
An Update on Microbial Biosynthesis of β-Caryophyllene, a Sesquiterpene with Multi-Pharmacological Properties


1
Institute of Chemical Engineering, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria
2
ICAR—Central Institute of Research on Buffaloes, Hisar 125001, Haryana, India
*
Author to whom correspondence should be addressed.
Fermentation 2024, 10(1), 60; https://doi.org/10.3390/fermentation10010060
Submission received: 9 December 2023
/
Revised: 3 January 2024
/
Accepted: 12 January 2024
/
Published: 15 January 2024
(This article belongs to the Special Issue New Research on Strains Improvement and Microbial Biosynthesis)
Abstract
:The sesquiterpene β-caryophyllene (BCP) is a major component of various plant essential oils, to which it confers a unique spicy aroma. It is mainly used as a fragrance additive in the food, cosmetic and perfume industries, with an annual consumption ranging between 100 and 1000 metric tons worldwide. Recently, BCP has attracted attention as a promising precursor for the production of high-density fuels and for its various biological activities and pharmacological effects. These include antioxidant, anti-inflammatory, anticancer, immune–modulatory, and many other activities. Due to its underlying mechanisms, β-caryophyllene interacts with various human receptors, including CB2 of the endocannabinoid system, which defines it as a phytocannabinoid with therapeutic potential for certain serious conditions. Due to β-caryophyllene’s high utility, various green and sustainable strategies for its production in microorganisms have been developed. This article provides an update on the state-of-the-art in this field to identify directions for further development to extend the compound’s potential.
1. Introduction
The naturally occurring sesquiterpene (E)-β-caryophyllene (BCP) has a distinctive spicy and woody scent and is “generally recognized as safe” [1]. As a result, it is frequently utilized as a flavoring and fragrance agent in the food and cosmetics industries [2,3,4]. Moreover, this terpene is an increasingly common functional ingredient in products, with therapeutic benefits for conditions of pain, neurological diseases and metabolic problems [5,6,7]. These activities are principally related to BCP’s cannabinoid nature, which manifests in the selective activation of cannabinoid type 2 (CB2) receptors [2,7,8]. Caryophyllene is also a promising precursor for the production of advanced biofuels, as well as a potent antibacterial and insecticidal agent [9,10,11].
Global BCP consumption ranges from 100 to 1000 metric tons annually, with a prognostic market value of USD 390 million for 2033 [4,12]. Commercially available β-caryophyllene originates mostly from plants and is offered at a variety of prices and purity levels. It is primarily derived from hops (Humulus lupulus L.), black pepper (Piper nigrum L.), and clove (Syzygium aromaticum L.) [3,5]. Two other promising sources are the diesel tree (Copaifera langsdorffii Desf.) and the Siamese weed (Chromolaena odorata L.), both of which are utilized to produce bioenergy [5]. Even with advances in extraction technology, there are still a number of issues that make producing BCP on an industrial scale challenging. These include dependence on the genetic diversity and phenotypic flexibility of plants, as well as a limited supply of plant material [3,5,9]. Chemical synthesis, in contrast, may be used to manufacture BCP in high quantities and with a comparatively stable quality, but it usually entails a multistep process that is expensive, time-consuming, labor-intensive, and not environmentally benign. This is clearly demonstrated in the first synthetic techniques proposed by Cory et al. [13]. Synthetic biology, as an alternative, can be employed to produce value-added sesquiterpenoids in a sustainable manner [9,14,15,16,17,18]. Microorganisms are ideal hosts for this purpose since they grow quickly, require little workspace and water resources, and can generate all isoprenoid precursors in a natural way [18]. So far, the microbial production of BCP has been primarily studied in Saccharomyces cerevisiae and Escherichia coli using recently developed genetic manipulation strategies at the metabolic and transcriptional levels [9,14,15,16,17,18]. Both hosts have provided promising results, with E. coli CAR2 producing the highest titer of 5.1 g/L [19]. Regardless of this recent progress and keeping in mind that the microbial synthesis of BCP is still in its infancy, extensive research is required to meet industrial demand for this versatile chemical. To that end, it is critical to outline the field’s future prospects.
Reviewing the accomplishments and weaknesses in regulating the intricate mevalonic acid (MVA) and 2-C-methyl-D-erythritol 4-phosphate (MEP) biochemical pathways involved in the microbial synthesis of BCP might shed light on its prospects for building efficient cell factories. We began by investigating these mechanisms in plants because they naturally produce BCP. Then, we focused on metabolic engineering strategies, which are the most widely applied methods for producing BCP in S. cerevisiae and E. coli. In addition, we discovered three other potentially useful microbial hosts: the photosynthetic cyanobacteria Rhodobacter capsulatus [20], Synechocystis sp. [21], and Synechococcus elongatus [22].
2. Biosynthesis of BCP in Plants
BCP is a phytoalexin-like molecule that plays a significant role in plant defense against herbivores and pathogens as well as allelopathic interactions [23,24,25]. This sesquiterpene (C15H25), like all other isoprenoids, is generated by higher plants from isopentenyl pyrophosphate (IPP).
2.1. Biosynthesis of IPP (Upstream Module)
The well-documented biosynthesis of IPP evolves in two independent pathways: in the cytosol via mevalonic acid (MVA) and in the plastids via 2-C-methyl-D-erythritol 4-phosphate (MEP) [24,25,26,27,28,29,30,31]. The initial substrates of each route are transformed into IPP via a series of enzyme-catalyzed reactions (Figure 1).
The MVA pathway begins with the condensation of two molecules acetyl-CoA-catalyzed by acetyl-CoA acetyltransferase (AACT) [32]. The resulting acetoacetyl-CoA interacts with another molecule of acetyl-CoA, which is carried out by hydroxymethylglutaryl-CoA synthase (HMGS) to give 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA), which is then converted to mevalonic acid (MVA). The formation of mevalonic acid is the rate-limiting step of this biosynthetic pathway, and it is catalyzed by 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR). Consequently, HMGR, as a key enzyme, is a frequent subject of modifications intended to enhance the MVA pathway leading to isoprenoid synthesis [26,27,29,32]. Mevalonic acid subsequently undergoes two-step phosphorylation followed by decarboxylation until the sequential formation of mevalonic acid 5-phosphate (MVAP), 5-diphosphomevalonic acid (MVAPP), and finally IPP. The reactions are mediated by mevalonate kinase (MK), phosphomevalonate kinase (PMK), and mevalonate diphosphate decarboxylase (MDD), respectively. Part of IPP is then isomerized by isopentenyl diphosphate isomerase (IDI) to create dimethylallyl diphosphate (DMAPP). In addition, an alternative route to IPP has been proposed, which includes the decarboxylation of MVAP to isopentenyl phosphate (IP) by phosphomevalonate decarboxylase (PMD), followed by the phosphorylation of IP to IPP by isopentenyl phosphate kinase (IPK) [29,31].
The MEP pathway is initiated by two rate-limiting steps: the condensation of pyruvate and glyceraldehyde 3-phosphate (G3P) to 1-deoxy-D-xylulose 5-phosphate (DXP), catalyzed by 1-deoxy-D-xylulose-5-phosphate synthase (DXS), and the reduction of DXP to 2-C-methyl-D-erythritol 4-phosphate (MEP) by 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR) [32]. MEP is converted to 4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol (CD-ME) by 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase (MCT). The newly formed molecule is then phosphorylated to 4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol 2-phosphate (CD-ME2P) by 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase (CMK). The cyclization of CD-ME2P by 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MDS) leads to 2-C-methyl-D-erythritol 2,4-cyclodiphosphate (ME-cPP), which is subsequently converted to 1-hydroxy-2-methyl-2-butenyl 4-diphosphate (HMBPP) utilizing 4-hydroxy-3-methylbut-2-enyldiphosphate synthase (HDS). In the final step, HMBPP uses 4-hydroxy-3-methylbut-2-enyl diphosphate reductase (HDR) to produce a mixture of IPP and DMAPP.
2.2. BCP Synthesis (Downstream Module)
The two isomers IPP and DMAPP are the building blocks of all terpenoids and can be fused together by prenyltransferases to produce linear precursors of variable lengths [24,25,31]. They first undergo a head-to-tail condensation catalyzed by geranyl diphosphate synthase (GPPS) to produce geranyl diphosphate (GPP), the monoterpenoid precursor (Figure 1). Meanwhile, GPP interacts with another molecule of IPP under the catalytic action of farnesyl diphosphate synthase (FPPS) to form farnesyl pyrophosphate (FPP). FPP gives rise to all sesquiterpenoids by undergoing a number of cyclization, ionization, and rearrangement reactions mediated by a specialized class of enzymes called sesquiterpene synthases [33]. To yield BCP, (E,E)-FPP forms (E,E)-α-humulyl carbocation, a portion of which rearranges into α-humulene, a ring-open isomer of BCP, while the remainder cyclizes to a caryophyllene cation [34,35]. This cation rearranges into (E)-β-caryophyllene, which can further isomerize to its Z-β-form, isocaryophyllene. In fact, BCP is typically found in plants in mixtures with trace levels of its isomers, α-humulene and isocaryophyllene, which serve identical ecological functions [24,34,36].
To date, over 300 plant sesquiterpene synthases have been characterized and stored on an online database [33]. Some of these enzymes, which catalyze the biosynthesis of BCP, have been employed in the genetic engineering of models and ornamental plants as well as crops, such as Artemisia annua L. [37], Arabidopsis thaliana L. Heynh. [38], Nicotiana benthamiana Domin [23], Phoebe bournei (Hemsl.) Yang [39], Dendrobium officinale Lindl. [40], Oenanthe javanica (Blume) DC. [41], Zea mays L. [42], Oryza sativa L. [43], Gossypium hirsutum L. [44], Daucus carota L. [45], etc. In general, the regulation of gene expression promoting BCP production has been found to be crucial for the plant response to pathogens and herbivores, thus offering a sustainable pest management strategy [42,44,46,47,48].
3. Microbial Biosynthesis of BCP
Metabolic engineering strategies for the synthesis of BCP have been primarily developed in two model microorganisms, the yeast S. cerevisiae and the Gram-negative bacterium E. coli [14,15,17]. Both are well-characterized, robust hosts for the efficient and large-scale production of industrially important chemicals [32,51,52]. The photosynthetic bacteria R. capsulatus [20], Synechocystis sp. [21], and S. elongatus [22] were also tested, with different degrees of success. Unlike plants, which produce IPP via two biosynthetic routes, yeast solely uses the MVA pathway, whereas most Gram-negative bacteria employ the MEP pathway [26,27,32].
3.1. S. cerevisiae
The most extensively utilized strategy for producing sesquiterpenes in yeast is the regulation of the enzyme balance across the entire MVA pathway [53,54]. The main goal of modifying the upstream module is to bypass the rate-limiting step of mevalonic acid synthesis, allowing for efficient IPP and DMAPP production [14,15,29]. The supply of these isomers must be regulated to avoid excessive levels that, due to their toxicity, can impair cell growth [14,15,29]. The downstream module is manipulated in order to boost the flow to FPP, the sesquiterpenoid precursor, and to use appropriate terpene synthases to generate a high titer of the desired products [29].
The rate-limiting reaction of the MVA pathway in S. cerevisiae is mediated by two isozymes, encoded by HMGR1 and HMGR2 [14,29,51]. To facilitate the FPP synthesis, one or both of these genes is typically combined with other gene elements, such as IDI1 and ERG20, which encode the IDI and FPS enzymes, and selected promoters in a recyclable integration cassette [49]. This ensures their unlimited sequential integration at any locus in the yeast genome [49,55]. To produce BCP, Ignea et al. inserted a cassette carrying HMGR2, IDI1, and ERG20 into S. cerevisiae EG60, followed by plasmid-driven co-overexpression of the HMG2 (K6R) variant, IDI1, and the S. pomifera L. terpene synthase SpP330 [49]. As a result, they were able to obtain a sesquiterpene mixture containing 11.25% BCP. To better overcome the rate-limiting step, the same scientists engineered a S. cerevisiae strain with the HMGR2 (K6R)-encoded enzyme additionally stabilized via tandem heterozygous deletion of three genes involved in the degradation of the endoplasmic reticulum transmembrane protein, Ubc7p, Ssm4p, and Pho86p [50]. After overexpression of S. fruticosa L. terpene synthase Sf126, the final strain AM109 produced 125 mg/L BCP in a shake flask. Thus, the yield of the desired product increased 11-fold above the wild strain. Traces of the BCP isomer, α-humulene, were also detected.
Li et al. boosted the endogenous MVA pathway in S. cerevisiae BY4742 by knocking down the transcriptional regulator GAL80 [56]. This modification, which is known to activate the GAL1/GAL10 bidirectional promoter in the absence of galactose [57], enabled the integration of two expression cassettes. The first was constructed from ERG10 (encoding AACT), ERG13 (encoding HMGS), tHMG1, ERG12 (encoding MK), and ERG8 (encoding PMK) and inserted at delta loci. The second cassette, which included the genes tHMG1, ERG19 (encoding MDD), IDI1, and ERG20, was integrated at the rDNA site. The derived sesquiterpene chassis strain SQTBY03 boosted the availability of FPP by 458-fold. The yeast cell factories of CPLBY01 were then built by overexpressing codon-optimized QHS1 from A. annua into the SQTBY03 strain for de novo synthesis of BCP at a titer of 250.4 mg/L in a shake flask. After 140 h of fed-batch fermentation with CPLBY01, a mixture of 2949.1 mg/L BCP and 109.8 mg/L α-humulene from glucose was obtained [56]. Meanwhile, truncated HMG1 genes, which produce an enzyme lacking the membrane-binding region, are frequently overexpressed in S. cerevisiae to increase the isoprenoid titer [14,29].
In their quest to optimize the MVA pathway, Lu et al. discovered that Enterococcus faecalis EfHMGR expression was greater than that of S. cerevisiae tHMGR1 [58]. Based on this finding, they developed a sophisticated multistep strategy for driving the flux in the direction of squalene and BCP synthesis. To that end, S. cerevisiae SQ3-4, which shows high sterol pathway activity, was engineered to carry overexpressed genes related to enhanced MVA pathways (ERG10, ERG13, EfHMGR, ERG12, ERG8, ERG19, IDI1, ERG20, and ERG9); reduced nicotinamide adenine dinucleotide phosphate (NADPH) regeneration (gapC); increased acetyl-CoA supply (CDC19 + PDC1); and improved alanine metabolism (GAD1, PAN5, and CAB1). A squalene-producing strain, SQ3-5, was obtained by down-regulating ERG1 in S. cerevisiae SQ3-4. Overexpression of the A. annua QHS1 gene in S. cerevisiae SQ3-4 resulted in the strain SQ3-4-CPS, which produced 11.86 ± 0.09 mg/L BCP in a shake flask. In a subsequent study, Lu et al. employed directed evolution to optimize A. annua β-caryophyllene synthase [59]. One of the resulting variants, the E353D mutant, exhibited 35% higher catalytic activity than the wild type and operated in significantly wider pH and temperature ranges. The novel S. cerevisiae chassis, showing higher metabolic flux of FPP synthesis and a higher capacity for transmembrane transport of β-caryophyllene, was constructed by overexpressing β-alanine-metabolism-related genes GAD1, PAN6, and CAB1; MVA-pathway-related genes EfHMGR, ERG12, ERG8, CUP1 (promoter), ERG19, IDI1, and ERG20; and the ATP-binding cassette transporter gene variant STE6T1025N. After 48 h of cultivation in a test tube, the final strain, YEH4-ECPSE353D, produced 70.45 mg/L BCP, 2.93 times more than the strain SQ3-4-CPS [58,59]. Ultimately, a 96 h fed-batch fermentation process yielded a BCP yield of 594.05 mg/L at a synthesis rate of 6.19 mg/L/h [59]. The conversion rate of glucose was 3.54 mg/g.
Godara et al. successfully combined metabolic engineering with adaptive laboratory evolution (ALE) to increase BCP production in S. cerevisiae BY4741 [60]. ALE is an experimental technique commonly used to adapt industrial strains to produce specialized metabolites under selective environmental conditions [53,61,62]. Following this approach, the cell growth of BY4741 was stimulated in an oxidative environment by exploiting the antioxidant capacity of BCP; i.e., the strain produced more antioxidants for survival due to induced gene mutations [60]. Godara et al. first deleted the cytosol catalase gene (CTT1) to reduce yeast resistance to hydrogen peroxide. Then, modified variants of the strain, carrying the codon-optimized QHS1 gene from A. annua, tHMG1, HMG2(K6R), UPC2-1, and ERG20, were constantly or intermittently exposed to growth-permissible amounts of hydrogen peroxide. Those that produced larger levels of BCP were more resistant to the induced oxidative stress and performed better in the periodic challenge strategy. Only mutations in the intergenic region of MST27/tR(UCU)G1 and a nonsynonymous mutation in STE6 were found to be beneficial for product formation. The mechanism of their contribution remains unclear, although a mutation in STE6, a known ABC transporter, may be involved in BCP export. This combined strategy increased BCP synthesis in isolated mutants four-fold, and their cell growth correlated positively with yields. An additional increase in flux to BCP in the most developed mutant, YAG117, produced a titer of 104.7 ± 6.2 mg/L in a test tube.
Another strategy to overcome the intricate regulation of the MVA pathway towards terpenoid production is to rewire the yeast central carbon metabolism [15,18,29]. In this context, the implementation of a synthetic malonic acid-acetoacetyl-CoA (MAAC) metabolic route, which increases the supply of acetyl-CoA, was successful in producing BCP in S. cerevisiae YQ-7 in a stable and effective manner [63]. Liang et al. obtained this strain from S. cerevisiae CEN.PK2-1D by sequential integrating the A. annua QHS1 gene, tHMG1, IDI1, ERG20, ERG13, and ERG10, and finally a cassette for constitutive and strong expression of the malonate transporter Mae I (from Schizosaccharomyces pombe), MatB (encoding malonyl-CoA synthetase), and ACCS (encoding acetoacetyl-CoA synthase). The fermentation conditions were optimized in a shake flask by adding Mg2+, which improved the catalytic activity of QHS1. The highest BCP titer of 328 ± 13.44 mg/L was obtained with six rounds of glucose supplementation after 144 h of fermentation. This yield was 13-fold higher than that of the control strain.
For clarification’s sake, the most successful strategies for producing BCP in S. cerevisiae are depicted in Figure 2. The carbon source is always glucose, except for the strain YQ-7, which used malonic acid [64]. Precise comparison of the efficacy of the applied approaches is difficult due to the different experimental settings employed and lack of uniformity for data presentation. However, rewiring of the yeast central carbon metabolism seems to surpass the other established strategies and definitely needs further attention. Importantly, the majority of research in this area has been conducted in the last three years, and the results provide a highly promising foundation for obtaining economically relevant S. cerevisiae strains for BCP synthesis.
3.2. E. coli
Engineering E. coli strains for BCP synthesis often involves optimizing the native MEP pathway or designing a hybrid MVA metabolic route [14,15,16,64]. The preferred approach commences with the identification and improvement in exogenous terpene synthases, given that this bacterium lacks the ability to produce them internally.
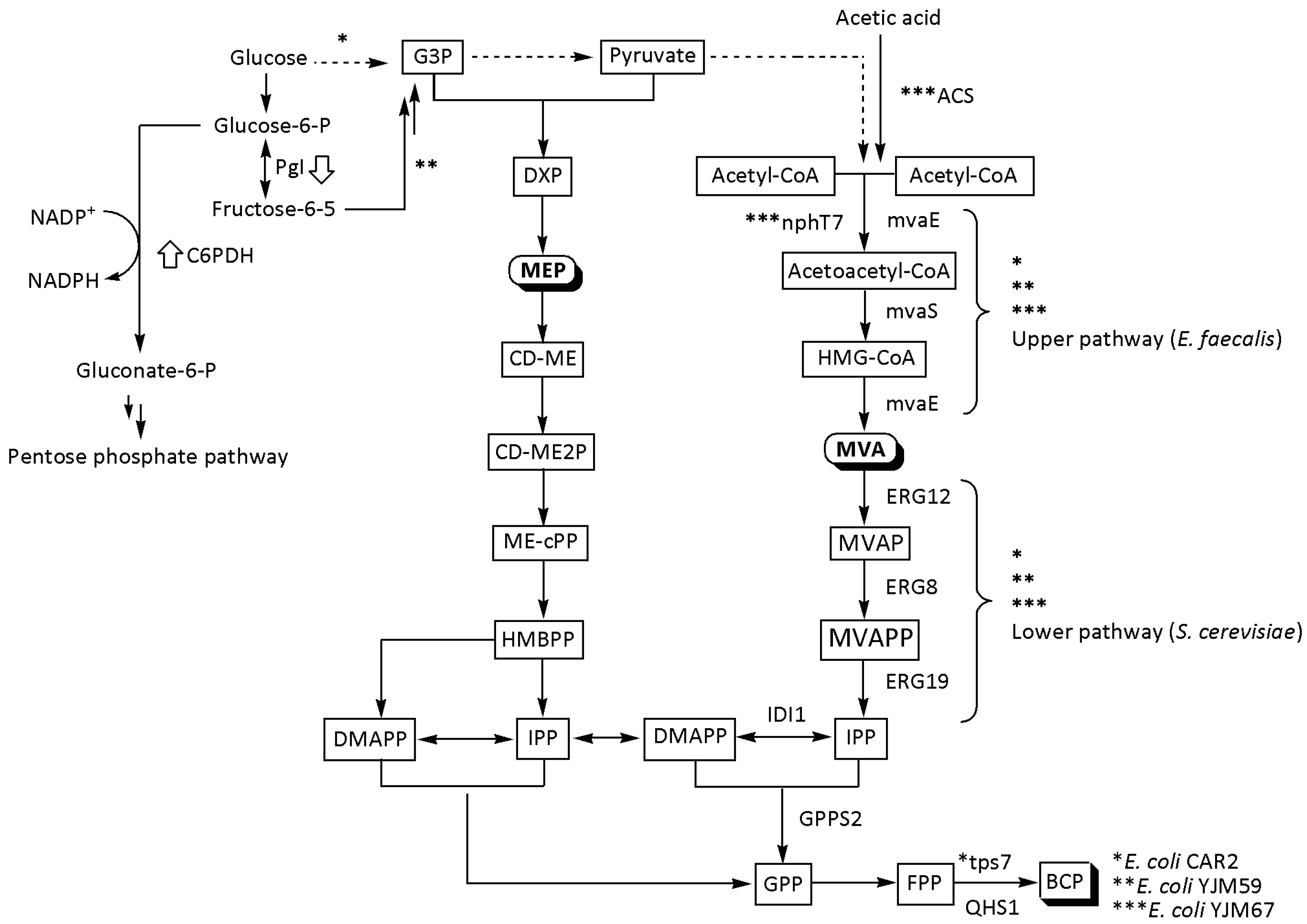
Yang et al. tested the efficiency of three codon-optimized terpene synthase genes for BCP synthesis in E. coli BL21 (DE3) [9]. In this regard, they demonstrated that QHS1 from A. annua was the best choice compared to TPS21 from A. thaliana and TPS23 from Z. perennis. After integrating QHS1, Yang et al. modified the strain’s MEP metabolism by overexpressing the ispG gene (encoding HDS). The ispG gene is known to effectively reduce ME-cPP efflux and therefore increase downstream terpenoid synthesis [14,15]. This procedure raised the BCP yield 2-fold over the control strain with a native MEP pathway. Further overexpression of the dxs gene, which encodes the rate-limiting DXS enzyme, resulted in an additional 1.67-fold increase. Under shake flask conditions, the resulting strain YJM54 produced BCP with a titer of 350 ± 18 µg/L. The parent strain was also engineered to harbor β-caryophyllene synthase, hybrid MVA, and native MEP pathways. In the resulting YJM55 strain, the hybrid MVA cluster was created by combining the upper E. faecalis pathway and the lower S. cerevisiae pathway. This approach led to a dramatic 142.9-fold increase in the BCP production to 15 ± 0.75 mg/L when compared to the control strain. The yield was subsequently raised to 52 ± 2.2 mg/L and then to 220 ± 6 mg/L in the final YJM59 strain by integrating GPPS2 gene from Abies grandis (Douglas ex D. Don) Lindley, encoding GPPS, and deletion of the phosphoglucose isomerase gene (pgi), respectively. The final two steps were designed to increase the supply of GPPS and NADPH. After 60 h, the fed-batch fermentation with E. coli YJM59 yielded 1.52 g/L BCP from glucose [9]. The conversion efficiency of glucose to BCP (g/g) was 1.69%. The BCP yield was about 6.7% of the theoretical yield (25.2%), indicating that this modified E. coli strain has a high potential for large-scale synthesis of (E)-β-caryophyllene [9].
Cheng et al. discovered a new gene of β-caryophyllene synthase (TPS7) by analyzing the genome of N. tabacum L. using bioinformatics methods [19]. By overexpressing the tps7 gene in E. coli BL21 (DE3), they were able to build a recombinant CAR1 strain carrying an additional MVA gene cluster from E. faecalis. This strategy resulted in the synthesis of 16.1 mg/L BCP in a shake flask after 24 h of cultivation. However, during the fermentation process, cell growth was impaired by the accumulation of large amounts of BCP in the intracellular space. Only 20% of the product was released in the culture broth, while its extracellular concentration gradually decreased during the course of the reaction due to the volatility of this terpene. On the other hand, overexpression of the MVA pathway resulted in an abundance of FPP in the fermentation broth, where farnesyl acetate, probably derived from FPP, was also detected in high amounts as a byproduct. This compound is also toxic to cells and inhibits DNA replication [19,29]. To solve the problem of reduced cell growth and loss of BCP, the downstream genes of the S. cerevisiae MVA pathway were inserted into the E. coli CAR1 genome. The newly obtained strain CAR2 produced 100.3 mg/L BCP in a shake flask after 24 h of fermentation. This manipulation increased CAR2 biomass 1.5-fold relative to the CAR1 strain in the face of dramatically reduced farnesyl acetate. During fed-batch fermentation, the rate of BCP synthesis increased after 4 h and reached its peak yield of 4319 mg/L 60 h after induction. By including n-dodecane in the fermentation broth for in situ BCP extraction, a 20% increase in the production was achieved while limiting caryophyllene loss due to volatilization. The maximum production of BCP was 5142 mg/L from glucose with a productivity of 80.3 mg/L/h at 64 h after induction [19]. In the high-cell-density fermentation, the efficiency of the metabolically modified strain in converting glucose to BCP (g/g) was 4.4%.
Yang et al. took up the challenge to produce BCP in E. coli utilizing acetic acid as the sole carbon source [65]. Such a synthesis requires extensive metabolic engineering due to the fact that acetic acid has an inhibitory effect on microorganisms, reducing their growth rates and thus limiting product concentrations [66,67]. Aiming to create a robust strain, Yang et al. first modified E. coli BL21(DE3) by co-overexpressing an optimized QHS1 from A. annua and acetyl-CoA synthase (ACS) from Acetobacter pasteurianus [65]. However, this approach using the native MEP pathway of E. coli proved insufficient to produce the desired chemical. The strain was further improved by introducing the GPPS2 gene from A. grandis to increase GPP supply, resulting in 102 ± 9 μg/L of BCP after 24 h of cultivation. Based on their previous experience [9], the authors constructed a recombinant strain YJM66 containing a hybrid MVA pathway to direct the flux to the FPP. The strain YJM66 was obtained by co-overexpression of three plasmids carrying QHS1 from A. annua, mvaE, and mvaS from E. faecalis and the GPPS2 gene from A. grandis; ERG12, ERG8, ERG19, and IDI1 from S. cerevisiae; and ACS from A. pasteurianus in E. coli BL21(DE3). This procedure resulted in an 8-fold rise in the BCP yield to 8 ± 0.75 mg/L. The strain was further optimized by overexpressing an AACS-encoding gene (nphT7) from a Streptomyces sp. strain CL190 to increase the acetoacetyl-CoA supply in the cells. As a result, the BCP yield increased to 22 ± 1.8 mg/L after 24 h of cultivation in 60 mM acetate medium. The final strain, YJM67, produced 1.05 g/L BCP in a fed batch after 72 h of fermentation [65]. The conversion efficiency of acetic acid (g/g) to BCP was 2.1%.
Another alternative carbon source, the hydrolysate of the marine microalgae Nannochloropsis sp., has recently been used for the manufacture of value-added terpenoids as an advanced biofuel in E. coli mutant strains [68,69]. Although this method is designed to produce terpenoid combinations, they are enriched in BCP (20%), and proper tuning of the E. coli genome and fermentation conditions may represent an alternative avenue for BCP production. Furthermore, microalgae constitute a promising renewable resource for the sustainable manufacture of bioproducts [70].
The most successful tactic for producing BCP in E. coli (Figure 3) was developed and patented by a group of researchers who worked hard to optimize the strains [9,19,65,71,72]. The results obtained are very promising and seem to be one step closer to the synthesis of BCP on a multigram scale. Additionally, the team has outlined several goals for future research, including, but not limited to, improving the catalytic activity of TPS7, fine-tuning the expression levels of several inter- and intra-modules of the metabolic pathway, and optimizing the fermentation step in real time with online monitoring [19]. It is worth noting here that a new efficient chromatographic method for the purification of β-caryophyllene from E. coli supernatant has recently been developed [73]. It uses the polymeric material Rensa® RP from Biotage, Uppsala, Sweden as an adsorbent and the green solvent ethanol as an eluent. In one step, this system provides BCP with >99% purity and >80% yield. This method is considered superior to liquid–liquid extraction for microbially produced terpenes.
3.3. R. capsulatus
The photosynthetic species Rhodobacter is a promising alternative host for BCP production. Individuals are able to accumulate FPP through their native MEP biosynthetic pathway and also contain an intrinsic carotenoid biosynthetic pathway that can be engineered for sesquiterpene synthesis [20,21]. They also form an extended intracytoplasmic membrane system, thus providing a naturally extended reservoir for membrane-bound enzymes and terpenes [21]. Another advantage of Rhodobacter species is their ability for phototropic growth in a low-cost minimal medium at relatively high growth rates, allowing for the use of sunlight as an energy source for sustainable cultivation and production processes [21,74]. Recent studies have shown that the engineered biosynthesis of the isoprenoid precursor can lead to a strong increase in the formation of sesquiterpenoids in R. capsulatus [74] and R. sphaeroides [75].
Hilgers et al. performed several steps to synthesize β-caryophyllene in R. capsulatus [20]. First, they inserted a sequence of the A. annua QHS1 gene into the pRhon5Hi-2 vector carrying the strong nifH gene promoter for heterologous gene expression. The constructed the vectors pRhon5Hi-2-QHS1 and pRhon5Hi-2-QHS1-ispA; the latter, containing a copy of the ispA gene, was introduced into the wild-type SB1003 strain as well as the SB1003-MVA strain, which carried the chromosomally integrated MVA pathway genes derived from the zeaxanthin-producing bacterium Paracoccus zeaxanthinifaciens. The strains were incubated under anaerobic phototrophic conditions. In strain SB1003, QHS1 expression produced detectable amounts of BCP. Surprisingly, co-expression of QHS1 and ispA in the same strain did not result in an increase in BCP production. On the other hand, co-expression of QHS1 and ispA in R. capsulatus SB1003-MVA resulted in a 300% increase in BCP synthesis compared to the control strain. The highest BCP titer, 90 ± 19 mg/L, was obtained after three days of cultivation in hangate tubes under flask illumination of the SB1003-MVA strain expressing pRhon5Hi-2-QHS1-ispA. By using infrared light and screw-neck vials for growth, the yield improved to 139 ± 31 mg/L, with a volumetric productivity of 1.93 ± 0.44 mg/L/h. This research, which is a result of engineering the isoprenoid precursor supply as well as improving growth conditions, successfully established R. capsulatus as a heterologous host for BCP synthesis.
It is completely logical to expect a similar study in R. sphaeroides, which has already been used for the synthesis of another sesquiterpene, amorphadiene [75]. But for now, this possibility remains a niche for future research.
3.4. Synechocystis sp.
Cyanobacteria are extremely adaptable, fast-growing photosynthetic prokaryotes that are able to transform carbon dioxide into sugar and water and are well suited for the manufacture of terpenoids [76]. They produce a lot of carotenoids and chlorophyll on their own and can be engineered to synthesize sesquiterpenoids [76,77].
The cyanobacteria from Synechocystis sp. were initially described as a promising platform for the synthesis of sesquiterpenoids in 2011 when BCP was generated by a PCC6803 strain containing the A. annua QHS1 gene in its genome [21]. In the pertinent work, Reinsvold et al. designed cloning primers to amplify the QHS1 1644 bp open reading frame and place it next to the strong psbAII promoter at the location of the multiple cloning of the psbAII locus in PSBAII-KS plasmid. In a PCC6803 strain transformed by plasmid PSBAII-KS, the resultant pPSBAIIQHS1 vector was inserted using a homologous recombination procedure in the presence of sucrose. The stabilized QSH1 gene enhanced the bacterium’s natural MEP biosynthesis pathway, resulting in a BCP growth rate of approximately 46.4 ± 2.9 ng/mL/week under standard growth conditions.
Synechocystis has been abandoned as an alternative for the production of BCP despite the recent usage of the PCC6803 strain in the successful production of other sesquiterpenes, such as (E)-α-bisabolene, valencene, and santalene [78,79,80]. For their synthesis, some of the native MEP pathway bottlenecks in cyanobacteria were overcome by introducing heterologous enzymes and/or deleting selected genes, but cultivation was also examined under photoautotrophic conditions and on alternate sources of nitrogen. Undoubtedly, the positive outcome of these strategies can be applied to improve the production of BCP in Synechocystis sp.
3.5. S. elongatus
Synechococcus is a unicellular cyanobacterium that is widespread in the marine environment. Recently, some two physiologically diverse model strains have been demonstrated to synthesize the sesquiterpenes bisabolene and farnesene [81].
A group of Chinese scientists succeeded in genetically modifying S. elongatus UTEX 2973 to obtain a BCP-producing strain [82]. They constructed three vectors, pSI-ispA-gpps-idi1sc, pSII-idi1sc, and pSIII-tps21, and transferred them in the parent strain to obtain Synechococcus NSI-gpps-ispA-idi1sc-II-idi1sc-III-tps21. The final strain utilizing the catalytic activity of TPS21 from A. thaliana produced 212.37 µg/L BCP in a photobioreactor for 96 h; the output per unit time was increased 8 times. A patent claim is based on this innovation.
4. A Summary of Findings
The growing industrial demand for BCP as a result of its numerous applications in various sectors naturally drew the attention of synthetic biology. Despite the fact that such a “green” approach to manufacturing this added-value sesquiterpene is still in its early stages, significant effort has been put forward in recent years to establish an efficient technique for synthesizing BCP in the two most well-known microbial hosts, S. cerevisiae and E. coli. The best outcomes were obtained by resolving bottlenecks in the relevant metabolic pathways by balancing enzyme expression and deleting or down-regulating competing processes utilizing modern technology. Table 1 shows the most efficient strains in terms of BCP titer obtained in a shake flask or test tube, in ascending order from top to bottom, along with the time of fermentation, if reported. Fed-batch fermentation was also carried out with some of the S. cerevisiae strains and all of the E. coli strains. The table also includes the results of single investigations with three different microbial hosts, R. capsulatus, Synechocystis sp., and S. elongatus. As a result, the summarized data are an amalgam of several BCP microbiological synthesis strategies.
The development of efficient strains of S. cerevisiae was most effective when the full homologous MVA route was enhanced. Typically, this was accomplished by inserting expression cassettes encoding the necessary enzymes (Table 1, Figure 2). Improving the rate-limiting enzyme is usually achieved by using a truncated form of HMGR1 (CPLBY01 and YQ-7 strains) [56,67], the stable variant HMGR2 (K6R) (AM109 strain) [50], or a combination of tHMGR1 and HMGR2 (K6R) (P11M1 strain) [60], or by substituting the S. cerevisiae tHMGR1 with the more effective E. faecalis EfHMGR (YEH4-ECPSE353D strain) [59]. Another important goal is to optimize and maintain the gene that encodes the intended terpene synthase. This is most typically QSH1 from A. annua, which can be improved at the translational level through methods such as codon optimization, directed evolution, and insertion into recyclable expression cassettes. However, Sf126 from S. fruticosa (AM109 strain) was also successfully applied [50]. Furthermore, better transmembrane transport of BCP was accomplished by either incorporating the ATP-binding cassette transporter gene variation STE6T1025N (YEH4-ECPSE353D strain) [59] or mutation in STE6, a recognized ABC transporter (P11M1 strain) [60]. Two intriguing approaches to improving BCP production are the ALE technique, which leverages BCP’s natural ability to scavenge peroxy radicals (P11M1 strain) [60] and rewiring the central carbon metabolism to use malonic acid as a carbon source, thereby increasing acetyl-CoA supply (YQ-7 strain) [63].
Engineering E. coli for efficient BCP production requires two critical steps: the selection of an appropriate terpene synthase and the integration of a hybrid MVA pathway in the bacterial genome (Table 1, Figure 3). As E. coli lacks terpene synthases, genes encoding such enzymes in plants were inserted into the parental strains. The expression of QHS1 from A. annua (YJM67 and YJM59 strains) [9,65] and tps7 from N. tabacum (CAR2 strain) [19] was shown to be the most efficient. Although the native MEP pathway has a larger theoretical output based on carbon stoichiometry, the exogenous MVA pathway produces terpenoids in E. coli more effectively. Furthermore, a hybrid MVA cluster formed by combining the upper E. faecalis route with the lower S. cerevisiae pathway provided a better solution (YJM59 strain) [9]. The expression of the GPPS2 gene from A. grandis, which encodes GPPS, in conjunction with pgi deletion, which increases NADPH supply, enhanced the BCP titer generated by the same strain. Two patents cover the design of vectors and plasmids used in these experiments [71,72]. It was also demonstrated that E. coli can be engineered to synthesize BCP utilizing acetic acid as the only carbon source (YJM67 strain) [65].
Amongst the alternative bacterial hosts (Table 1), R. capsulatus was engineered to produce BCP by combining MVA pathway genes from the carotenoid-producing bacteria P. zeaxanthinifaciens with QHS1 from A. annua and the ispA gene (SB1003-MVA strain) [20]. The yield was raised by cultivation under phototrophic conditions using UV and particularly IR light. Another photosynthetic bacterium, Synechocystis sp., containing the stabilized QHS1 gene from A. annua was able to synthesize BCP on a nanogram scale (PCC6803 strain) [21], but no further attempts to increase the yield were made despite the strain’s success in producing other sesquiterpenoids. Experiments were instead carried out in the Synechocystis phylogenetic neighbor S. elongatus. The modified S. elongatus strain outperformed Synechocystis by exploiting the catalytic activity of A. thaliana TPS21, although it still produced BCP in poor yields [82]. The experiment’s findings are protected by a patent, and it is hoped that the strain will be further refined.
5. Conclusions and Future Perspectives
Despite advances in microbial BCP production in S. cerevisiae and E. coli, this important sesquiterpene has yet to be synthesized on an industrial scale. In this context, numerous tactics and tools will be used to improve the BCP titer. The focus will remain on optimizing core metabolic pathways and building novel hybrid clusters, but more artificial pathways will be explored, and suitable enzymes will be discovered [14,15,17,29,55,83]. Retrosynthesis technologies will be supplemented with high-throughput procedures enabling the automated design of microbial strain engineering to aid in these efforts [54,83,84,85]. Efforts will also be made to find more efficient terpene synthases [86], particularly in plants rich in BCP; to conduct membrane engineering for increased storage capacity [54,83,87,88]; to induce or overexpress efflux pumps, which have broad substrate specificity and export toxic metabolites outside the cells [85,89,90]; and to explore additional opportunities for applying the ALE technique [54,83,91,92]. In situ product extraction will receive more attention [54,83], especially since it has proven effective for improving the BCP titer in E. coli [19]. Most likely, preparative chromatography will be studied more thoroughly as a method for isolating β-caryophyllene with high purity and yield from fermentation broth. An impetus for such research could be the recently established chromatographic method for microbially generated BCP that uses a system of the polymeric substance Rensa® RP from Biotage, Uppsala, Sweden and ethanol and is more efficient than liquid–liquid extraction [73].
Other approaches will aim to modify the energy of and reduce the power balance in microbial cells [54,83]. Hopefully, some new carbon sources will be accepted as well. Among the promising candidates are xylose, glycerol, carbon dioxide, isopentenol, and marine microalgae and sugarcane bagasse hydrolysates [54,83]. Finally, learning from the progress in the microbial synthesis of other sesquiterpenes will allow for improved BCP titers in photosynthetic bacteria [75,78,79,80] as well as the exploration of other microbial hosts, particularly the Gram-negative bacteria Cupriavidus necator and Methylobacterium extorquens, which have been reported to produce the BCP isomer α-humulene in yields of 11 mg/L and 1.65 g/L, respectively [93].
This review paper outlines the accomplishments, difficulties, and prospects in the microbial synthesis of (E)-β-caryophyllene, demonstrating that more work remains to be done before reaching commercial-scale production.
Author Contributions
Conceptualization, D.B. and L.T.; methodology, L.T. and C.S.; software, L.T.; validation, L.T. and C.S.; formal analysis, L.T. and C.S.; investigation, L.T. and C.S.; resources, L.T. and C.S.; data curation, C.S.; writing—original draft preparation, C.S.; writing—review and editing, L.T. and D.B.; visualization, L.T.; supervision, D.B.; project administration, not applicable; funding acquisition, not applicable; L.T. and C.S. contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- CFR—Code of Federal Regulations Title 21. 2023. Volume 3. 21CFR172.515. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=172.515&SearchTerm=caryophyllene (accessed on 20 September 2023).
- Francomano, F.; Caruso, A.; Barbarossa, A.; Fazio, A.; La Torre, C.; Ceramella, J.; Mallamaci, R.; Saturnino, C.; Lacopetta, D.; Sinicropi, M.S. β-Caryophyllene: A sesquiterpene with countless biological properties. Appl. Sci. 2019, 9, 5420. [Google Scholar] [CrossRef]
- Caryophyllene Market Analysis—Industry Size, Share, Research Report, Insights, COVID-19 Impact, Statistics, Trends, Growth and Forecast 2023–2030. Available online: https://markwideresearch.com/caryophyllene-market/ (accessed on 21 September 2023).
- Caryophyllene Market by Purity, Application, End Use and Region. Forecast 2023 to 2033. Available online: https://www.futuremarketinsights.com/reports/caryophyllene-market (accessed on 20 September 2023).
- Maffei, M.E. Plant natural sources of the endocannabinoid (E)-β-caryophyllene: A systematic quantitative analysis of published literature. Int. J. Mol. Sci. 2020, 21, 6540. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Yuan, S.; Li, L.; Zheng, J.; Zhao, D.; Wang, C.; Wang, H.; Liu, X.; Liu, J. Application of terpenoid compounds in food and pharmaceutical products. Fermentation 2023, 9, 119. [Google Scholar] [CrossRef]
- Blake, K. Beta-Caryophyllene: A review of current research. Altern. Complement. Ther. 2021, 27, 222–226. [Google Scholar] [CrossRef]
- Hashiesh, H.M.; Sharma, C.; Goyal, S.N.; Sadek, B.; Jha, N.K.; Kaabi, J.A.; Ojha, S. A focused review on CB2 receptor-selective pharmacological properties and therapeutic potential of β-caryophyllene, a dietary cannabinoid. Biomed. Pharmacother. 2021, 140, 111639. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Z.; Guo, L.; Du, J.; Bae, H.J. Biosynthesis of β-caryophyllene, a novel terpene-based high-density biofuel precursor, using engineered Escherichia coli. Renew. Energy 2016, 99, 216–223. [Google Scholar] [CrossRef]
- Myszka, K.; Schmidt, M.T.; Majcher, M.; Juzwa, W.; Czaczyk, K. β-Caryophyllene-rich pepper essential oils suppress spoilage activity of Pseudomonas fluorescens KM06 in fresh-cut lettuce. LWT Food Sci. Technol. 2017, 83, 118–126. [Google Scholar] [CrossRef]
- Ma, S.; Jia, R.; Guo, M.; Qin, K.; Zhang, L. Insecticidal activity of essential oil from Cephalotaxus sinensis and its main components against various agricultural pests. Ind. Crops Prod. 2020, 150, 112403. [Google Scholar] [CrossRef]
- Api, A.M.; Belsito, D.; Botelho, D.; Bruze, M.; Burton, G.A., Jr.; Buschmann, J.; Cancellieri, M.A.; Dagli, M.L.; Date, M.; Dekant, W.; et al. RIFM fragrance ingredient safety assessment. β-Caryophyllene, CAS Registry Number 87-44-5. Food Chem. Toxicol. 2022, 159 (Suppl. 1), 112707. [Google Scholar] [CrossRef]
- Hancock, E.N.; Wahl, J.M.; Brown, M.K. Recent advances in the synthesis of gem-dimethylcyclobutane natural products. Nat. Prod. Rep. 2019, 36, 1383–1393. [Google Scholar] [CrossRef]
- Navale, G.R.; Dharne, M.S.; Shinde, S.S. Metabolic engineering and synthetic biology for isoprenoid production in Escherichia coli and Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2021, 105, 457–475. [Google Scholar] [CrossRef]
- Liu, C.L.; Xue, K.; Yang, Y.; Liu, X.; Li, Y.; Lee, T.S.; Bai, Z.; Tan, T. Metabolic engineering strategies for sesquiterpene production in microorganism. Crit. Rev. Biotechnol. 2022, 42, 73–92. [Google Scholar] [CrossRef] [PubMed]
- Klaus, O.; Hilgers, F.; Nakielski, A.; Hasenklever, D.; Jaeger, K.E.; Axmann, I.M.; Drepper, T. Engineering phototrophic bacteria for the production of terpenoids. Curr. Opin. Biotechnol. 2022, 77, 102764. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhi, H.; Fang, Z.; Zhang, P. Genetic engineering of yeast, filamentous fungi and bacteria for terpene production and applications in food industry. Food Res. Int. 2021, 147, 110487. [Google Scholar] [CrossRef] [PubMed]
- Schempp, F.M.; Drummond, L.; Buchhaupt, M.; Schrader, J. Microbial cell factories for the production of terpenoid flavor and fragrance compounds. J. Agric. Food Chem. 2018, 66, 2247–2258. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Zhang, K.; Guo, J.; Yang, Q.; Li, Y.; Xian, M.; Zhang, R. Highly efficient biosynthesis of β-caryophyllene with a new sesquiterpene synthase from tobacco. Biotechnol. Biofuels Bioprod. 2022, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Hilgers, F.; Habash, S.S.; Loeschcke, A.; Ackermann, Y.S.; Neumann, S.; Heck, A.; Klaus, O.; Hage-Hülsmann, J.; Grundler, F.M.W.; Jaeger, K.E.; et al. Heterologous production of β-caryophyllene and evaluation of its activity against plant pathogenic fungi. Microorganisms 2021, 9, 168. [Google Scholar] [CrossRef]
- Reinsvold, R.E.; Jinkerson, R.E.; Radakovits, R.; Posewitz, M.C.; Basu, C. The production of the sesquiterpene β-caryophyllene in a transgenic strain of the cyanobacterium Synechocystis. J. Plant Physiol. 2011, 168, 848–852. [Google Scholar] [CrossRef]
- Lei, C.; Shubin, L.; Sun, T.; Zhang, W. Synechococcus Genetically Engineered Bacteria for Biosynthesizing Caryophyllene and Its Construction Methods and Applications. CN111394383A, 10 July 2020. [Google Scholar]
- Muthusamy, S.; Vetukuri, R.R.; Lundgren, A.; Ganji, S.; Zhu, L.H.; Brodelius, P.E.; Kanagarajan, S. Transient expression and purification of β-caryophyllene synthase in Nicotiana benthamiana to produce β-caryophyllene in vitro. PeerJ 2020, 8, e8904. [Google Scholar] [CrossRef]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. Int. J. Mol. Sci. 2021, 22, 5710. [Google Scholar] [CrossRef]
- Abbas, F.; Ke, Y.; Yu, R.; Yue, Y.; Amanullah, S.; Jahangir, M.M.; Fan, Y. Volatile terpenoids: Multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 2017, 246, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv. Biochem. Eng. Biotechnol. 2015, 148, 63–106. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zha, W.; Li, W.; Wang, J.; You, A. Advances in the biosynthesis of terpenoids and their ecological functions in plant resistance. Int. J. Mol. Sci. 2023, 24, 11561. [Google Scholar] [CrossRef]
- Wang, Q.; Quan, S.; Xiao, H. Towards efficient terpenoid biosynthesis: Manipulating IPP and DMAPP supply. Bioresour. Bioprocess. 2019, 6, 6. [Google Scholar] [CrossRef]
- Zhou, F.; Pichersky, E. More is better: The diversity of terpene metabolism in plants. Curr. Opin. Plant Biol. 2020, 55, 1–10. [Google Scholar] [CrossRef]
- Nagegowda, D.A.; Gupta, P. Advances in biosynthesis, regulation, and metabolic engineering of plant specialized terpenoids. Plant Sci. 2020, 294, 110457. [Google Scholar] [CrossRef]
- Wang, C.; Liwei, M.; Park, J.B.; Jeong, S.H.; Wei, G.; Wang, Y.; Kim, S.W. Microbial platform for terpenoid production: Escherichia coli and yeast. Front. Microbiol. 2018, 9, 2460. [Google Scholar] [CrossRef]
- Durairaj, J.; Di Girolamo, A.; Bouwmeester, H.J.; de Ridder, D.; Beekwilder, J.; van Dijk, A.D. An analysis of characterized plant sesquiterpene synthases. Phytochemistry 2019, 158, 157–165. [Google Scholar] [CrossRef]
- Di Sotto, A.; Mancinelli, R.; Gullì, M.; Eufemi, M.; Mammola, C.L.; Mazzanti, G.; Di Giacomo, S. Chemopreventive potential of caryophyllane sesquiterpenes: An overview of preliminary evidence. Cancers 2020, 12, 3034. [Google Scholar] [CrossRef]
- He, S.M.; Wang, X.; Yang, S.C.; Dong, Y.; Zhao, Q.M.; Yang, J.L.; Cong, K.; Zhang, J.J.; Zhang, G.H.; Wang, Y.; et al. De novo transcriptome characterization of Rhodomyrtus tomentosa leaves and identification of genes involved in α/β-pinene and β-caryophyllene biosynthesis. Front. Plant Sci. 2018, 9, 1231. [Google Scholar] [CrossRef] [PubMed]
- Boncan, D.A.T.; Tsang, S.S.K.; Li, C.; Lee, I.H.T.; Lam, H.M.; Chan, T.F.; Hui, J.H.L. Terpenes and terpenoids in plants: Interactions with environment and insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef]
- Cai, Y.; Jia, J.W.; Crock, J.; Lin, Z.X.; Chen, X.Y.; Croteau, R. A cDNA clone for β-caryophyllene synthase from Artemisia annua. Phytochemistry 2002, 61, 523–529. [Google Scholar] [CrossRef]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, J.; Han, S.; Chong, S.L.; Meng, G.; Song, M.; Wang, Y.; Zhou, S.; Liu, C.; Lou, L.; et al. The chromosome-scale genome of Phoebe bournei reveals contrasting fates of terpene synthase (TPS)-a and TPS-b subfamilies. Plant Commun. 2022, 3, 100410. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Sun, X.; Li, D.; Wei, G.; Liu, L.; Chen, F.; Cai, Y.; Fan, H. Terpenoid biosynthesis in Dendrobium officinale: Identification of (E)-β-caryophyllene synthase and the regulatory MYB genes. Ind. Crops Prod. 2022, 182, 114875. [Google Scholar] [CrossRef]
- Feng, K.; Kan, X.Y.; Yan, Y.J.; Wang, Y.; Sun, N.; Yang, Z.Y.; Zhao, S.P.; Wu, P.; Li, L.J. Identification and characterization of terpene synthase OjTPS1 involved in β-caryophyllene biosynthesis in Oenanthe javanica (Blume) DC. Ind. Crops Prod. 2023, 192, 115998. [Google Scholar] [CrossRef]
- Kollner, T.G.; Held, M.; Lenk, C.; Hiltpold, I.; Turlings, T.C.J.; Gershenzon, J.; Degenhardta, J. A maize (E)-β-caryophyllene synthase implicated in indirect defense responses against herbivores is not expressed in most American maize varieties. Plant Cell 2008, 20, 482–494. [Google Scholar] [CrossRef]
- Wang, L.; Xu, G.; Li, L.; Ruan, M.; Bennion, A.; Wang, G.L.; Li, R.; Qu, S. The OsBDR1-MPK3 module negatively regulates blast resistance by suppressing the jasmonate signaling and terpenoid biosynthesis pathway. Proc. Natl. Acad. Sci. USA 2023, 120, e2211102120. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, G.; Huang, X.; Guo, H.; Su, X.; Han, L.; Zhang, Y.; Qi, Z.; Xiao, Y.; Cheng, H. Overexpression of the caryophyllene synthase gene GhTPS1 in cotton negatively affects multiple pests while attracting parasitoids. Pest Manag. Sci. 2020, 76, 1722–1730. [Google Scholar] [CrossRef]
- Yahyaa, M.; Tholl, D.; Cormier, G.; Jensen, R.; Simon, P.W.; Ibdah, M. Identification and characterization of terpene synthases potentially involved in the formation of volatile terpenes in carrot (Daucus carota L.) roots. J. Agric. Food Chem. 2015, 63, 4870–4878. [Google Scholar] [CrossRef]
- Nawade, B.; Shaltiel-Harpaz, L.; Yahyaa, M.; Kabaha, A.; Kedoshim, R.; Bosamia, T.C.; Ibdah, M. Characterization of terpene synthase genes potentially involved in black fig fly (Silba adipata) interactions with Ficus carica. Plant Sci. 2020, 298, 110549. [Google Scholar] [CrossRef] [PubMed]
- Frank, L.; Wenig, M.; Ghirardo, A.; van der Krol, A.; Vlot, A.C.; Schnitzler, J.P.; Rosenkranz, M. Isoprene and β-caryophyllene confer plant resistance via different plant internal signaling pathways. Plant Cell Environ. 2021, 44, 1151–1164. [Google Scholar] [CrossRef]
- Huang, A.C.; Osbourn, A. Plant terpenes that mediate below-ground interactions: Prospects for bioengineering terpenoids for plant protection. Pest Manag. Sci. 2019, 75, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Ignea, C.; Cvetkovic, I.; Loupassaki, S.; Kefalas, P.; Johnson, C.B.; Kampranis, S.C.; Makris, A.M. Improving yeast strains using recyclable integration cassettes, for the production of plant terpenoids. Microb. Cell Fact. 2011, 10, 4. [Google Scholar] [CrossRef]
- Ignea, C.; Trikka, F.A.; Kourtzelis, I.; Argiriou, A.; Kanellis, A.K.; Kampranis, S.C.; Makris, A.M. Positive genetic interactors of HMG2 identify a new set of genetic perturbations for improving sesquiterpene production in Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Nong, F.T.; Wang, Y.Z.; Yan, C.X.; Gu, Y.; Song, P.; Sun, X.M. Strategies for efficient production of recombinant proteins in Escherichia coli: Alleviating the host burden and enhancing protein activity. Microb. Cell Fact. 2022, 21, 191. [Google Scholar] [CrossRef]
- Bureau, J.A.; Oliva, M.E.; Dong, Y.; Ignea, C. Engineering yeast for the production of plant terpenoids using synthetic biology approaches. Nat. Prod. Rep. 2023, 40, 1822–1848. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, R.; Yang, Q.; Zhang, J.; Zhao, Y.; Zheng, Y.; Yang, J. Recent advances in the biosynthesis of isoprenoids in engineered Saccharomyces cerevisiae. Adv. Appl. Microbiol. 2021, 114, 1–35. [Google Scholar] [CrossRef]
- Daletos, G.; Katsimpouras, C.; Stephanopoulos, G. Novel strategies and platforms for industrial isoprenoid engineering. Trends Biotechnol. 2020, 38, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, C.; Wang, P.; Yan, X.; Zhou, Z. Production of sesquiterpenoids α-neoclovene and β-caryophyllene by engineered Saccharomyces cerevisiae. Synth. Biol. 2021, 2, 792–803. [Google Scholar] [CrossRef]
- Elison, G.L.; Xue, Y.; Song, R.; Acar, M. Insights into bidirectional gene expression control using the canonical GAL1/GAL10 promoter. Cell Rep. 2018, 25, 737–748.e4. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhou, C.; Guo, X.; Du, Z.; Cheng, Y.; Wang, Z.; He, X. Enhancing fluxes through the mevalonate pathway in Saccharomyces cerevisiae by engineering the HMGR and β-alanine metabolism. Microb. Biotechnol. 2022, 15, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Deng, H.; Zhou, C.; Du, Z.; Guo, X.; Cheng, Y.; He, X. Enhancement of β-caryophyllene biosynthesis in Saccharomyces cerevisiae via synergistic evolution of β-caryophyllene synthase and engineering the chassis. ACS Synth. Biol. 2023, 12, 1696–1707. [Google Scholar] [CrossRef]
- Godara, A.; Kao, K.C. Adaptive laboratory evolution of β-caryophyllene producing Saccharomyces cerevisiae. Microb. Cell Fact. 2021, 20, 106. [Google Scholar] [CrossRef]
- Hirasawa, T.; Maeda, T. Adaptive laboratory evolution of microorganisms: Methodology and application for bioproduction. Microorganisms 2023, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Mavrommati, M.; Daskalaki, A.; Papanikolaou, S.; Aggelis, G. Adaptive laboratory evolution principles and applications in industrial biotechnology. Biotechnol. Adv. 2022, 54, 107795. [Google Scholar] [CrossRef]
- Liang, B.; Yang, Q.; Zhang, X.; Zhao, Y.; Liu, Y.; Yang, J.; Wang, Z. Switching carbon metabolic flux for enhancing the production of sesquiterpene-based high-density biofuel precursor in Saccharomyces cerevisiae. Biotechnol. Biofuels Bioprod. 2023, 16, 124. [Google Scholar] [CrossRef]
- Ward, V.C.A.; Chatzivasileiou, A.O.; Stephanopoulos, G. Metabolic engineering of Escherichia coli for the production of isoprenoids. FEMS Microbiol. Lett. 2018, 365, fny079. [Google Scholar] [CrossRef]
- Yang, J.; Nie, Q. Engineering Escherichia coli to convert acetic acid to β-caryophyllene. Microb. Cell Fact. 2016, 15, 74. [Google Scholar] [CrossRef] [PubMed]
- Kutscha, R.; Pflügl, S. Microbial upgrading of acetate into value-added products-examining microbial diversity, bioenergetic constraints and metabolic engineering approaches. Int. J. Mol. Sci. 2020, 21, 8777. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lama, S.; Agrawal, D.; Kumar, V.; Park, S. Acetate as a potential feedstock for the production of value-added chemicals: Metabolism and applications. Biotechnol. Adv. 2021, 49, 107736. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Davis, R. One-pot bioconversion of algae biomass into terpenes for advanced biofuels and bioproducts. Algal Res. 2016, 17, 316–320. [Google Scholar] [CrossRef]
- Wu, W.; Liu, F.; Davis, R.W. Engineering Escherichia coli for the production of terpene mixture enriched in caryophyllene and caryophyllene alcohol as potential aviation fuel compounds. Metab. Eng. Commun. 2018, 6, 13–21. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Yang, J.; Wang, X.; Yi, X.; Nie, Q. A Method of Synthesizing Beta-Caryophyllene by Microbial Catalysis and a Reconstituted Cell Capable of Synthesizing the Beta-Caryophyllene. CN104120141A, 29 October 2014. [Google Scholar]
- Zhang, R.; Li, Y.X.; Zhang, K.; Yang, A.; Wang, Y. Engineering Bacterium for Producing β-Caryophyllene and Construction Method and Application Thereof. CN111004763A, 14 April 2020. [Google Scholar]
- Grozdev, L.; Kaiser, J.; Berensmeier, S. One-step purification of microbially produced hydrophobic terpenes via process chromatography. Front. Bioeng. Biotechnol. 2019, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Troost, K.; Loeschcke, A.; Hilgers, F.; Özgür, A.Y.; Weber, T.M.; Santiago-Schübel, B.; Svensson, V.; Hage-Hülsmann, J.; Habash, S.S.; Grundler, F.M.W.; et al. Engineered Rhodobacter capsulatus as a phototrophic platform organism for the synthesis of plant sesquiterpenoids. Front. Microbiol. 2019, 10, 1998. [Google Scholar] [CrossRef]
- Orsi, E.; Folch, P.L.; Monje-López, V.T.; Fernhout, B.M.; Turcato, A.; Kengen, S.W.M.; Eggink, G.; Weusthuis, R.A. Characterization of heterotrophic growth and sesquiterpene production by Rhodobacter sphaeroides on a defined medium. J. Ind. Microbiol. Biotechnol. 2019, 46, 1179–1190. [Google Scholar] [CrossRef]
- Satta, A.; Esquirol, L.; Ebert, B.E. Current metabolic engineering strategies for photosynthetic bioproduction in cyanobacteria. Microorganisms 2023, 11, 455. [Google Scholar] [CrossRef]
- Herold, R.A.; Bryan, S.J. Engineered terpenoid production in Synechocystis sp. PCC 6803 under different growth conditions. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rodrigues, J.S.; Lindberg, P. Metabolic engineering of Synechocystis sp. PCC 6803 for improved bisabolene production. Metab. Eng. Commun. 2021, 12, e00159. [Google Scholar] [CrossRef] [PubMed]
- Dietsch, M.; Behle, A.; Westhoff, P.; Axmann, I.M. Metabolic engineering of Synechocystis sp. PCC 6803 for the photoproduction of the sesquiterpene valencene. Metab. Eng. Commun. 2021, 13, e00178. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Garin, V.; Chenebault, C.; Diaz-Santos, E.; Vincent, M.; Sassi, J.F.; Cassier-Chauvat, C.; Chauvat, F. Exploring the potential of the model cyanobacterium Synechocystis PCC 6803 for the photosynthetic production of various high-value terpenes. Biotechnol. Biofuels Bioprod. 2022, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xu, X.; Wu, Y.; Sun, H.; Luan, G.; Lu, X. Conversion of carbon dioxide into valencene and other sesquiterpenes with metabolic engineered Synechocystis sp. PCC 6803 cell factories. GCB Bioenergy 2023, 15, 1154–1165. [Google Scholar] [CrossRef]
- Lei, C.; Shubin, L.; Sun, T.; Zhang, W. Synechococcus Genetic Engineering Bacteria for Biosynthesizing Caryophyllene, and Construction Method and Application Thereof. CN111394383B, 4 October 2020. [Google Scholar]
- Rinaldi, M.A.; Ferraz, C.A.; Scrutton, N. Alternative metabolic pathways and strategies to high-titre terpenoid production in Escherichia coli. Nat. Prod. Rep. 2022, 39, 90–118. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.H.; Kaufmann-Malaga, B.B.; Lerman, J.A.; Dougherty, D.P.; Zhang, Y.; Kilbo, A.L.; Wilson, E.H.; Ng, C.Y.; Erbilgin, O.; Curran, K.C.; et al. An Automated scientist to design and optimize microbial strains for the industrial production of small molecules. bioRxiv 2023. [Google Scholar] [CrossRef]
- Gurdo, N.; Volke, D.C.; McCloskey, D.; Nikel, P.I. Automating the design-build-test-learn cycle towards next-generation bacterial cell factories. New Biotechnol. 2023, 74, 1–15. [Google Scholar] [CrossRef]
- Kwak, S.; Crook, N.; Yoneda, A.; Ahn, N.; Ning, J.; Cheng, J.; Dantas, G. Functional mining of novel terpene synthases from metagenomes. Biotechnol. Biofuels 2022, 15, 104. [Google Scholar] [CrossRef]
- Jin, K.; Xia, H.; Liu, Y.; Li, J.; Du, G.; Lv, X.; Liu, L. Compartmentalization and transporter engineering strategies for terpenoid synthesis. Microb. Cell Fact. 2022, 21, 92. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, H.; Chen, X.; Liu, L. Engineering microbial membranes to increase stress tolerance of industrial strains. Metab. Eng. 2019, 53, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Lozada, N.J.H.; Pfleger, B.F. Efflux systems in bacteria and their metabolic engineering applications. Appl. Microbiol. Biotechnol. 2015, 99, 9381–9393. [Google Scholar] [CrossRef] [PubMed]
- Osborne, M.G.; Geiger, C.J.; Corzett, C.H.; Kram, K.E.; Finkel, S.E. Removal of toxic volatile compounds in batch culture prolongs stationary phase and delays death of Escherichia coli. Appl. Environ. Microbiol. 2021, 87, e0186021. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.L.; Li, J.; Nong, F.T.; Yan, C.X.; Ma, W.; Zhu, X.F.; Zhang, L.; Sun, X.M. Application of adaptive laboratory evolution in lipid and terpenoid production in yeast and microalgae. ACS Synth. Biol. 2023, 12, 1396–1407. [Google Scholar] [CrossRef]
- Ting, W.W.; Ng, I.S. Adaptive laboratory evolution and metabolic regulation of genetic Escherichia coli W3110 toward low-carbon footprint production of 5-aminolevulinic acid. J. Taiwan Inst. Chem. Eng. 2022, 141, 104612. [Google Scholar] [CrossRef]
- Moser, S.; Pichler, H. Identifying and engineering the ideal microbial terpenoid production host. Appl. Microbiol. Biotechnol. 2019, 103, 5501–5516. [Google Scholar] [CrossRef]
Figure 1.
Biosynthesis of BCP and its isomers in plants. AACT—acetyl-CoA acetyltransferase; HMGS—hydroxymethylglutaryl-CoA synthase; HMGR—3-hydroxy-3-methylglutaryl-CoA reductase; MK—mevalonate kinase; PMK—phosphomevalonate kinase; MDD—mevalonate diphosphate decarboxylase; IDI—isopentenyl diphosphate isomerase; PMD—phosphomevalonate decarboxylase; IPK—isopentenyl phosphate kinase; DXS—1-deoxy-D-xylulose-5-phosphate synthase; DXR—1-deoxy-D-xylulose-5-phosphate reductoisomerase; MCT—2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; CMK—4-diphosphocytidyl-2-C-methyl-D-erythritol kinase; MDS—2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; HDS—4-hydroxy-3-methylbut-2-enyldiphosphate synthase; HDR—4-hydroxy-3-methylbut-2-enyl diphosphate reductase; GPPS—geranyl diphosphate synthase; and FPPS—farnesyl diphosphate synthase.
Figure 1.
Biosynthesis of BCP and its isomers in plants. AACT—acetyl-CoA acetyltransferase; HMGS—hydroxymethylglutaryl-CoA synthase; HMGR—3-hydroxy-3-methylglutaryl-CoA reductase; MK—mevalonate kinase; PMK—phosphomevalonate kinase; MDD—mevalonate diphosphate decarboxylase; IDI—isopentenyl diphosphate isomerase; PMD—phosphomevalonate decarboxylase; IPK—isopentenyl phosphate kinase; DXS—1-deoxy-D-xylulose-5-phosphate synthase; DXR—1-deoxy-D-xylulose-5-phosphate reductoisomerase; MCT—2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; CMK—4-diphosphocytidyl-2-C-methyl-D-erythritol kinase; MDS—2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; HDS—4-hydroxy-3-methylbut-2-enyldiphosphate synthase; HDR—4-hydroxy-3-methylbut-2-enyl diphosphate reductase; GPPS—geranyl diphosphate synthase; and FPPS—farnesyl diphosphate synthase.
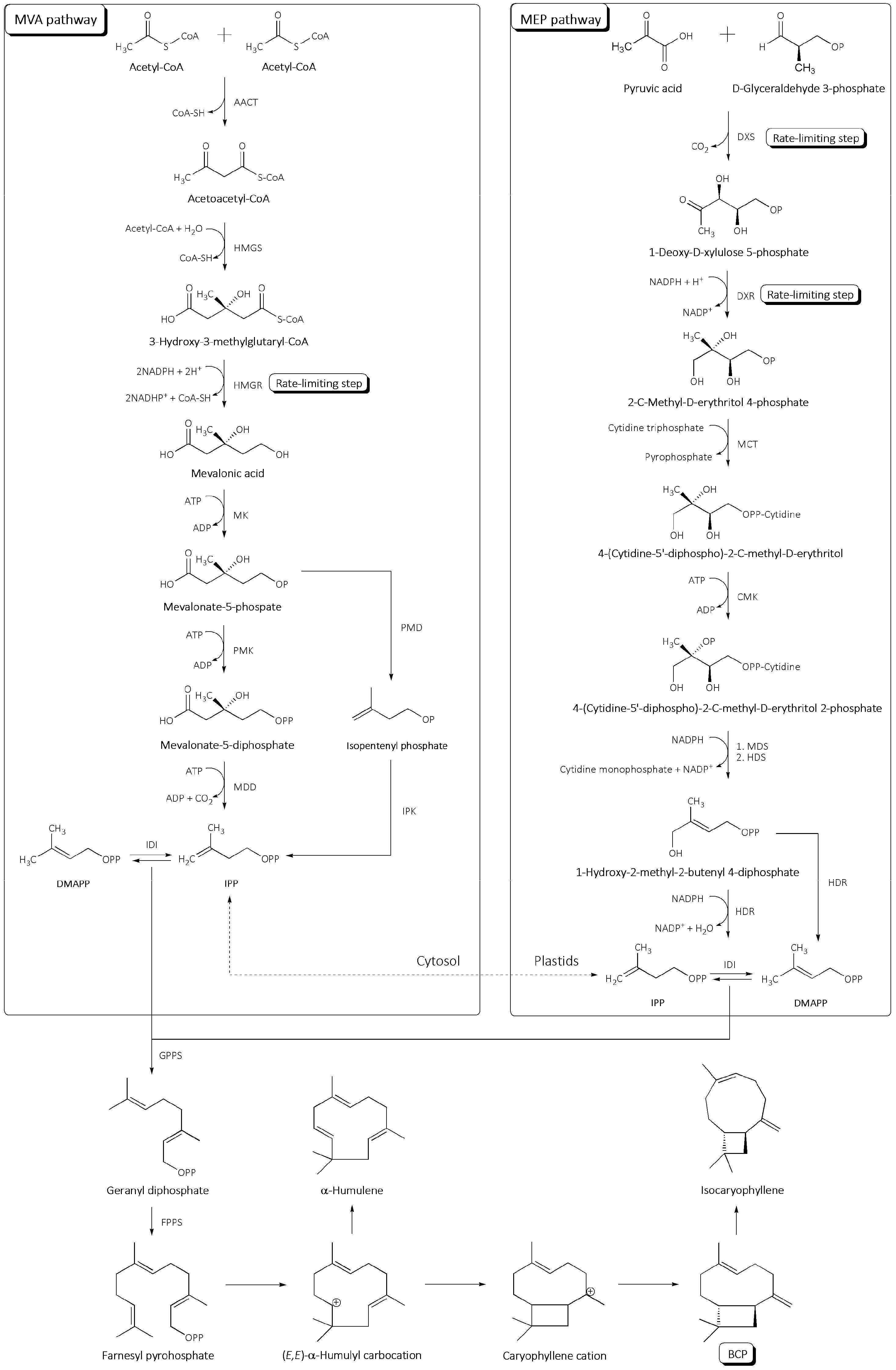
Figure 2.
Summary of the genetic manipulations of S. cerevisiae strains for BCP production. β-Alanine-metabolism-related enzymes: GAD1 (glutamate decarboxylase), PAN6 (pantothenate synthase), and CAB1 (pantothenate kinase); MVA pathway enzymes: ERG10 (acetyl-CoA acetyltransferase), ERG13 (hydroxymethylglutaryl-CoA synthase), tHMG1 (truncated 3-hydroxy-3-methylglutaryl-CoA reductase), EfHMGR (3-hydroxy-3-methylglutaryl-CoA reductase from E. faecalis), HMG2 (K6R) (stable variant of 3-hydroxy-3-methylglutaryl-CoA reductase), ERG12 (mevalonate kinase), ERG8 (phosphomevalonate kinase), ERG19 (mevalonate diphosphate decarboxylase), and IDI1 (isopentenyl diphosphate isomerase); Malonic acid-acetoacetyl-CoA pathway enzymes: MatB (malonyl-CoA synthase), and MatA (malonyl-CoA decarboxylase); downstream module enzymes: ERG20 (farnesyl diphosphate synthase), ERG9 (squalene synthase), QHS1 (terpene synthase from A. annua), QHS1E353D—QHS1 mutant, and Sf126 (terpene synthase from S. fruticosa); MVA pathway intermediates: HMG-CoA—3-hydroxy-3-methylglutaryl-CoA; MVA—mevalonic acid; MVAPP—5-diphosphomevalonic acid; IPP— and DMAPP—dimethylallyl diphosphate; downstream module intermediates: GPP—geranyl diphosphate and FPP—farnesyl pyrophosphate. BCP transportation genes: Ste6p (ATP-binding cassette transporter gene variant STE6T1025N) and Ste6 (ABC transporter).
Figure 2.
Summary of the genetic manipulations of S. cerevisiae strains for BCP production. β-Alanine-metabolism-related enzymes: GAD1 (glutamate decarboxylase), PAN6 (pantothenate synthase), and CAB1 (pantothenate kinase); MVA pathway enzymes: ERG10 (acetyl-CoA acetyltransferase), ERG13 (hydroxymethylglutaryl-CoA synthase), tHMG1 (truncated 3-hydroxy-3-methylglutaryl-CoA reductase), EfHMGR (3-hydroxy-3-methylglutaryl-CoA reductase from E. faecalis), HMG2 (K6R) (stable variant of 3-hydroxy-3-methylglutaryl-CoA reductase), ERG12 (mevalonate kinase), ERG8 (phosphomevalonate kinase), ERG19 (mevalonate diphosphate decarboxylase), and IDI1 (isopentenyl diphosphate isomerase); Malonic acid-acetoacetyl-CoA pathway enzymes: MatB (malonyl-CoA synthase), and MatA (malonyl-CoA decarboxylase); downstream module enzymes: ERG20 (farnesyl diphosphate synthase), ERG9 (squalene synthase), QHS1 (terpene synthase from A. annua), QHS1E353D—QHS1 mutant, and Sf126 (terpene synthase from S. fruticosa); MVA pathway intermediates: HMG-CoA—3-hydroxy-3-methylglutaryl-CoA; MVA—mevalonic acid; MVAPP—5-diphosphomevalonic acid; IPP— and DMAPP—dimethylallyl diphosphate; downstream module intermediates: GPP—geranyl diphosphate and FPP—farnesyl pyrophosphate. BCP transportation genes: Ste6p (ATP-binding cassette transporter gene variant STE6T1025N) and Ste6 (ABC transporter).
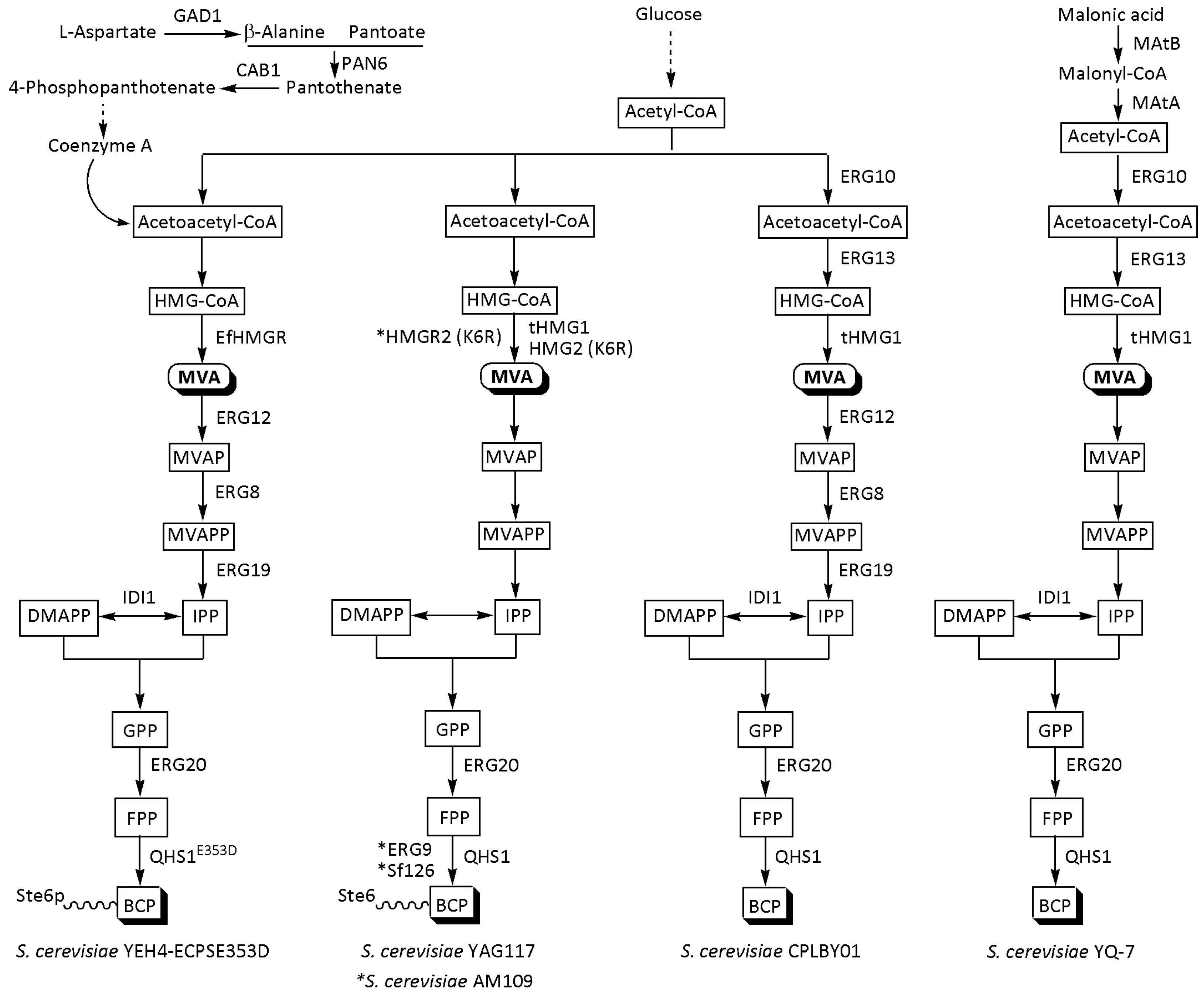
Figure 3.
Three main routes to BCP synthesis in E. coli using hybrid MVA pathway. Pentose phosphate pathway enzymes: G6PDH (glucose-6-phosphate dehydrogenase) and Pgi (phosphoglucose isomerase); acetic-acid-related enzymes: ACS—acetyl-CoA synthase from A. pasteurianus and nphT7 (acetoacetyl CoA synthase from Streptomyces sp. strain CL190); heterologous MVA pathway enzymes: MvaE (acetyl-CoA acetyltransferase/3-hydroxy-3-methylglutaryl-CoA reductase) and MvaS (3-hydroxy-3-methylglutaryl-CoA synthase) from E. faecalis; ERG12 (mevalonate kinase), ERG8 (phosphomevalonate kinase), ERG19 (mevalonate pyrophosphate decarboxylase), and IDI1 (isopentenyl diphosphate isomerase) from S. cerevisiae; downstream module enzymes: GPPS2 (geranyl diphosphate synthase from A. grandis), tps7 (β-caryophyllene synthase from N. tabacum), and QHS1 (terpene synthase from A. annua). MVA pathway intermediates, HMG-CoA—3-hydroxy-3-methylglutaryl-CoA; MVA—mevalonic acid; MVAPP—5-diphosphomevalonic acid; IPP—isopentenyl pyrophosphate and DMAPP—dimethylallyl diphosphate. MEP pathway intermediates: G3P—glyceraldehyde 3-phosphate; DXP—1-deoxy-D-xylulose 5-phosphate; MEP—2-C-methyl-D-erythritol 4-phosphate; CD-ME—4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol; CD-ME2P—4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol 2-phosphate; ME-cPP—2-C-methyl-D-erythritol 2,4-cyclodiphosphate; HMBPP—1-hydroxy-2-methyl-2-butenyl 4-diphosphate; IPP—isopentenyl pyrophosphate; DMAPP—dimethylallyl diphosphate. Downstream module intermediates: GPP—geranyl diphosphate and FPP—farnesyl pyrophosphate.
Figure 3.
Three main routes to BCP synthesis in E. coli using hybrid MVA pathway. Pentose phosphate pathway enzymes: G6PDH (glucose-6-phosphate dehydrogenase) and Pgi (phosphoglucose isomerase); acetic-acid-related enzymes: ACS—acetyl-CoA synthase from A. pasteurianus and nphT7 (acetoacetyl CoA synthase from Streptomyces sp. strain CL190); heterologous MVA pathway enzymes: MvaE (acetyl-CoA acetyltransferase/3-hydroxy-3-methylglutaryl-CoA reductase) and MvaS (3-hydroxy-3-methylglutaryl-CoA synthase) from E. faecalis; ERG12 (mevalonate kinase), ERG8 (phosphomevalonate kinase), ERG19 (mevalonate pyrophosphate decarboxylase), and IDI1 (isopentenyl diphosphate isomerase) from S. cerevisiae; downstream module enzymes: GPPS2 (geranyl diphosphate synthase from A. grandis), tps7 (β-caryophyllene synthase from N. tabacum), and QHS1 (terpene synthase from A. annua). MVA pathway intermediates, HMG-CoA—3-hydroxy-3-methylglutaryl-CoA; MVA—mevalonic acid; MVAPP—5-diphosphomevalonic acid; IPP—isopentenyl pyrophosphate and DMAPP—dimethylallyl diphosphate. MEP pathway intermediates: G3P—glyceraldehyde 3-phosphate; DXP—1-deoxy-D-xylulose 5-phosphate; MEP—2-C-methyl-D-erythritol 4-phosphate; CD-ME—4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol; CD-ME2P—4-(cytidine-5′-diphospho)-2-C-methyl-D-erythritol 2-phosphate; ME-cPP—2-C-methyl-D-erythritol 2,4-cyclodiphosphate; HMBPP—1-hydroxy-2-methyl-2-butenyl 4-diphosphate; IPP—isopentenyl pyrophosphate; DMAPP—dimethylallyl diphosphate. Downstream module intermediates: GPP—geranyl diphosphate and FPP—farnesyl pyrophosphate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main characteristics, fermentation type, and time of the microbial strains with best BCP titers.
Table 1.
Main characteristics, fermentation type, and time of the microbial strains with best BCP titers.
| Strain | Main Characteristics | Fermentation Type | Titer (mg/L) | Time (h) | Reference |
|---|---|---|---|---|---|
| S. cerevisiae | MVA pathway | ||||
| YEH4-ECPSE353D | β-Alanine metabolism genes (GAD1, PAN6, and CAB1); MVA genes (EfHMGR, ERG12, ERG8, CUP1, ERG19, IDI1, and ERG20); improved transmembrane transport of BCP (STE6T1025N), and 353D mutant of QHS1 (A. annua) | Test tube | 70.45 | 48 | [59] |
| Fed-batch | 594.05 | 96 | |||
| YAG117 | Deleted cytosol catalase gene (CTT1) to reduce yeast resistance to hydrogen peroxide, overexpressed codon-optimized QHS1 (A. annua), tHMG1, HMG2 (K6R), UPC2-1, and ERG20; mutations in MST27/tR(UCU)G1 and STE6 | Test tube | 104.7 ± 6.2 | n.m. | [60] |
| AM109 | HMG2 (K6R), deleted (Ubc7p, Ssm4p, and Pho86p), ERG9, and Sf126 (S. fruticosa) | Shake flask | 125.0 | * n.m. | [50] |
| CPLBY01 | MVA genes (ERG10, ERG13, tHMG1, ERG12, ERG8; tHMG1, ERG19, IDI1, and ERG20), and codon-optimized QHS1 (A. annua) | Shake flask | 250.4 | n.m. | [56] |
| Fed-batch | 2949.1 | 140 | |||
| YQ-7 | MVA genes (QHS1 from A. annua, tHMG1, IDI1, ERG20, ERG13, and ERG10), and artificial synthetic malonic acid-acetoacetyl-CoA metabolic pathway (Mae I from Schizosaccharomyces pombe, MatB, and ACCS) | Shake flask | 328 ± 13.44 | 144 | [63] |
| E. coli | Hybrid MVA pathway | ||||
| YJM67 | From acetic acid; QHS1 (A. annua), mvA and mvS (E. faecalis), GPPS2 (A. grandis), ERG12, ERG8, ERG19, and IDI1 (S. cerevisiae), ACS (A. pasteurianus), and nphT7 (Streptomyces sp.) | Shake flask | 22 ± 1.8 | 24 | [65] |
| Fed-batch | 1050 | 72 | |||
| CAR2 | mvaE and mvaS (E. faecalis), ERG12, ERG8, ERG19, and IDI1 (S. cerevisiae), tps7 (N. tabacum), and T7 promoter, Cm R | Shake flask | 100.3 | 24 | [19] |
| Fed-batch | 4319 | 60 | |||
| In situ fed-batch | 5142 | 64 | |||
| YJM59 | QHS1 (A. annua), mvA, and mvaS (E. faecalis), GPPS2 (A. grandis), C6PDH (E. coli), ERG12, ERG8, ERG19, and IDI1 (S. cerevisiae), and deleted pgi gene | Shake flask | 220 ± 6 | n.m. | [9] |
| Fed-batch | 1520.0 | 60 | |||
| R. capsulatus | Heterologous MVA pathway | ||||
| SB1003-MVA | MVA genes (P. zeaxanthinifaciens), QHS1 (A. annua), and ispA | Gas-tight hangate tubes, UV | 90 ± 19 | 72 | [20] |
| Screw-neck vials, IR | 139 ± 31 | 72 | |||
| Synechocystis sp. | MEP pathway | ||||
| PCC6803 | Stabilized QSH1 (A. annua) | Shake flask | 46.4 × 10−5 ± 2.9 | Week | [21] |
| S. elongatus | MEP pathway | ||||
| NSI-gpps-ispA-idi1sc-II-idi1sc-III-tps21 | gpps-ispA-idi1sc-II-idi1sc-III-tps21, where tps21 encodes TPS21 (A. thaliana) | Photobioreactor | 212.37 × 10−3 | 96 | [82] |
* n.m.—not mentioned.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsigoriyna, L.; Sango, C.; Batovska, D. An Update on Microbial Biosynthesis of β-Caryophyllene, a Sesquiterpene with Multi-Pharmacological Properties. Fermentation 2024, 10, 60. https://doi.org/10.3390/fermentation10010060
AMA Style
Tsigoriyna L, Sango C, Batovska D. An Update on Microbial Biosynthesis of β-Caryophyllene, a Sesquiterpene with Multi-Pharmacological Properties. Fermentation. 2024; 10(1):60. https://doi.org/10.3390/fermentation10010060
Chicago/Turabian StyleTsigoriyna, Lidia, Chakarvati Sango, and Daniela Batovska. 2024. "An Update on Microbial Biosynthesis of β-Caryophyllene, a Sesquiterpene with Multi-Pharmacological Properties" Fermentation 10, no. 1: 60. https://doi.org/10.3390/fermentation10010060
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.