
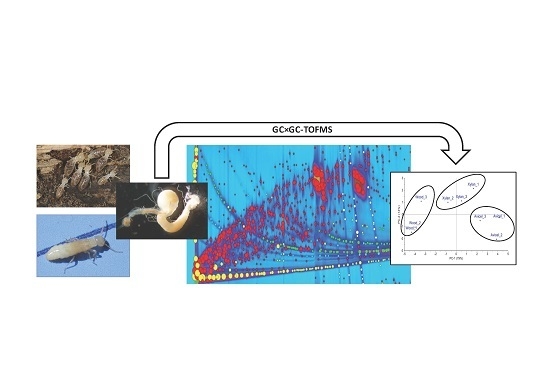
GC×GC-TOFMS for the Analysis of Metabolites Produced by Termites (Reticulitermes flavipes) Bred on Different Carbon Sources

 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Termites, Culture Conditions and Artificial Diets
2.2. Chemicals
2.3. Sample Preparation and Derivatization
2.4. GC×GC-TOFMS Analysis
2.5. Data Processing and Statistical Analyses
3. Results and Discussion
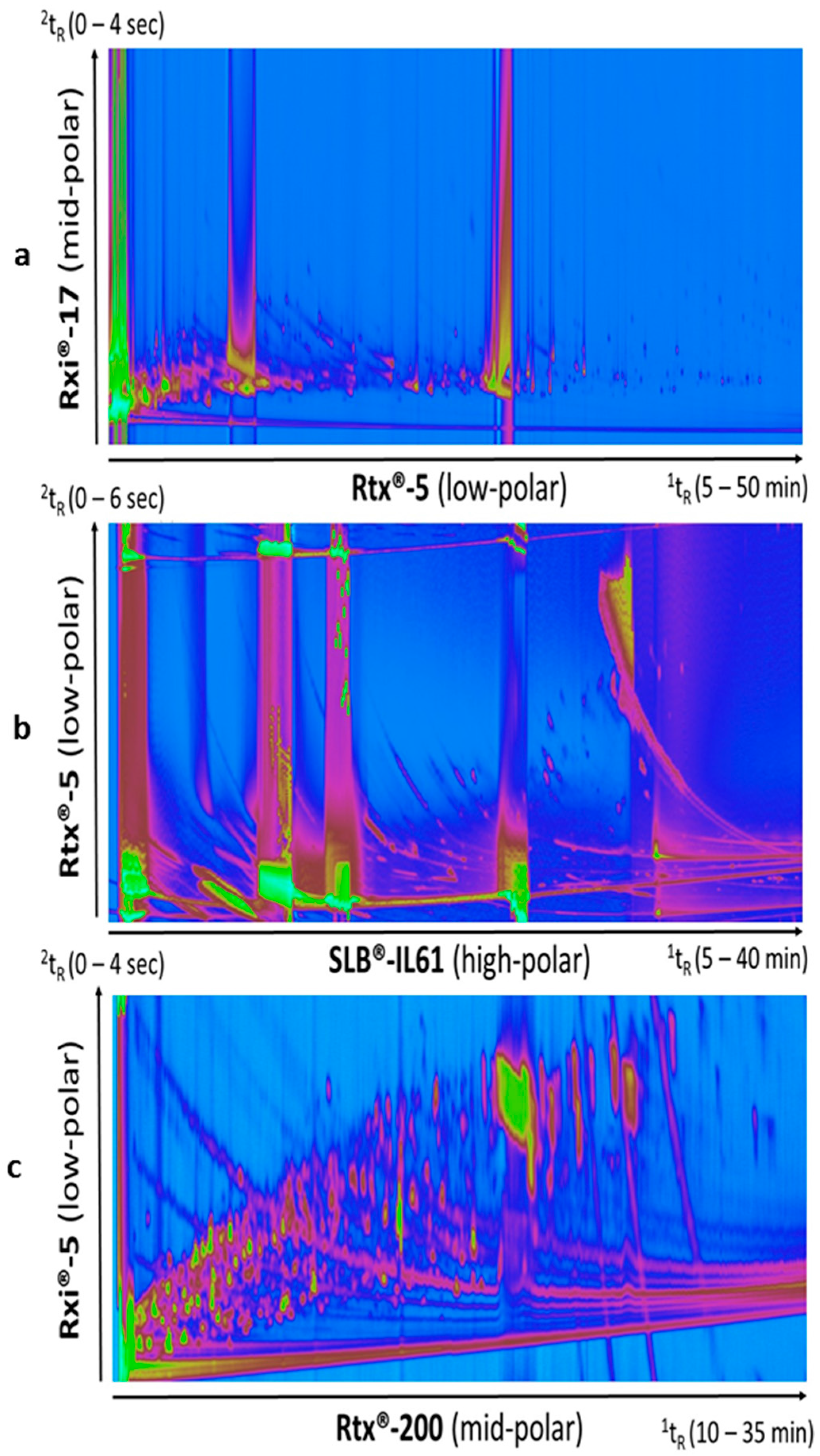
3.1. Test of Different Column Sets
3.2. Filtering Out Column Bleed and Noise in Derivatized Samples
3.3. Principal Component Analysis of Metabolite Profiles of Termites Cultured on Different Carbon Sources
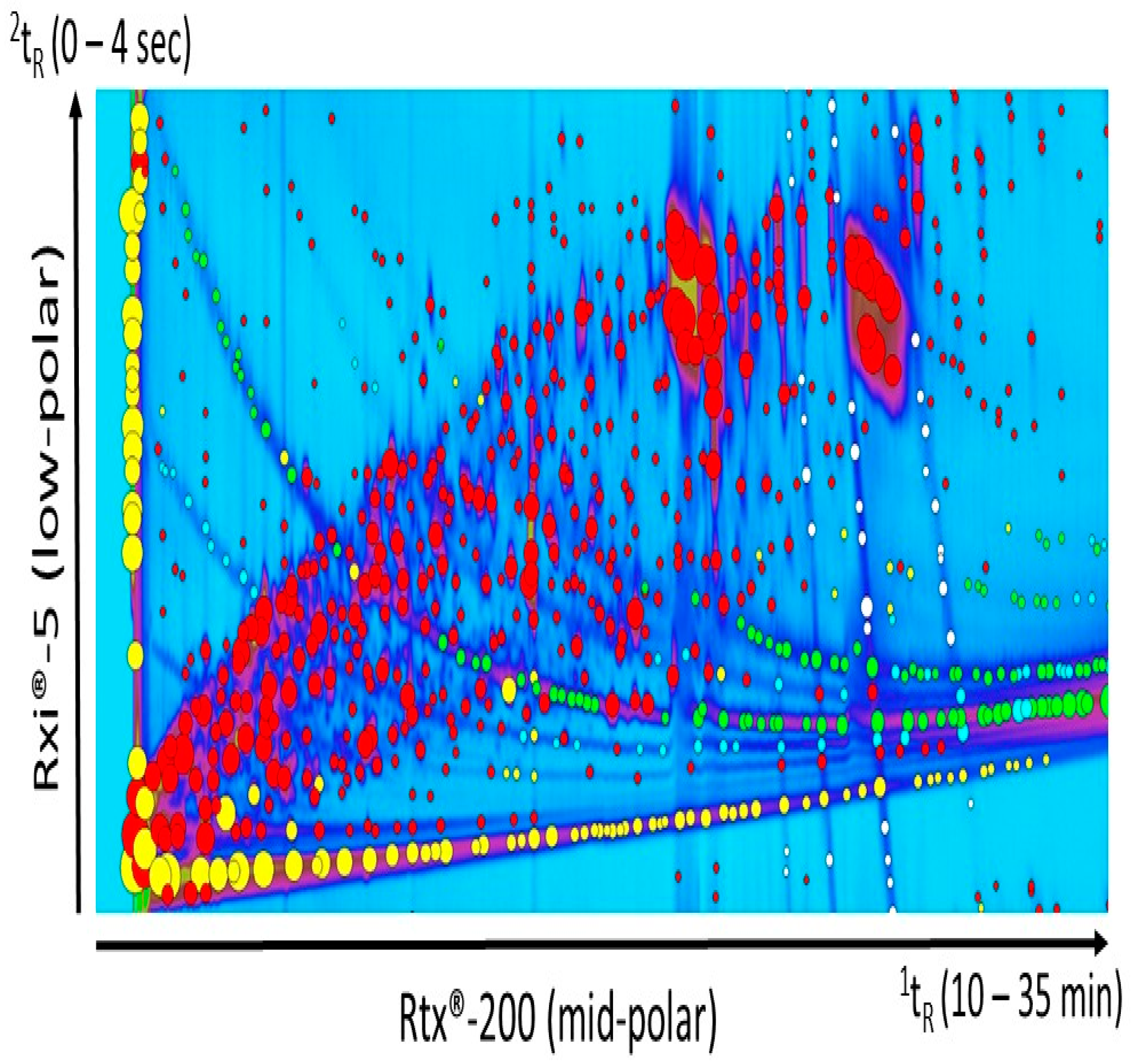
3.4. Identification of Diet-Specific Compounds
3.4.1. Wood Diet
3.4.2. Xylan Diet
3.4.3. Cellulose Diet
3.5. Influence of Pentoses Versus Hexoses on Metabolite Profile
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ohkuma, M. Termite symbiotic systems: Efficient bio-recycling of lignocellulose. Appl. Microbiol. Biotechnol. 2003, 61, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chandel, A.K.; da Silva, S.S. Sustainable degradation of lignocellulosic biomass—Techniques, applications and commercialization. InTech 2013. [Google Scholar] [CrossRef]
- Breznak, J.A.; Brune, A. Role of microorganisms in the digestion of lignocellulose by termites. Ann. Rev. Entomol. 1994, 39, 453–487. [Google Scholar] [CrossRef]
- Kudo, T. Termite-microbe symbiotic system and its efficient degradation of lignocellulose. Biosci. Biotechnol. Biochem. 2009, 73, 2561–2567. [Google Scholar] [CrossRef] [PubMed]
- Hongoh, Y. Toward the functional analysis of uncultivable, symbiotic microorganisms in the termite gut. Cell. Mol. Life Sci. 2011, 68, 1311–1325. [Google Scholar] [CrossRef] [PubMed]
- Brune, A.; Emerson, D.; Breznak, J. The termite gut microflora as an oxygen sink: Microelectrode determination of oxygen and pH gradients in guts of lower and higher termites. Appl. Environ. Microbiol. 1995, 61, 2681–2687. [Google Scholar] [PubMed]
- Sethi, A.; Slack, J.M.; Kovaleva, E.S.; Buchman, G.W.; Scharf, M.E. Lignin-associated metagene expression in a lignocellulose-digesting termite. Insect Biochem. Mol. Biol. 2013, 43, 91–101. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Ivanova, N.; Kirton, E.; Allgaier, M.; Bergin, C.; Scheffrahn, R.H.; Kyrpides, N.C.; Warnecke, F.; Tringe, S.G.; Hugenholtz, P. Comparative metagenomic and metatranscriptomic analysis of hindgut paunch microbiota in wood- and dung-feeding higher termites. PLoS ONE 2013, 8, e61126. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, J.; Millet, C.; Tarayre, C.; Brasseur, C.; Destain, J.; Vandenbol, M.; Thonart, P.; Portetelle, D.; De Pauw, E.; Haubruge, E.; et al. Symbiont diversity in Reticulitermes santonensis (Isoptera: Rhinotermitidae): Investigation strategy through Proteomics. Environ. Entomol. 2013, 42, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Syrenne, R.; Sun, S.; Yuan, J.S. Exploration of Natural Biomass Utilization Systems (NBUS) for advanced biofuel—From systems biology to synthetic design. Curr. Opin. Biotechnol. 2014, 27, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, G.; Tsuboi, Y.; Kihara, K.; Saitou, S.; Moriya, S.; Lo, N.; Kikuchi, J. Metabolomic profiling of 13C-labelled cellulose digestion in a lower termite: Insights into gut symbiont function. Proc. R. Soc. B. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Ralston-Hooper, K.; Jannasch, A.; Adamec, J.; Sepulveda, M. The use of two-dimensional gas chromatography-time-of-flight mass spectrometry (GC×GC-TOF-MS) for metabolomic analysis of polar metabolites. Methods Mol. Biol. 2011, 708, 205–211. [Google Scholar] [PubMed]
- Kamleh, M.A.; Dow, J.A.; Watson, D.G. Applications of mass spectrometry in metabolomic studies of animal model and invertebrate systems. Brief. Funct. Genom. Proteom. 2009, 8. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.G., Jr.; Welch, S.M.; Turner, W.E.; Sjödin, A.; Focant, J.-F. Cryogenic zone compression for the measurement of dioxins in human serum by isotope dilution at the attogram level using modulated gas chromatography coupled to high resolution magnetic sector mass spectrometry. J. Chromatogr. A 2011, 1218, 3274–3281. [Google Scholar] [CrossRef] [PubMed]
- Brune, A. Termite guts: The world’s smallest bioreactors. Trends Biotechnol. 1998, 16, 16–21. [Google Scholar] [CrossRef]
- Evans, T.A.; Forschler, B.T.; Grace, J.K. Biology of invasive termites: A worldwide review. Ann. Rev. Entomol. 2013, 58, 455–474. [Google Scholar] [CrossRef] [PubMed]
- Lewis, V.R.; Sutherland, M.; Haverty, M.I. Pest Notes: Subterranean and Other Termites. UC ANR Publication 7415 (2014). Available online: http://www.ipm.ucdavis.edu/PDF/PESTNOTES/pntermites.pdf (accessed on 9 March 2016).
- Schauer, C.; Thompson, C.L.; Brune, A. The bacterial community in the gut of the cockroach Shelfordella lateralis reflects the close evolutionary relatedness of cockroaches and termites. Appl. Environ. Microbiol. 2012, 78. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.H.; Lee, D.Y.; Skogerson, K.; Wohlgemuth, G.; Choi, I.G.; Fiehn, O.; Fiehn, O.; Kim, K.H. Global metabolic profiling of plant cell wall polysaccharide degradation by Saccharophagus degradans. Biotechnol. Bioeng. 2010, 105, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Wachsmuth, C.J.; Vogl, F.C.; Oefner, P.J.; Dettmer, K. Gas Chromatographic Techniques in Metabolomics. In Chromatographic Methods in Metabolomics; Hyotylainen, T., Wiedmer, S., Eds.; RCS Publishing: Cambridge, UK, 2013; pp. 87–105. [Google Scholar]
- Magel, E. Physiology of cambial growth, storage of reserves and heartwood formation. In Trends in European Forest Tree Physiology Research; Huttunen, S., Heikkila, H., Bucher, J., Sundberg, B., Jarvis, P., Matyssek, R., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 19–32. [Google Scholar]
- Austin, J.; Szalanski, A.; Scheffrahn, R.; Messenger, M.; Dronnet, S.; Bagnères, A.G. Genetic evidence for the synonymy of two Reticulitermes species: Reticulitermes flavipes and Reticulitermes santonensis. Ann. Entomol. Soc. Am. 2005, 98, 395–401. [Google Scholar] [CrossRef]
- Breznak, J.A. Ecology of prokaryotic microbes in the guts of wood- and litter-feeding termites. In Termites: Evolution, Sociality, Symbiosis, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 209–231. [Google Scholar]
- Potrikus, C.J.; Breznak, J.A. Gut bacteria recycle uric acid nitrogen in termites: A strategy for nutrient conservation. Proc. Natl. Acad. Sci. USA 1981, 78, 4601–4605. [Google Scholar] [CrossRef] [PubMed]
- Brune, A.; Ohkuma, M. Role of the Termite gut Microbiota in Symbiotic digestion. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 439–475. [Google Scholar]
- Machida, M.; Kitade, O.; Miura, T.; Matsumoto, T. Nitrogen recycling through proctodeal trophallaxis in the Japenese damp-wood termite Hodotermopsis japonica (Isoptera). Insectes Soc. 2001, 48, 52–56. [Google Scholar] [CrossRef]
- Brune, A.; Miambi, E.; Breznak, J.A. Roles of oxygen and the intestinal microflora in the metabolism of lignin-derived phenylpropanoids and other monoaromatic compounds by termites. Appl. Environ. Microbiol. 1995, 61, 2688–2695. [Google Scholar] [PubMed]
- Meijer, S.; Panagiotou, G.; Olsson, L.; Nielsen, J. Physiological characterization of xylose metabolism in Aspergillus niger under oxygen-limited conditions. Biotechnol. Bioeng. 2007, 98, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghany, S.E.; Day, I.; Heuberger, A.L.; Broeckling, C.D.; Reddy, A.S.N. Metabolic engineering of Arabidopsis for butanetriol production using bacterial genes. Metab. Eng. 2013, 20, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.; Molefe, M.N.; Frost, J.W. Microbial synthesis of the energetic material precursor 1,2,4-butanetriol. J. Am. Chem. Soc. 2003, 125, 12998–12999. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Filloux, A. Pseudomonas-A Model System in Biology; Springer: New York, NY, USA, 2007. [Google Scholar]
- Buslov, D.K.; Kaputski, F.N.; Sushko, N.I.; Torgashev, V.I.; Solov’eva, L.V.; Tsarenkov, V.M.; Zubets, O.V.; Larchenko, L.V. Infrared spectroscopic analysis of the structure of xylans. J. Appl. Spectrosc. 2009, 76, 801–805. [Google Scholar] [CrossRef]
- Kumar, A.; Bachhawat, A.K. Pyroglutamic acid: Throwing light on a lightly studied metabolite. Curr. Sci. 2012, 102, 288–297. [Google Scholar]
- Ingram, L.O.; Buttke, T.M. Effects of alcohols on micro-organisms. Adv. Microb. Physiol. 1984, 25, 253–300. [Google Scholar] [PubMed]
- Chi, Z.; Kohlwein, S.D.; Paltauf, F. Role of phosphatidylinositol (PI) in ethanol production and ethanol tolerance by a high ethanol producing yeast. J. Ind. Microbiol. Biotechnol. 1999, 22, 58–63. [Google Scholar] [CrossRef]
- Furukawa, K.; Kitano, H.; Mizoguchi, H.; Hara, S. Effect of cellular inositol content on ethanol tolerance of Saccharomyces cerevisiae in sake brewing. J. Biosci. Bioeng. 2004, 98, 107–113. [Google Scholar] [CrossRef]
- Krause, E.L.; Villa-García, M.J.; Henry, S.A.; Walker, L.P. Determining the effects of inositol supplementation and the opi1 mutation on ethanol tolerance of Saccharomyces cerevisiae. Ind. Biotechnol. 2007, 3, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Yamaguchi, M.; Morinaga, T.; Kinehara, M.; Ikeuchi, M.; Ashida, H.; Fujita, Y. Myo-inositol catabolism in Bacillus subtilis. J. Biol. Chem. 2008, 283, 10415–10424. [Google Scholar] [CrossRef] [PubMed]
- Tarayre, C.; Brognaux, A.; Brasseur, C.; Bauwens, J.; Millet, C.; Mattéotti, C.; Destain, J.; Vandenbol, M.; Portetelle, D.; De Pauw, E.; et al. Isolation and cultivation of a xylanolytic Bacillus Subtilis extracted from the gut of the termite Reticulitermes santonensis. Appl. Biochem. Biotechnol. 2013, 171, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Scharf, M.E. Omic research in termites: An overview and a roadmap. Front. Genet. 2015, 6. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | Amines | Organic Acids | Sugars | Others |
|---|---|---|---|---|
| Alanine | 2-Piperidone | 2-Ketogluconic acid | Arabitinol | 1,2,4-butanetriol |
| Asparagine | Cadaverine | 2-Ketoglutaric acid | Erythrose | Phosphate |
| Aspartic acid | Nicotinamide | 2-Ketovaline | Fructose | Phosphite |
| Beta-Alanine | N-Methylvaleramide | Benzoic acid | Galactose | |
| Glutamic acid | Putrescine | Caproic acid | Glucose | |
| Glutamine | Citric acid | Inositol Isomer | ||
| Glycine | Fumaric acid | Inositol myo- | ||
| Isoleucine | Glyceric acid | Meso-Erythritol | ||
| Leucine | Hydroxyphenylacetic acid | Rhamnose | ||
| N-acetyl-lysine | Lactic acid | Xylose | ||
| 5-oxoproline | Malic acid | |||
| Ornithine | Oxalic acid | |||
| Phenylalanine | Phenylacetic acid | |||
| Serine | Phenylpropanoic acid | |||
| Threonine | Pipecolinic acid | |||
| Tyrosine | Succinic acid | |||
| Valine | Uric acid |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brasseur, C.; Bauwens, J.; Tarayre, C.; Millet, C.; Mattéotti, C.; Thonart, P.; Destain, J.; Francis, F.; Haubruge, E.; Portetelle, D.; et al. GC×GC-TOFMS for the Analysis of Metabolites Produced by Termites (Reticulitermes flavipes) Bred on Different Carbon Sources. Separations 2016, 3, 19. https://doi.org/10.3390/separations3020019
Brasseur C, Bauwens J, Tarayre C, Millet C, Mattéotti C, Thonart P, Destain J, Francis F, Haubruge E, Portetelle D, et al. GC×GC-TOFMS for the Analysis of Metabolites Produced by Termites (Reticulitermes flavipes) Bred on Different Carbon Sources. Separations. 2016; 3(2):19. https://doi.org/10.3390/separations3020019
Chicago/Turabian StyleBrasseur, Catherine, Julien Bauwens, Cédric Tarayre, Catherine Millet, Christel Mattéotti, Philippe Thonart, Jacqueline Destain, Frédéric Francis, Eric Haubruge, Daniel Portetelle, and et al. 2016. "GC×GC-TOFMS for the Analysis of Metabolites Produced by Termites (Reticulitermes flavipes) Bred on Different Carbon Sources" Separations 3, no. 2: 19. https://doi.org/10.3390/separations3020019