4. Discussion
The most important meat quality indicator is the pH value of meat [
39]. Post slaughter, anaerobic metabolic decomposition of the glycogen in the muscles causes the production of lactic acid and a subsequent reduction in pH. This may lead to denaturization of muscle proteins, which ultimately results in meat with poor water-holding capacity and (in extreme cases) pale, soft and exudative (PSE) meat [
40]. Regardless of diet and muscle, there was a significant decrease in pH on d 1 and d 7 postmortem. This finding was expected, as the only source of energy for muscle following the exsanguination of an animal is anaerobic glycolysis, which demands the conversion of glycogen to lactic acid in the postmortem muscle [
41,
42]. Glycogen is the substrate for energy production. During the first day postmortem, glycogen is converted into lactic acid and energy. The role of glycogen in skeletal muscle is thought to be influenced less by nutrition and more by the effects of stress or the energy demands of muscle [
43]. The formation of lactate causes a reduction in postmortem pH [
44], therefore, the decrease in pH on day 1 indicated the accumulation of lactic acid. Similar to the present study, several other researchers have reported similar pH values with supplementation of bypass fat and oils in sheep [
45,
46,
47,
48] and beef cattle [
49]. In the present study, operations such as loading, transportation and handling at the slaughterhouse were carried out appropriately, without having a negative impact on animals, therefore, the pH values remained within the normal range for meat [
41,
50].
The water-holding capacity (WHC) is the capability of meat to retain its inherent water during storage and processing [
41], which is an important factor that influences the quantity and yield of the meat [
51,
52] and is considered one of the important economic criteria for the meat processing industry and consumers [
53]. Most of the water is held in the interfilament spaces within the myofilament lattice. The pH, sarcomere length, ionic strength, osmotic pressure and pre- or post-rigor status of the muscle are the factors which determine the volume of interfilament spaces. This, in turn, controls the amount of water present in the meat [
54]. The presence of collagenase enzymes that fragment the connective tissues and myofibrillar proteins are responsible for improving the water-holding capacity of the meat [
55].
The drip loss and cooking loss of the LD muscles in sheep were not influenced by the diets in the present study. The findings are supported by Bhatt et al. [
56] who found similar water-holding capacities in lambs fed with RBF. Sutter et al. [
45] also found similar results for WHC and cooking loss in lambs with supplementation of RBF, coconut oil and oilseeds. Similarly, no differences in WHC were found with or without RBF supplementation in cattle [
57]. Similar findings were reported by several studies [
46,
48,
58]. In contrast to the present study, Oliveira et al. [
49] found a significant difference in the water-holding capacity, as the WHC was higher in linseed oil supplementation in comparison to RBF in cattle. They reported that there was no obvious explanation for this, indicating that it required further investigation. The pH of the meat can also affect other meat quality traits, including thawing loss, water-holding capacity, cooking loss and shear force [
48]. Hence, similar pH in the present study yielded similar water-holding capacity and cooking loss. The aging time increased drip loss and cooking loss values in the current study, irrespective of the treatments.
Cooking loss is an accumulation of liquid and soluble matters lost from meat during cooking [
41] and is important, since the remaining water in the cooked product is the main contributor to the sensation of juiciness [
50]. Similar to the present study, there were no significant differences found for cooking loss with protected and unprotected lipid supplementation in cattle [
49,
59,
60,
61]. On the contrary, there was an increase in cooking loss found with RBF supplementation in lambs [
56] and several other studies reported significantly different cooking loss with RBF supplementation [
47,
57,
62,
63].
Tenderness is among the most important attributes of meat quality as it affects the eating satisfaction of a consumer [
50]. No significant effect of diet was observed for the shear force at day 0 and day 7 but the diet influenced shear force on day 1 postmortem in the present study. This difference on day 1 could possibly be due to differences in pH on day 1, as meat pH can affect shear force values [
48]. Similar to the present study, Bhatt et al. [
63] observed no significant effect of RBF supplementation on the shear force in the meat of lambs. The findings of the present study are also supported by several other studies who reported similar shear force values with protected fat supplementation [
45,
46,
57,
58,
59,
64]. In contrast to the present study, significant differences were observed in shear force values in cattle [
61]. It was reported that sheep meat with shear force values lower than 2.27 kg/cm
2 was classified as soft; meat with values between 2.28 and 3.63 kg/cm
2 was classified as intermediately tender [
48]. However, in the present study, the shear force values were below 2, indicating tender meat. Busboom et al. [
65] reported that sheep meat fat became firmer as the levels of capric, myristic, palmitic and stearic acids increased in the diet. This could be the possible reason for tender meat in this study as our diets contained high levels of palmitic and stearic acids. The postmortem aging process significantly influenced shear force values. The reduction in shear force as the aging day progressed could be attributed to the weakening of myofibrillar structures by endogenous muscle proteinases [
41]. This finding is consistent with the study conducted by Andrade et al. [
57], which found a decrease as the aging day progressed with RBF supplementation when compared to the diet without RBF. The decrease in shear force during postmortem aging was also observed in mutton [
66] and chevon [
67]. The shear force values in the
longissimus dorsi muscle decreased, ultimately resulting in improved tenderness with postmortem aging day from 0 to 7 in the present study.
Color is one of the most important meat quality attributes since it is the first characteristic evaluated by consumers and is an indicator of freshness. Therefore, it directly affects the final purchasing choice of the consumer [
41,
42]. Meat color is influenced by the animal’s age, weight, exercise, and nutrition as well as the pH of the meat. The color is most affected by the amount and chemical state of the principal pigment, myoglobin; the higher the concentration, the darker the meat [
64]. The lightness (
L*) and yellowness (
b*) of
longissimus dorsi muscle was not influenced by diet on postmortem aging day 0 and day 7. These were influenced, however (
p < 0.05), on day 1, whereas, the redness (
a*) was not influenced by the diet. This significant difference observed on postmortem aging day 1 was possibly due to the significant effects of diet seen in pH on day 1. The same effect was also observed in shear force values. The findings are in corroboration with Andrade et al. [
57] who found a significant difference in meat lightness (
L*) and redness (
a*) in
longissimus lumborum muscle with RBF supplementation in cattle. On the other hand, no significant differences were found in lightness (
L*), redness (
a*) or yellowness (
b*) in loin muscle [
49]. The lightness (
L*) values in the present study are similar to the values reported by Awawdeh et al. [
46] for lambs, while the redness (
a*) values in the present study were less than those observed by them. Thus, the slightly darker color of the meat in the present study was due to the age of animal (mature animals were slaughtered for this work).
Ruminant products are typically high in saturated fatty acids (SFA), followed by monounsaturated fatty acids (MUFA) and polyunsaturated fatty acids (PUFA). Red meats from ruminants are higher in the ratio of saturated fatty acids (SFA) than unsaturated fatty acids (UFA) compared to that of meat products from monogastric animals and fishes [
68,
69]. Lipid supplementation is the main method used to modify the fatty acid profile of red meat [
70,
71,
72]. This was clearly demonstrated in the present study, in which significant improvements in desirable fatty acids were observed in tissues of Dorper sheep with supplementation of prilled fat with lecithin.
The most abundant (37% of total fatty acids) fatty acid found in
longissimus dorsi (LD) muscle in sheep was oleic acid (C18:1n-9), irrespective of the dietary treatment, and its concentration was influenced by the diet. The findings are in agreement with the results reported by Andrade et al. [
57] who found C18:1n-9 in the most abundant concentration in rearing and fattening periods (39.75 and 41.05%) regardless of treatment with supplementation of RBF in cattle. Similarly, Oliveira et al. [
49] reported C18:1n-9 ranging 37% to 40% in
longissimus thoracis muscle with two different oils and RBF supplementation. Likewise, several other studies also found C18:1n-9 in the most abundant quantity in muscles with RBF and dietary oil supplementation [
59,
70,
73,
74]. The highest concentration of C18:1n-9 in the present study was recorded in the diet containing PFL, which could be attributed to the presence of lecithin. This finding is supported by the study conducted by Li et al. [
75] who found that supplementation of soy lecithin increased the concentration of C18:1n-9 in the
longissimus dorsi muscle of cattle compared to the control diet; however, the difference was nonsignificant.
There was no significant difference in the concentration of palmitic acid (C16:0) in muscle and liver with RBF supplementation. Similar observations were reported in the liver and foreshank muscles of sheep [
72]. The concentrations of stearic acid (C18:0) in LD muscle and liver with RBF supplementation did not significantly differ, which was in agreement with the study reported by Warner et al. [
4] who observed similar concentration of C18:0 with RBF supplementation in LD muscle in cattle. However, the concentration of C18:0 could not have any negative effect on the meat as, when consumed by humans, C18:0 is transformed into C18:1n-9, a fatty acid that does not carry any cardiovascular risks [
76]. The findings of the present study contradict several studies who found a reduction in C18:0 with dietary oil in ruminants [
77,
78].
The concentration of lauric acid (C12:0) was not affected by dietary treatments in LD muscle and liver. Similar findings were observed in different tissues in cattle [
78]. The concentrations of myristic acid (C14:0) and pentadecanoic acid (C15:0) in LD muscle differed significantly. This finding is in agreement with those of Gilbert et al. [
59] who also reported a significant reduction with RBF supplementation, as compared to other diets in steers. Lima et al. [
60] also reported similar concentrations of C12:0, C14:0 C15:0 and C17:0 with RBF supplementation in cattle.
The concentrations of linoleic acid (C18:2n-6) and linolenic acid (C18:2n-3) increased significantly in LD muscle with RBF supplementation as compared to the diet without RBF. There was no significant increase observed in the liver. Conversely, a higher concentration of C18:2n-6 was observed with RBF supplementation in cattle [
49]. Our findings are in concordance with the findings of Gómez et al. [
79] who found a nonsignificant effect for C18:2n-6 and a significant effect for C18:3n-3; those effects were similar in liver and LD muscles, respectively, observed in the present study. Contrary to the present study, the study conducted by Gilbert et al. [
59] reported significantly higher concentration of C18-2n-6 in the control group as compared to the animals fed RBF. Additionally, the present study is supported by another study which evaluated different RBF in the muscles of cattle and found that two of the RBF increased the concentration of C18:2n-6 and C18:3n-3 compared to the control group [
80]. Thus, the increase in the concentrations of C18:2n-6 and C18:3n-3 in the diet with RBF (PF, PFL and CaS) (as compared to the diet without RBF (CON)) in the
longissimus dorsi muscle and liver could be the effect of rumen bypass fat. Inertness of RBF allowed them to pass intact through the rumen, allowing its absorption in the small intestine and subsequent deposition in the tissue [
49].
No significant differences were observed in the levels of CLA
Cis-9
Trans-11 or CLA
Trans-10
Cis-12 in LD muscle of Dorper sheep fed RBF. The concentration of CLA
Trans-10
Cis-12 in the liver increased significantly with RBF supplementation compared to the diet without RBF. This increased level of CLA is supported by Oliveira et al. [
49] who found a significant increase in the concentrations of CLA with oil supplementation and attributed this increase to the presence of larger levels of C18:2n-6 in the diets which is a precursor of CLA. In the present study, the diet with calcium soap (CaS) was high in C18:2n-6—this could be the reason for significantly increased CLA
Trans-10
Cis-12 concentration in the liver in the group fed with calcium soap (CaS). Moreover, our results showed no significant increase in the concentration of CLA
Cis-9
Trans-11, probably because of the fatty acid composition of diet. The RBF supplemented diets (PF and PFL) were not rich in C18:2n-6 compared to the diet without RBF (CON) and CaS, and thus could not cause a significant increase in the concentration of CLA.
The n-6:n-3 ratio in the tissues examined (including the muscle and the liver) ranged from 2.32 to 3.56 with RBF supplementation in the present study. According to the Department of Health and Social Security DHSS, (1994), n-6:n-3 ratio values lower than 4.0 are desirable for the prevention of cardiovascular diseases. The values observed in the present study were even lower than these desirable values, denoting that the meat from the studied animals may be in a potentially healthier category.
In general, the supplementation of RBF significantly decreased concentrations of SFA and increased the concentrations of UFA, MUFA and PUFA, compared to the diet without RBF. These findings are supported by several studies which reported a decrease in SFA and an increase in UFA as the result of oil supplementation (whether protected or unprotected) [
49,
70,
71,
77,
78]. Furthermore, RBF supplementation increased the concentrations of C18:1n-9, CLA
Trans-10
Cis-12, C18-2n-6 and C18-3n-3 and reduced the n-6:n-3 ratio, which is beneficial to human health. These fatty acids reduce the levels of LDL cholesterol in the blood, freeing Dorper sheep meat fed with RBF from negative effects.
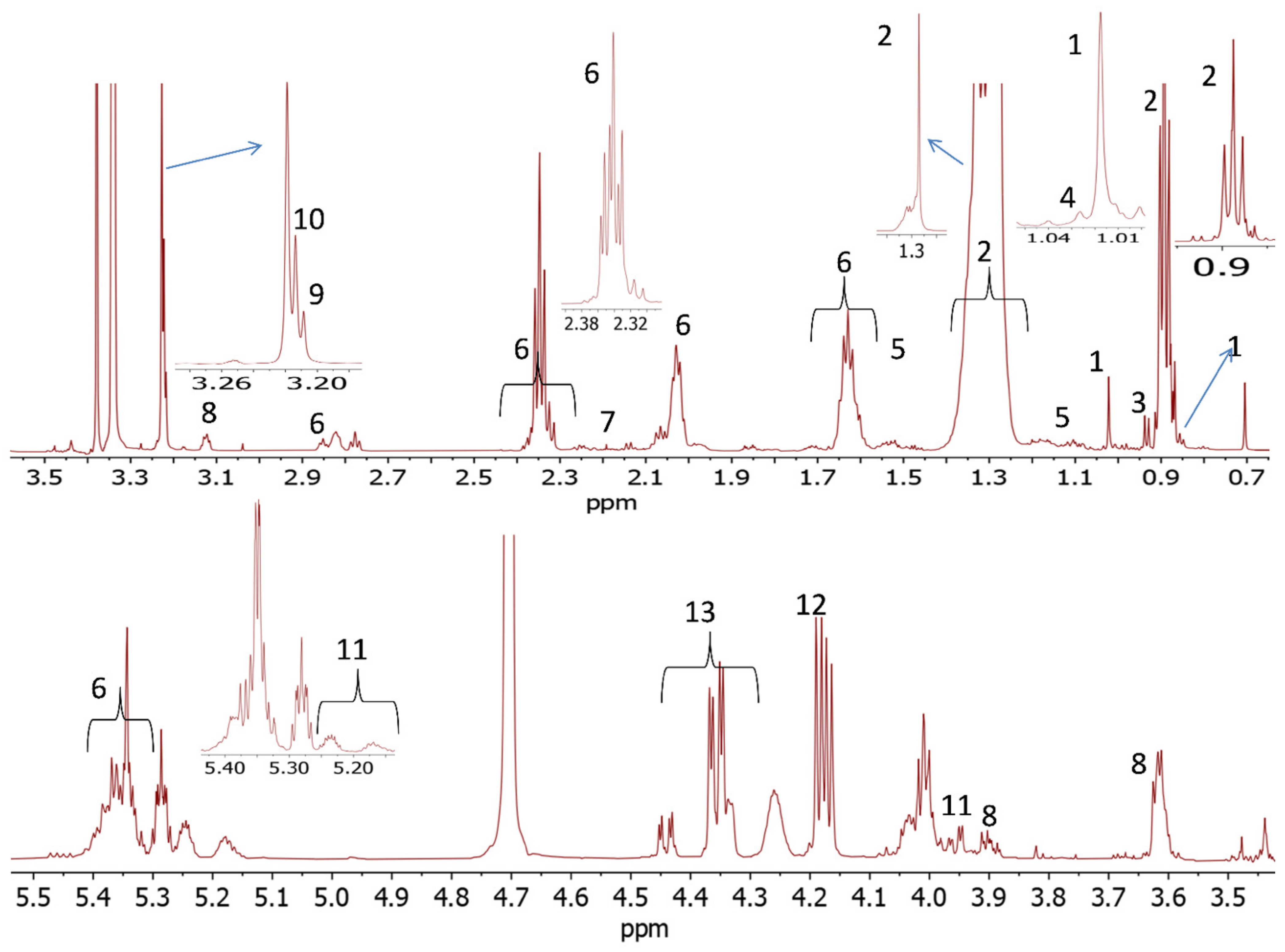
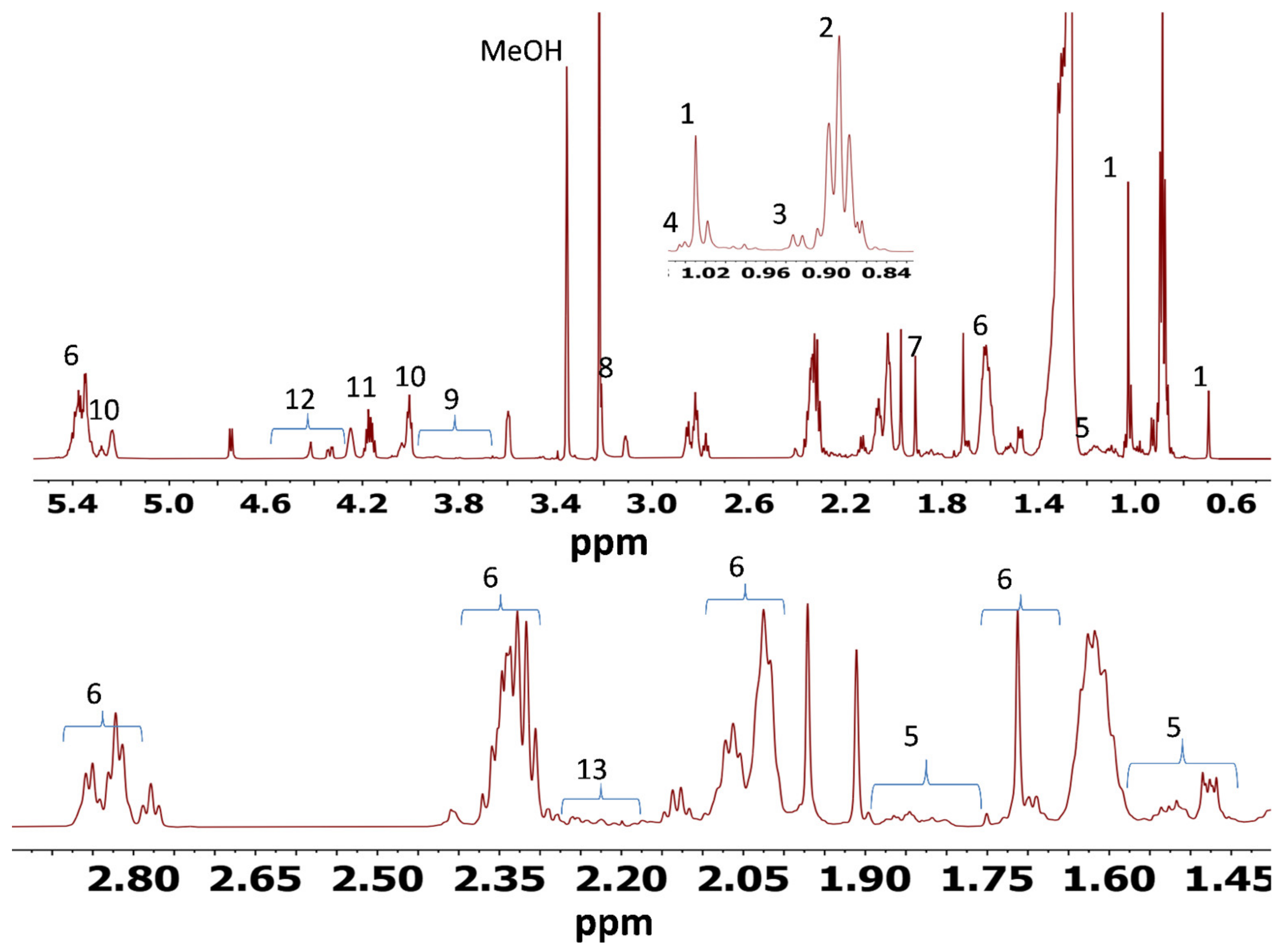
The NMR-based metabolomics proved to be an effective technique for examination of the responses to dietary treatments in muscle and liver tissues of sheep. A simple 1D spectrum enabled preliminary pattern recognition analysis and identification of metabolites. In the present study, NMR also met other significant benefits for metabolomics studies, e.g., simple sample preparation, quick spectra acquisition and the nondestructive nature of the method, allowing samples to be used with other techniques, if necessary.
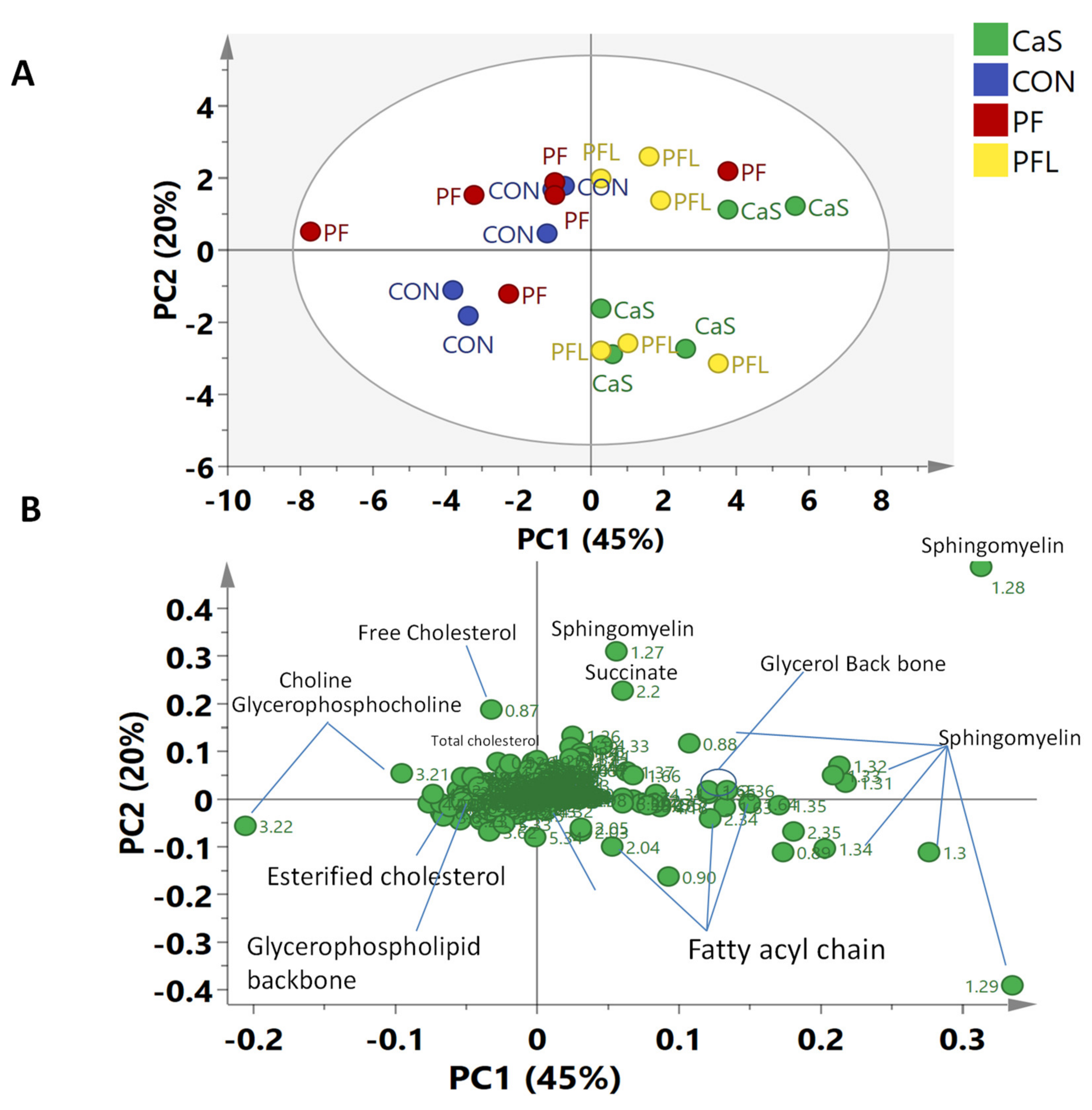
There have been only a few reported studies on the effects of dietary treatments on muscle and liver tissue metabolites. Palma et al. [
36] conducted similar studies on the effects of nutritional treatments on the muscle and liver metabolites in three different breeds of sheep (Merino, Damara and Dorper sheep). On the basis of multivariate data analysis (MvDA), they did not find a clear separation between treatment groups for either muscles or liver; however, they observed some differences in muscle samples among breeds. Furthermore, they were unable to find differences—especially in the Dorper breed—between the two nutritional treatment groups with good quality parameters in liver and muscle tissues. On the other hand, a clear separation was observed in the present study in the metabolomes of Dorper sheep muscle tissues supplemented with different rumen bypass fats.
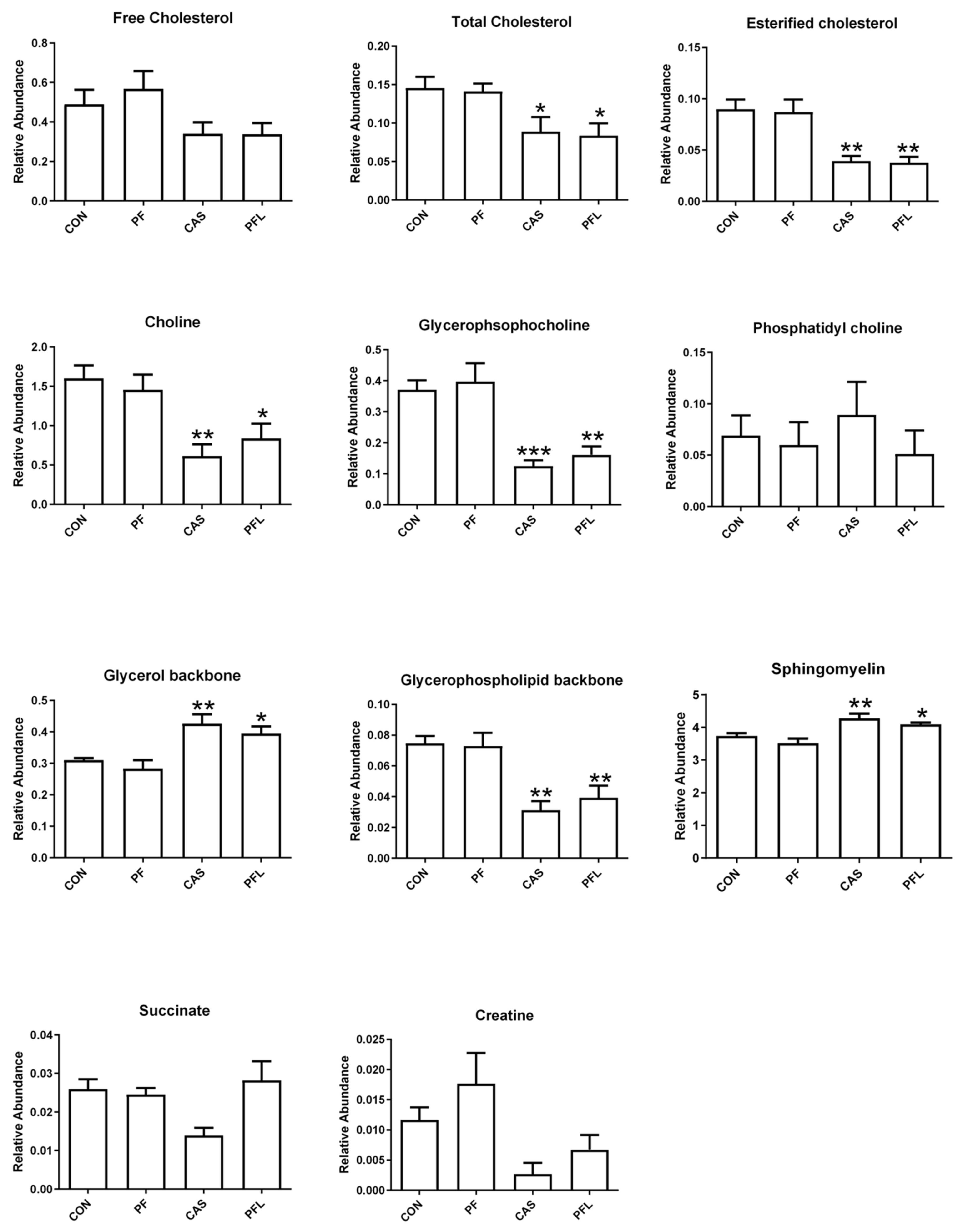
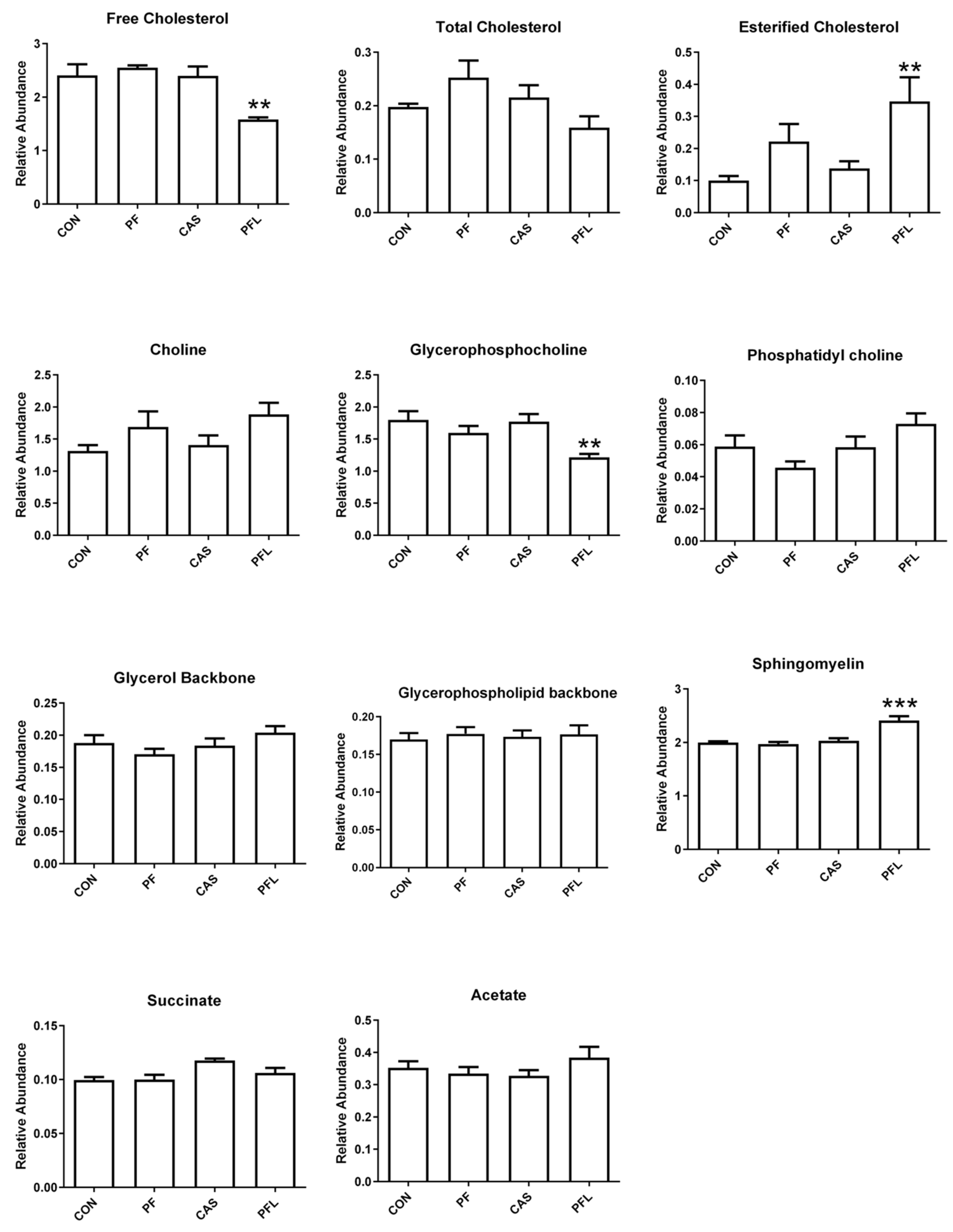
The key discriminating metabolites identified in muscle tissues were choline, creatine, esterified cholesterol, fatty acyl chain, free cholesterol, glycerol backbone, glycerophospholipid backbone, glycerophosphocholine, multiple cholesterol protons, phosphatidyl choline, sphingomyelin, succinate and total cholesterol. The PFL and CaS had higher levels of the glycerol, sphingomyelin, succinate, and fatty acyl chain, whereas CON and PF were characterized by high contents of free cholesterol, esterified cholesterol, total cholesterol, choline, creatine, glycerophosphocholine and glycerophospholipid backbone.
Creatine has been known to be involved in energy production in muscle tissues. Increased levels of creatine in the present study in PFL and CaS groups were reported. As reported by Jeong et al. [
81] increased creatine level in muscles may delay postmortem lactate formation and decreases in pH, potentially improving the water-holding capacity. Similarly, increased levels of creatine in muscles of feed-restricted sheep groups was reported to generate energy by promoting gluconeogenesis and glycogenolysis [
36]. Likewise, higher concentrations of glycerophosphocholine in feed-restricted sheep were also reported to be involved in some regulatory processes and to play an important role in muscle control (as the metabolite is the storage form of choline and the source of a methyl group) [
82]. Similar to these results, an increasing trend was observed in the levels of creatine and glycerophosphocholine in CON and PF groups in this experiment.
The PFL and CaS diets significantly decreased the concentrations of free cholesterol, total cholesterol and esterified cholesterol in muscle tissues compared with CON and PF diets. The research showed that a diet consisting mainly of red meat is rich in total fats, saturated fats and cholesterol and leads to concerns regarding the risk of coronary heart disease and atherosclerosis linked with high dietary levels of lipid and cholesterol [
83]. Therefore, a reduction in cholesterol levels in PFL and CaS diets was supported by the reduction in UFA in these diets in the present study. Cholesterol comes from food, then enters the digestive tract and the small intestine to be absorbed by enterocytes of the small intestine mucosa. Next, they undergo esterification into cholesterol esters. After that, lipoprotein cholesterol esters form chylomicrons, then get into the flow of lymph and end up in the bloodstream [
84]. In the present study, the inclusion of bypass fat decreased the meat cholesterol and fat content of sheep meat, when compared to the control group. The amount of lipid in meat can vary extensively, depending on many factors, e.g. the animal species, the diet eaten by the animal, the degree of trimming of fat from the muscle during the various handling phases, the particular cut of meat and the cooking or processing techniques used [
85]. In the present study, low levels of cholesterol in PFL and CaS diets suggested that the meat from the Dorper sheep fed with diets with PFL and CaS had lower cholesterol and were ultimately safer and healthier for consumption by humans.
Contrary to muscle tissues, no discriminating effect of diet on the metabolome of liver tissues was found. Dorper sheep have been reported to resist dehydration and quickly recover water weight loss as soon as water is available after dehydration [
86]. Likewise, different feeding systems (including extensive, semiextensive and intensive systems) were unable to affect the growth performance of Dorper sheep and also affirmed to have very little effect on meat quality parameters [
87]. Compared to Merino and Damara sheep, Dorper sheep showed few changes in muscle and liver metabolomes [
36].


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}