Genome-Wide Identification, Expression Pattern Analysis and Evolution of the Ces/Csl Gene Superfamily in Pineapple (Ananas comosus)

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Identification of Ces /Csl Genes in Pineapple
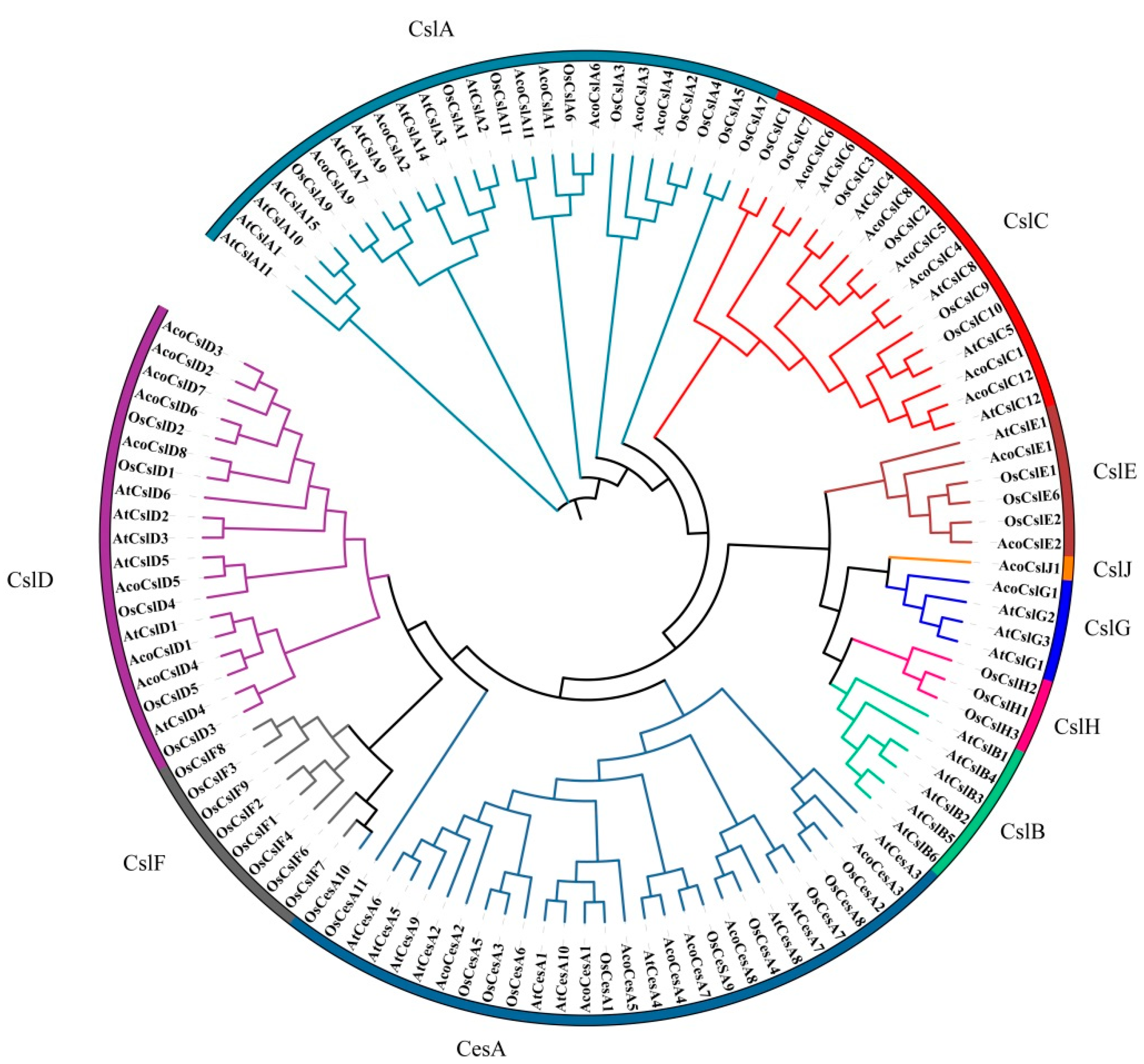
2.2. Phylogenetic Analysis of the Pineapple Ces/Csl Gene Superfamily
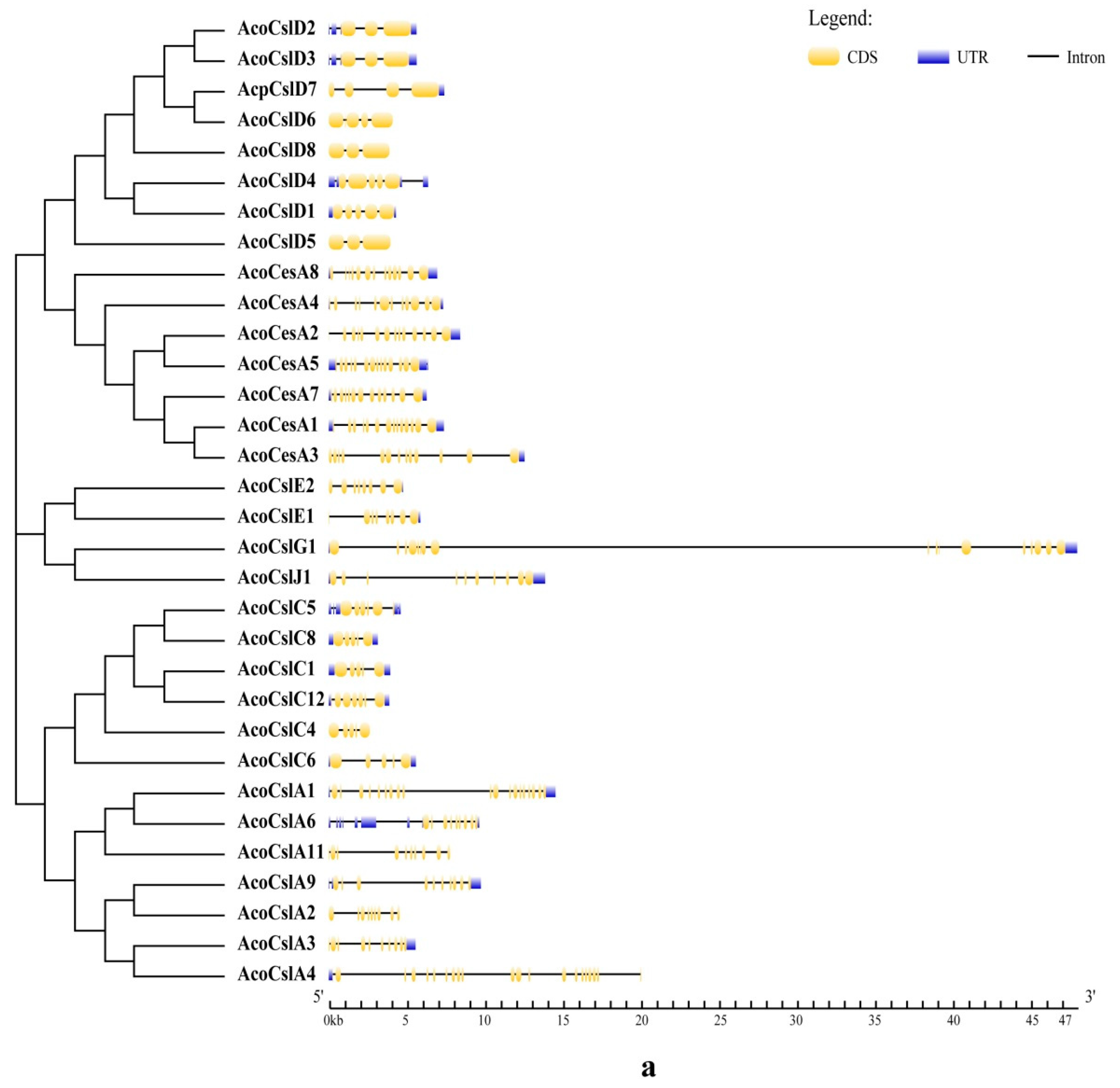
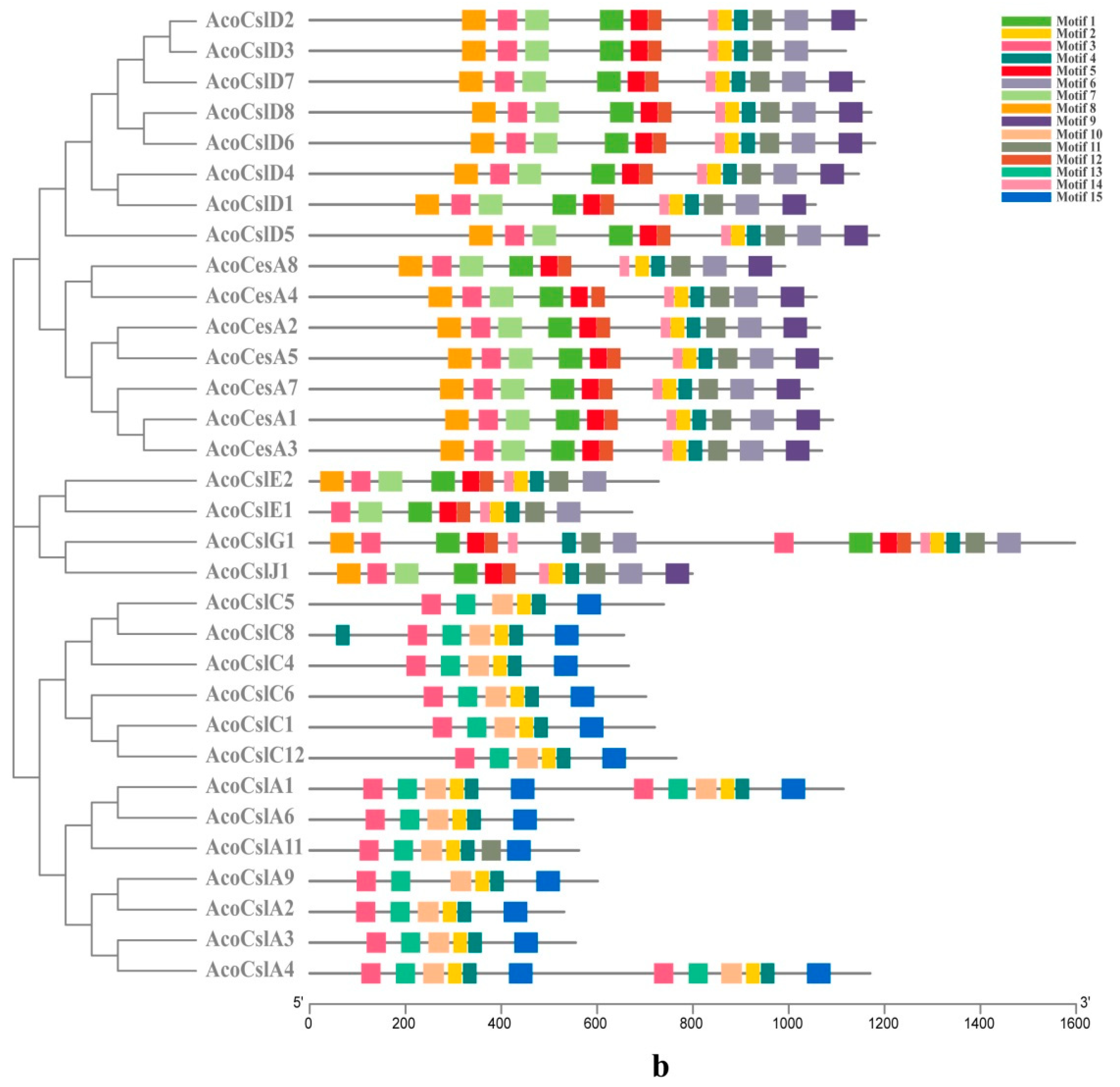
2.3. Gene Structure Analysis and Conserved Motif Identification
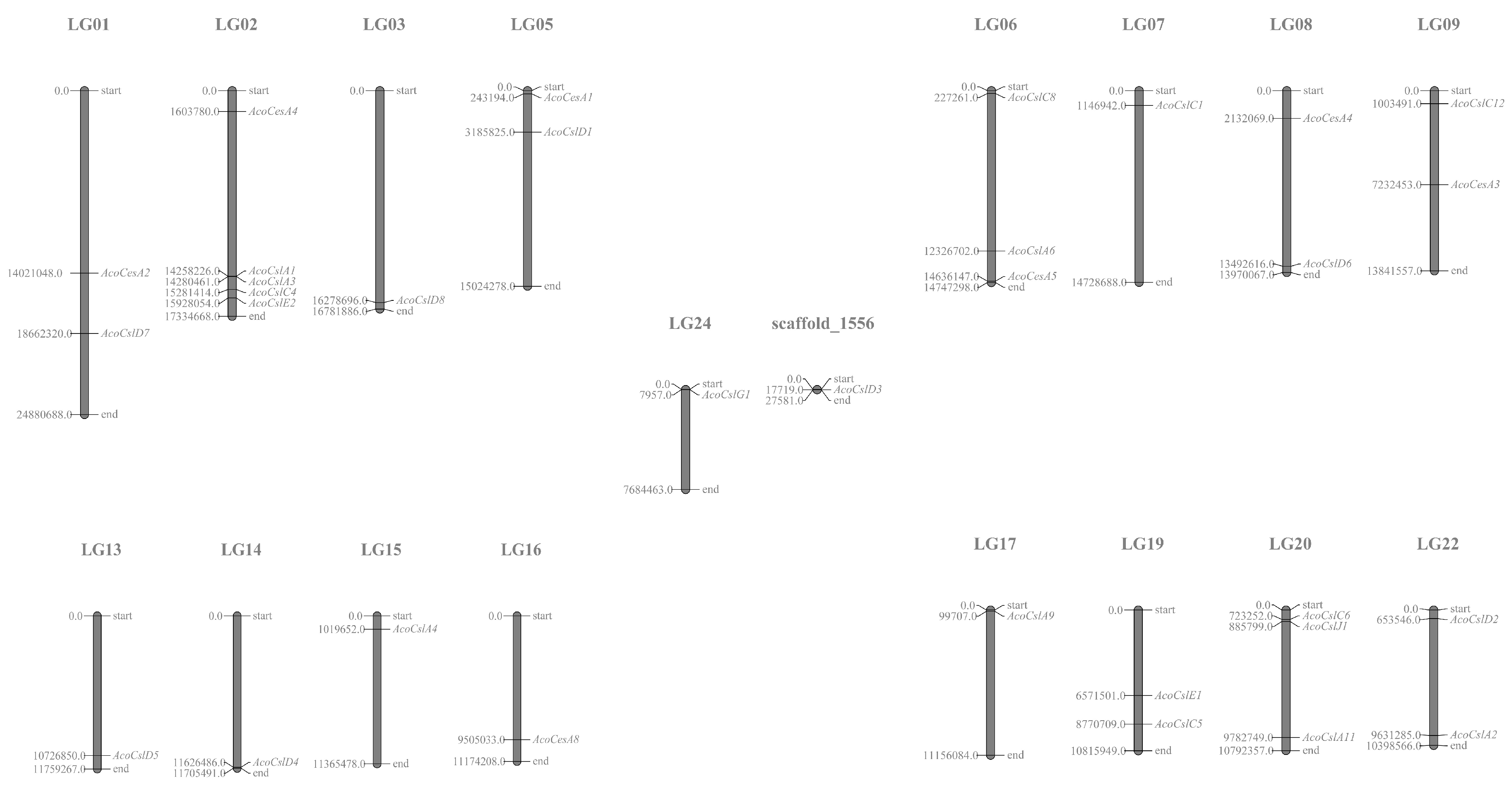
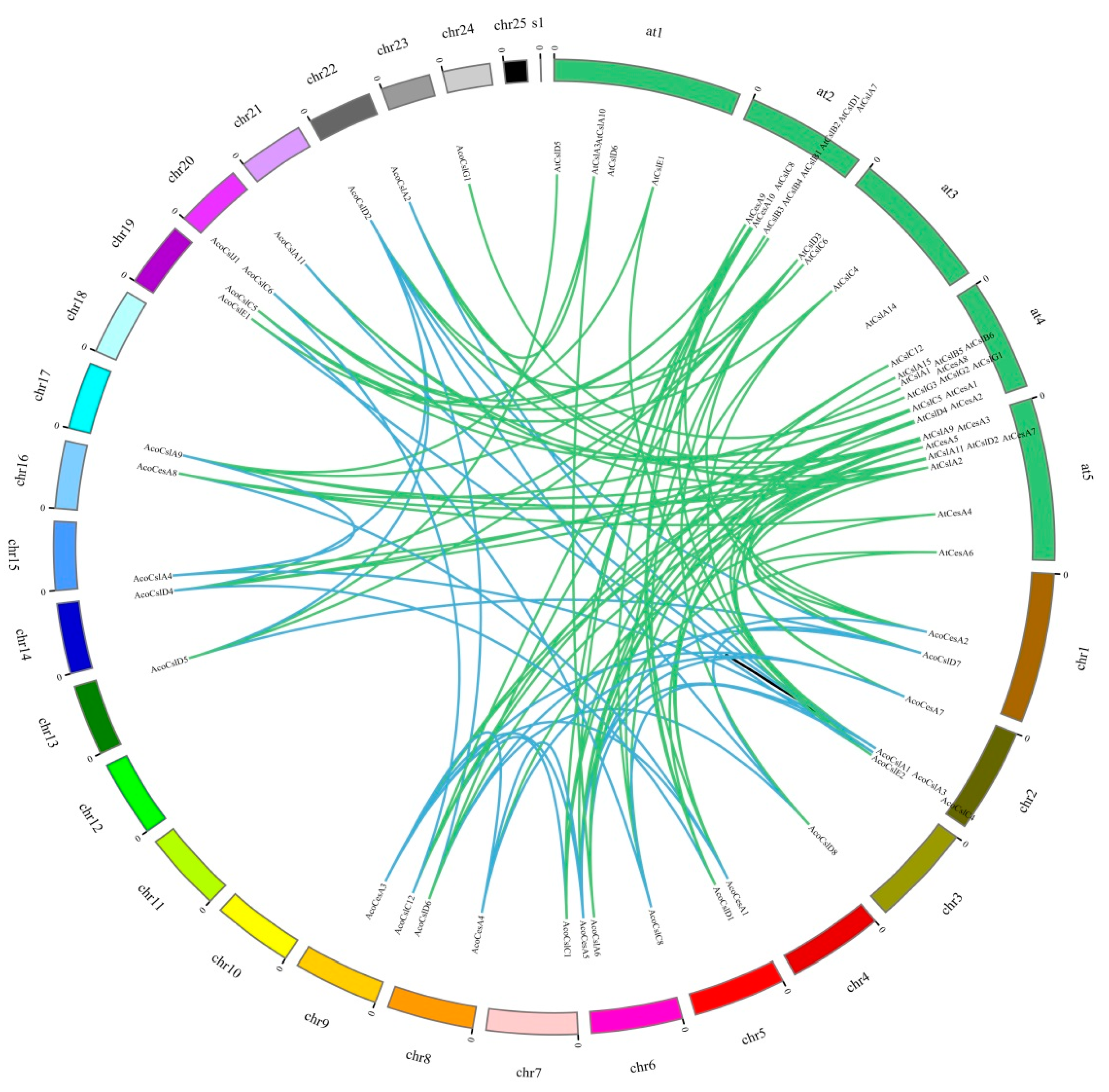
2.4. Synteny Analysis of Pineapple Ces/Csl Genes
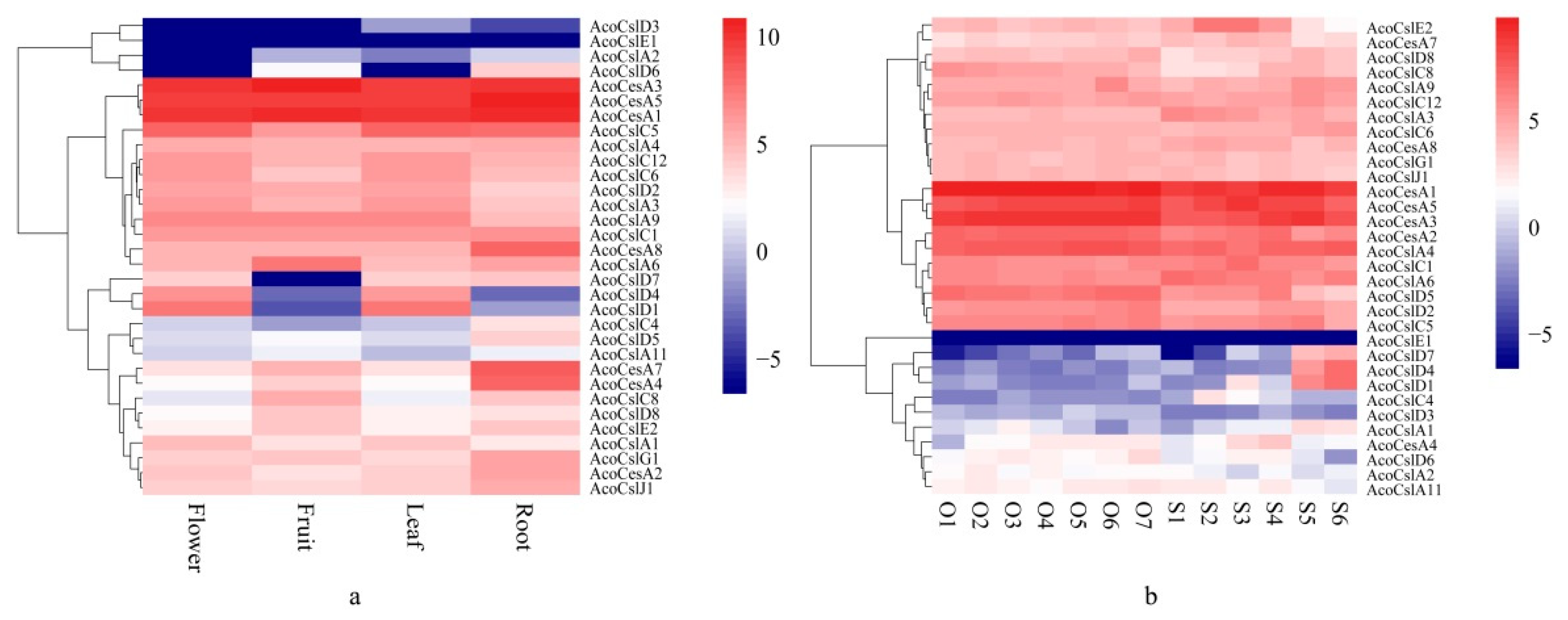
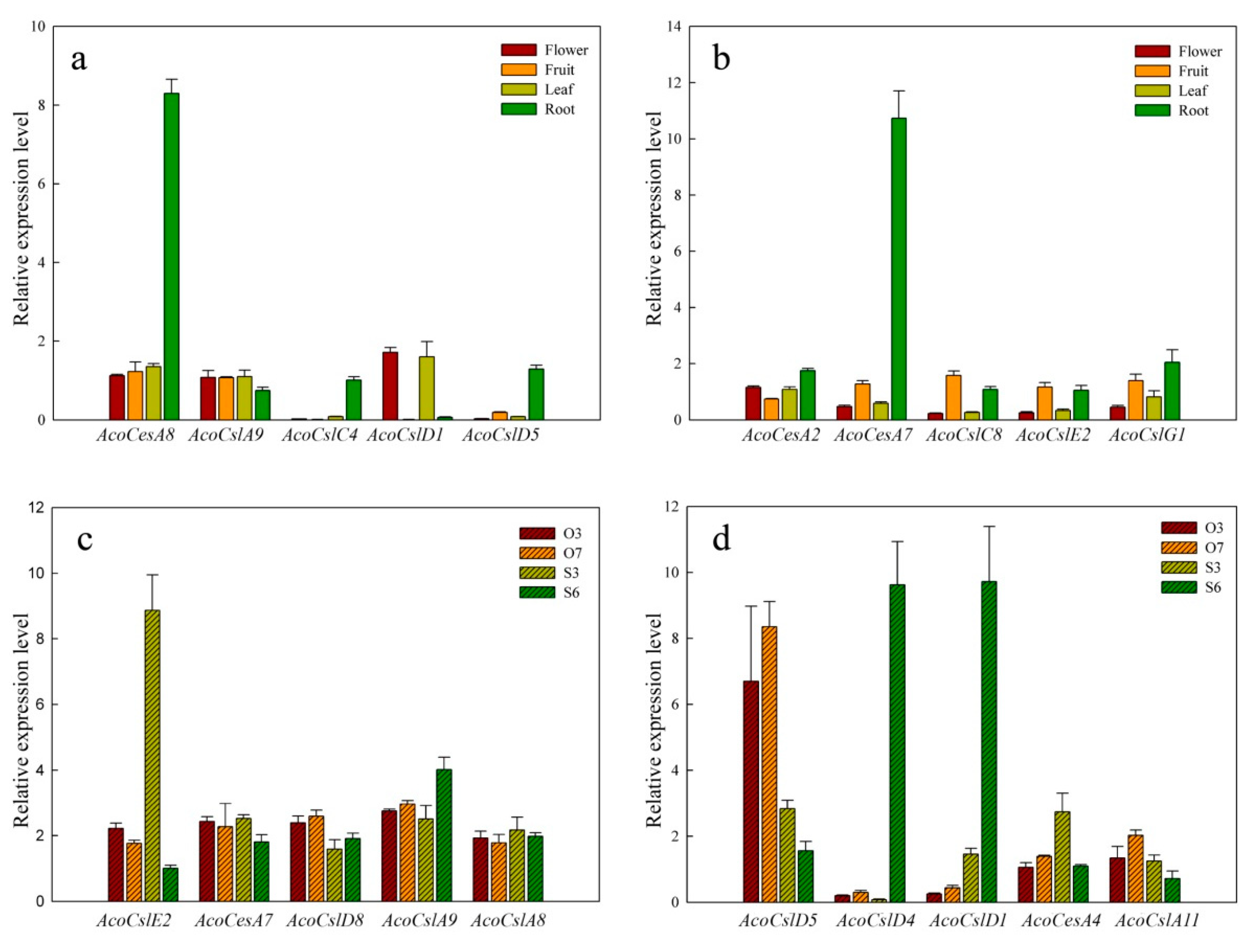
2.5. Expression Patterns of AcoCes/Csl Genes in Four Different Tissues
2.6. Expression of AcoCes/Csl Genes during Gametophyte Development
3. Discussion
4. Materials and Methods
4.1. Identification of Ces/Csl in Pineapple
4.2. Physicochemical Properties and Phylogenetic Analysis
4.3. Conserved Motifs Analysis of Pineapple Ces/Csl Protein
4.4. Chromosome Localization and Gene Structural Analysis of Pineapple Ces/Csl Genes
4.5. Synteny Analysis of Pineapple Ces/Csl Genes
4.6. RNA-Seq and qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Aco | Ananas comosus |
| At | Arabidopsis thaliana |
| Ces | Cellulose synthase |
| Csl | Cellulose synthase-like |
References
- Griffiths, J.S.; North, H.M. Sticking to cellulose: exploiting Arabidopsis seed coat mucilage to understand cellulose biosynthesis and cell wall polysaccharide interactions. New Phytol. 2017, 214, 959–966. [Google Scholar] [PubMed] [Green Version]
- Delmer, D.P. CELLULOSE BIOSYNTHESIS: Exciting Times for A Difficult Field of Study. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 245–276. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.E.; Burn, J.E.; Hocart, C.H. Towards the mechanism of cellulose synthesis. Trends Plant Sci. 2002, 7, 461–467. [Google Scholar] [CrossRef]
- Saxena, I.M.; Brown, R.M. Cellulose biosynthesis: Current views and evolving concepts. Ann. Bot.-London 2005, 96, 9–21. [Google Scholar]
- Somerville, C. Cellulose synthesis in higher plants. Annu. Rev. Cell. Dev. Biol. 2006, 22, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Crowell, E.F.; Gonneau, M.; Stierhof, Y.D.; Höfte, H.; Vernhettes, S. Regulated trafficking of cellulose synthases. Curr. Opin. Plant Biol. 2010, 13, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Li, S.D.; Gu, Y. Cellulose synthase complexes: composition and regulation. Front. Plant Sci. 2012, 3, 75. [Google Scholar] [CrossRef]
- Li, S.D.; Bashline, L.; Zheng, Y.Z.; Xin, X.R.; Huang, S.X.; Kong, Z.S.; Kim, S.H.; Cosgrove, D.J.; Gu, Y. Cellulose synthase complexes act in a concerted fashion to synthesize highly aggregated cellulose in secondary cell walls of plants. Proc. Natl. Acad. Sci. USA 2016, 113, 11348–11353. [Google Scholar] [CrossRef] [Green Version]
- Bessueille, L.; Bulone, V. A survey of cellulose biosynthesis in higher plants. Plant Biotechnol. 2008, 25, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Guerriero, G.; Fugelstad, J.; Bulone, V. What Do We Really Know about Cellulose Biosynthesis in Higher Plants? J. Integr. Plant Biol. 2010, 52, 161–175. [Google Scholar]
- Lerouxel, O.; Cavalier, D.M.; Liepman, A.H.; Keegstra, K. Biosynthesis of plant cell wall polysaccharides - a complex process. Curr. Opin. Plant Biol. 2006, 9, 621–630. [Google Scholar] [PubMed]
- Richmond, T.A.; Somerville, C.R. The Cellulose Synthase Superfamily. Plant Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, A.; Schwerdt, J.G.; Shirley, N.J.; Khor, S.F.; Neumann, K.; O’Donovan, L.A.; Lahnstein, J.; Collins, H.M.; Henderson, M.; Fincher, G.B.; et al. Revised Phylogeny of the Cellulose Synthase Gene Superfamily: Insights into Cell Wall Evolution. Plant Physiol. 2018, 177, 1124–1141. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.B.; Huang, J.L.; Xu, Y. The cellulose synthase superfamily in fully sequenced plants and algae. Bmc Plant Biol. 2009, 9, 99. [Google Scholar]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar]
- Morgan, J.L.W.; Strumillo, J.; Zimmer, J. Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature 2013, 493, U181–U192. [Google Scholar]
- Arioli, T.; Peng, L.; Betzner, A.S.; Burn, J.; Wittke, W.; Herth, W.; Camilleri, C.; Hofte, H.; Plazinski, J.; Birch, R.; et al. Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 1998, 279, 717–720. [Google Scholar]
- Cano-Delgado, A.; Penfield, S.; Smith, C.; Catley, M.; Bevan, M. Reduced cellulose synthesis invokes lignification and defense responses in Arabidopsis thaliana. Plant J. 2003, 34, 351–362. [Google Scholar]
- Fagard, M.; Desnos, T.; Desprez, T.; Goubet, F.; Refregier, G.; Mouille, G.; McCann, M.; Rayon, C.; Vernhettes, S.; Höfte, H. PROCUSTE1 Encodes a Cellulose Synthase Required for Normal Cell Elongation Specifically in Roots and Dark-Grown Hypocotyls of Arabidopsis. Plant Cell 2000, 12, 2409–2423. [Google Scholar] [CrossRef]
- Beeckman, T.; Przemeck, G.K.; Stamatiou, G.; Lau, R.; Terryn, N.; De Rycke, R.; Inze, D.; Berleth, T. Genetic complexity of cellulose synthase a gene function in Arabidopsis embryogenesis. Plant Physiol. 2002, 130, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.M.; Zeef, L.A.H.; Ellis, J.; Goodacre, R.; Turner, S.R. Identification of novel genes in Arabidopsis involved in secondary cell wall formation using expression profiling and reverse genetics. Plant Cell 2005, 17, 2281–2295. [Google Scholar] [CrossRef] [PubMed]
- Timmers, J.; Vernhettes, S.; Desprez, T.; Vincken, J.P.; Visser, R.G.F.; Trindade, L.M. Interactions between membrane-bound cellulose synthases involved in the synthesis of the secondary cell wall. Febs Lett. 2009, 583, 978–982. [Google Scholar] [PubMed] [Green Version]
- Wang, X.; Cnops, G.; Vanderhaeghen, R.; De Block, S.; Van Montagu, M.; Van Lijsebettens, M. AtCSLD3, a cellulose synthase-like gene important for root hair growth in arabidopsis. Plant Physiol. 2001, 126, 575–586. [Google Scholar]
- Hazen, S.P.; Scott-Craig, J.S.; Walton, J.D. Cellulose synthase-like genes of rice. Plant Physiol. 2002, 128, 336–340. [Google Scholar] [CrossRef]
- Burton, R.A.; Wilson, S.M.; Hrmova, M.; Harvey, A.J.; Shirley, N.J.; Stone, B.A.; Newbigin, E.J.; Bacic, A.; Fincher, G.B. Cellulose synthase-like CslF genes mediate the synthesis of cell wall (1,3;1,4)-beta-D-glucans. Science 2006, 311, 1940–1942. [Google Scholar] [CrossRef]
- Liu, X.L.; Liu, L.F.; Niu, Q.K.; Xia, C.A.; Yang, K.Z.; Li, R.; Chen, L.Q.; Zhang, X.Q.; Zhou, Y.H.; Ye, D. MALE GAMETOPHYTE DEFECTIVE 4 encodes a rhamnogalacturonan II xylosyltransferase and is important for growth of pollen tubes and roots in Arabidopsis. Plant J. 2011, 65, 647–660. [Google Scholar]
- Saxena, I.M.; Lin, F.C.; Brown, R.M. Cloning and sequencing of the cellulose synthase catalytic subunit gene of Acetobacter xylinum. Plant Mol. Biol. 1990, 15, 673–683. [Google Scholar] [CrossRef]
- Saxena, I.M.; Brown, R.M., Jr.; Fevre, M.; Geremia, R.A.; Henrissat, B. Multidomain architecture of beta-glycosyl transferases: implications for mechanism of action. J. Bacteriol. 1995, 177, 1419–1424. [Google Scholar] [Green Version]
- Wong, H.C.; Fear, A.L.; Calhoon, R.D.; Eichinger, G.H.; Mayer, R.; Amikam, D.; Benziman, M.; Gelfand, D.H.; Meade, J.H.; Emerick, A.W. Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proc. Natl. Acad. Sci. USA 1990, 87, 8130–8134. [Google Scholar] [CrossRef] [PubMed]
- Pear, J.R.; Kawagoe, Y.; Schreckengost, W.E.; Delmer, D.P.; Stalker, D.M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 12637–12642. [Google Scholar] [PubMed]
- Appenzeller, L.; Doblin, M.; Barreiro, R.; Wang, H.Y.; Niu, X.M.; Kollipara, K.; Carrigan, L.; Tomes, D.; Chapman, M.; Dhugga, K.S. Cellulose synthesis in maize: isolation and expression analysis of the cellulose synthase (CesA) gene family. Cellulose 2004, 11, 287–299. [Google Scholar] [CrossRef]
- Burton, R.A.; Shirley, N.J.; King, B.J.; Harvey, A.J.; Fincher, G.B. The CesA gene family of barley. Quantitative analysis of transcripts reveals two groups of co-expressed genes. Plant Physiol. 2004, 134, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Joshi, C.P.; Bhandari, S.; Ranjan, P.; Kalluri, U.C.; Liang, X.; Fujino, T.; Samuga, A. Genomics of cellulose biosynthesis in poplars. New Phytol. 2004, 164, 53–61. [Google Scholar]
- Djerbi, S.; Lindskog, M.; Arvestad, L.; Sterky, F.; Teeri, T.T. The genome sequence of black cottonwood (Populus trichocarpa) reveals 18 conserved cellulose synthase (CesA) genes. Planta 2005, 221, 739–746. [Google Scholar] [PubMed]
- Nairn, C.J.; Haselkorn, T. Three loblolly pine CesA genes expressed in developing xylem are orthologous to secondary cell wall CesA genes of angiosperms. New Phytol. 2005, 166, 907–915. [Google Scholar] [PubMed]
- Wang, L.Q.; Guo, K.; Li, Y.; Tu, Y.Y.; Hu, H.Z.; Wang, B.R.; Cui, X.C.; Peng, L.C. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. Bmc Plant Biol. 2010, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Sundari, B.K.R.; Dasgupta, M.G. Isolation of developing secondary xylem specific cellulose synthase genes and their expression profiles during hormone signalling in Eucalyptus tereticornis. J. Genet. 2014, 93, U403–U437. [Google Scholar]
- Guerriero, G.; Legay, S.; Hausman, J.F. Alfalfa Cellulose Synthase Gene Expression under Abiotic Stress: A Hitchhiker’s Guide to RT-qPCR Normalization. PLoS ONE 2014, 9, e103808. [Google Scholar]
- Suzuki, S.; Li, L.; Sun, Y.H.; Chiang, V.L. The cellulose synthase gene superfamily and biochemical functions of xylem-specific cellulose synthase-like genes in Populus trichocarpa. Plant Physiol. 2006, 142, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xiong, G.Y.; Li, R.; Cui, J.J.; Tang, D.; Zhang, B.C.; Pauly, M.; Cheng, Z.K.; Zhou, Y.H. Rice cellulose synthase-like D4 is essential for normal cell-wall biosynthesis and plant growth. Plant J. 2009, 60, 1055–1069. [Google Scholar] [PubMed] [Green Version]
- Hepton, A.; Hodgson, A.S.; Bartholomew, D.P.; Paull, R.E.; Rohrbach, K.G. The Pineapple: Botany, Production and Uses; CABI Publishing: Wallingford, UK, 2003; pp. 4895–4902. [Google Scholar]
- Ming, R.; VanBuren, R.; Wai, C.M.; Tang, H.; Schatz, M.C.; Bowers, J.E.; Lyons, E.; Wang, M.L.; Chen, J.; Biggers, E.; et al. The pineapple genome and the evolution of CAM photosynthesis. Nat. Genet. 2015, 47, 1435–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.P.; Miao, C.Y.; Chen, R.K.; Ming, R. Genome-Wide Comparative Analysis of Microsatellites in Pineapple. Trop. Plant Biol. 2016, 9, 117–135. [Google Scholar]
- Paull, R.; Chen, N.; Ming, R.; Wai, C.; Shirley, N.; Schwerdt, J.; Bulone, V. Carbon Flux and Carbohydrate Gene Families in Pineapple. Trop. Plant Biol. 2016, 9, 200–213. [Google Scholar]
- Wai, C.M.; Powell, B.; Ming, R.; Min, X.J. Analysis of Alternative Splicing Landscape in Pineapple (Ananas comosus). Trop. Plant Biol. 2016, 9, 150–160. [Google Scholar]
- Zhang, X.D.; Liang, P.P.; Ming, R. Genome-Wide Identification and Characterization of Nucleotide-Binding Site (NBS) Resistance Genes in Pineapple. Trop. Plant Biol. 2016, 9, 187–199. [Google Scholar]
- Zheng, Y.; Li, T.; Xu, Z.N.; Wai, C.M.; Chen, K.; Zhang, X.T.; Wang, S.P.; Ji, B.; Ming, R.; Sunkar, R. Identification of microRNAs, phasiRNAs and Their Targets in Pineapple. Trop. Plant Biol. 2016, 9, 176–186. [Google Scholar]
- Song, S.W.; Hao, L.Y.; Zhao, P.; Xu, Y.; Zhong, N.Q.; Zhang, H.J.; Liu, N. Genome-wide Identification, Expression Profiling and Evolutionary Analysis of Auxin Response Factor Gene Family in Potato (Solanum tuberosum Group Phureja). Sci. Rep.-UK 2019, 9. [Google Scholar]
- Xing, H.Y.; Pudake, R.N.; Guo, G.G.; Xing, G.F.; Hu, Z.R.; Zhang, Y.R.; Sun, Q.X.; Ni, Z.F. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. Bmc Genomics 2011, 12, 178. [Google Scholar]
- Han, Y.; Li, X.; Cheng, L.; Liu, Y.; Wang, H.; Ke, D.; Yuan, H.; Zhang, L.; Wang, L. Genome-Wide Analysis of Soybean JmjC Domain-Containing Proteins Suggests Evolutionary Conservation Following Whole-Genome Duplication. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Azam, S.M.; Liu, Y.; Rahman, Z.U.; Ali, H.; Yan, C.; Wang, L.; Priyadarshani, S.V.G.N.; Hu, B.; Huang, X.; Xiong, J. Identification, Characterization and Expression Profiles of Dof Transcription Factors in Pineapple (Ananas comosus L). Trop. Plant Biol. 2018, 11, 49–64. [Google Scholar] [CrossRef]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant cell wall biosynthesis: genetic, biochemical and functional genomics approaches to the identification of key genes. Plant Biotechnol. J. 2006, 4, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Farrokhi, N.; Bacic, A.; Fincher, G.B. Plant cell wall polysaccharide biosynthesis: real progress in the identification of participating genes. Planta 2005, 221, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Dwivany, F.M.; Yulia, D.; Burton, R.A.; Shirley, N.J.; Wilson, S.M.; Fincher, G.B.; Bacic, A.; Newbigin, E.; Doblin, M.S. The CELLULOSE-SYNTHASE LIKE C (CSLC) Family of Barley Includes Members that Are Integral Membrane Proteins Targeted to the Plasma Membrane. Mol. Plant 2009, 2, 1025–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doblin, M.S.; Pettolino, F.A.; Wilson, S.M.; Campbell, R.; Burton, R.A.; Fincher, G.B.; Newbigin, E.; Bacic, A. A barley cellulose synthase-like CSLH gene mediates (1,3;1,4)-beta-D-glucan synthesis in transgenic Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5996–6001. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.J.; Yoo, C.M.; Mutwil, M.; Jensen, J.K.; Hou, G.; Blaukopf, C.; Sorensen, I.; Blancaflor, E.B.; Scheller, H.V.; Willats, W.G. Functional analysis of the cellulose synthase-like genes CSLD1, CSLD2, and CSLD4 in tip-growing Arabidopsis cells. Plant Physiol. 2008, 148, 1238–1253. [Google Scholar] [CrossRef]
- Nobles, D.R.; Romanovicz, D.K.; Brown, R.M., Jr. Cellulose in cyanobacteria. Origin of vascular plant cellulose synthase? Plant Physiol. 2001, 127, 529–542. [Google Scholar] [CrossRef]
- Doblin, M.S.; Kurek, I.; Jacob-Wilk, D.; Delmer, D.P. Cellulose biosynthesis in plants: from genes to rosettes. Plant Cell Physiol. 2002, 43, 1407–1420. [Google Scholar] [CrossRef]
- Liepman, A.H.; Wilkerson, C.G.; Keegstra, K. Expression of cellulose synthase-like (Csl) genes in insect cells reveals that CslA family members encode mannan synthases. Proc. Natl. Acad. Sci. USA 2005, 102, 2221–2226. [Google Scholar] [CrossRef] [PubMed]
- Dhugga, K.S.; Barreiro, R.; Whitten, B.; Stecca, K.; Hazebroek, J.; Randhawa, G.S.; Dolan, M.; Kinney, A.J.; Tomes, D.; Nichols, S.; et al. Guar seed beta-mannan synthase is a member of the cellulose synthase super gene family. Science 2004, 303, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Liepman, A.H.; Cavalier, D.M. The CELLULOSE SYNTHASE-LIKE A and CELLULOSE SYNTHASE-LIKE C families: recent advances and future perspectives. Front. Plant Sci. 2012, 3, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocuron, J.C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A gene from the cellulose synthase-like C family encodes a beta-1,4 glucan synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550–8555. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Gao, G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, P.; Li, Y.; Zhao, L.; Hou, Z.; Yan, M.; Hu, B.; Liu, Y.; Azam, S.M.; Zhang, Z.; Rahman, Z.U. Genome-Wide Identification and Expression Profiling of ATP-Binding Cassette (ABC) Transporter Gene Family in Pineapple (Ananas comosus(L.) Merr.) Reveal the Role ofAcABCG38 in Pollen Development. Front. Plant Sci. 2017, 8, 2150. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr | Length | Amino Acids | Introns | Exons | Isoelectric Point (pI) | Molecular Weight (MW) |
|---|---|---|---|---|---|---|---|---|
| AcoCesA1 | Aco014283 | LG05 | 7376 | 1104 | 13 | 14 | 6.72 | 122.4 |
| AcoCesA2 | Aco018229 | LG01 | 8424 | 1077 | 13 | 14 | 7.92 | 120.7 |
| AcoCesA3 | Aco024230 | LG09 | 12,555 | 1081 | 12 | 13 | 8.31 | 119.6 |
| AcoCesA4 | Aco014585 | LG08 | 7332 | 1070 | 11 | 12 | 7.61 | 119.9 |
| AcoCesA5 | Aco018552 | LG06 | 6358 | 1102 | 13 | 14 | 7.04 | 123.2 |
| AcoCesA7 | Aco012076 | LG02 | 6278 | 1062 | 12 | 13 | 6.33 | 118.9 |
| AcoCesA8 | Aco006039 | LG16 | 6951 | 1003 | 12 | 13 | 5.93 | 111.7 |
| AcoCslA1 | Aco001096 | LG02 | 14,534 | 1126 | 19 | 19 | 8.99 | 127.2 |
| AcoCslA2 | Aco006900 | LG22 | 4546 | 536 | 8 | 9 | 9.19 | 60.9 |
| AcoCslA3 | Aco001095 | LG02 | 5559 | 560 | 9 | 10 | 8.8 | 62.9 |
| AcoCslA4 | Aco004149 | LG15 | 20,041 | 1184 | 20 | 20 | 8.8 | 133.1 |
| AcoCslA6 | Aco002889 | LG06 | 9648 | 555 | 15 | 9 | 8.56 | 63 |
| AcoCslA9 | Aco016682 | LG17 | 9750 | 607 | 10 | 10 | 9.16 | 68.4. |
| AcoCslA11 | Aco014689 | LG20 | 7773 | 568 | 9 | 10 | 7.83 | 64.1 |
| AcoCslC1 | Aco004974 | LG07 | 3939 | 727 | 4 | 5 | 8.4 | 81.1 |
| AcoCslC4 | Aco000968 | LG02 | 2630 | 673 | 4 | 5 | 8.85 | 77.1 |
| AcoCslC5 | Aco008242 | LG19 | 4607 | 747 | 8 | 6 | 8.86 | 84.7 |
| AcoCslC6 | Aco013494 | LG20 | 5582 | 709 | 4 | 5 | 8.67 | 78.7 |
| AcoCslC8 | Aco011603 | LG06 | 3136 | 663 | 4 | 5 | 8.99 | 74.8 |
| AcoCslC12 | Aco008598 | LG09 | 3881 | 773 | 6 | 7 | 6.15 | 85.7 |
| AcoCslD1 | Aco004435 | LG05 | 4309 | 1068 | 4 | 5 | 7.62 | 116.9 |
| AcoCslD2 | Aco015969 | LG22 | 5627 | 1174 | 4 | 3 | 7.49 | 129.2 |
| AcoCslD3 | Aco030607 | scaffold_1556 | 5631 | 1131 | 4 | 3 | 6.74 | 124.7 |
| AcoCslD4 | Aco017129 | LG14 | 6362 | 1159 | 6 | 5 | 6.46 | 127.5 |
| AcoCslD5 | Aco013738 | LG13 | 3953 | 1201 | 2 | 3 | 8.45 | 130.9 |
| AcoCslD6 | Aco016995 | LG08 | 4094 | 1193 | 4 | 5 | 7.9 | 130.5 |
| AcoCslD7 | Aco025070 | LG01 | 7403 | 1170 | 3 | 4 | 6.7 | 128.9 |
| AcoCslD8 | Aco017291 | LG03 | 3892 | 1185 | 2 | 3 | 7.08 | 130.2 |
| AcoCslE1 | Aco026335 | LG19 | 5869 | 680 | 7 | 8 | 6.99 | 76.2 |
| AcoCslE2 | Aco000884 | LG02 | 4750 | 736 | 7 | 8 | 8.02 | 81.9 |
| AcoCslG1 | Aco013153 | LG24 | 47,982 | 1615 | 16 | 17 | 7.92 | 178.6 |
| AcoCslJ1 | Aco013513 | LG20 | 13,869 | 807 | 9 | 10 | 8.91 | 89.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, S.; Cheng, H.; Zhang, J.; Aslam, M.; Yan, M.; Hu, A.; Lin, L.; Ojolo, S.P.; Zhao, H.; Priyadarshani, S.V.G.N.; et al. Genome-Wide Identification, Expression Pattern Analysis and Evolution of the Ces/Csl Gene Superfamily in Pineapple (Ananas comosus). Plants 2019, 8, 275. https://doi.org/10.3390/plants8080275
Cao S, Cheng H, Zhang J, Aslam M, Yan M, Hu A, Lin L, Ojolo SP, Zhao H, Priyadarshani SVGN, et al. Genome-Wide Identification, Expression Pattern Analysis and Evolution of the Ces/Csl Gene Superfamily in Pineapple (Ananas comosus). Plants. 2019; 8(8):275. https://doi.org/10.3390/plants8080275
Chicago/Turabian StyleCao, Shijiang, Han Cheng, Jiashuo Zhang, Mohammad Aslam, Maokai Yan, Anqi Hu, Lili Lin, Simon Peter Ojolo, Heming Zhao, S.V.G.N. Priyadarshani, and et al. 2019. "Genome-Wide Identification, Expression Pattern Analysis and Evolution of the Ces/Csl Gene Superfamily in Pineapple (Ananas comosus)" Plants 8, no. 8: 275. https://doi.org/10.3390/plants8080275