Can Zymomonas mobilis Substitute Saccharomyces cerevisiae in Cereal Dough Leavening?


Dipartimento di Scienze per gli Alimenti, la Nutrizione, l’Ambiente, Università degli Studi di Milano, 20133 Milan, Italy
*
Author to whom correspondence should be addressed.
Foods 2018, 7(4), 61; https://doi.org/10.3390/foods7040061
Submission received: 28 March 2018
/
Revised: 12 April 2018
/
Accepted: 13 April 2018
/
Published: 16 April 2018
(This article belongs to the Special Issue Grain-based Foods: Processing, Properties, and Heath Attributes)
Abstract
:Baker’s yeast intolerance is rising among Western populations, where Saccharomyces cerevisiae is spread in fermented food and food components. Zymomonas mobilis is a bacterium commonly used in tropical areas to produce alcoholic beverages, and it has only rarely been considered for dough leavening probably because it only ferments glucose, fructose and sucrose, which are scarcely present in flour. However, through alcoholic fermentation, similarly to S. cerevisiae, it provides an equimolar mixture of ethanol and CO2 that can rise a dough. Here, we propose Z. mobilis as a new leavening agent, as an alternative to S. cerevisiae, overcoming its technological limit with different strategies: (1) adding glucose to the dough formulation; and (2) exploiting the maltose hydrolytic activity of Lactobacillus sanfranciscensis associated with Z. mobilis. CO2 production, dough volume increase, pH value, microbial counts, sugars consumption and ethanol production were monitored. Results suggest that glucose addition to the dough lets Z. mobilis efficiently leaven a dough, while glucose released by L. sanfranciscensis is not so well fermented by Z. mobilis, probably due to the strong acidification. Nevertheless, the use of Z. mobilis as a leavening agent could contribute to increasing the variety of baked goods alternative to those leavened by S. cerevisiae.
1. Introduction
In the last decades, the research in human nutrition has aimed both at improving food safety and demonstrating new healthy properties of foods or ingredients. Particularly in the grain cereals area, great attention has been paid to the study of sourdough microbial ecology [1,2] and to the positive effects of lactic acid bacteria (LAB) and yeast fermentation on the technological characteristics of dough. Therefore, several contributions have led to the enhancement of baked products by using a sourdough technology, which also matches consumer’s choices in terms of their preference towards the valorisation of traditional products that can be certified [3]. Organic acid production impacts on sourdough texture and product shelf-life, with acetic acid also displaying anti-ropiness and antifungal activities [4,5]. Acidification also helps to activate endogenous cereal proteases that release peptides and amino acids related to flavour formation [6]. The production of bacteriocins allows microorganisms to control the sourdough ecosystem [7], while the synthesis of homo-polysaccharides delays firmness and staling [6]. Sourdough fermentation can also have positive nutritional implications by biodegrading phytates, thus increasing mineral bioavailability, and by lowering the glycaemic response to the consumption of baked goods [6].
Nevertheless, adverse food reactions, such as baker’s yeast intolerance, have recently been increasing among Western population [8]. Apart from in well-known alcoholic beverages, such as beer, wine and cider, and baked goods, S. cerevisiae is also used in savoury spreads, as a food supplement in ‘multi-vitamin’ preparations and ‘probiotics’ in animal feed [9], and even in vaccine production [10]. It is therefore clear that we are often exposed to yeast parietal components [11].
Several studies report that an adverse response to baker’s yeast occurs in a proportion of patients with Inflammatory Bowel Disease (IBD). In particular, in patients with Crohn’s disease (CD), S. cerevisiae is recognized as an antigen, and anti-S. cerevisiae antibodies (ASCAs), directed against the cell wall mannan (phosphopeptidomannan) of yeast, have been identified as an important serological marker of this pathogenesis. However, the determination of ASCAs is also reliable in other autoimmune disorders besides CD [12]. Environmental factors such as food antigens may play an important role in the pathogenesis of autoimmune disorders [10,13] and obesity [9]. Although there is scarce literature on allergy-hypersensitivity to yeasts, some clinical conditions might benefit from reduced exposure to these microorganisms [14].
Based on these considerations, the study of new microbial resources to be applied in leavened goods may be considered of actual relevance; in this context, the possibility of replacing S. cerevisiae is noteworthy. The use of Zymonomas mobilis as leavening agent can contribute to an increase in the variety of bakery products alternative to those leavened by yeast in order to meet the specific demands of consumers. Z. mobilis can therefore be an interesting candidate to create a new food area of yeast-free baked goods. This bacterium is commonly used in tropical areas as a fermenting agent of plant saps to obtain alcoholic beverages such as pulque [15]. Z. mobilis ferments only glucose, fructose and sucrose, and through alcoholic fermentation it provides an equimolar mixture of ethanol and CO2 that can theoretically leaven a dough [16], just like S. cerevisiae. The narrow range of fermentable substrates is a technological limit of Z. mobilis vs S. cerevisiae that may be overcome by: (1) adding a fermentable sugar to the dough formulation; or (2) exploiting maltose hydrolytic activity of Lactobacillus sanfranciscensis associated with Z. mobilis. This unconventional association has been investigated as a model system (higher cell concentration and leavening temperature, shorter leavening time) in a previous paper [17]. The present research aims to compare Z. mobilis leavening performance when glucose is added to the dough both with its fermentative ability when Z. mobilis is in association with L. sanfranciscensis and in doughs formulated and processed similarly to a type I sourdough.
2. Materials and Methods
2.1. Microorganisms and Maintenance
Z. mobilis subs. mobilis type strain DSM 424 (DSMZ: Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH) and L. sanfranciscensis DSM 20663 were used in this study.
Z. mobilis was maintained in liquid DSM medium, while biomass production was carried out in liquid IC G20 medium (as previously reported) [16]. Both media contain bacto-peptone (Costantino SpA, Turin, Italy) 10 g/L and glucose (Sigma Aldrich, St. Louis, MO, USA) 20 g/L, while they differ for yeast extract (Costantino SpA) 10 g/L present in DSM medium and of casein enzymatic hydrolysate (Costantino SpA) 10 g/L in IC G20. For both media, the pH was set at 6.8, and sterilization occurred at 112 °C for 30 min.
L. sanfranciscensis was maintained and cultivated in MRSm medium as reported elsewhere [17]. Cultures were incubated at 30 °C in stationary conditions for 16–24 h. Stock cultures of both microorganisms were stored at −80 °C in the same media (DSM for Z. mobilis and MRSm for L. sanfranciscensis) added with 20% (v/v) glycerol (VWR International, Leuven, Belgium).
2.2. Biomass Production
Z. mobilis was cultured in 1 L flasks containing 600 mL of liquid IC G20 medium, inoculated with 5% (v/v) of a 9 h pre-culture grown in DSM medium. L. sanfranciscensis was grown in 1 L flasks containing 600 mL of MRSm medium, inoculated with 2% (v/v) of a 24 h pre-grown culture in the same medium. Cultures were incubated at 30 °C in stationary conditions for 16 h for Z. mobilis and 24 h for L. sanfranciscensis.
The determination of the cell biomass was performed by spectrophotometric measurement (OD 600 nm, 6705 UV-Vis Spectrophotometer, Jenway, UK). For each strain, at 16 h in the case of Z. mobilis and 24 h for L. sanfranciscensis, a calibration curve was built (OD 600 vs. CFU (colony-forming unit)/mL) to determine the proper culture volume to add in the dough preparation (cell concentration expressed as Log CFU/g dough).
2.3. Dough Production and Analytical Determinations
Doughs were prepared with 333 g of a commercial type 0 Manitoba wheat flour (Simec SpA, Santa Giusta, Oristano, Italy) and 167 mL of distilled water, with or without addition of 1 or 5% (w/w flour) glucose. Z. mobilis was added alone (7 Log CFU/g dough) or with L. sanfranciscensis (5 Log CFU/g dough) yielding to 100:1 ratio Zymomonas:Lactobacillus cells. Ingredients were mixed in a food mixer (CNUM5ST, Bosch, Stuttgart, Germany) at speed 1 for 6 min. The dough was divided into 3 sections, treated as follows and then incubated at 26 °C:
- -
- 400 g, inserted into a 1 L graduate cylinder to evaluate the dough volume increase up to 24 h of leavening;
- -
- 25 g, inserted into a double chamber flask connected with a graduate burette filled with acidified water, to evaluate the total amount of CO2 produced during leavening [16];
- -
- The remaining sample was left to leaven into a Becker; samples were taken at appropriate intervals to determine dough pH, microbial counts and to carry out HPLC (high performance liquid chromatography) analysis.
Each analysis was performed at 0, 8, 16 and 24 h of leavening time.
2.4. Evaluation of Dough Volume Increase and Total CO2 Production
The increase in the dough volume (mL) was evaluated at appropriate time intervals through the record of the level reached by the dough inside the graduate cylinder. CO2 production (mL) was monitored by measuring the level reached by the liquid present inside the burette connected to the double chamber flask.
2.5. Determination of the Microbial Populations in Doughs
Approximately 10 g of dough sample were decimally diluted in sterile peptone water (10 g/L Bacto-peptone (Costantino SpA), pH 6.8) and homogenized in a Stomacher 400 Circulator (Seward, Worthing, UK) for 5 min at 260 rpm. The appropriate dilutions were plated onto MRSm agar (MRSm broth added with agar 15 g/L) for the determination of L. sanfranciscensis population, as well as onto DSM agar (DSM broth added with agar 15 g/L) for Z. mobilis. Plates were incubated at 30 °C for 3 d in anaerobic conditions. Aerobic bacterial count (ABC) was determined by pour plating in Tryptic Soy Agar (TSA, Scharlab, Barcelona, Spain) after incubation at 30 °C for 48–72 h. The enumeration of yeasts and moulds were carried out in Yeast Glucose Chloramphenicol Agar (YGC-Scharlab, Barcelona, Spain) plates after incubating at 25 °C for 3–5 day.
2.6. HPLC Analyses and pH Monitoring
Maltose and glucose consumption, as well as ethanol production during leavening, were measured through an HPLC system (L 7000, Merck Hitachi, Tokyo, Japan) as reported by Musatti et al. [17]. Briefly, 4 mL of homogenized dough samples were centrifuged (Eppendorf 5804 (Hamburg, Germany), 10,600× g, 10 min) and supernatants were filtered (0.45 µm syringe filter, VWR International, Radnor, PA, USA) before HPLC analysis. Data refer to 1 g dough (mg/g dough).
Dough pH was monitored at different intervals on the integral undiluted dough sample (pH-meter Eutech Instruments pH 510, Toronto, ON, Canada).
2.7. Statistical Analysis
All samples were prepared and analysed at least in triplicate. The effect of two factors, such as % glucose addition or L. sanfranciscensis co-inoculation, on some fermentation parameters were investigated by ANOVA according to the general linear model. Results of microbiological counts were transformed in the respective decimal logarithms to match a normal distribution of values. Data were processed with Statgraphic R Plus 5.1 for Windows (StatPoint, Inc., Herndon, VA, USA). When the effect was significant (p < 0.05), differences between means were separated by LSD test of multiple comparisons.
3. Results and Discussion
3.1. Trials with Glucose Addition into Dough
Dough samples were prepared with or without glucose addition (1% and 5% w/w). When glucose was not added, Z. mobilis fermented only the glucose amount naturally present in the flour (around 2.01 ± 0.59 mg/g dough). However, even if there are some hydrolytic enzymatic activities in the dough due to the presence of endogenous amylases, the low glucose concentration does not allow adequate CO2 production to obtain a suitable dough volume increase by Zymomonas, especially in the first times of incubation.
The need to add a fermentable carbon source to the flour, in order to obtain a leavening of the dough, had already been highlighted in a previous work [16]. Actually, the results confirmed that the addition of glucose increases the CO2 production (p = 0.001), and that in the three tested conditions mean values became statistically different at 16 h leavening time (p = 0.007) (Figure 1). Similarly, the addition of glucose allowed the doubling of the dough volume within the considered incubation time. As expected, the highest CO2 production is related to the highest dough volume increase; in particular, with 5% glucose, the mean value of dough volume reached more than 850 mL (p = 0.019) with respect to an average of 815 or 735 mL with 1% or without glucose addition, respectively. CO2 production was also related to bacterial growth; when no glucose is added, Z. mobilis grew approximately 1.3 Log CFU/g in 24 h, and around 1.6–2 Log CFU/g in the presence of 1% and 5% glucose, respectively. The performances obtained in dough samples with the two glucose concentrations were not significantly different between them, but both were statistically different from those obtained without glucose (p < 0.001).
Results from HPLC analysis confirmed the increase of maltose during leavening time (p < 0.001) due to hydrolytic activity of the flour amylases and Z. mobilis inability to use this sugar (Table 1). At up to 8 h of incubation, the ethanol formation was not statistically different (p = 0.414), even if the three tested conditions had different levels of glucose. Then, glucose was mainly consumed between 8 and 16 h, producing CO2 and ethanol in higher amounts in samples to which 5% glucose was added, as expected.
3.2. Bacterial Association Z. mobilis-L. sanfranciscensis
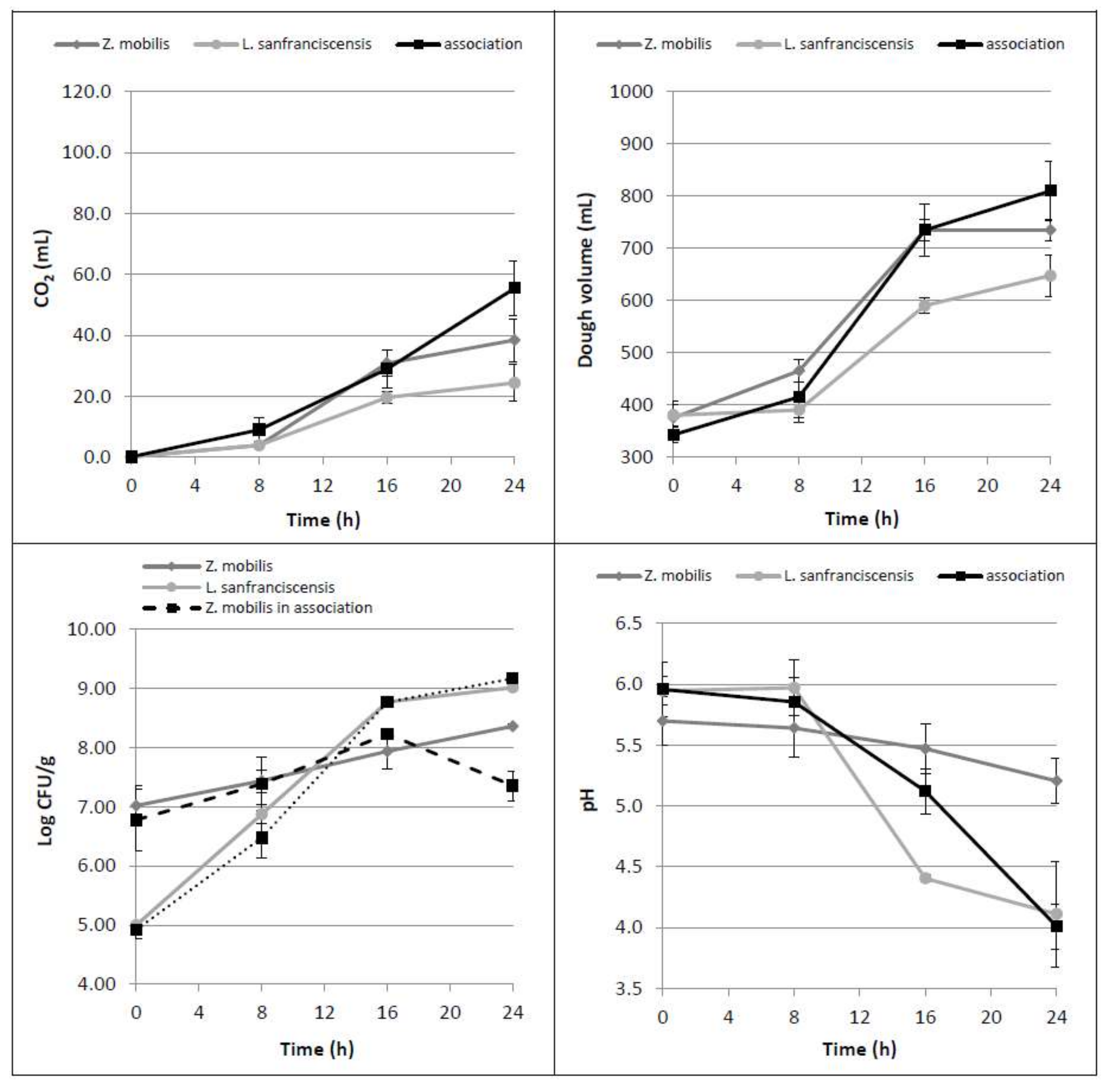
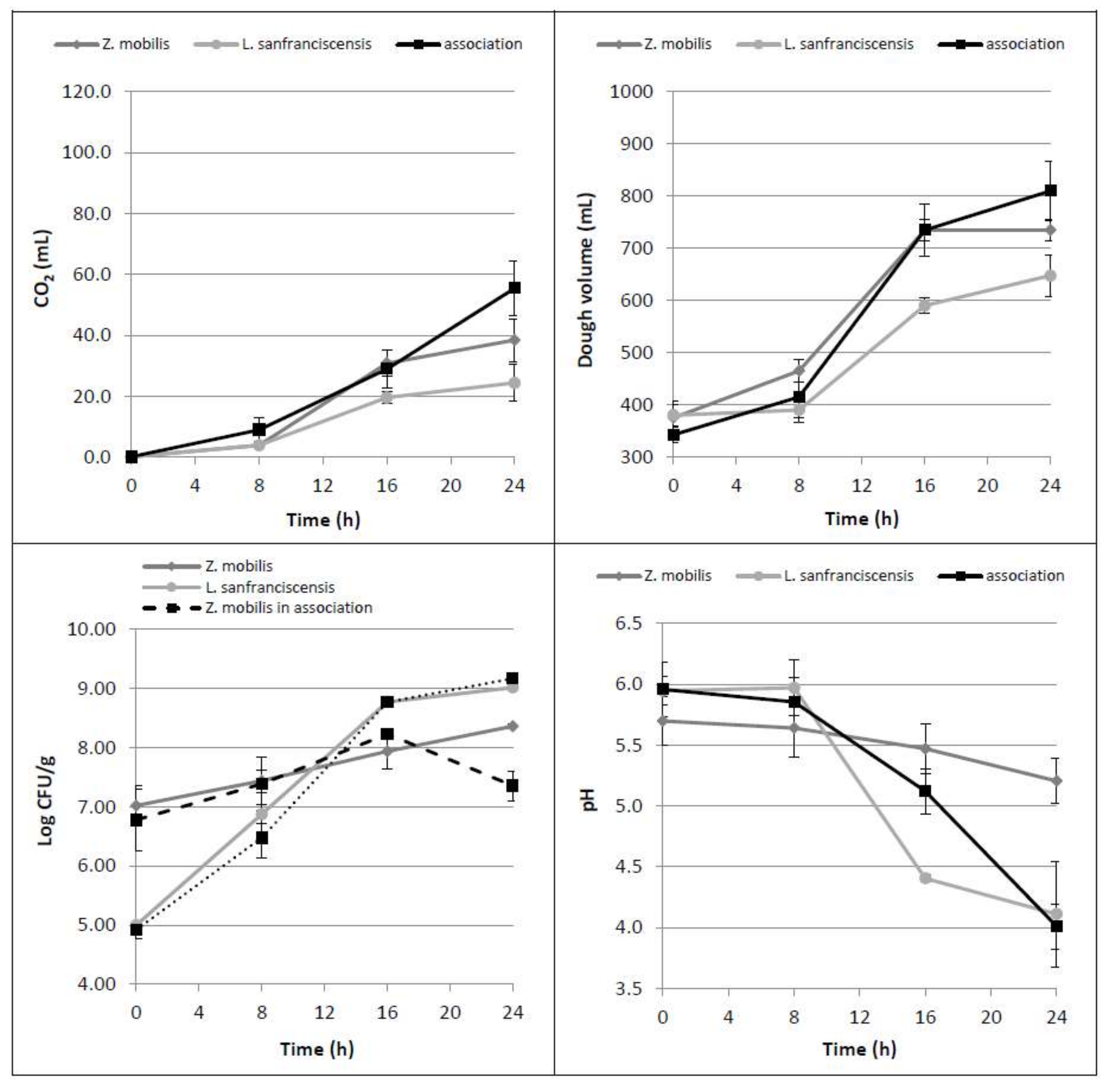
The association of Zymomonas with lactic acid bacteria has already been described in various food products, especially in some fermented drinks [18,19,20,21]. From this perspective, the possibility of obtaining a gradual glucose release in the dough exploiting the maltose hydrolytic activity of Lactobacillus sanfranciscensis was investigated [17].
When L. sanfranciscensis and Z. mobilis were inoculated together, the mean values of CO2 production and dough volume increase did not significantly differ from those obtained with the use of Z. mobilis alone (Figure 2). Dealing with the single leavened samples, dough volumes at 16 and 24 h were found to be statistically different from those observed by using L. sanfranciscensis alone (p = 0.023 and 0.024, respectively). These results indicate that the contribution of the two microorganisms in association is not additive. Respect to the trials performed with glucose addition, in which the dough volumes nearly doubled in the first 8 h, the dough volume increased less than 20% in the case of the microbial association. As regards the trends of acidification and bacterial counts during the incubation time, the obtained data proved to be strongly affected by the presence of L. sanfranciscensis: the pH decreased to values of around 4 and the LAB growth (plus 4 Log CFU/g) was not influenced by the presence of Zymomonas. On the contrary, Z. mobilis, when grown in association with L. sanfranciscensis, statistically reduced (p = 0.002) its cell concentration from 16 h leavening onward. This behavior is probably due to the strong acidification of the medium produced by L. sanfranciscensis, able to affect both Z. mobilis vitality and fermentation ability.
HPLC data confirmed that L. sanfranciscensis consumed maltose (p = 0.012) and released glucose (p = 0.003) in the dough [4], that it is not totally consumed by Z. mobilis (Table 2).
In summary, these results highlight that when inoculated alone, Z. mobilis is able to consume all the glucose present in a dough, while when coupled with L. sanfranciscensis, its fermentative performance decreases. Furthermore, the presence of L. sanfranciscensis did not lead to a significant ethanol yield increase, even if it can consume the available maltose in flour and release glucose.
4. Conclusions
The results obtained demonstrate that Z. mobilis is able to efficiently leaven a dough when glucose is present in the dough formulation. On the other hand, although the metabolic activities of LAB have positive effects on the structural and sensorial properties of the baked product [5,6], the traditional back-slopping sourdough technology [22] cannot be proposed due to the accelerated acidification of the dough impairing the growth of Zymomonas. In fact, preliminary trials have evidenced that, independently of the initial cell ratio between the two bacteria, L. sanfranciscensis always became the prevailing microbial population. This disproportion with Z. mobilis increased with refreshments, thus giving strongly acidified and poorly leavened doughs.
Future trials will be aimed at investigating microbial association with other LAB or the use of dough formulations naturally enriched of sugars fermentable by Z. mobilis. In this context, sucrose can also be considered an interesting alternative to glucose; nevertheless, the strain leavening performance with this carbon source has to be evaluated.
Acknowledgments
This work was founded by Bando Linea R&S per Aggregazioni, Regione Lombardia, Programma Operativo Regionale 2014–2020, Strategia “InnovaLombardia” (D.G.R. No. 2448/2014) Project number 145007.
Author Contributions
A.M. and C.P. conceived and designed the experiments; A.M. and C.M. performed the experiments; R.F. analysed the data; R.F. contributed reagents/materials/analysis tools; M.R. and C.P. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Food Sci. Technol. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Picozzi, C.; D’Anchise, F.; Foschino, R. PCR detection of Lactobacillus sanfranciscensis in sourdough and Panettone baked product. Eur. Food Res. Technol. 2006, 222, 330–335. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [Google Scholar] [CrossRef]
- Arendt, E.K.; Ryan, L.A.M.; Bello, F.D. Impact of sourdough on the texture of bread. Food Microbiol. 2007, 24, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Angioloni, A.; Romani, S.; Gaetano Pinnavaia, G.; Dalla Rosa, M. Characteristics of bread making doughs: Influence of sourdough fermentation on the fundamental rheological properties. Eur. Food Res. Technol. 2006, 222, 54–57. [Google Scholar] [CrossRef]
- Messens, W.; De Vuyst, L. Inhibitory substances produced by Lactobacilli isolated from sourdoughs—A review. Int. J. Food Microbiol. 2002, 72, 31–43. [Google Scholar] [CrossRef]
- Mansueto, P.; Montalto, G.; Pacor, M.L.; Esposito-Pellitteri, M.; Ditta, V.; Lo Bianco, C.; Leto-Barone, S.M.; Di Lorenzo, G. Food allergy in gastroenterologic diseases: Review of literature. World J. Gastroenterol. 2006, 12, 7744–7752. [Google Scholar] [CrossRef] [PubMed]
- Salamati, S.; Martins, C.; Kulseng, B. Baker’s yeast (Saccharomyces cerevisiae) antigen in obese and normal weight subjects. Clin. Obes. 2015, 5, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Perricone, R.; Blank, M.; Perricone, C.; Shoenfeld, Y. Anti-Saccharomyces cerevisiae autoantibodies in autoimmune diseases: From bread baking to autoimmunity. Clin. Rev. Allergy Immunol. 2013, 45, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Sicard, D.; Legras, J.-L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. Comptes Rendus Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Israeli, E.; Grotto, I.; Gilburd, B.; Balicer, R.D.; Goldin, E.; Wiik, A.; Shoenfeld, Y. Anti-Saccharomyces cerevisiae and antineutrophil cytoplasmic antibodies as predictors of inflammatory bowel disease. Gut 2005, 54, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- Muratori, P.; Muratori, P.; Muratori, L.; Guidi, M.; Maccariello, S.; Pappas, G.; Ferrari, R.; Gionchetti, P.; Campieri, M.; Bianchi, F.B. Anti-Saccharomyces cerevisiae antibodies (ASCA) and autoimmune liver diseases. Clin. Exp. Immunol. 2003, 132, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.A.; Tadros, S.; Bansal, A.S. Beer, Cider, and Wine Allergy. Case Rep. Immunol. 2017, 2017, 7958924. [Google Scholar] [CrossRef] [PubMed]
- Sahm, H.; Bringer-Meyer, S.; Sprenger, G.A. Proteobacteria: Alpha and Beta Subclasses. In The Prokaryotes, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: Berlin, Germany, 2006; Volume 5, ISBN 978-0-387-25476-0. [Google Scholar]
- Musatti, A.; Rollini, M.; Sambusiti, C.; Manzoni, M. Zymomonas mobilis: Biomass production and use as a dough leavening agent. Ann. Microbiol. 2015, 65, 1583–1589. [Google Scholar] [CrossRef]
- Musatti, A.; Mapelli, C.; Foschino, R.; Picozzi, C.; Rollini, M. Unconventional bacterial association for dough leavening. Int. J. Food Microbiol. 2016, 237, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Alcántara-Hernández, R.J.; Rodríguez-Álvarez, J.A.; Valenzuela-Encinas, C.; Gutiérrez-Miceli, F.A.; Castañón-González, H.; Marsch, R.; Ayora-Talavera, T.; Dendooven, L. The bacterial community in “taberna” a traditional beverage of Southern Mexico. Lett. Appl. Microbiol. 2010, 51, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.; Giles-Gómez, M.; Hernández, G.; Córdova-Aguilar, M.S.; López-Munguía, A.; Gosset, G.; Bolívar, F. Analysis of bacterial community during the fermentation of pulque, a traditional Mexican alcoholic beverage, using a polyphasic approach. Int. J. Food Microbiol. 2008, 124, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.N.; Ibekwe, V.I.; Anyanwu, B.N. Investigation of some physicochemical and microbial succession parameters of palm wine. J. Food Technol. 2006, 4, 308–312. [Google Scholar]
- Valadez-Blanco, R.; Bravo-Villa, G.; Santos-Sánchez, N.F.; Velasco-Almendarez, S.I.; Montville, T.J. The Artisanal Production of Pulque, a Traditional Beverage of the Mexican Highlands. Probiotics Antimicrob. Proteins 2012, 4, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, S.A.; Hertel, C. Impact of ecological factors on the stability of microbial associations in sourdough fermentation. Food Microbiol. 2011, 28, 583–589. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Time course of CO2 production (mL), dough volume increase (mL), microbial growth of Z. mobilis (Log CFU/g) and dough pH in the three tested conditions (0%, 1%, 5% w/w glucose).
Figure 1.
Time course of CO2 production (mL), dough volume increase (mL), microbial growth of Z. mobilis (Log CFU/g) and dough pH in the three tested conditions (0%, 1%, 5% w/w glucose).
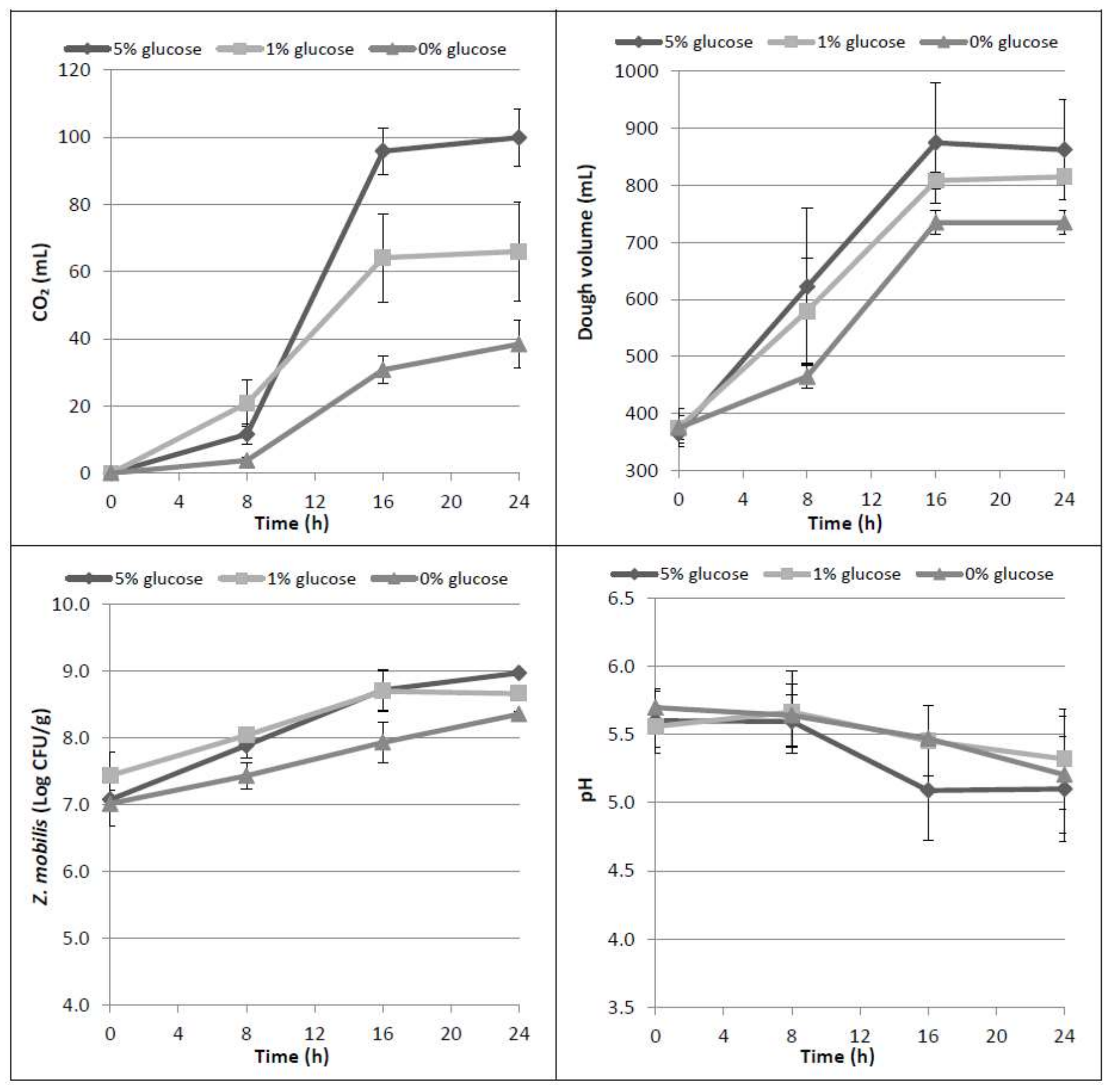
Figure 2.
Time course of CO2 production (mL), dough volume increase (mL), microbial growth (Log CFU/g) as well as dough pH, in doughs leavened by Z. mobilis and L. sanfranciscensis alone or by their association.
Figure 2.
Time course of CO2 production (mL), dough volume increase (mL), microbial growth (Log CFU/g) as well as dough pH, in doughs leavened by Z. mobilis and L. sanfranciscensis alone or by their association.

{kind=link}
{kind=link}
Table 1.
Maltose, glucose and ethanol concentrations (expressed in terms of mg/g dough, mean and standard deviation (St. dev.)) present at 0, 8, 16 and 24 h in doughs leavened by Z. mobilis with 0%, 1%, 5% (w/w) of glucose added respect to the flour.
Table 1.
Maltose, glucose and ethanol concentrations (expressed in terms of mg/g dough, mean and standard deviation (St. dev.)) present at 0, 8, 16 and 24 h in doughs leavened by Z. mobilis with 0%, 1%, 5% (w/w) of glucose added respect to the flour.
| Glucose (% w/w flour) | Time (h) | Maltose (mg/g) | Glucose (mg/g) | Ethanol (mg/g) | |||
|---|---|---|---|---|---|---|---|
| Mean | St. dev. | Mean | St. dev. | Mean | St. dev. | ||
| 0 | 0 | 10.50 | 0.96 | 1.96 | 0.06 | 0.00 | 0.00 |
| 8 | 14.59 | 1.67 | 1.12 | 0.13 | 1.14 | 0.13 | |
| 16 | 17.71 | 3.16 | 0.23 | 0.33 | 2.79 | 0.49 | |
| 24 | 15.69 | 2.65 | 0.00 | 0.00 | 3.38 | 0.33 | |
| 1 | 0 | 10.55 | 1.11 | 8.72 | 0.41 | 0.00 | 0.00 |
| 8 | 15.48 | 5.22 | 3.42 | 0.62 | 1.84 | 0.41 | |
| 16 | 19.00 | 1.59 | 1.23 | 0.54 | 4.03 | 1.02 | |
| 24 | 21.43 | 4.49 | 0.92 | 0.44 | 3.96 | 0.77 | |
| 5 | 0 | 8.33 | 0.95 | 35.72 | 2.95 | 0.00 | 0.00 |
| 8 | 15.72 | 3.29 | 34.56 | 1.62 | 0.72 | 1.01 | |
| 16 | 18.57 | 2.84 | 2.66 | 1.48 | 9.73 | 0.13 | |
| 24 | 20.80 | 2.92 | 1.46 | 0.40 | 13.65 | 2.46 | |
Table 2.
Maltose, glucose and ethanol concentrations (expressed in terms of mg/g dough, mean and standard deviation) present at 0, 8, 16 and 24 h in doughs leavened by Z. mobilis, L. sanfranciscensis and their association.
Table 2.
Maltose, glucose and ethanol concentrations (expressed in terms of mg/g dough, mean and standard deviation) present at 0, 8, 16 and 24 h in doughs leavened by Z. mobilis, L. sanfranciscensis and their association.
| Microorganism | Time(h) | Maltose (mg/g) | Glucose (mg/g) | Ethanol (mg/g) | |||
|---|---|---|---|---|---|---|---|
| Mean | St. dev. | Mean | St. dev. | Mean | St. dev. | ||
| Lactobacillus sanfranciscensis (5 Log CFU/g) | 0 | 11.33 | 1.46 | 2.08 | 1.01 | 0.00 | 0.00 |
| 8 | 16.68 | 0.66 | 3.20 | 0.82 | 0.00 | 0.00 | |
| 16 | 9.43 | 0.60 | 4.01 | 0.28 | 0.00 | 0.00 | |
| 24 | 10.80 | 0.80 | 4.54 | 0.52 | 0.00 | 0.00 | |
| Zymomonas mobilis (7 Log CFU/g) | 0 | 10.50 | 0.96 | 1.96 | 0.06 | 0.00 | 0.00 |
| 8 | 14.59 | 1.67 | 1.12 | 0.13 | 1.14 | 0.13 | |
| 16 | 17.71 | 3.16 | 0.23 | 0.33 | 2.79 | 0.49 | |
| 24 | 15.69 | 2.65 | 0.00 | 0.00 | 3.38 | 0.33 | |
| L. sanfranciscensis coupled with Z. mobilis (5–7 Log CFU/g) | 0 | 8.96 | 1.82 | 1.04 | 0.56 | 0.00 | 0.00 |
| 8 | 14.21 | 2.70 | 0.92 | 0.31 | 0.75 | 0.28 | |
| 16 | 13.58 | 0.94 | 0.56 | 0.79 | 3.73 | 0.70 | |
| 24 | 12.33 | 2.76 | 0.82 | 0.35 | 4.91 | 1.23 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Musatti, A.; Mapelli, C.; Rollini, M.; Foschino, R.; Picozzi, C. Can Zymomonas mobilis Substitute Saccharomyces cerevisiae in Cereal Dough Leavening? Foods 2018, 7, 61. https://doi.org/10.3390/foods7040061
AMA Style
Musatti A, Mapelli C, Rollini M, Foschino R, Picozzi C. Can Zymomonas mobilis Substitute Saccharomyces cerevisiae in Cereal Dough Leavening? Foods. 2018; 7(4):61. https://doi.org/10.3390/foods7040061
Chicago/Turabian StyleMusatti, Alida, Chiara Mapelli, Manuela Rollini, Roberto Foschino, and Claudia Picozzi. 2018. "Can Zymomonas mobilis Substitute Saccharomyces cerevisiae in Cereal Dough Leavening?" Foods 7, no. 4: 61. https://doi.org/10.3390/foods7040061
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.