
Microbial Community Shifts during Biogas Production from Biowaste and/or Propionate

Abstract
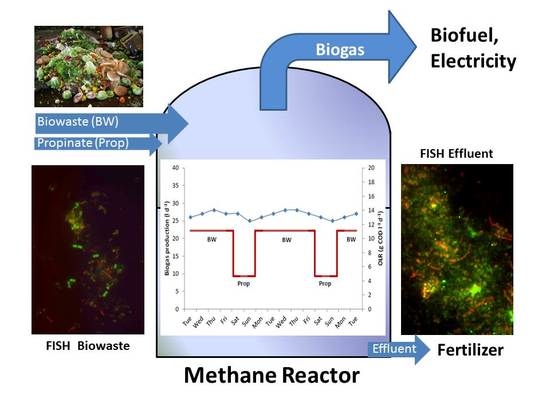
:
Bullet Points
- Anaerobic digestion of biowaste + propionate
- Periodic biowaste replacement by propionate
- Propionate-oxidizing bacteria and methanogenic archaea
- Constant biogas production with different substrates
- Community changes during different operational conditions
1. Introduction
2. Materials and Methods
2.1. Source of Fresh Biowaste and Digester Residues for Inoculation of Laboratory Reactors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Average Values |
|---|---|
| Total solids, TS (%) | 6.1 ± 0.5 |
| Volatile solids, VS (%) | 5.2 ± 0.4 |
| Chemical oxygen demand, COD (g·L−1) | 94–113 |
| Total Kjeldahl Nitrogen, TKN (g·L−1) | 2.2 ± 0.2 |
| NH4+-Nitrogen (g·L−1) | 0.5 ± 0.1 |
| pH | 4.5 |
| Acetate (g·L−1) | 3.1 ± 0.3 |
| Propionate (g·L−1) | 2.7 ± 0.3 |
| n-Butyrate (g·L−1) | 1.5 ± 0.2 |
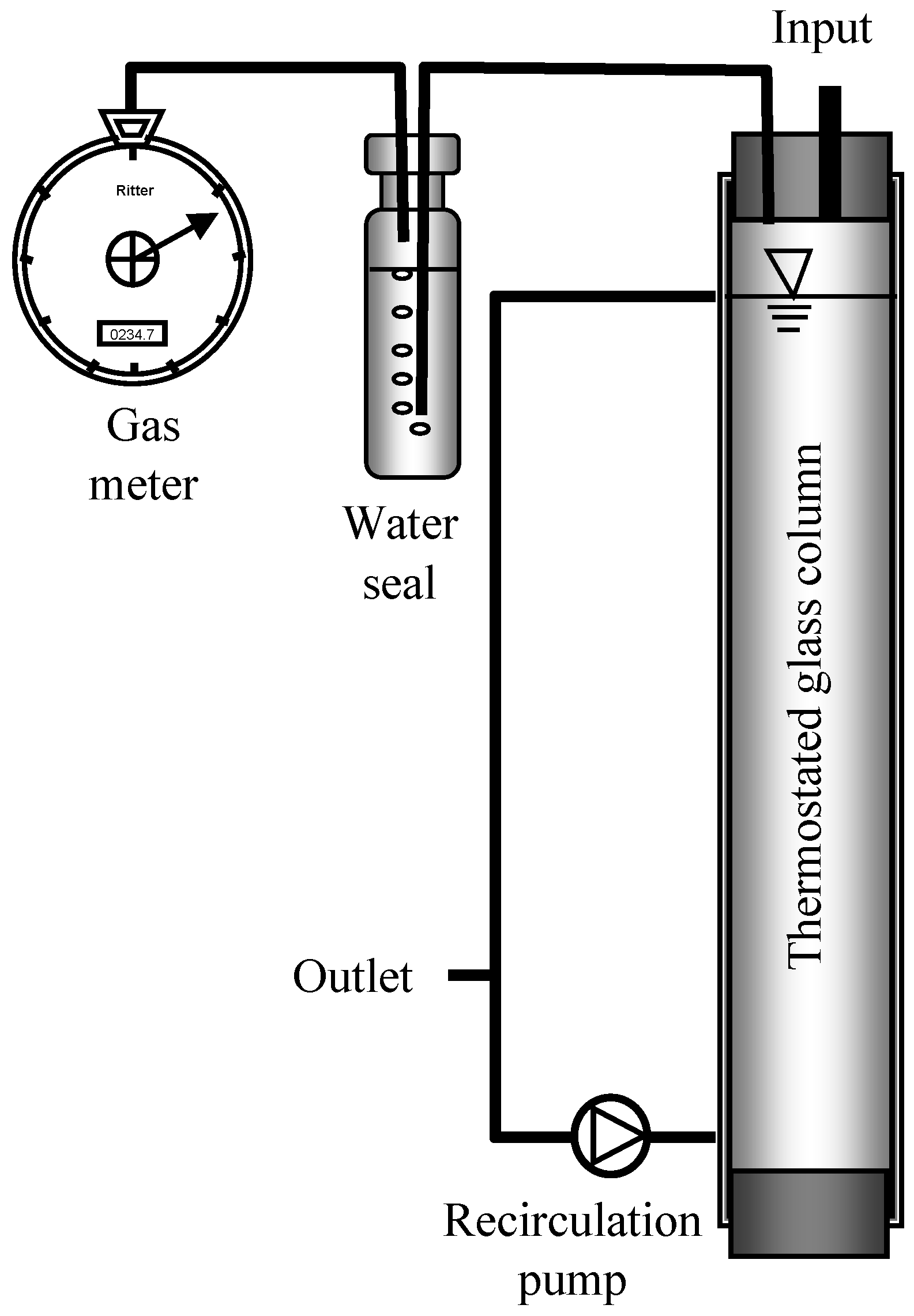
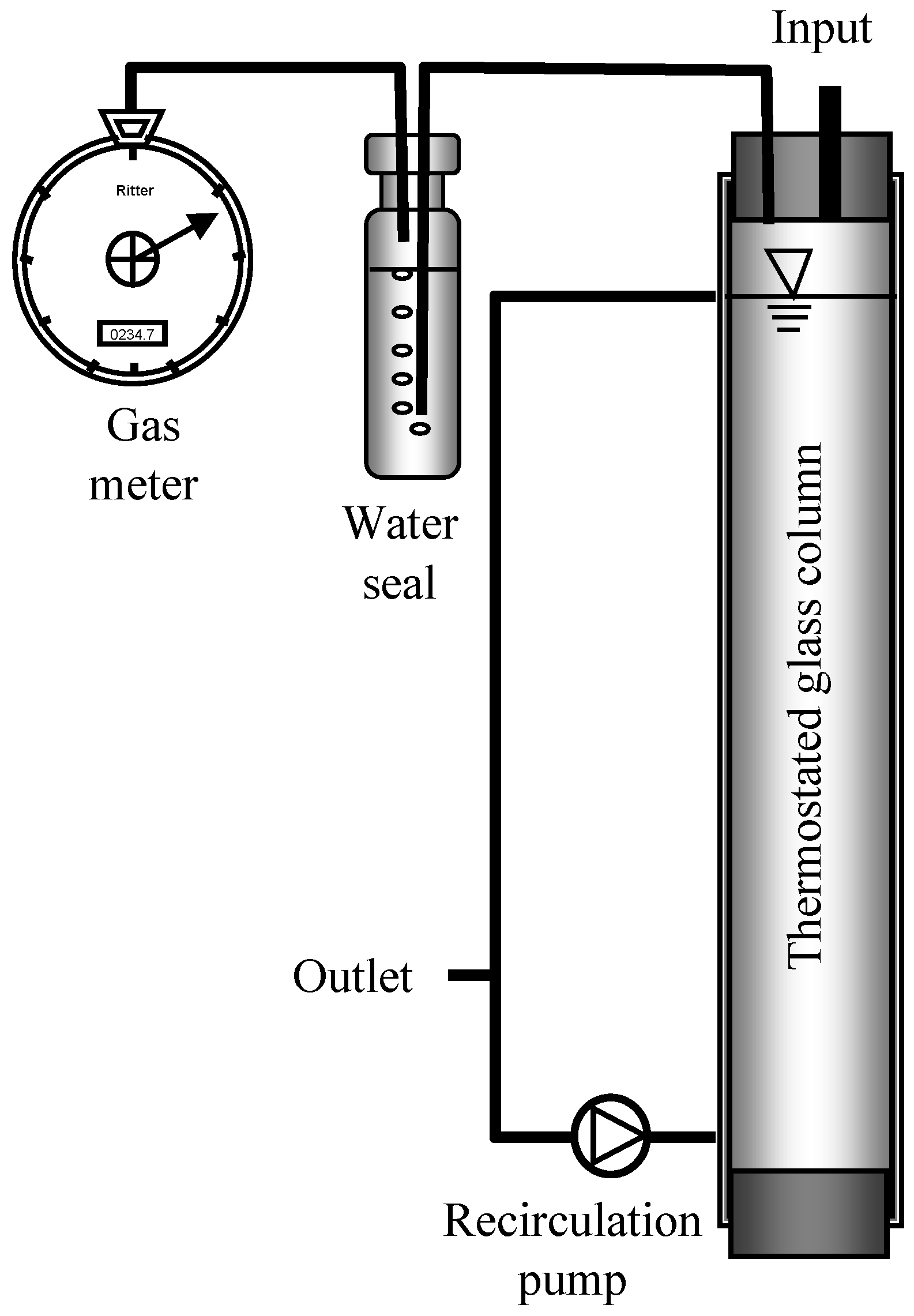
2.2. Reactor Setup, Feeding, and Incubation Conditions
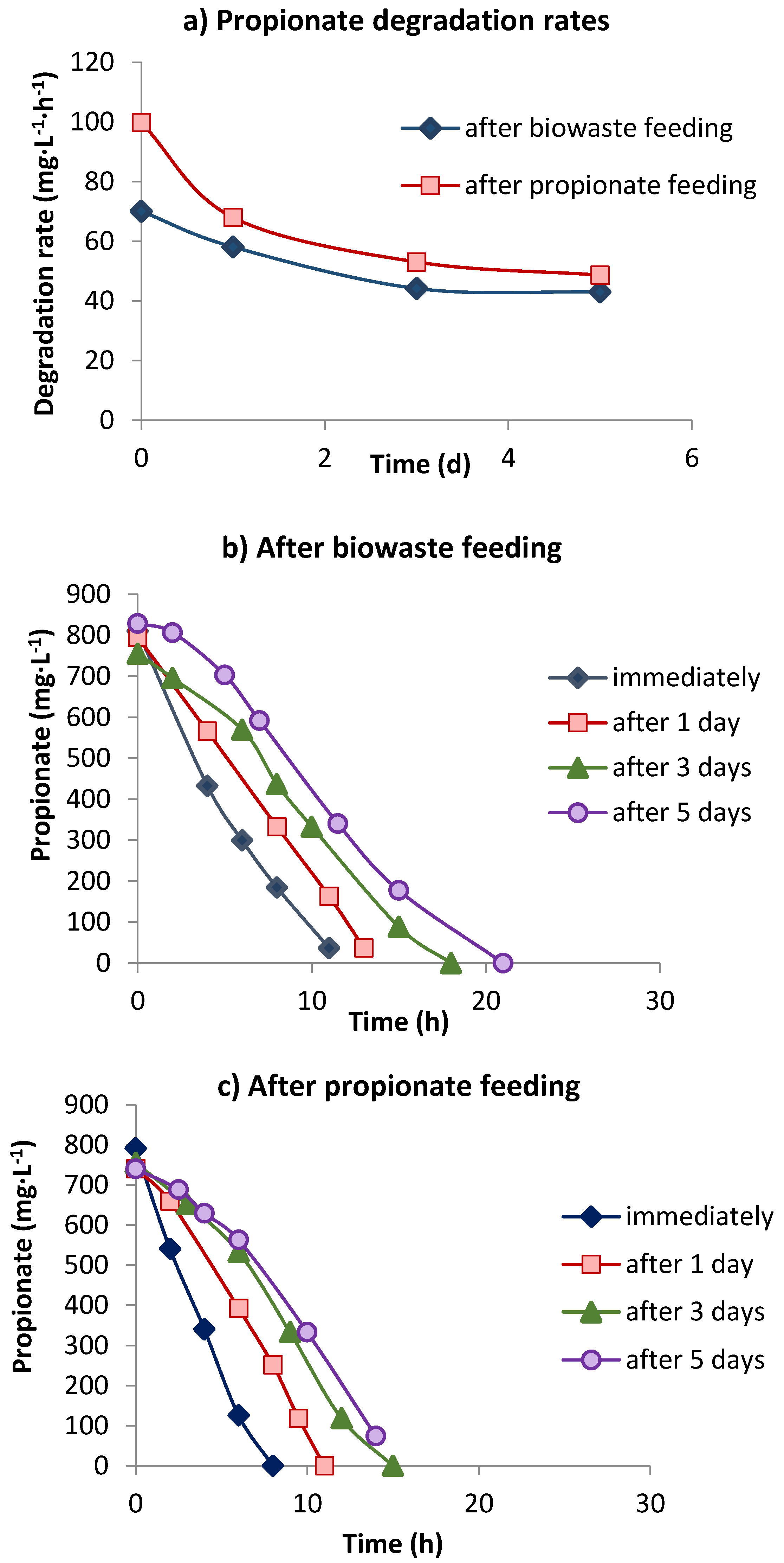
| Time (days) | OLR (kg COD·m−3·d−1) | Propionate Addition (g·L−1) | Degradation Rate (mg·L−1·h−1) |
|---|---|---|---|
| 2 | 12 | - | 40.4 b |
| 12 | 12 | - | 41.4 c |
| 30 | 12 +1 a | 0.7 | 54.8 b |
| 72 | 14 + 2.5 a | 1.5 | 70.8 b |
| 94 | 14 + 3.0 a | 1.9 | 99.9 b |
| 117 | 14 + 4.0 a | 2.5 | 109.2 b |
2.3. Analyses
2.4. Characterization of the Biowaste Community by Fluorescence in situ Hybridization (FISH)
2.5. Community Density Calculation
3. Results
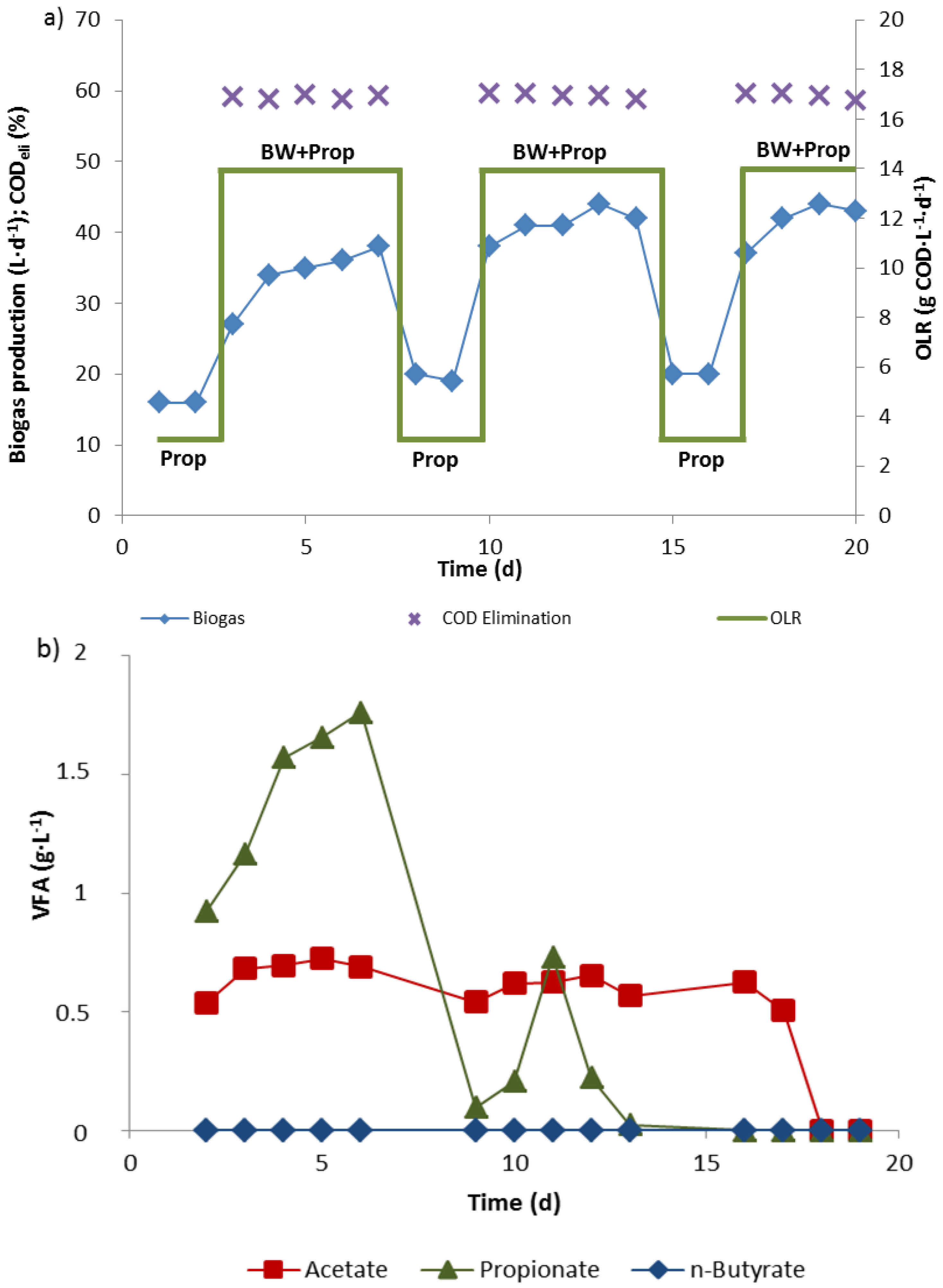
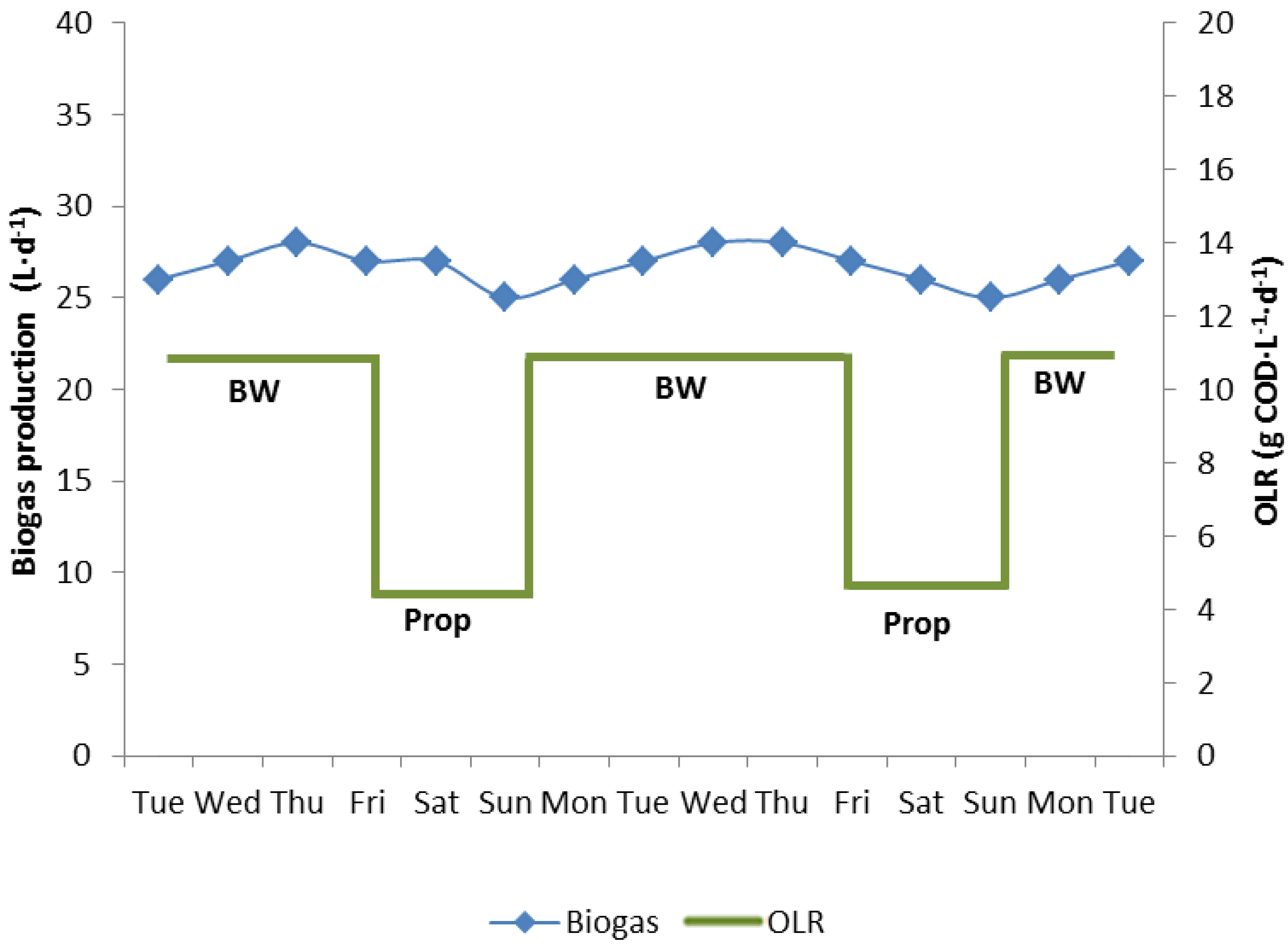
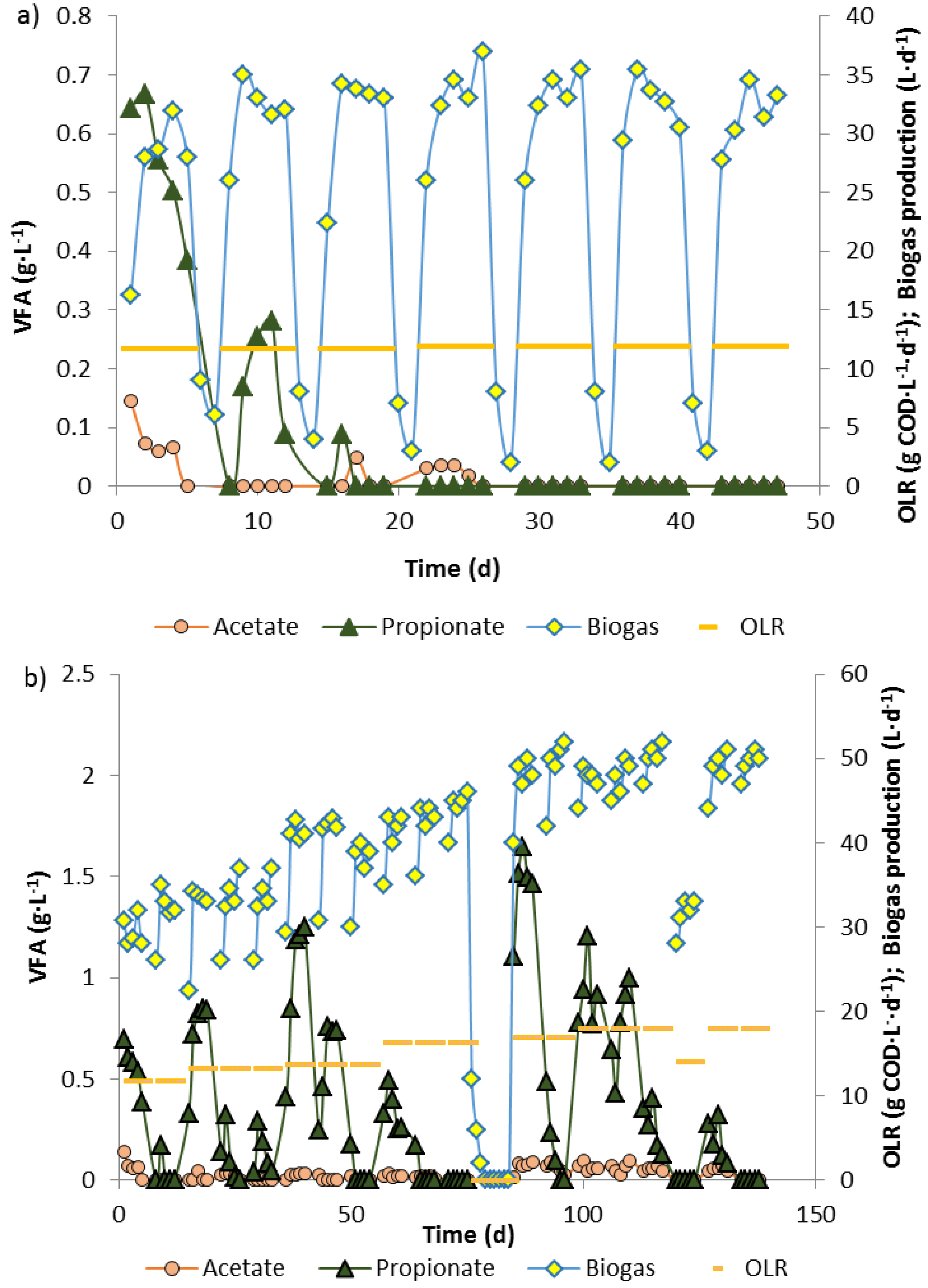
3.1. Biowaste Digestion and Co-Digestion Strategies of Propionate
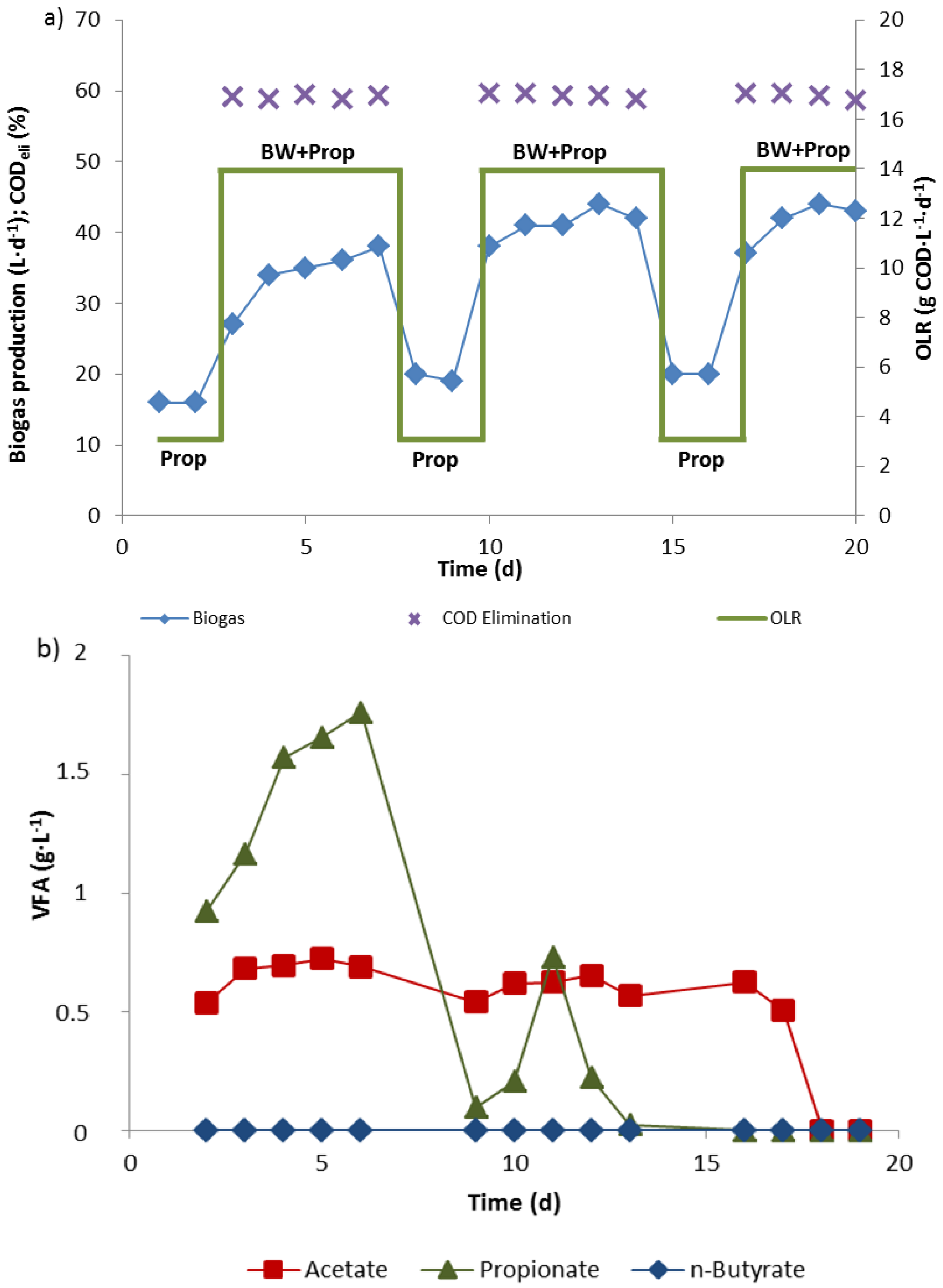
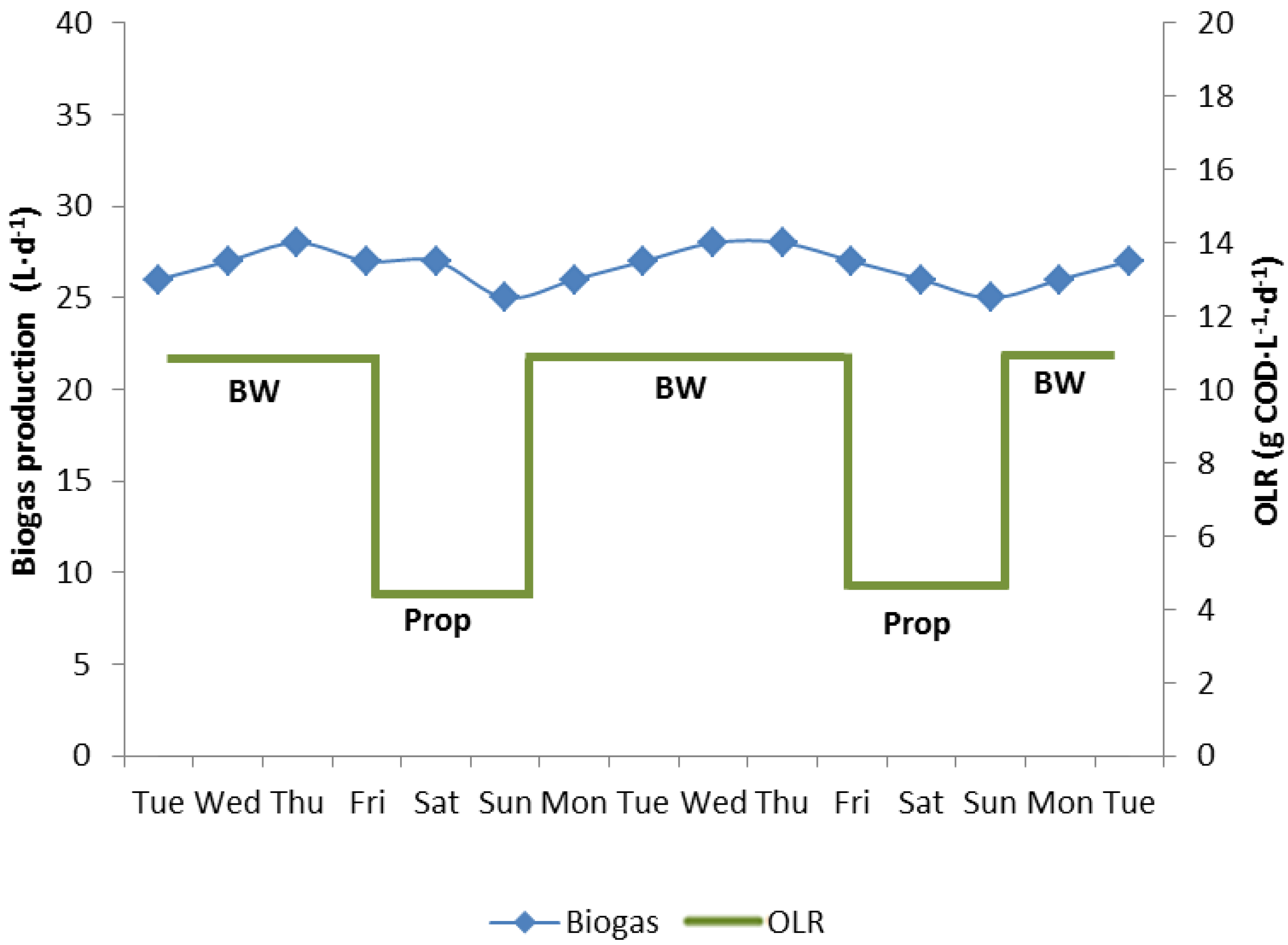
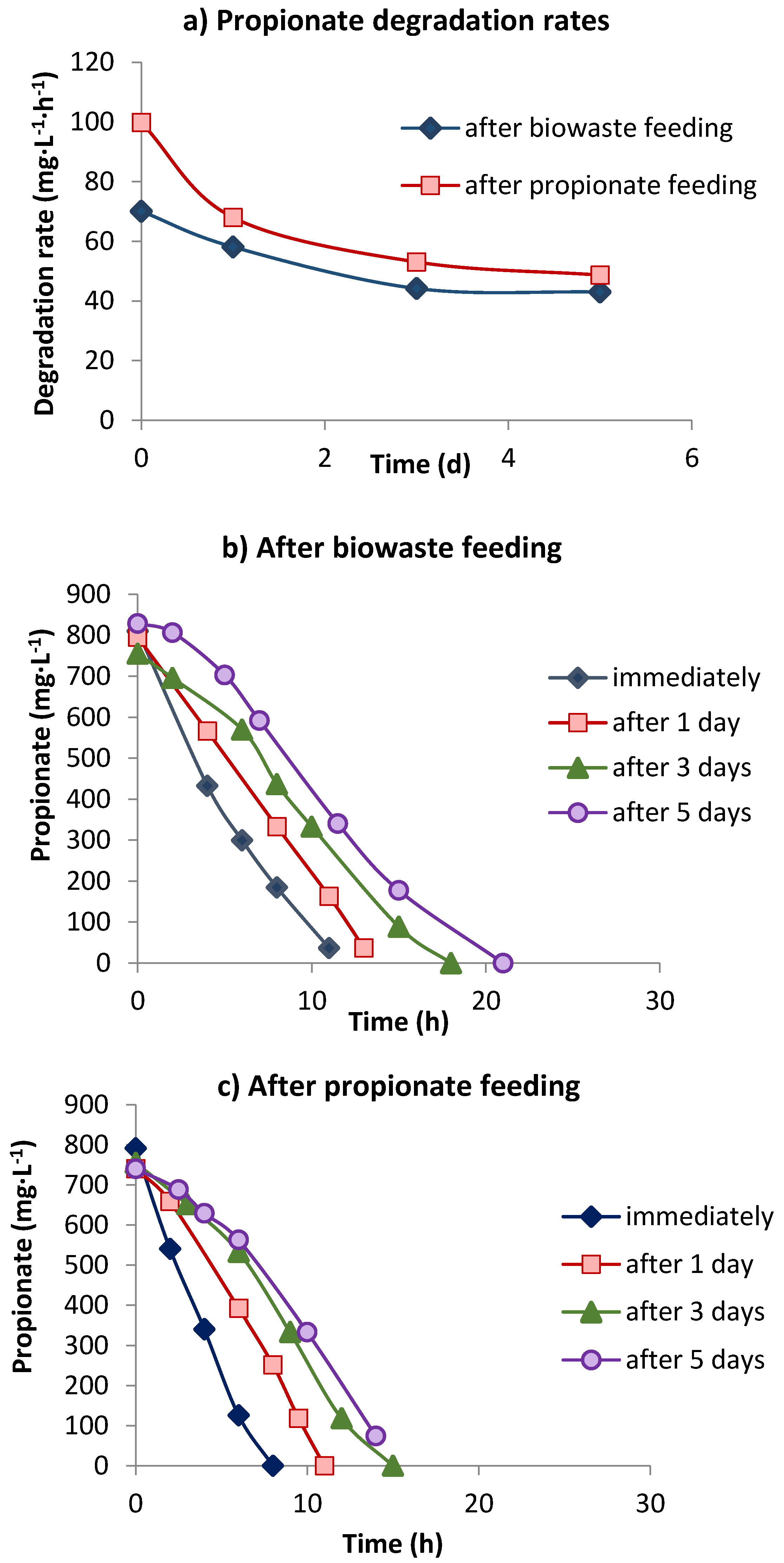
3.2. Propionate Degradation Rates
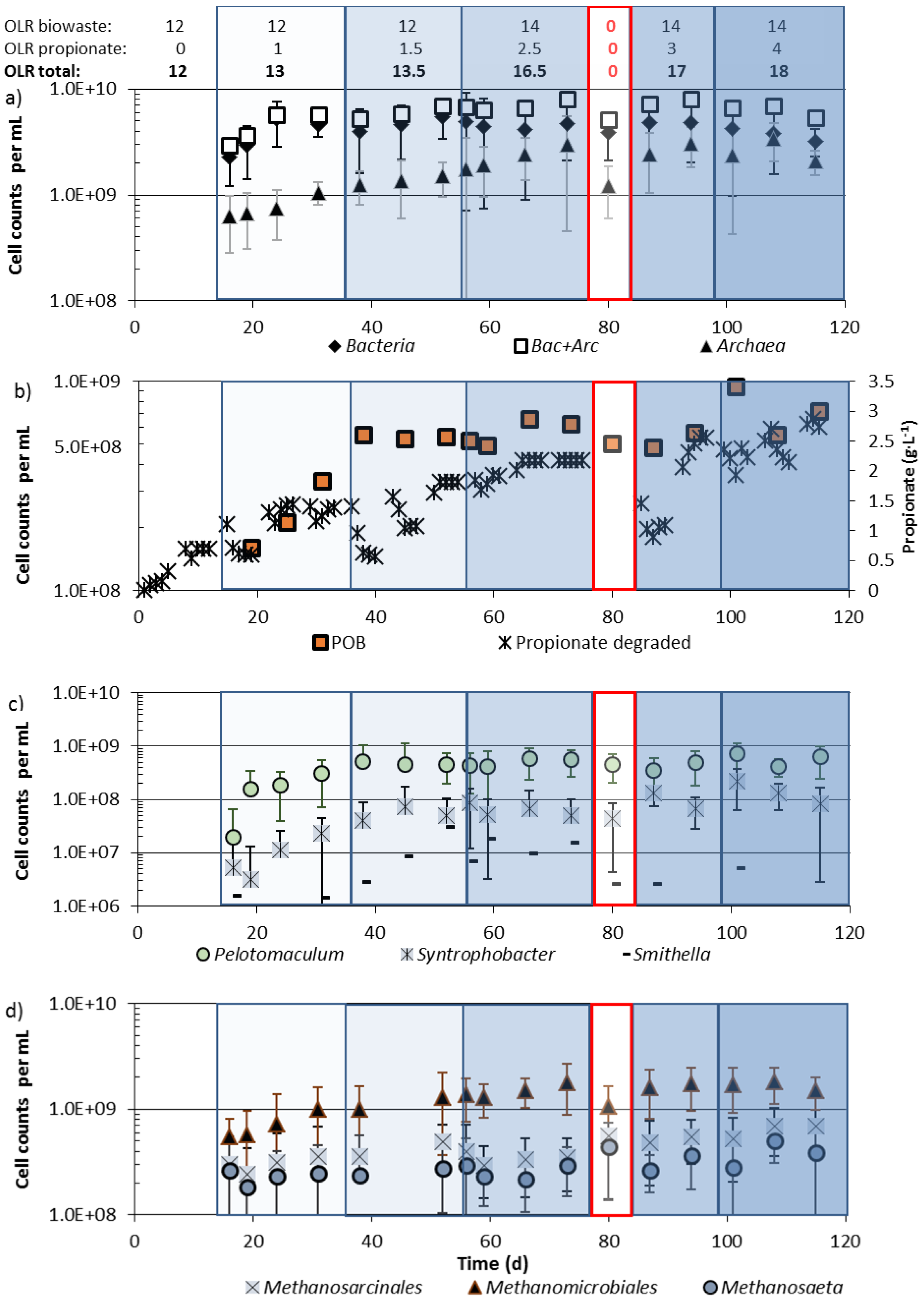
3.3. Community Changes during Biowaste and Propionate Degradation
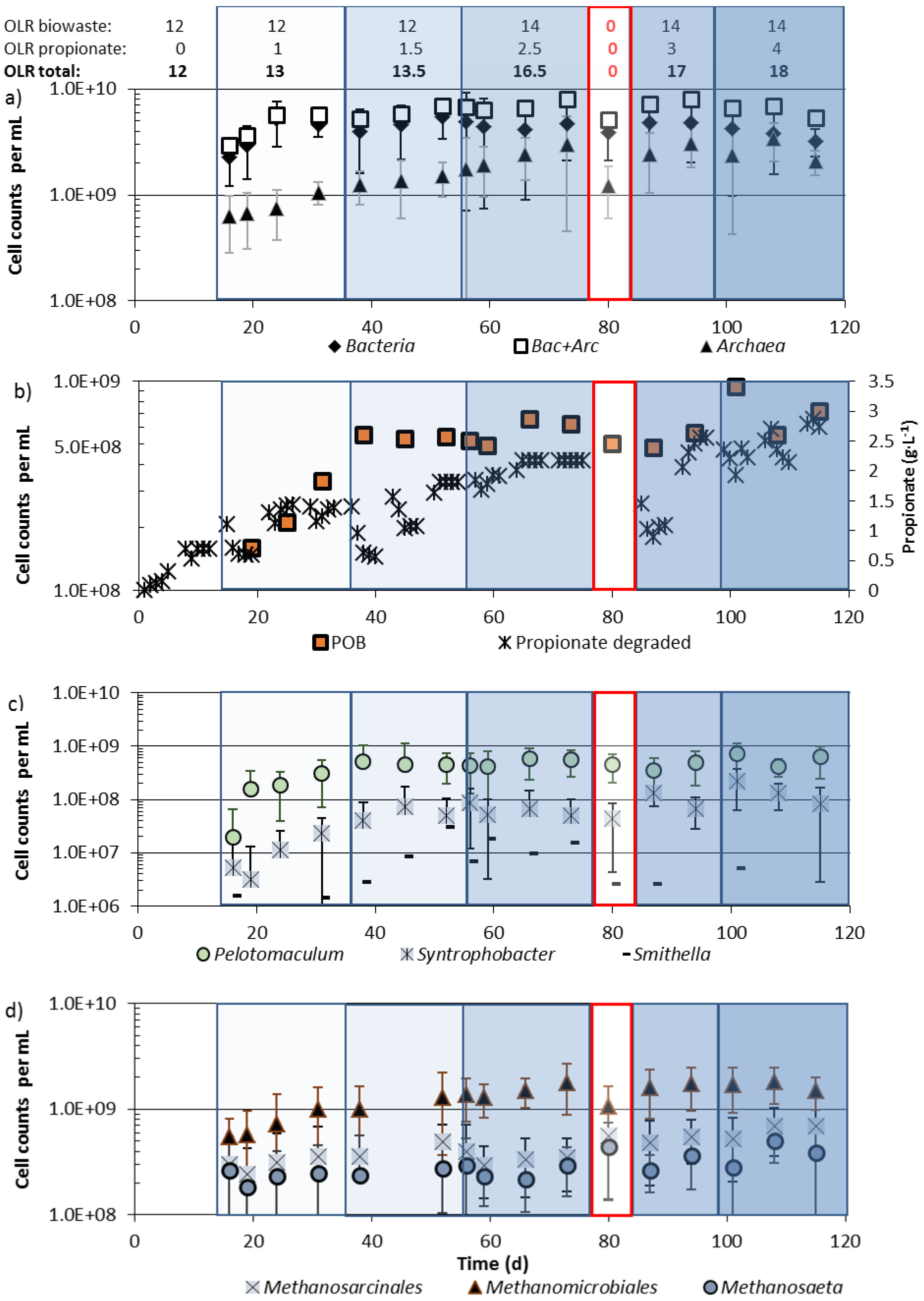
4. Discussion
4.1. Physiological Aspects of Biowaste and/or Propionate Degradation
4.2. Community Shifts during Propionate Degradation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McMahon, K.; Theng, D.; Stams, A.; Mackie, R.; Raskin, L. Microbial population dynamics during start up and overload conditions of anaerobic digesters treating municipal solid waste and sewage sludge. Biotechnol. Bioeng. 2004, 87, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Gallert, C.; Henning, A.; Winter, J. Scale-up of anaerobic digestion of the organic fraction from domestic wastes. Water Res. 2003, 37, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Gallert, C.; Winter, J. Bacterial metabolism in wastewater treatment systems. In Environmental Biotechnology—Concepts and Applications; Jördening, H.J., Winter, J., Eds.; Wiley-VCH: Weinheim, Germany, 2005; pp. 1–48. [Google Scholar]
- Gallert, C.; Winter, J. Propionic acid accumulation and degradation during start-up of anaerobic biowaste digestion. Bioresour. Technol. 2008, 99, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ban, Q.; Zhang, L.; Jha, A.K. Syntrophic propionate degradation in anaerobic digestion: A review. Int. J. Agric. Biol. 2012, 14, 843–850. [Google Scholar]
- Moertelmaier, C.; KIT, Karlsruhe Institute of Technology, Karlsruhe, Germany. Propionate-Oxidizing Bacteria in Anaerobic Biowaste Digesters. Unpublished Data. 2015. [Google Scholar]
- Moertelmaier, C.; Li, C.; Winter, J.; Gallert, C. Fatty acid metabolism and population dynamics in a wet biowaste digester during re-start after revision. Bioresour. Technol. 2014, 166, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Mörtelmaier, C.; Winter, J.; Gallert, C. Effect of moisture of municipal biowaste on start-up and efficiency of mesophilic and thermophilic dry anaerobic digestion. Bioresour. Technol. 2014, 168, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Gallert, C.; Winter, J. Verfahren der Abwasserreinigung. In Industrielle Mikrobiologie; Sahm, H., Antranikian, G., Stahmann, K.P., Takors, R., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2013; pp. 271–295. [Google Scholar]
- De Bok, F.; Harmsen, H.; Plugge, C.; de Vries, M.; Akkermans, A.; de Vos, W.; Stams, A. The first true obligately syntrophic propionate-oxidizing bacterium, Pelotomaculum schinkii sp. nov., co-cultured with Methanospirillum hungatei, and emended description of the genus Pelotomaculum. Int. J. Syst. Evolut. Microbiol. 2005, 55, 1697–1703. [Google Scholar] [CrossRef]
- Felchner-Zwirello, M.; Winter, J.; Gallert, C. Interspecies distances between propionic acid degraders and methanogens in syntrophic consortia for optimal hydrogen transfer. Appl. Microbiol. Biotechnol. 2013, 97, 9193–9205. [Google Scholar] [CrossRef] [PubMed]
- Shigematsu, T.; Era, S.; Mizuno, Y.; Ninomiya, K.; Kamegawa, K.; Morimura, S.; Kima, K. Microbial community of a mesophilic propionate-degrading methanogenic consortium in chemostat cultivation analyzed based on 16S rRNA and acetate kinase genes. Appl. Microbiol. Biotechnol. 2006, 72, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Ban, Q.; Li, J.; Zhang, L.; Jha, A.K.; Zhang, Y. Quantitative analysis of previously identified propionate-oxidizing bacteria and methanogens at different temperatures in an UASB reactor containing propionate as a sole carbon source. Appl. Biochem. Biotechnol. 2013, 171, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Ariesyady, H.; Ito, T.; Yoshiguchi, K.; Okabe, S. Phylogenetic and functional diversity of propionate-oxidizing bacteria in an anaerobic digester sludge. Appl. Microbiol. Biotechnol. 2007, 75, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Rui, J.; Pei, Z.; Sun, X.; Zhang, S.; Yan, Z.; Wang, Y.; Liu, X.; Zheng, T.; Li, X. Straw- and slurry-associated prokaryotic communities differ during co-fermentation of straw and swine manure. Appl. Microbiol. Biotechnol. 2014, 98, 4771–4780. [Google Scholar] [CrossRef] [PubMed]
- Hori, T.; Haruta, S.; Ueno, Y.; Ishii, M.; Igarashi, Y. Dynamic transition of a methanogenic population in response to the concentration of volatile fatty acids in a thermophilic anaerobic digester. Appl. Environ. Microbiol. 2006, 72, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Nayono, S.E.; Gallert, C.; Winter, J. Foodwaste as a co-substrate in a fed-batch anaerobic biowaste digester for constant biogas supply. Water Sci. Technol. 2009, 59, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- APHA. AWWA; WEF. In Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC; New York, NY, USA, 2005. [Google Scholar]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucloetide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [PubMed]
- Daims, H.; Lücker, S.; Wagner, M. Daime, a novel image analysis programe for microbial ecology and biofilm research. Environ. Microbiol. 2006, 8, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Pullammanppallil, P.C.; Chynoweth, D.P.; Lyberatos, G.; Svoronos, S.A. Stable performance of anaerobic digestion in the presence of a high concentration propionic acid. Bioresour. Technol. 2001, 78, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Amani, T.; Nosrati, M.; Mousavi, S.M.; Kermanshahi, R.K. Study of syntrophic anaerobic digestion of volatile fatty acids using enriched cultures at mesophilic conditions. Int. J. Environ. Res. 2011, 5, 333–342. [Google Scholar]
- Nayono, S.E.; Gallert, C.; Winter, J. Co-digestion of press water and food waste in a biowaste digester for improvement of biogas production. Bioresour. Technol. 2010, 101, 6998–7004. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Kao, J.; Stabnikova, O. Digestion of pre-treated food waste in a hybrid anaerobic solid–liquid (HASL) system. J. Chem. Technol. Biotechnol. 2006, 81, 345–351. [Google Scholar] [CrossRef]
- Angenent, L.; Shihwu, S.; Raskin, L. Methanogenic population dynamics during startup of a full-scale anaerobic sequencing batch reactor treating swine waste. Water Res. 2002, 36, 4648–4654. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Moertelmaier, C.; Winter, J.; Gallert, C. Co-digestion of wheat and rye bread suspensions with the source-sorted municipal biowaste fraction. Waste Manag. 2015. revised. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Moertelmaier, C.; Winter, J.; Gallert, C. Microbial Community Shifts during Biogas Production from Biowaste and/or Propionate. Bioengineering 2015, 2, 35-53. https://doi.org/10.3390/bioengineering2010035
Li C, Moertelmaier C, Winter J, Gallert C. Microbial Community Shifts during Biogas Production from Biowaste and/or Propionate. Bioengineering. 2015; 2(1):35-53. https://doi.org/10.3390/bioengineering2010035
Chicago/Turabian StyleLi, Chaoran, Christoph Moertelmaier, Josef Winter, and Claudia Gallert. 2015. "Microbial Community Shifts during Biogas Production from Biowaste and/or Propionate" Bioengineering 2, no. 1: 35-53. https://doi.org/10.3390/bioengineering2010035