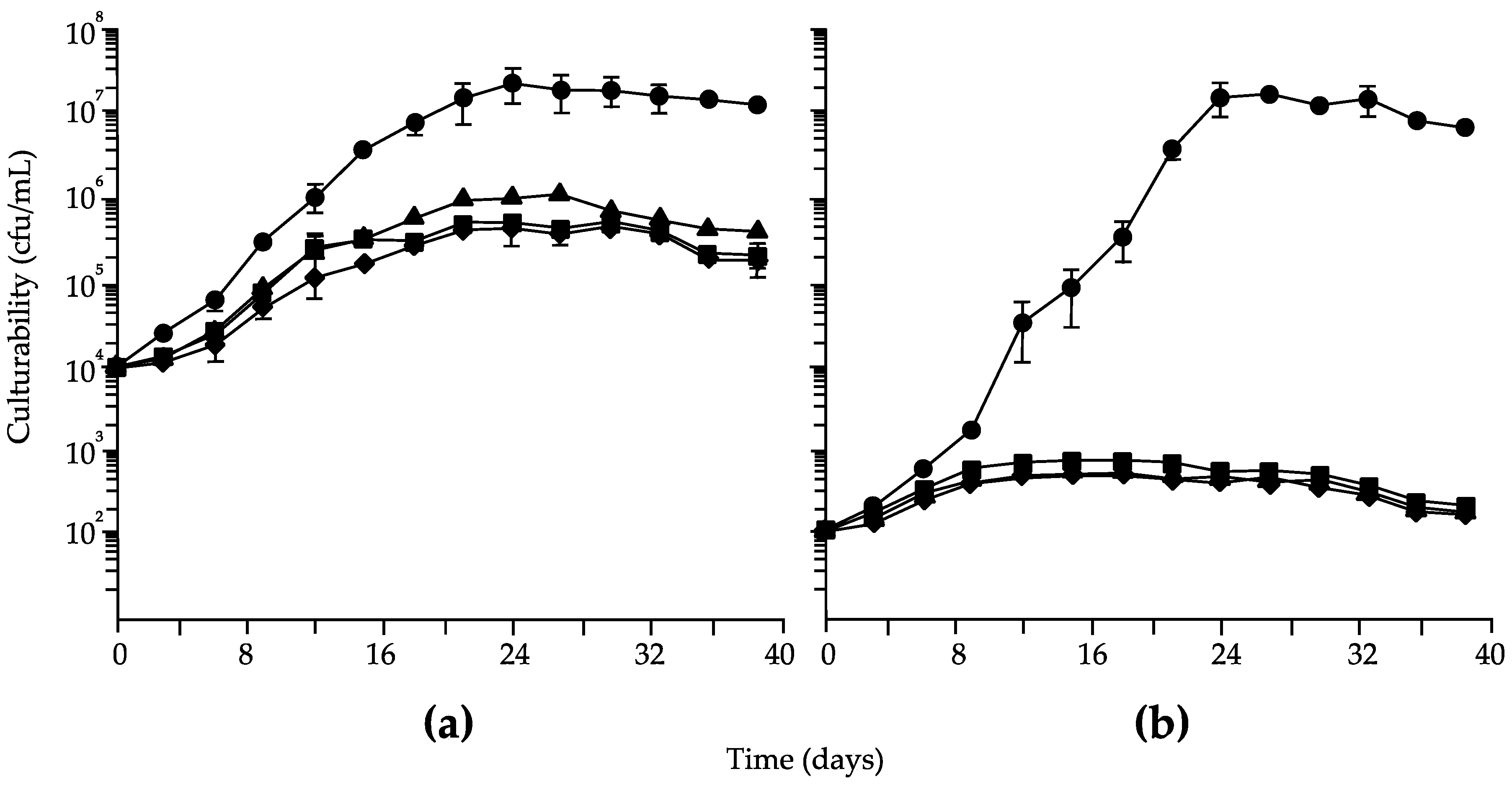
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MWCO | Molecular Weight Cut–Off |
| cfu | colony forming units |
References
- Fugelsang, K.C.; Edwards, C.G. Wine Microbiology: Practical Applications and Procedures, 2nd ed.; Springer Science and Business Media: New York, NY, USA, 2007. [Google Scholar]
- Chatonnet, P.; Duborudieu, D.; Boidron, J.N.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Fugelsang, K.C.; Zoecklein, B.W. Population dynamics and effects of Brettanomyces bruxellensis strains on Pinot noir (Vitis vinifera L.) wines. Am. J. Enol. Vitic. 2003, 54, 294–300. [Google Scholar]
- Romano, A.; Perello, M.C.; Lonvaud-Funel, A.; Sicard, G.; de Revel, G. Sensory and analytical re-evaluation of “Brett character”. Food Chem. 2009, 114, 15–19. [Google Scholar] [CrossRef]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N. The influence of Brettanomyces/Dekkera sp. yeasts and lactic acid bacteria on the ethylphenol content of red wines. Am. J. Enol. Vitic. 1995, 46, 463–468. [Google Scholar]
- Barata, A.; Pagliara, D.; Piccininno, T.; Tarantino, F.; Ciardulli, W.; Malfeito-Ferreira, M.; Loureiro, V. The effect of sugar concentration and temperature on growth and volatile phenol production by Dekkera bruxellensis in wine. FEMS Yeast Res. 2008, 7, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.C.; Bohlscheid, J.C.; Edwards, C.G. Impact of available nitrogen and sugar concentration in musts on alcoholic fermentation and subsequent wine spoilage by Brettanomyces bruxellensis. Food Microbiol. 2015, 46, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Conterno, L.; Joseph, C.M.L.; Arvik, T.J.; Henick-Kling, T.; Bisson, L.F. Genetic and physiological characterization of Brettanomyces bruxellensis strains isolated from wines. Am. J. Enol. Vitic. 2006, 57, 139–147. [Google Scholar]
- Burkholder, P.R.; McVeigh, I.; Moyer, D. Studies on some growth factors of yeasts. J. Bacteriol. 1944, 48, 385–391. [Google Scholar] [PubMed]
- Van der Walt, J.P.; van Kerken, A.E. The wine yeasts of the Cape. Part II. The occurrence of Brettanomyces intermedius and Brettanomyces schanderlii in South African table wines. Antonie Leeuwenhoek 1959, 25, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Wickerham, L.J. Taxonomy of Yeasts; United States Department of Agriculture: Washington, DC, USA, 1951.
- Crauwels, S.; Zhu, B.; Steensels, J.; Busschaert, P.; Samblanx, G.D.; Marchal, K.; Willems, K.A.; Verstrepen, K.J.; Lievens, B. Assessing genetic diversity among Brettanomyces yeasts by DNA fingerprinting and whole-genome sequencing. Appl. Environ. Microbiol. 2014, 80, 4398–4413. [Google Scholar] [CrossRef] [PubMed]
- Conterno, L.; Aprea, E.; Franceschi, P.; Viola, R.; Vrhovsek, U. Overview of Dekkera bruxellensis behaviour in an ethanol-rich environment using untargeted and targeted metabolomic approaches. Food Res. Int. 2013, 51, 670–678. [Google Scholar] [CrossRef]
- Chandra, M.; Barata, A.; Ferreira-Dias, S.; Malfeito-Ferreira, M.; Loureiro, V. A response surface methodology study on the role of factors affecting growth and volatile phenol production by Brettanomyces bruxellensis ISA 2211 in wine. Food Microbiol. 2014, 42, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Caldeira, J.; Boteheiro, R.; Pagliara, D.; Malfeito-Ferreira, M.; Loureiro, V. Survival patterns of Dekkera bruxellensis in wines and inhibitory effect of sulfur dioxide. Int. J. Food Microbiol. 2008, 121, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.L.; Umiker, N.L.; Arneborg, N.; Edwards, C.G. Identification and characterization of Dekkera bruxellensis, Candida pararugosa, and Pichia guilliermondii isolated from commercial red wines. Food Microbiol. 2009, 26, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, J.; Eberhard, T.; Schnürer, J.; Passoth, V. Fermentation characteristic of Dekkera bruxellensis strains. Appl. Microbiol. Biotechnol. 2010, 87, 1487–1497. [Google Scholar] [CrossRef] [PubMed]
- Leistner, L. Basic aspects of food preservation by hurdle technology. Int. J. Food Microbiol. 2000, 55, 181–186. [Google Scholar] [CrossRef]
- Zuehlke, J.M.; Edwards, C.G. Impact of sulfur dioxide and temperature on culturability and viability of Brettanomyces in wine. J. Food Prot. 2013, 76, 2024–2030. [Google Scholar] [CrossRef] [PubMed]
- Umiker, N.L.; DeScenzo, R.; Lee, J.; Edwards, C.G. Removal of Brettanomyces bruxellensis from red wine using membrane filtration. J. Food Proc. Pres. 2013, 37, 799–805. [Google Scholar] [CrossRef]
- Brandam, C.; Castro-Martine, C.; Delia, M.L.; Ramon-Portugal, F.; Strehaiano, P. Effect of temperature on Brettanomyces bruxellensis: Metabolic and kinetic aspects. Can. J. Microbiol. 2008, 54, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzzon, R.; Nardin, T.; Micheletti, O.; Nicolini, G.; Larcher, R. Antimicrobial activity of ozone. Effectiveness against the main wine spoilage microorganisms and evaluation of impact on simple phenols in wine. Aust. J. Grape Wine Res. 2013, 19, 180–188. [Google Scholar] [CrossRef]
- Puertolas, E.; Lopez, N.; Condon, S.; Raso, J.; Alvarez, I. Pulsed electric fields inactivation of wine spoilage yeast and bacteria. Int. J. Food Microbiol. 2009, 130, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Lustrato, G.; Vigentini, I.; de Leonardis, A.; Alfano, G.; Tirelli, A.; Foschino, R.; Ranalli, G. Inactivation of wine spoilage yeast Dekkera bruxellensis using low electric current treatment. J. Appl. Microbiol. 2010, 109, 594–604. [Google Scholar] [PubMed]
- Petrova, B.; Cartwright, Z.M.; Edwards, C.G. Effectiveness of chitosan preparations against Brettanomyces bruxellensis grown in culture media and red wines. J. Int. Sci. Vigne Vin 2016, 50, 49–57. [Google Scholar] [CrossRef]
- Renouf, V.; Strehaiano, P.; Lonvaud-Funel, A. Effectiveness of dimethyldicarbonate to prevent Brettanomyces bruxellensis growth in wine. Food Cont. 2008, 19, 208–216. [Google Scholar] [CrossRef]
- Zuehlke, J.M.; Glawe, D.A.; Edwards, C.G. Efficacy of dimethyl dicarbonate against yeasts associated with Washington State grapes and wines. J. Food Proc. Pres. 2015, 39, 1016–1026. [Google Scholar] [CrossRef]
- Zuehlke, J.M.; Petrova, B.; Edwards, C.G. Advances in the control of wine spoilage by Zygosaccharomyces and Dekkera/Brettanomyces. Ann. Rev. Food Sci. Technol. 2013, 4, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.E.; Arroyo-López, F.N.; Garrido-Fernández, A.; Querol, A.; Mercado, L.A.; Ramirez, M.L.; Combina, M. Probabilistic model for the spoilage wine yeast Dekkera bruxellensis as a function of pH, ethanol and free SO2 using time as a dummy variable. Int. J. Food Microbiol. 2014, 170, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: Amsterdam, NY, USA, 2011; pp. 53–986. [Google Scholar]
- Buechsenstein, J.; Ough, C.S. SO2 determination by aeration-oxidation: A comparison with Ripper. Am. J. Enol. Vitic. 1978, 29, 161–164. [Google Scholar]
- Angyal, G. Biotin. In US Food and Drug Administration. Methods for the Microbiological Analysis of Selected Nutrients; Association of Official Analytical Chemists: Arlington, VA, USA, 1996; pp. 9–10. [Google Scholar]
- Madan, M.; Gulati, N. Organic growth factor requirements of some yeasts. Microbios 1980, 28, 167–172. [Google Scholar] [PubMed]
- Aguilar-Uscanga, M.G.; Delia, M.L.; Strehaiano, P. Nutritional requirements of Brettanomyces bruxellensis: Growth and physiology in batch and chemostat cultures. Can. J. Microbiol. 2000, 46, 1046–1050. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.P.; Greenfield, P.F. Ethanol and the fluidity of the yeast plasma membrane. Yeast 1987, 3, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.H.; Harrison, J.S. The Yeasts, 5th ed.; Academic Press: London, UK, 1971; Volume 2, pp. 3–60. [Google Scholar]
- Hertz, R.; Sebrell, W.H. Occurrence of avidin in the oviduct and secretions of the genital tract of several species. Science 1942, 96. [Google Scholar] [CrossRef] [PubMed]
- Korpela, J. Avidin, a high affinity biotin-binding protein, as a tool and subject of biological research. Med. Biol. 1984, 62, 5–26. [Google Scholar] [PubMed]
- Bohlscheid, J.C.; Fellman, J.K.; Wang, X.D.; Ansen, D.; Edwards, C.G. The influence of nitrogen and biotin interactions on the performance of Saccharomyces in alcoholic fermentations. J. Appl. Microbiol. 2007, 102, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Zarogiannis, S.; Liakopoulos, V.; Hatzoglou, C.; Vogiatzidis, K.; Salmas, M.; Stefanidis, I.; Gourgoulianis, K.; Molyvdas, P.A.; Lafis, S. Development of an egg-white bioassay for monitoring biotin levels in urine and serum. Anal. Sci. 2007, 23, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Hagen, K.M.; Keller, M.; Edwards, C.G. Survey of biotin, pantothenic acid, and assimilable nitrogen in winegrapes from the Pacific Northwest. Am. J. Enol. Vitic. 2008, 59, 432–436. [Google Scholar]
- Haider, K.; Frederick, L.R.; Flaig, W. Reactions between amino acid compounds and phenols during oxidation. Plant Soil. 1965, 22, 49–64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Omitted Vitamin | Strain | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E1 | I1a | |||||||||||
| 0% Ethanol | 10% Ethanol | 0% Ethanol | 10% Ethanol | |||||||||
| S1 * | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 | |
| None (Control) | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| p-Aminobenzoic acid | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Biotin | ++ d | + | - | ++ d | ++ d | + | ++ d | + | − | ++ d | ++ d | + |
| Folic acid | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| myo-Inositol | ++ | ++ | ++ | ++ d | ++ d | ++ d | ++ | ++ | ++ | ++ | ++ d | ++ d |
| Nicotinic acid | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Pantothenic acid | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Pyridoxine | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Riboflavin | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Thiamin | ++ | ++ | ++ | ++ d | ++ d | ++ d | ++ | ++ | ++ | ++ d | ++ d | ++ d |
| Ethanol (% v/v) | Omitted Vitamin | Strain | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1b | N2 | 2049 | 2091 | ||||||||||
| S1 * | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 | S1 | S2 | S3 | ||
| 0% | None (Control) | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Biotin | ++ | ++ d | + | ++ d | ++ d | + | ++ d | ++ d | + | ++ d | ++ d | + | |
| myo-Inositol | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| Thiamin | ++ | + | - | ++ d | ++ | ++ | ++ d | ++ | ++ | ++ | ++ | ++ | |
| 10% | None (Control) | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ |
| Biotin | ++ | ++ d | + | ++ d | ++ d | + | ++ | ++ d | + | ++ | ++ d | - | |
| myo-Inositol | ++ d | ++ d | ++ d | ++ | ++ d | ++ d | ++ | ++ | ++ | ++ | ++ | ++ | |
| Thiamin | ++ d | ++ d | ++ d | ++ d | ++ d | ++ d | ++ | ++ d | ++ d | ++ | + | − | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Von Cosmos, N.H.; Edwards, C.G. Use of Nutritional Requirements for Brettanomyces bruxellensis to Limit Infections in Wine. Fermentation 2016, 2, 17. https://doi.org/10.3390/fermentation2030017
Von Cosmos NH, Edwards CG. Use of Nutritional Requirements for Brettanomyces bruxellensis to Limit Infections in Wine. Fermentation. 2016; 2(3):17. https://doi.org/10.3390/fermentation2030017
Chicago/Turabian StyleVon Cosmos, Nicolas H., and Charles G. Edwards. 2016. "Use of Nutritional Requirements for Brettanomyces bruxellensis to Limit Infections in Wine" Fermentation 2, no. 3: 17. https://doi.org/10.3390/fermentation2030017
APA StyleVon Cosmos, N. H., & Edwards, C. G. (2016). Use of Nutritional Requirements for Brettanomyces bruxellensis to Limit Infections in Wine. Fermentation, 2(3), 17. https://doi.org/10.3390/fermentation2030017