
Minor Wheat Protein Fractions Analysis by Using Capillary Zone Electrophoresis


Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Materials
2.2. Extraction of CM Protein Fraction
2.3. Capillary Zone Electrophoresis (CZE)
2.4. Chemicals
3. Results and Discussion
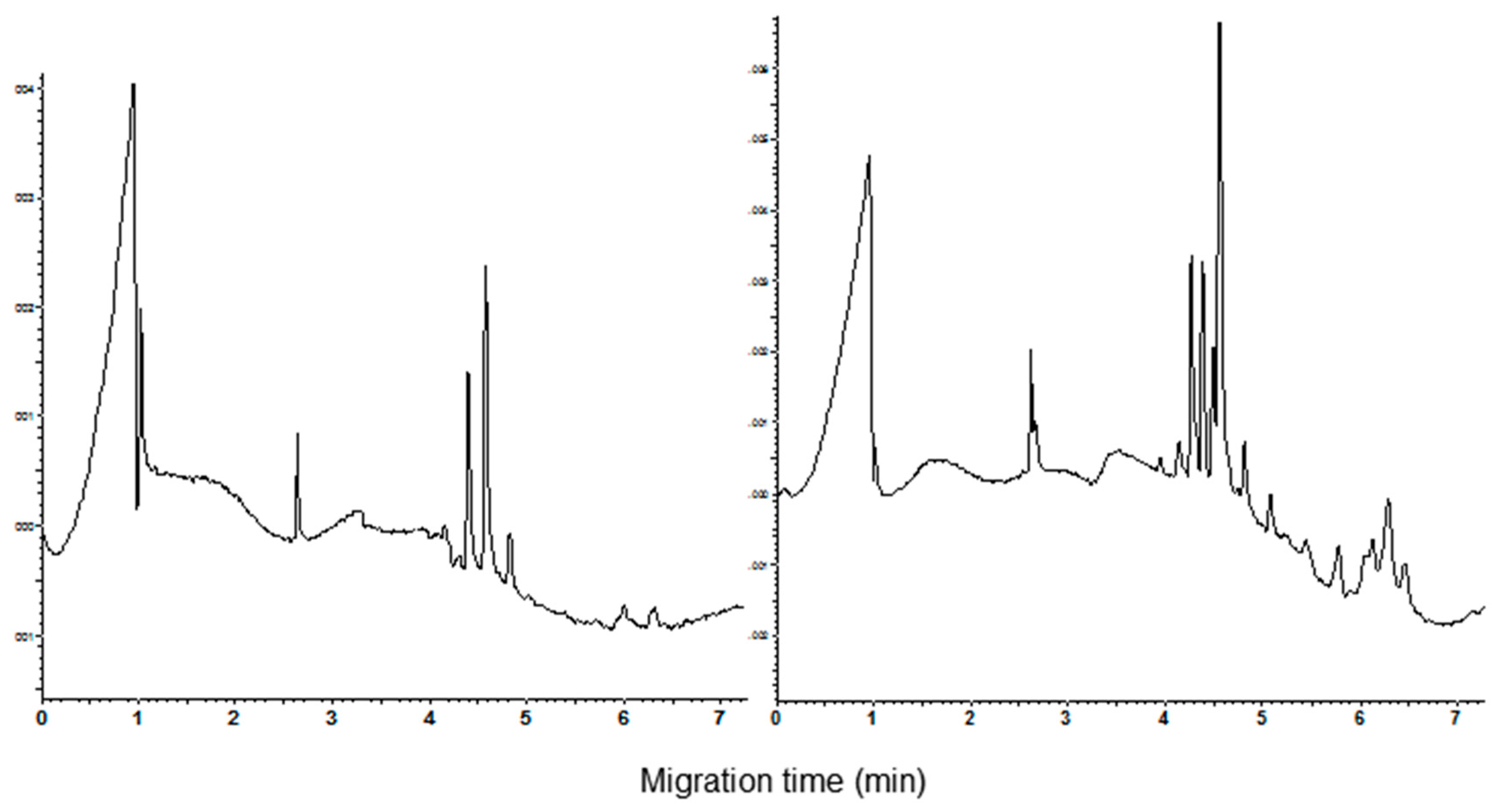
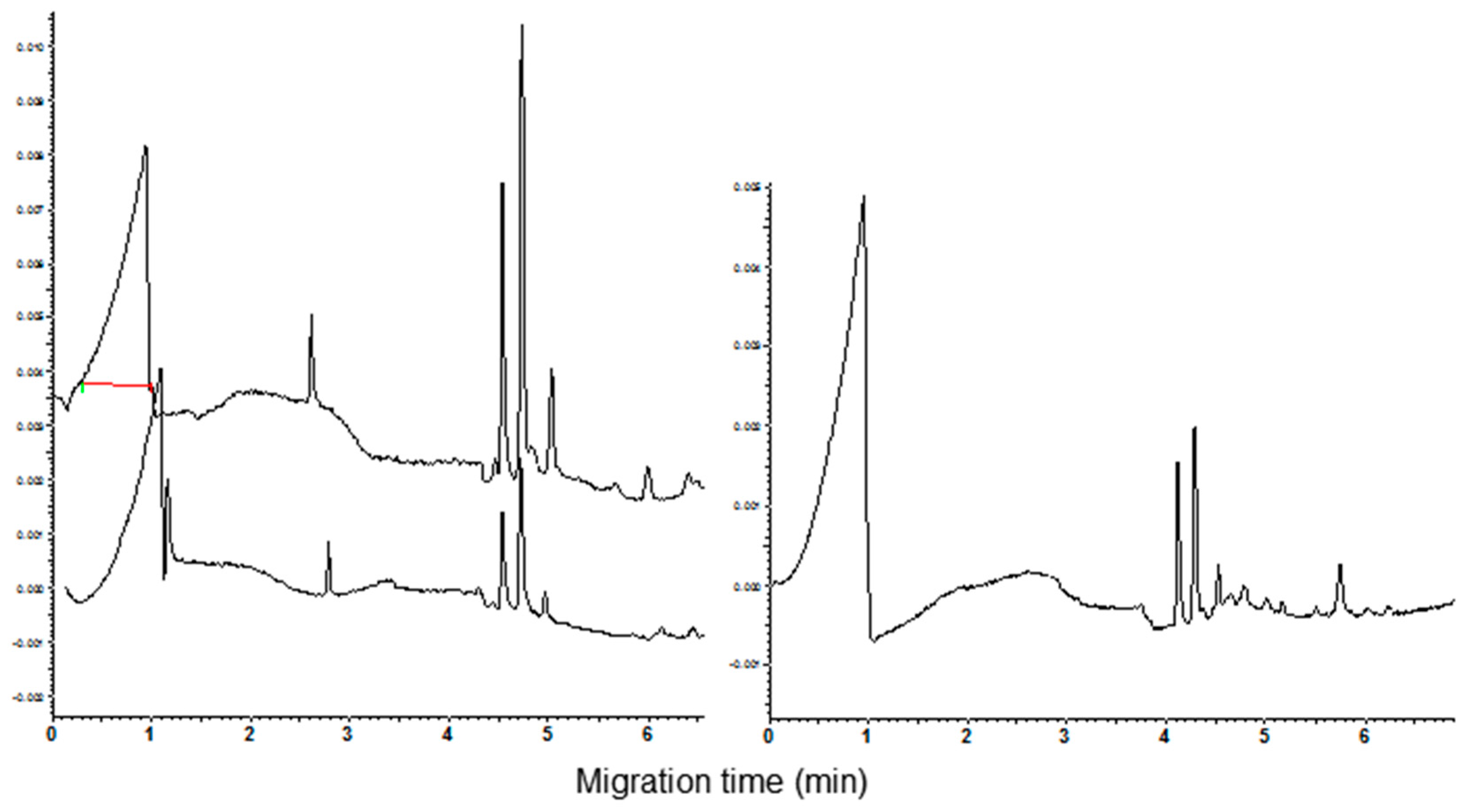
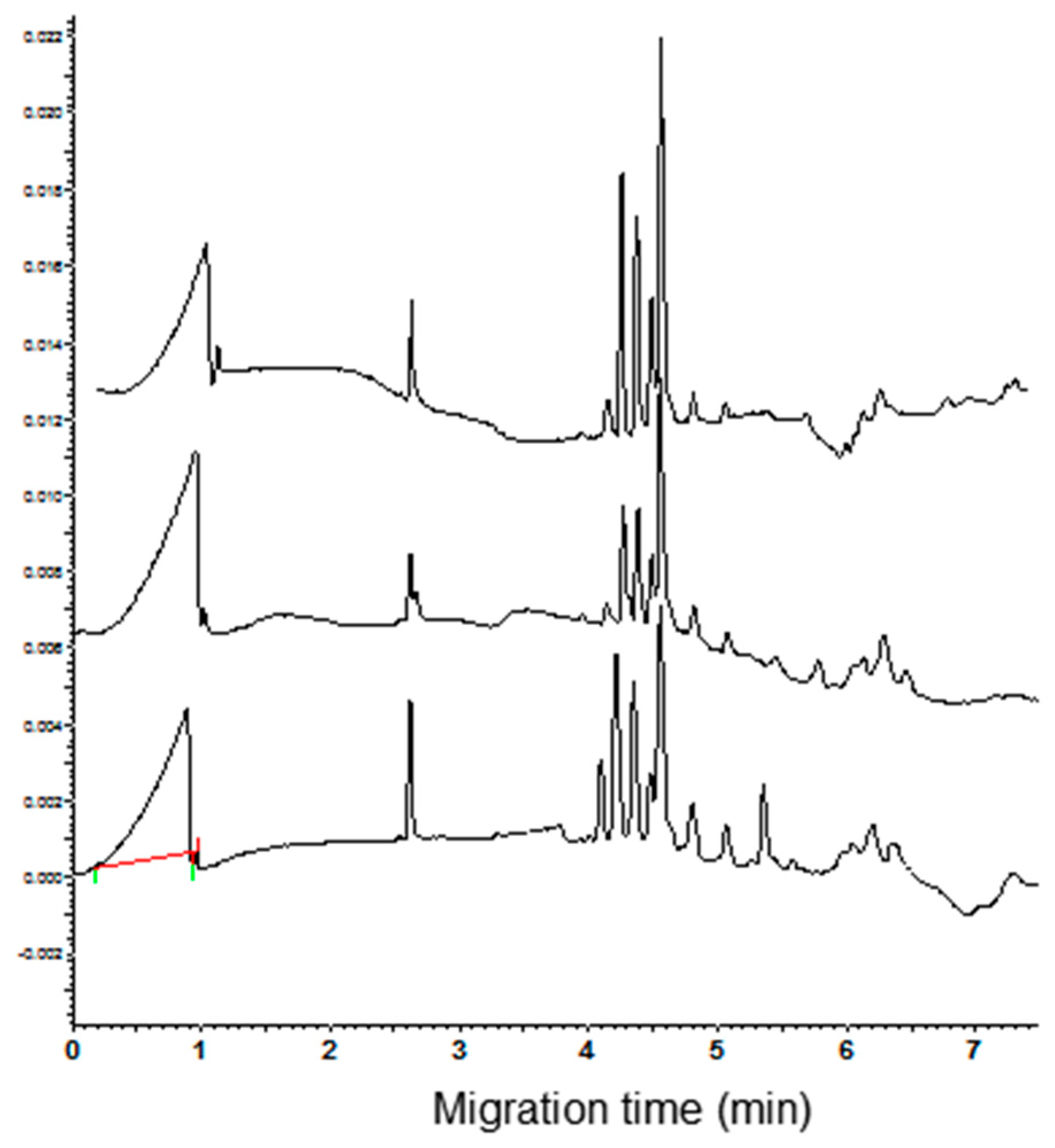
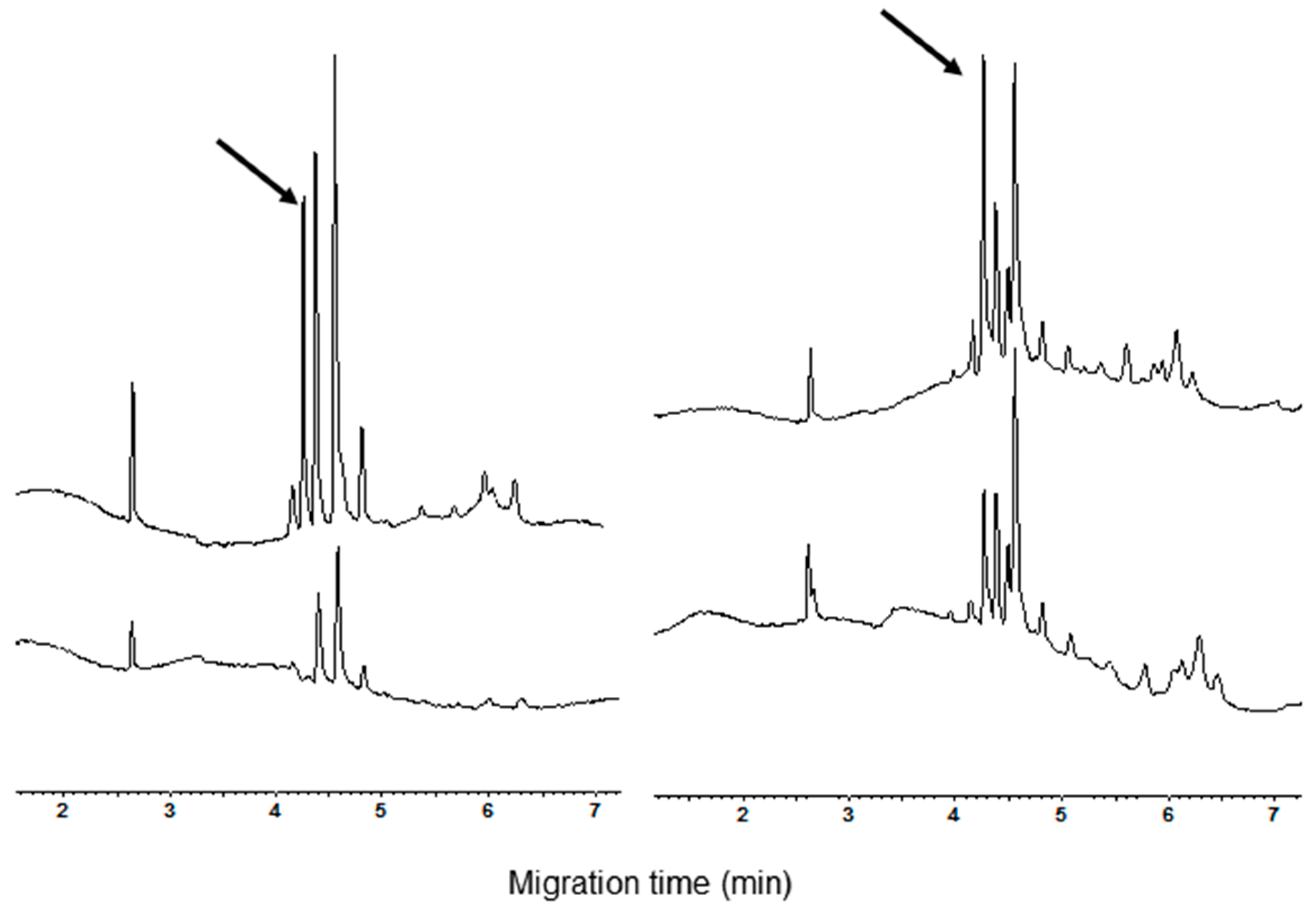
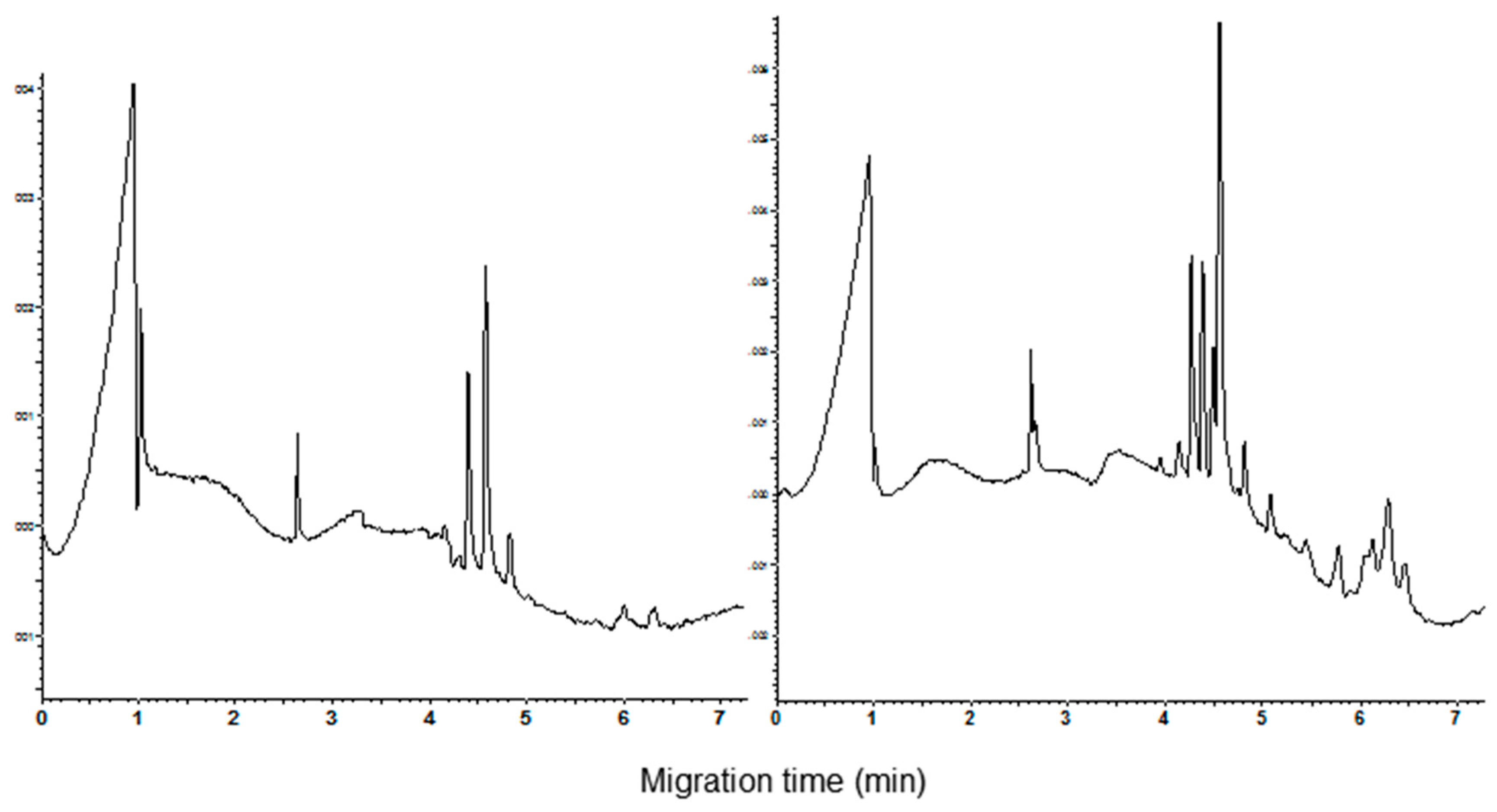
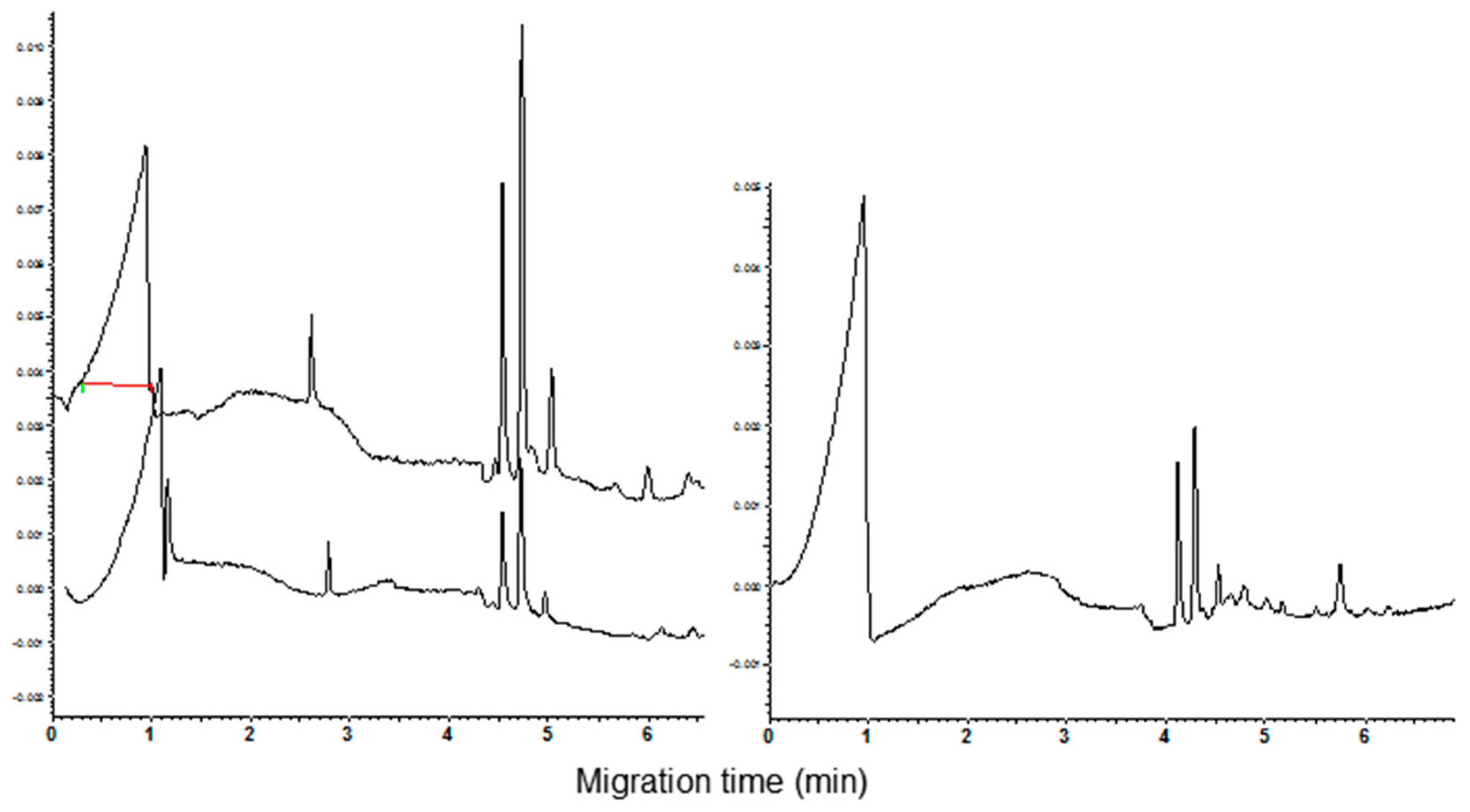
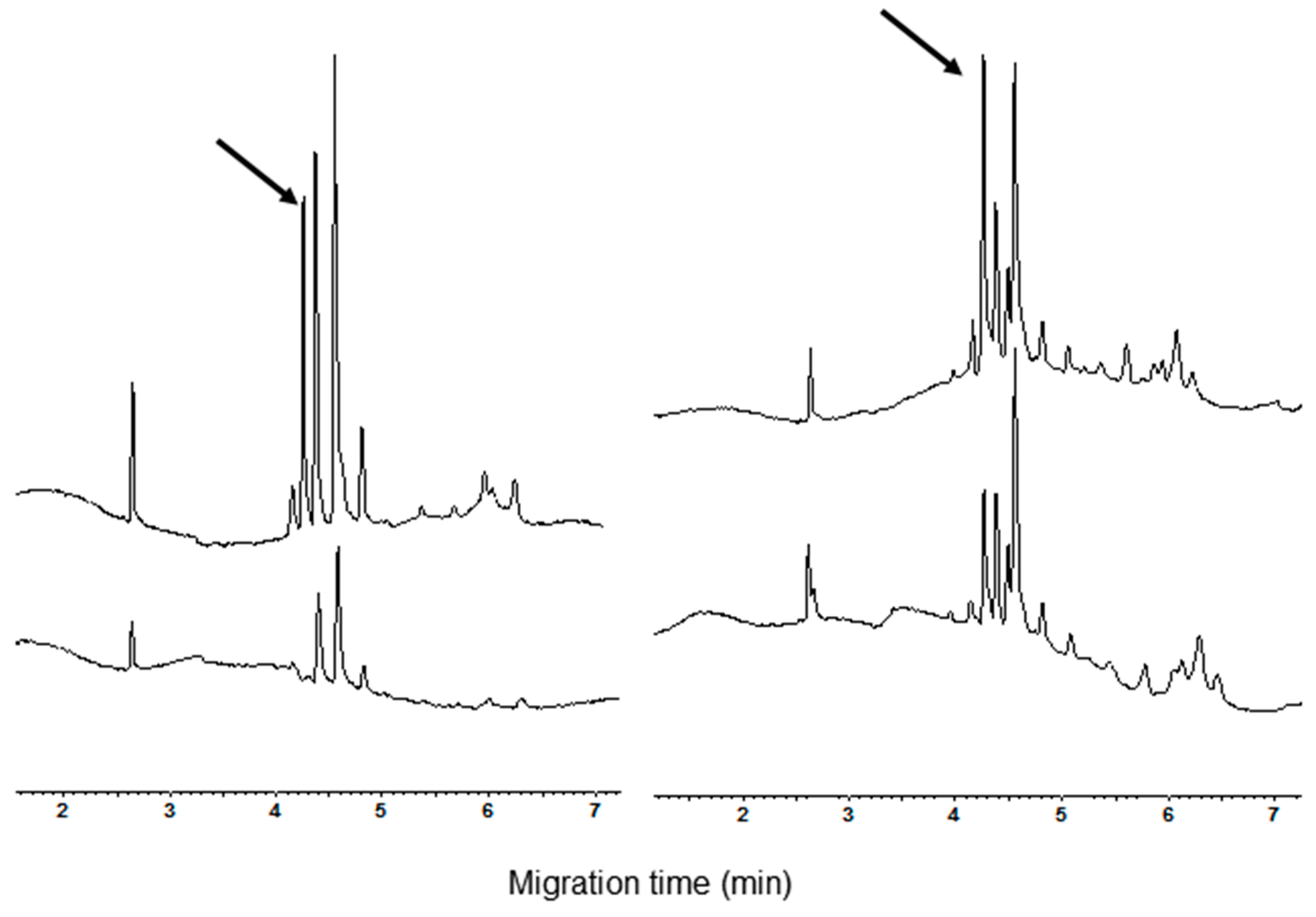
3.1. Polyacrylamide Gel Electrophoresis and Capillary Zone Electrophoresis of CM Extracts
3.2. Qualitative and Quantitative Analysis of CM Peaks
4. Conclusions
Conflicts of Interest
References
- Tatham, A.S.; Shewry, P.R. Allergenes in wheat and related cereals. Clin. Exp. Allergy 2008, 38, 1712–1726. [Google Scholar] [PubMed]
- Pietzak, M. Celiac disease, wheat allergy, and gluten sensitivity: When gluten free is not a fad. J. Parenter. Enter. Nutr. 2012, 36, 68S–75S. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Grossi-de-Sá, M.F. Plant alpha-amylase inhibitors and their interaction with insect alpha-amylases. Structure, function and potential for crop protection. Eur. J. Biochem. 2002, 269, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Bloch, C.; Silva, C.P.; Grossi de Sá, M.F. Activity of wheat α-amylase inhibitors towards bruchid α-amylases and structural explanation of observed specificities. Eur. J. Biochem. 2000, 267, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Fontanini, D.; Capocchi, A.; Muccilli, V.; Saviozzi, F.; Cunsolo, V.; Saletti, R.; Foti, S.; Galleschi, L. Dimeric inhibitors of human salivary alpha-amylase from emmer (Triticum dicoccon Schrank) seeds. J. Agric. Food Chem. 2007, 55, 10452–10460. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, G.; Quirce, S.; Diaz-Perales, A. Wheat allergens associated with baker’s asthma. J. Investig. Allergol. Clin. Immunol. 2011, 21, 81–92. [Google Scholar] [PubMed]
- Carbonero, P.; Garcia-Olmedo, F. A multigene family of trypsin/alpha-amylase inhibitors from cereals. In Seed Proteins; Shewry, P.R., Casey, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 617–633. [Google Scholar]
- Salcedo, G.; Sanchez-Monge, R.; Garcia-Casado, G.; Amentia, A.; Gomez, L.; Barber, D. The cereal α-amylase/trypsin inhibitor family associated with bakers’ asthma and food allergy. In Plant Food Allergens; Mill, E.N.C., Shewry, P.R., Eds.; Blackwell Publishing Company: Oxford, UK, 2004; pp. 70–86. [Google Scholar]
- Zoccatelli, G.; Pellegrina, C.D.; Mosconi, S.; Consolini, M.; Veneri, G.; Chignola, R.; Peruffo, A.; Rizzi, C. Full-fledged proteomic analysis of bioactive wheat amylase inhibitors by a 3-D analytical technique: Identification of new heterodimeric aggregation states. Electrophoresis 2007, 28, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Dolnik, V. Capillary electrophoresis of proteins 2005–2007. Electrophoresis 2008, 29, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Bean, S.R.; Tilley, M. Separation of water soluble proteins from cereals by free zone capillary electrophoresis. Cereal Chem. 2003, 80, 505–515. [Google Scholar] [CrossRef]
- Piergiovanni, A.R. Extraction and separation of water-soluble proteins from different wheat species by acidic capillary electrophoresis. J. Agric. Food Chem. 2007, 55, 3850–3856. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Ma, W.; Chen, J.; Wang, K.; Li, J.; Wang, S.; Bekes, F.; Appels, R.; Yan, Y. Characterization and comparative analysis of wheat high molecular weight glutenin subunits by SDS-PAGE, RP-HPLC, HPCE and MALDI-TOF-MS. J. Agric. Food Chem. 2010, 58, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Bean, S.R.; Lookhart, G.L. Ultrafast capillary electrophoretic analysis of cereal storage proteins and its applications to protein characterisation and cultivar differentiation. J. Agric. Food Chem. 2000, 48, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Piergiovanni, A.R. Estimating gliadin and albumin variation at intra and inter-accession level in the USDA Oriental wheat (Triticum. turanicum. L. subsp. turanicum (Jakubz.) (A. Löve & D. Löve) collection using capillary zone electrophoresis. Cereal Chem. 2009, 86, 37–43. [Google Scholar]
- Scholz, E.; Ganzler, K.; Gergely, S.; Salgo, A. Use of capillary electrophoresis to monitor wheat maturation. Chromatographia 2002, 56, 127–130. [Google Scholar] [CrossRef]
- Charmet, G.; Robert, N.; Branlard, G.; Linossier, L.; Martre, P.; Triboit, E. Genetic analysis of dry matter and nitrogen accumulation and protein composition in wheat kernels. Theor. Appl. Genet. 2005, 111, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Kitta, K.; Ohnishi-Kameyama, M.; Ogawa, T.; Kawamoto, S. Detection of low-molecular weight allergens resolved on two-dimensional electrophoresis with acid-urea polyacrylamide gel. Anal. Biochem. 2006, 351, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.R.; Wei, Y.M.; Yan, Z.H.; Zheng, Y.L. Detection of single nucleotide polymorphysms in 24 kDa dimeric alpha-amylase inhibitors from cultivated wheat and its diploid putative progenitors. Biochim. Biophys. Acta 2005, 1723, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.R.; Yan, Z.H.; Wei, Y.M.; Nevo, E.; Baum, B.R.; Zheng, Y.L. Molecular charactersation of dimeric alpha-amylase inhibitor genes in wheat and development of genome allele-specific primers for the genes located on chromosome 3BS and 3DS. J. Cereal Sci. 2006, 43, 360–368. [Google Scholar] [CrossRef]
- Baur, X.; Posch, A. Characterized allergens causing bakers’ asthma. Allergy 1998, 53, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Maeda, K.; Murayama, R.; Di Magno, E.P. Character of a wheat amylase inhibitor preparation and effects on fasting human pancreaticobiliary secretions and hormones. Gastroenterology 1996, 111, 1313–1320. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cv. Name | α-AI Type III (µg/mg) |
|---|---|
| Chinese spring | 1.2 |
| Rio | 2.1 |
| Chopin | 2.0 |
| Autonomia | 2.4 |
| Bolero | 1.7 |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piergiovanni, A.R. Minor Wheat Protein Fractions Analysis by Using Capillary Zone Electrophoresis. Separations 2016, 3, 17. https://doi.org/10.3390/separations3020017
Piergiovanni AR. Minor Wheat Protein Fractions Analysis by Using Capillary Zone Electrophoresis. Separations. 2016; 3(2):17. https://doi.org/10.3390/separations3020017
Chicago/Turabian StylePiergiovanni, Angela Rosa. 2016. "Minor Wheat Protein Fractions Analysis by Using Capillary Zone Electrophoresis" Separations 3, no. 2: 17. https://doi.org/10.3390/separations3020017