Characteristics and Expression Pattern of MYC Genes in Triticum aestivum, Oryza sativa, and Brachypodium distachyon


State Key Laboratory of Crop Stress Biology for Arid Areas, College of Agronomy, Northwest A&F University, Yangling 712000, China
*
Authors to whom correspondence should be addressed.
Plants 2019, 8(8), 274; https://doi.org/10.3390/plants8080274
Submission received: 29 June 2019
/
Revised: 3 August 2019
/
Accepted: 6 August 2019
/
Published: 8 August 2019
(This article belongs to the Section Plant Genetics, Genomics and Biotechnology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Myelocytomatosis oncogenes (MYC) transcription factors (TFs) belong to basic helix-loop-helix (bHLH) TF family and have a special bHLH_MYC_N domain in the N-terminal region. Presently, there is no detailed and systematic analysis of MYC TFs in wheat, rice, and Brachypodium distachyon. In this study, 26 TaMYC, 7 OsMYC, and 7 BdMYC TFs were identified and their features were characterized. Firstly, they contain a JAZ interaction domain (JID) and a putative transcriptional activation domain (TAD) in the bHLH_MYC_N region and a BhlH region in the C-terminal region. In some cases, the bHLH region is followed by a leucine zipper region; secondly, they display tissue-specific expression patterns: wheat MYC genes are mainly expressed in leaves, rice MYC genes are highly expressed in stems, and B. distachyon MYC genes are mainly expressed in inflorescences. In addition, three types of cis-elements, including plant development/growth-related, hormone-related, and abiotic stresses-related were identified in different MYC gene promoters. In combination with the previous studies, these results indicate that MYC TFs mainly function in growth and development, as well as in response to stresses. This study laid a foundation for the further functional elucidation of MYC genes.
1. Introduction
bHLH (basic helix-loop-helix) family is the second largest plant transcription factor (TF) family. They are characterized by a bHLH domain, which consists of 50–60 amino acids that form two distinctive regions, the basic region and HLH (helix-loop-helix) region [1]. The basic region functions as a DNA-binding motif, while the HLH region forms two amphipathic α helices with a linking loop [1,2,3]. In the bHLH family, there are some special members, named myelocytomatosis oncogenes (MYC). These members are characterized by a so-called bHLH_MYC_N region in the N-terminal and a bHLH region in the C-terminal [1,2,4,5].
MYC genes function in various physiological and molecular processes, especially in growth and development. In Arabidopsis thaliana, MYC2 functions synergistically with MYC3 and MYC4 in regulating leaf senescence [6], root elongation [7], stamen development [8], seed production, and seed storage protein accumulation [8,9], as well as chlorophyll degradation [10]; other Arabidopsis MYC genes, like ALCATRAZ functions in cell separation in fruit dehiscence [11], SPATULA controls development of carpel margin tissues [12], and ABORTED MICROSPORES (AMS) plays a crucial role in tapetum cell development and pollen wall formation [13]. In rice (Oryza sativa), the orthologous AMS gene Tapetum Degeneration Retardation (TDR) is necessary for tapetum degradation and anther development [14]; OsMYC2 is expressed in all tissues, and highly expressed in the spikelets and floral organs [15]. It regulates spikelet development through the interaction with OsJAZ1 and the activation of the downstream gene OsMADS1 [16].
MYC genes are also involved in plant secondary metabolism. For example, overexpression of Arabidopsis MYC3 and MYC4 result in more anthocyanin accumulation [17], and MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis [18]. Wheat (Triticum aestivum) MYC1 and barley (Hordeum vulgare) Myc1 (HvAnt2) regulate anthocyanin synthesis in pericarp too [19,20]. Additionally, NbMYC2 regulates alkaloid biosynthesis [21], and SmMYC2 regulates the phenolic acid biosynthesis in Salvia miltiorrhiza [22], while TcJAMYC1/2/4 negatively regulate the expression of genes associating with paclitaxel biosynthesis in Taxus cuspidate [23].
Meanwhile, MYC genes play important roles in response to abiotic and biotic stresses. For example, the expression of AtMYC2 is induced by drought and salt stresses [24], and overexpressing this gene improves osmotic stress tolerance [25]. Arabidopsis MYC67 and MYC70 interact with ICE1 and negatively regulate cold tolerance in Arabidopsis [26]. Additionally, rice OsMYC2 negatively regulates JA-mediated resistance to a necrotrophic pathogen [27,28].
Especially, several MYC genes were reported to associate with JA (jasmonate) signal. Arabidopsis MYC2, MYC3, MYC4, and MYC5 are master regulators in JA signal pathway [17,27,29,30]. Arabidopsis JASMONATE-INSENSITIVE1 is essential for jasmonate-regulated defense responses [27]. In addition to regulating spikelet development via the JA signal [16], OsMYC2 mediates numerous defense-related transcriptional changes via JA signaling [31].
Wheat (T. aestivum) and rice (O. sativa) are important cereal crops, and B. distachyon is a grass model plant and has a close genetic relationship with wheat [32]. In this study, TaMYC, OsMYC, and BdMYC genes were characterized at the genome-wide level, and the expression patterns were analyzed. Meanwhile, the specific domain of MYC proteins and the cis-elements in the promoters were identified. Based on these results, we summarized the characteristics of MYC TFs. This study laid a foundation for further functional elucidation of MYC genes.
2. Results
2.1. Wheat, Rice, and B. distachyon MYC TFs
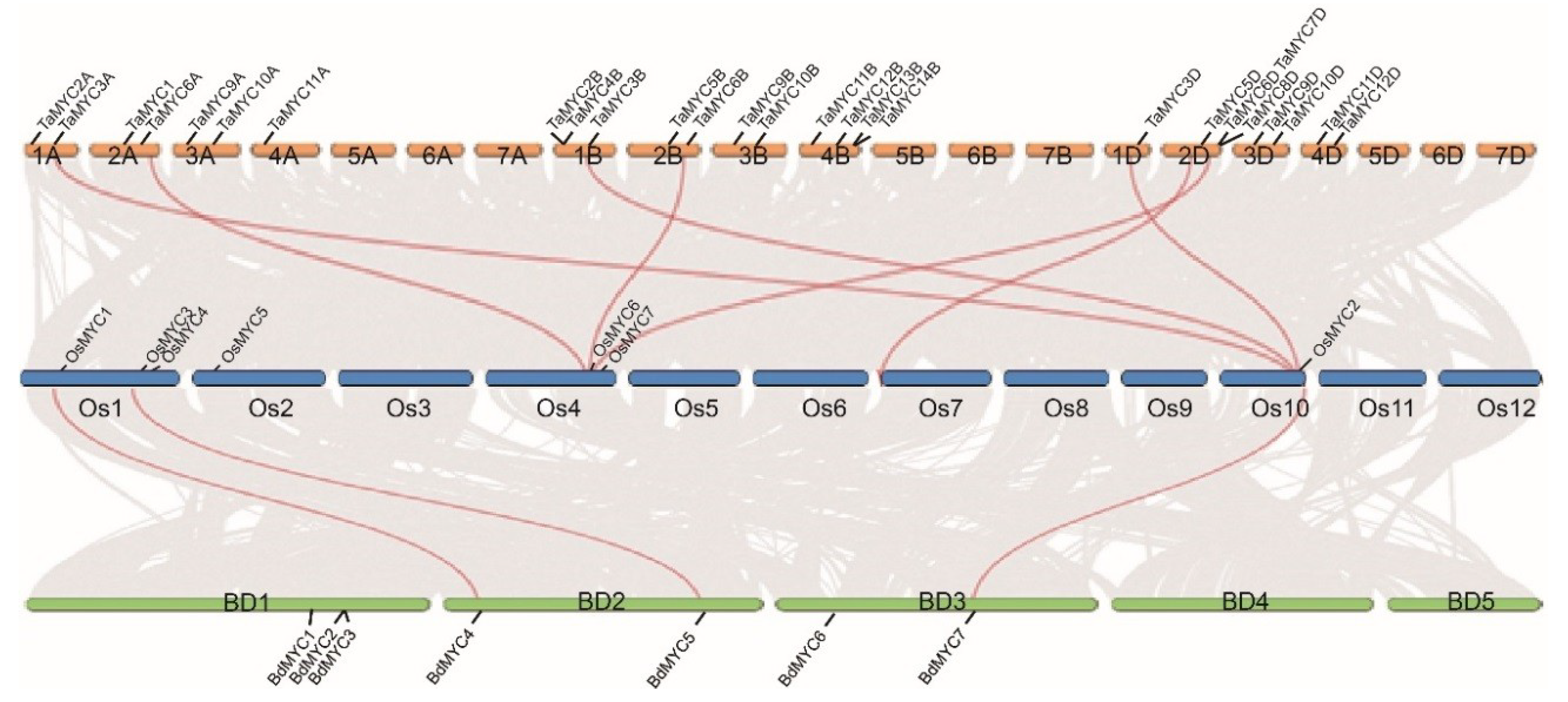
Totally, 26 putative TaMYC, 7 OsMYC, and 7 BdMYC genes were identified in the relevant genomes, respectively (Figure 1), including two reported genes: TaMYC1 and OsMYC2 [15,19]. Among these 40 genes, 34 genes were verified by ESTs (Expressed Sequence Tags) deposited in the NCBI database, and 18 TaMYC genes constitute 6 sets, every set includes three homologous genes in A, B, and D subgenomes, respectively.
Except for TaMYC1 and OsMYC2, other 25 TaMYC genes were named as TaMYC2A to TaMYC12B according to their distribution on chromosomes and genomic homology; other 6 OsMYC and 7 BdMYC genes were named as OsMYC1, OsMYC3 to OsMYC7, and BdMYC1 to BdMYC7 based on their chromosomal localization (Figure 1).
The physical features of these MYC TFs were predicted. In wheat, the protein length varies from 456 (TaMYC7A) to 699 (TaMYC4D) amino acids; the PI (Isoelectric Point) varies from 4.96 (TaMYC6D) to 8.73 (TaMYC9D); and the molecular weight varies from 49.17 kDa (TaMYC9A) to 63.69 kDa (TaMYC7D). In rice, the protein length varies from 473 (OsMYC7) to 904 (OsMYC4) amino acids; the PI varies from 4.68 (OsMYC5) to 8.36 (OsMYC3); and the molecular weight varies from 50.67 kDa (OsMYC3) to 97.39 kDa (OsMYC4). In B. distachyon, the protein length varies from 470 (BdMYC5) to 706 (BdMYC7); the PI varies from 4.53 (BdMYC6) to 7.22 (BdMYC5); and the molecular weight varies from 50.43 kDa (BdMYC5) to 76.24 kDa (BdMYC7). The protein subcellular localization was predicted by CELLO web server [33] and results showed that 26 MYC proteins locate in the nucleus, while other 14 MYC proteins locate in the chloroplast, cytoplasm, mitochondria. The detailed information is listed in Table S1.
2.2. Sequence Alignment and Phylogenetic Tree of MYC TFs
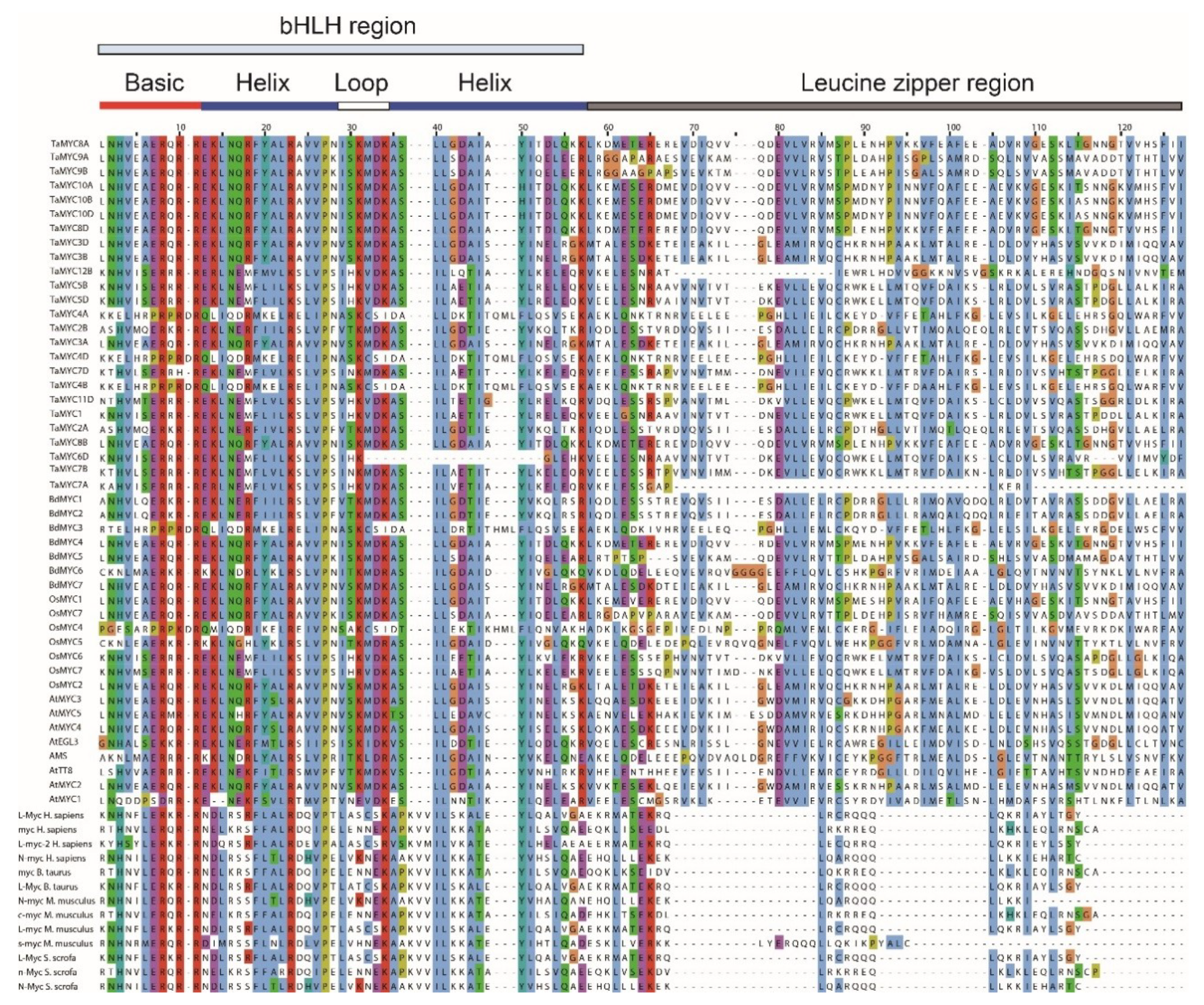
Based on previous studies [13,18,34,35,36,37,38], protein sequences of 13 known animal MYCs and 8 known Arabidopsis MYCs obtained from GenBank [39], and sequences of 40 MYCs in this study were used to perform sequence alignment. Multiple sequence alignment identified a distinct N-terminal bHLH_MYC_N region (Figure S1). As shown in Table S2, the consensus ratio of 56 conserved amino acid residues is more than 50% in the bHLH_MYC_N domains. Leu-100, Trp-113, Tyr-115, Trp-119, and Leu-147 are identical in these 40 MYC TFs, indicating that these amino acids are key component of bHLH_MYC_N domain. More importantly, one JAZ interaction domain (JID) and one putative transcriptional activation domain (TAD) were identified in the bHLH_MYC_N domain. Furthermore, a bHLH domain and a leucine zipper region were found in the C-terminal region (Figure 2 and Figure S1).
In animals, MYC proteins are key regulators of mammalian cell proliferation that activate genes as part of a heterodimeric complex with the protein Max [40]. As shown in Figure S2, the alignment of 6 known Max proteins (Genbank number: NP_660092.1 [Homo sapiens], XP_002613458.1 [Branchiostoma floridae], XP_008195890.1 [Tribolium castaneum], NP_510223.1 [Caenorhabditis elegans], NP_001099103.1 [Bos taurus], NP_002373.3 [Homo sapiens]) and other MYC proteins showed that they are highly conserved in bHLH domains, while poorly conserved in leucine zipper region.
According to the alignment of bHLH domains in rice and Arabidopsis bHLH proteins [41], some amino acids residues, such as Val-960 and Ala-962 in the basic region, Asn-972 and Val-981 in the first Helix, Ser-986, Lys-987, Met-1006, and Asp-1008 in the Loop, and Asp-1017 in the second Helix, are only found in plant MYC proteins (Figure 2 and Table S3). In addition, although the bHLH-ZIP domain is commonly present in some human c-Myc and Max proteins [42], such as myc_H. sapiens, myc_B. taurus, c-myc_M. musculus, n-Myc_S. scrofa in Figure 2, they were lowly conserved in the leucine zipper region.
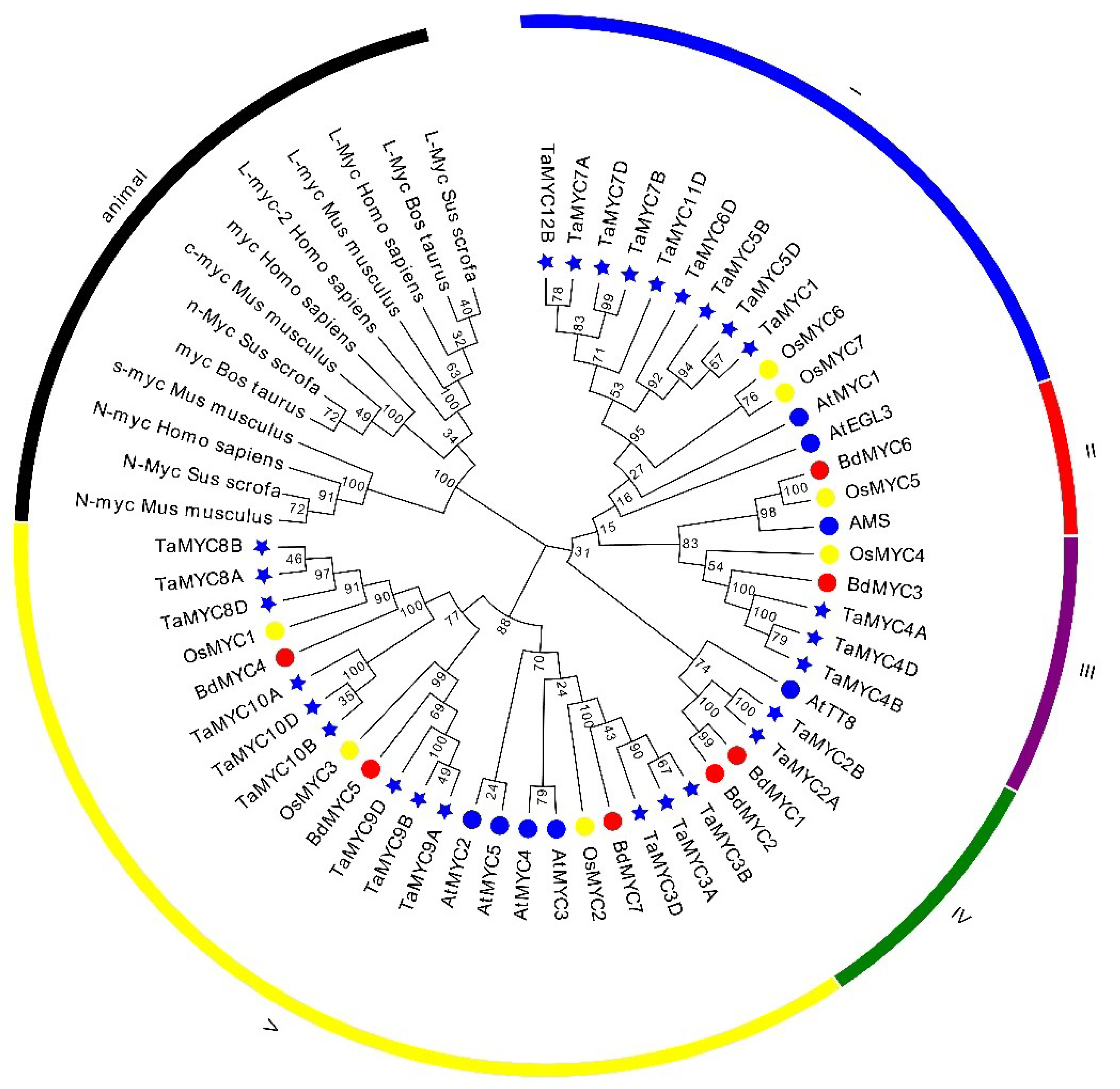
To understand the evolutionary relationships of MYC TFs, a neighbor-joining (NJ) phylogenetic tree was constructed based on the full-length alignment of 40 putative MYCs, 13 known animal MYCs, and 8 known Arabidopsis MYC. As shown in Figure 3, these MYC TFs are classified into six sub-groups with highly bootstrap values. The class V is the largest while the class II was the smallest. There are 9 TaMYCs, 2 OsMYCs, and 2 AtMYCs in class I; 1 OsMYC, 1 BdMYC, and 1 AtMYC in class II; 3 TaMYCs, 1 OsMYC, and 1 BdMYC in class III; 2 TaMYCs, 2 BdMYCs, 1 AtMYC in class IV; 12 TaMYCs, 3 OsMYCs, 3 BdMYCs, and 4 AtMYCs in class V; and 13 known animal MYCs in class “animal”.
2.3. Gene Structures and Conserved Motifs
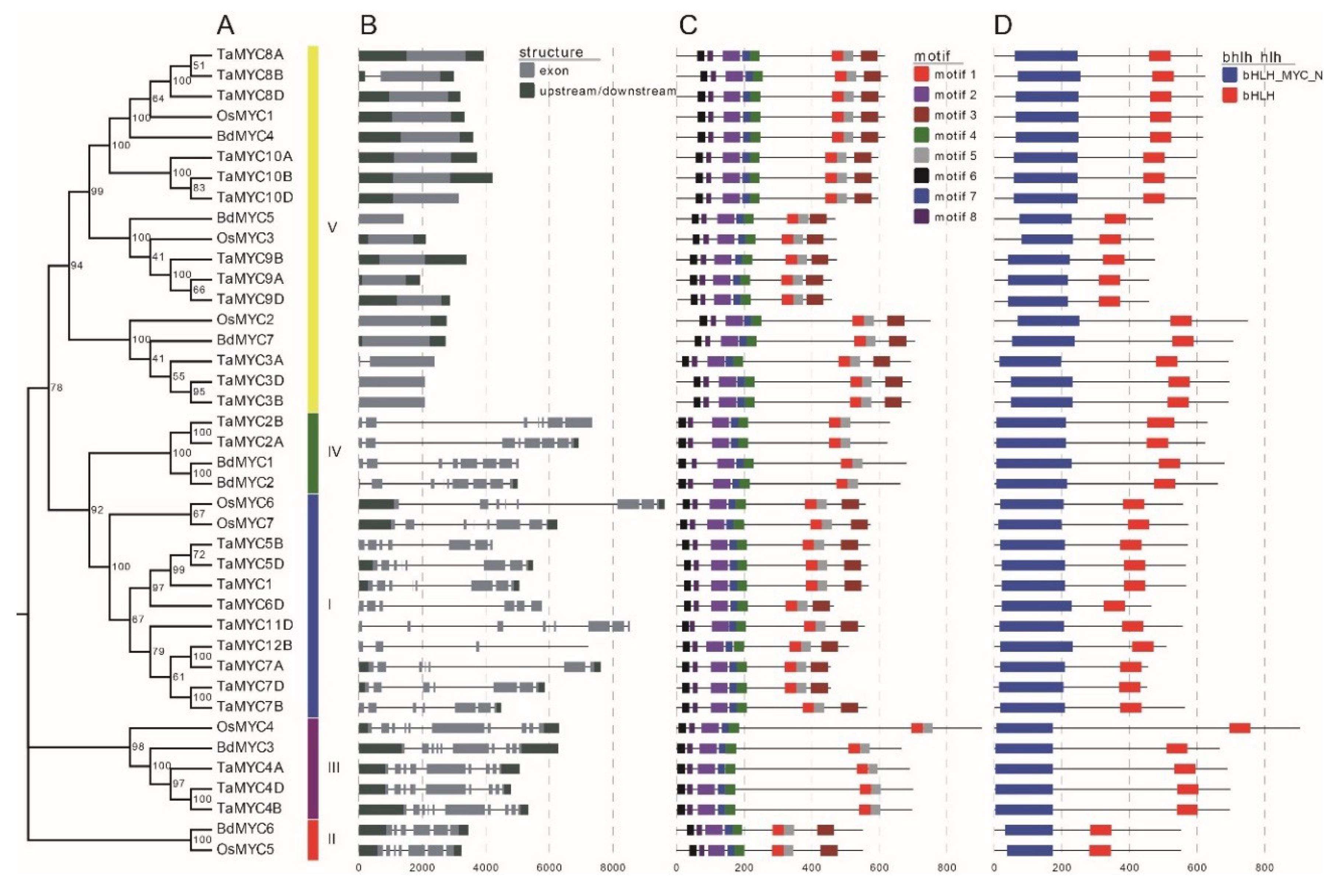
We used genomic DNA sequences and CDS (coding sequences) to analyze the gene structure (Figure 4B). Gene structures of TaMYC, OsMYC, and BdMYC genes are similar within the same subgroup. In wheat, the exon number ranges from 1 to 11. In rice and B. distachyon, the exon number ranges from 1 to 10. Notably, most class V members only have one exon.
Conserved motifs are helpful to understand the functions of MYC TFs. In this study, 8 conserved motifs were identified (Figure 4C). Motifs 2, 4, 6, 7, and 8 constitute the bHLH_MYC_N domain (Figure 4D). Among these five motifs, motifs 6 and 8 form JID and motif 2 composes TAD. JID consists of approximately 90 amino acids, including a specific motif (W-[TN]-Y-[AG]-[IVL]-[FYL]-W-X(6,19)-L-[GT]-W-[GK]-[DE]-G). TAD includes approximately 70 amino acids and a specific motif ([VL]-[TG]-[DEG]-[TA]-E-[WML]-[FY]-[FY]-X(2)-[SC]-[MA]-X(3)-F-X(4)-G-[LAG]-P-G-X(9)-W).
2.4. Synteny and Homologous Gene Pairs
Gene duplication events include tandem duplication and segmental duplication. We analyzed the gene duplication by using the MCScanX software [44]. As shown in Table S4, only one segmental duplication pair (TaMYC6D-TaMYC7D) was found.
In addition, orthologous between MYC genes in wheat, B. distachyon, A. thaliana, rice, H. vulgare, Sorghum bicolor, and Zea mays were also investigated (Table S5). A total of 12, 0, 19, 7, 22, and 28 orthologs and orthologous gene pairs between wheat and B. distachyon, A. thaliana, rice, H. vulgare, S. bicolor, and Z. mays (Table S5); 2, 5, 4, 6, and 7 orthologs and orthologous gene pairs between B. distachyon and A. thaliana, rice, H. vulgare, S. bicolor, and Z. mays; 6, 3, 3, and 2 orthologs and orthologous gene pairs between rice and A. thaliana, S. bicolor, H. vulgare, and Z. mays. These results suggested that MYC genes of monocots have strong relationships.
2.5. Identified Cis-Elements in MYC Gene Promoters
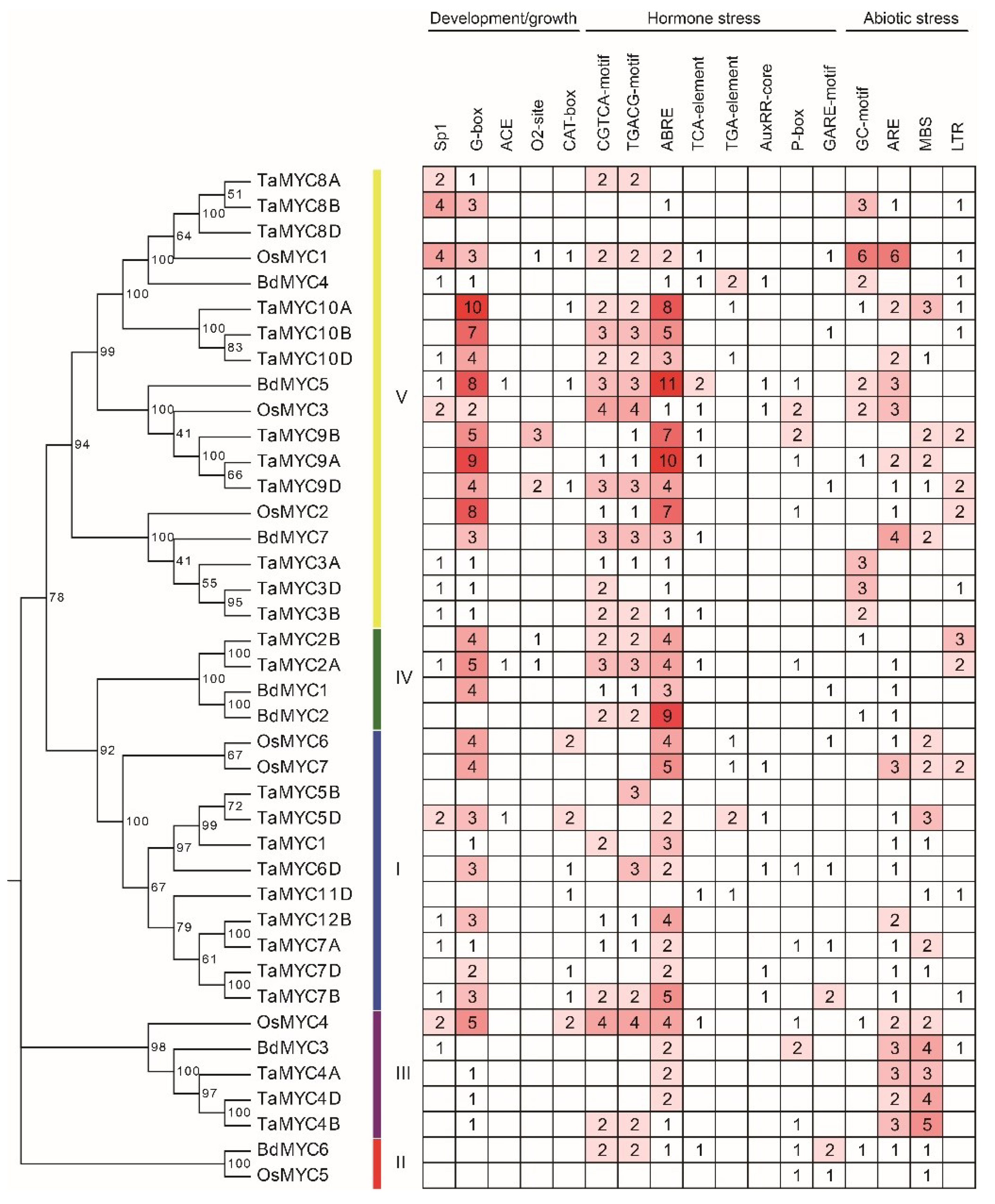
We also identified the cis-elements in the 2-kb promoters of MYC genes using the PlantCARE web tool [45]. As shown in Figure 5, three types of cis-elements, which are related to plant growth/development, hormone, and abiotic stresses, were identified. The cis-elements related to growth/development include light-responsive element G-box (CACGTC) [46] and Sp1 (GGGCGG) [47], the metabolism regulation related cis-element O2-site [48], and the meristem expression related cis-element CAT-box [49]. The cis-elements related to hormones include the methyl jasmonate (MeJA)-responsive elements CGTCA-motif [50] and TGACG-motif [51], the abscisic acid (ABA)-responsive element ABRE (ACGTG) [52], the gibberellin(GA)-responsive element GARE-motif [53], the auxin-responsive elements TGA-element [54] and AuxRR-core (GGTCCAT) [55], and the salicylic acid (SA)-responsive element TCA-element (CCATCTTTTT) [56]. The cis-elements associated with abiotic stresses include adaptive elements such as drought-inducibility element MBS (CAACTG) [57], low-temperature responsiveness element LTR (CCGAAA) [58], anaerobic induction ARE (AAACCA) [59], and anoxic specific inducibility element GC-motif (A/CGCCGCGCA) [60] were detected in a series of members. Combined with the phylogenetic tree, these results showed that the phylogenetically similar genes shared identical cis-elements.
2.6. Expression Profiles of MYC Genes
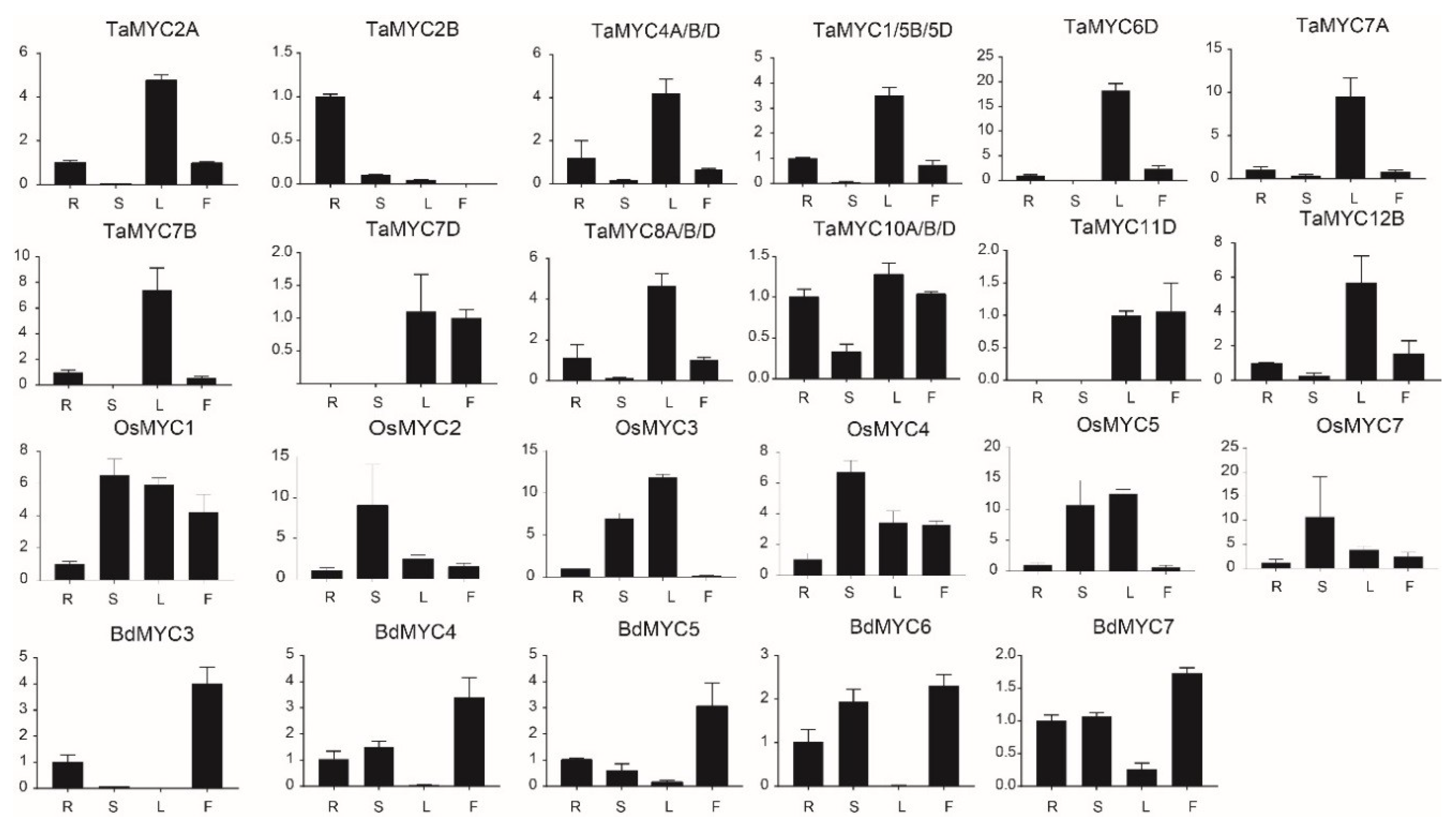
We analyzed the expression patterns of TaMYC, OsMYC, and BdMYC genes by quantitative real-time polymerase chain reaction (qRT-PCR). Among these 40 MYC genes, the expression of 9 genes (TaMYC3A/B/D, TaMYC9A/B/D, OsMYC6, BdMYC1, and BdMYC2) was not detected in all cases. As shown in Figure 6, MYC genes displayed tissue-specific expression. A total of 11 TaMYC genes (including TaMYC2A, TaMYC4A/B/D, TaMYC1/5B/5D, TaMYC6D, TaMYC7A, TaMYC7B, TaMYC7D, TaMYC8A/B/D, TaMYC10A/B/D, TaMYC11D, and TaMYC12B) were highly expressed in leaves, 4 OsMYC genes (OsMYC1, OsMYC2, OsMYC4, and OsMYC7) were highly expressed in stems, and 5 BdMYC genes (BdMYC3, BdMYC4, BdMYC5, BdMYC6, and BdMYC7) were highly expressed in inflorescences.
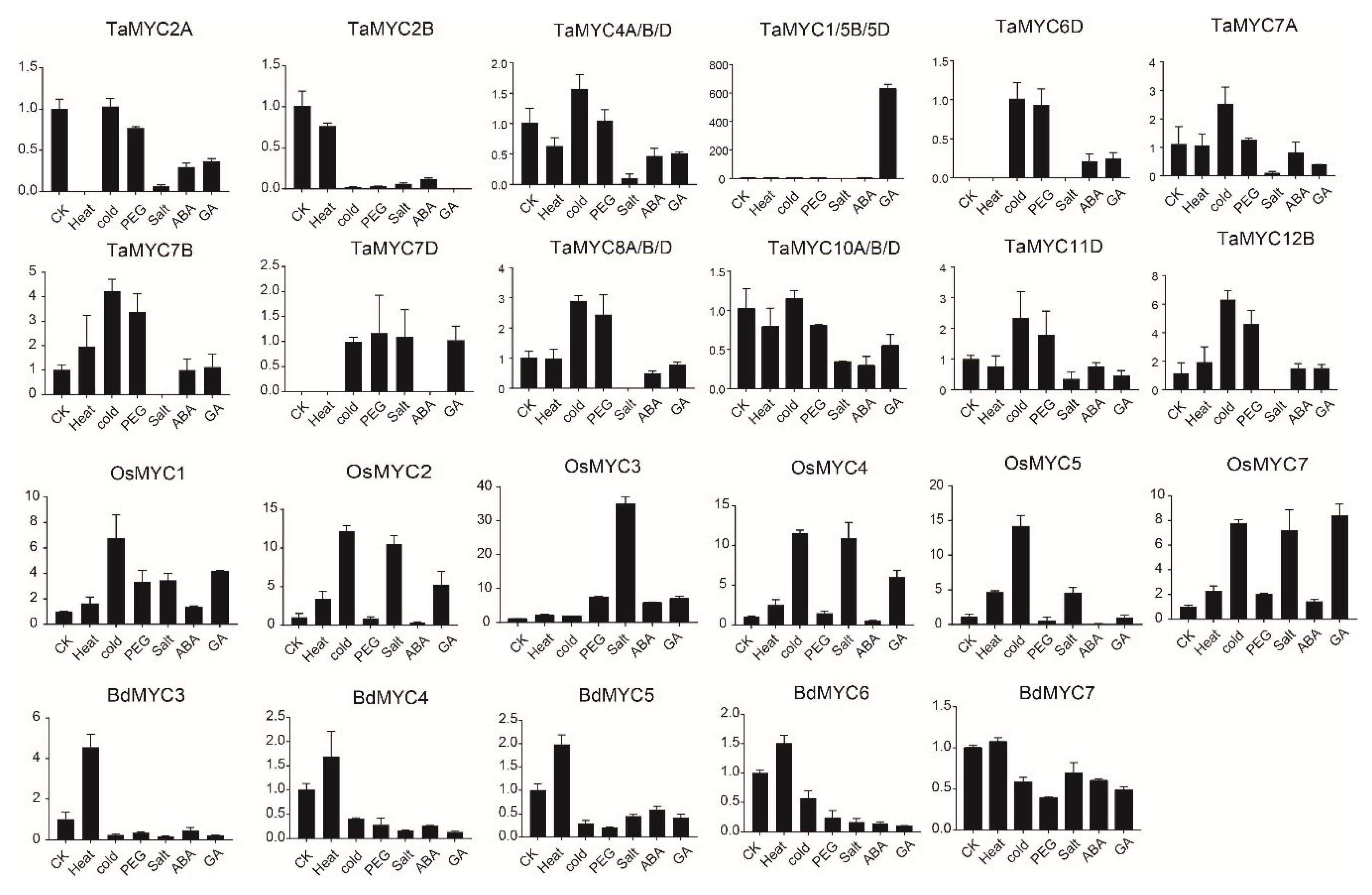
Additionally, we also analyzed the expression of these genes in two-week seedlings with different treatments (Figure 7). Although the results showed no regularity, there are some meaningful findings. In wheat, the expression of TaMYC1/5B/5D is induced by GA (Gibberellin), indicating the involvement in GA signal. TaMYC7A, TaMYC7B, and TaMYC7D displayed different expression patterns, implying that their functions are diversified. In rice, the expression of six expressed genes is upregulated by different abiotic stresses, suggesting their important roles. In B. distachyon, BdMYC3 is drastically induced by heat.
3. Discussion
3.1. The Characteristics of MYCs
bHLH TFs are the second largest class of plant TFs [61]. Plant MYC TFs belong to the bHLH superfamily. MYC proteins have one bHLH_MYC_N domain in the N-terminal region [1]. In this domain, Leu-100, Trp-113, Tyr-115, Trp-119, and Leu-147 amino acids residues are complete conserved in all TaMYC, BdMYC, and OsMYC proteins. The bHLH_MYC_N include two domains: JID and TAD. The former is necessary for interacting with JAZ proteins, while the latter is a putative transcriptional activation domain [2]. For example, Arabidopsis MYC2, MYC3, and MYC4 interact with the C-terminal JAS domain of JAZ proteins through JID, and AtMYC2 recruits the mediator complex required for transcription initiation through its TAD, which specifically interacts with the activator interaction domain [62,63]. We also identified one specific motif (W-[TN]-Y-[AG]-[IVL]-[FYL]-W-X(6,19)-L-[GT]-W-[GK]-[DE]-G) in JID and one specific motif ([VL]-[TG]-[DEG]-[TA]-E-[WML]-[FY]-[FY]-X(2)-[SC]-[MA]-X(3)-F-X(4)-G-[LAG]-P-G-X(9)-W) in TAD.
The C-terminal region of TaMYCs, BdMYCs, and OsMYCs is conserved and includes the typical basic region and HLH domain, and most MYCs contain leucine zipper (bZIP) domain (Figure 2 and Figure S2). Previous reports showed that the basic region of MYC proteins mainly recognizes CACGTG (G-box) sequence and CATGTG sequence, both E-box DNA-binding sites (5′-CANNTG-3′), and the bHLH domain are required for the heterodimer to bind to the G-box in target genes [2]. The bHLH-ZIP domain is present in human c-Myc and Max proteins [42], which have DNA-binding activity and has been predicted to mediate protein–protein interactions. The interaction of Max and c-Myc depends on the integrity of the c-Myc HLH-Zip domain, but not the basic region or other sequences outside the domain [42]. Different from animal MYCs, plants MYCs contain a longer (about 70 amino acids) leucine zipper. Furthermore, the C-terminal leucine zipper domain can form the dimer that affects the specificity of interaction with other TFs [64]. Compared to other bHLH proteins in plants [43,65], bHLH-ZIP domain only appeared on group B which MYC and Max exist.
Compared to bHLH genes, MYC genes have three special kinds of cis-elements in their gene promoters. These cis-elements associate with their special functions. Moreover, MYC genes show tissue-specific expression patterns. Additionally, different from other non-MYC-bHLH genes which have 2 or more exons [41,43,65], most class V MYC members, which is the largest class only, have one exon.
3.2. Functions of MYC TFs
Previous studies showed that MYC TFs function in plant development and growth [2,9,37]. For example, MYC proteins function as regulators in regulating plant seed production, root elongation, leaf senescence, and stamen development [6,7,8,9]; they also regulate plant secondary metabolism and are actively involved in hormone-mediated plant growth [21,66,67]. In this study, most MYC genes are expressed in roots, stems, leaves, and inflorescences (Figure 6). In combination with the identification of many cis-elements related to plant growth and development and hormone stress, these results further suggest their functions in growth and development.
The expression profiles of many MYC genes indicate their probable functions in response to abiotic and biotic stresses. For example, Arabidopsis MYC2, MYC3, MYC4, and MYC5 are induced by jasmonate [17,27,29,30]; MYC3 and MYC4 act additively with MYC2 to regulate defense against insect herbivory [30]; and MYC5 regulates JA-mediated plant defenses against herbivores and is involved in JA-regulated plant resistance to pathogens [37]. Overexpression of OsMYC2 results in bacterial blight resistance in rice [16]. In maize, the expression of two MYC genes, ZmbHLH103 and ZmbHLH104, is significantly upregulated under drought stress [65]. In this study, the expression of 6 OsMYC genes is induced by different abiotic stresses (Figure 7), further indicating that MYC genes participate in response to abiotic stresses. Consistently, many cis-elements related to abiotic stresses were identified.
Taken together, we can draw a conclusion that MYC TFs mainly function in growth/development, and in response to environmental stresses.
4. Materials and Methods
4.1. Identification of MYC TFs in Wheat, Rice, and B. distachyon
The wheat, rice, and B. distachyon genome sequences, protein sequences, coding sequences (CDS), and upstream 2-kb genomic DNA sequences were downloaded from Ensembl plants [68]. The chromosomal distribution of MYC genes was obtained from the wheat, rice, and B. distachyon genome annotations in Ensembl plants [68]. To identify the MYC TFs in wheat, rice, and B. distachyon, 7 known MYC proteins, including wheat TaMYC1, barley HvMyc1, rice OsMYC2, Arabidopsis MYC2, MYC3, MYC4, and AMS proteins sequences, were downloaded from Ensembl plants [68] and were used to build a hidden Markov model (HMM), then searched against the genome protein sequences of wheat, rice, B. distachyon, respectively, with a threshold of E< 1e-5 and the length of amino acid >200 aa (amino acid). Then, the HMM profile of the bHLH-MYC_N domain (PF14215) was downloaded from the Pfam database [69] to search against the protein sequences of wheat, rice, and B. distachyon with a threshold of E < 1e-5. After manual correction to remove the redundancy and alternative splice, the NCBI-CDD database (NCBI Conserved Domains Database) and SMART database (Simple Modular Architecture Research Tool) were used to confirm the putative MYC proteins. The ExPASy webserver [70] was used to predict the theoretical isoelectric point and molecular weight of TaMYCs, OsMYCs, and BdMYCs. To further verify the existence of MYC genes in wheat, rice, and B. distachyon, we performed BLASTN [71] to search for ESTs using the CDS of MYC genes. The CELLO web server [33] was used to predict the subcellular localization of MYC proteins.
4.2. Multiple Sequence Alignment and Phylogenetic Analysis of MYCs
An unrooted neighbor-joining (NJ) tree was constructed by MEGA 7.0 software with 1000 bootstrap replications and Jones–Taylor–Thornton model based on full-length protein sequence alignments was done by using the ClustalX 2.0 method [72,73]. The multiple sequence alignment was performed using the ClustalX 2.0 and visualized by Jalview [74], and the phylogenetic tree was visualized by Evolview [75].
4.3. Analysis of Gene Structures and Conserved Motifs
The exon–intron structure of MYC genes was graphically displayed by the Gene Structure Display Server [76] by using the CDS and DNA sequences of wheat, rice, and B. distachyon MYC genes. The protein sequences of MYCs were used to predict the conserved motifs by using the MEME online program [77] with the following parameters: optimum width of motifs set from 5 to 200 amino acids and maximum number of motifs set at 8. The gene structure and motifs were visualized by Evolview [75].
4.4. Analysis of Cis-Elements, Gene Duplication, and Synteny
The 2-kb upstream genomic DNA sequences of MYC genes were submitted to the PlantCARE online tool to identify the cis-elements [44]. Gene duplication and systeny analysis of MYCs in different species were done by using the MCScanX software [43] and visualized by Dual Systeny Plotter software by CJ-Chen [78].
4.5. Expression Profile Analysis
The cultivar of T. aestivum ‘Chinese Spring’, rice cultivar ‘Dongjin’, and B. distachyon Bd-21 were planted in an artificial climate chamber at 26/22 °C (day/night) with a photoperiod of 16/8 h (day/night). For tissues analysis, roots, stems, leaves, and inflorescences were collected at the heading stage. For different abiotic stresses, 2-week-old seedling plants were subjected to H2O (CK), heat (42 °C), cold (4 °C), drought (20% PEG6000), salt (200 mM NaCl), ABA (100 μM), and GA (100 μM) for 2 h, then whole plants were collected for RNA isolation. Total RNA was extracted using RNAiso Reagent (TaKaRa, Dalian, China) according to the manufacturer’s protocol and treated with DNaseI. The Transcriptor First Strand cDNA Synthesis Kit (Roche) was used to synthesize cDNA based on the manual. The QuantStudioTM Real-Time PCR Software (ThermoFisher Scientific) was used to carry out qRT-PCR, data acquisition, and analysis. The volume of each reaction was 15 μL containing 7.5 μL of SYBR Premix Ex Taq (TaKaRa, Dalian, China), 0.75 μL (10 pmol μL−1) each of forward and reverse primers, 0.5 μL of cDNA (5.0 ng μL−1), and 5.5 μL of ddH2O. The reference genes (Taactin, Osactin, Bdactin) [79,80,81] were used to normalize the expression of the TaMYC, OsMYC, and BdMYC genes and the 2(−ΔΔCt) analysis method was used to determine the relative expression level [82]. The following program was used for qRT-PCR: 50 °C for 2 min, 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 1 min in the PCR stage and 95 °C for 1 min, 95 °C for 15 s in the melt curve stage. Primers for 6 sets of TaMYC genes (TaMYC3A/B/D, TaMYC4A/B/D, TaMYC1/5B/5D, TaMYC8A/B/D, TaMYC9A/B/D, and TaMYC10A/B/D) were universal in each set, because of the highly conserved sequences in A, B, and D sub-genomes, thus the detected expression is the combination of three copies of homologous genes. The primers are listed in Table S6.
5. Conclusions
In this study, 26 TaMYC, 7 OsMYC, and 7 BdMYCTFs were identified and divided into five groups. The MYC proteins contained a JAZ interaction domain (JID) and a putative transcriptional activation domain (TAD) in the bHLH_MYC_N domain of the N-terminal region, the bHLH domain was found in the C-terminal region, some MYCs followed by a bHLH-ZIP domain. The expression profiles and identified three kinds of cis-elements indicate that MYC TFs function in plant development and in response to environmental stresses. Taken together, our results provide a solid foundation for further structural and functional investigations on MYCs.
Supplementary Materials
The following data are available online at https://www.mdpi.com/2223-7747/8/8/274/s1, Figure S1: The JAZ interaction domain (JID), putative transcriptional activation domain (TAD) were identified in the MYC proteins; Figure S2: The bHLH domain and leucine zipper domain were identified in the MYC proteins; Table S1: The detailed information of TaMYCs, OsMYCs, BdMYCs; Table S2: The consensus ratio of the conserved amino acid residues in the bHLH_MYC_N domains, Table S3: The consensus ratio of the conserved amino acid residues in the bHLH domains of TaMYCs, OsMYCs, and BdMYCs, Table S4: Wheat, rice, and B. distachyon MYC gene duplication events, Table S5: The homologous genes between plant MYC genes, Table S6: Primer used in this study.
Author Contributions
H.L. conceived and designed the project; S.C., H.Z., T.L., and X.N. performed main experiments; H.L. and S.C. analyzed the data. H.L. and S.C. wrote the manuscript. All authors have read and approved the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (31571657). The funding body did not exert influence on the design of the study, and collection, analysis, and interpretation of data, or in writing of the manuscript.
Conflicts of Interest
The authors have declared that they have no conflicts of interest.
References
- Nuno, P.; Liam, D. Origin and diversification of basic-helix-loop-helix proteins in plants. Mol. Biol. Evol. 2010, 27, 862–874. [Google Scholar]
- Kazan, K.; Manners, J.M. MYC2: The Master in Action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledent, V.; Vervoort, M. The basic helix-loop-helix protein family: Comparative genomics and phylogenetic analysis. Genome Res. 2001, 11, 754–770. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Liao, Y.C.; Lv, F.F.; Zhang, Z.; Sun, P.W.; Gao, Z.H.; Hu, K.P.; Sui, C.; Jin, Y.; Wei, J.H. Transcription Factor AsMYC2 Controls the Jasmonate-responsive Expression of ASS1 Regulating Sesquiterpene Biosynthesis in Aquilaria sinensis (Lour.) Gilg. Plant Cell Physiol. 2017, 58, 1924–1933. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Maeda, H.; Oguchi, T.; Yamaguchi, T.; Tanabe, N.; Ebana, K.; Yano, M.; Ebitani, T.; Izawa, T. The Birth of a Black Rice Gene and Its Local Spread by Introgression. Plant Cell 2015, 27, 2401–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiancong, Q.; Jiaojiao, W.; Huang, H.; Bei, L.; Hua, G.; Yule, L.; Susheng, S.; Daoxin, X. Regulation of Jasmonate-Induced Leaf Senescence by Antagonism between bHLH Subgroup IIIe and IIId Factors in Arabidopsis. Plant Cell 2015, 27, 1634–1649. [Google Scholar] [Green Version]
- Gasperini, D.; Chételat, A.; Acosta, I.F.; Goossens, J.; Pauwels, L.; Goossens, A.; Dreos, R.; Alfonso, E.; Farmer, E.E. Multilayered Organization of Jasmonate Signalling in the Regulation of Root Growth. PLoS Genet. 2015, 11, e1005300. [Google Scholar] [CrossRef]
- Tiancong, Q.; Huang, H.; Susheng, S.; Daoxin, X. Regulation of Jasmonate-Mediated Stamen Development and Seed Production by a bHLH-MYB Complex in Arabidopsis. Plant Cell 2015, 27, 1620–1633. [Google Scholar] [Green Version]
- Gao, C.; Qi, S.; Liu, K.; Li, D.; Jin, C.; Li, Z.; Huang, G.; Hai, J.; Zhang, M.; Chen, M. MYC2, MYC3, and MYC4 function redundantly in seed storage protein accumulation in Arabidopsis. Plant Physiol. Biochem. 2016, 108, 63–70. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, J.; Xie, Z.; Gao, J.; Ren, G.; Gao, S.; Zhou, X.; Kuai, B. Jasmonic acid promotes degreening via MYC2/3/4- and ANAC019/055/072-mediated regulation of major chlorophyll catabolic genes. Plant J. 2015, 84, 597–610. [Google Scholar] [CrossRef]
- Rajani, S.; Sundaresan, V. The Arabidopsis myc/bHLH gene ALCATRAZ enables cell separation in fruit dehiscence. Curr. Biol. 2001, 11, 1914–1922. [Google Scholar] [CrossRef] [Green Version]
- Heisler, M.; Atkinson, A.; Bylstra, Y.; Walsh, R.; Smyth, D. SPATULA, a gene that controls development of carpel margin tissues in Arabidopsis, encodes a bHLH protein. Development 2001, 128, 1089–1098. [Google Scholar] [PubMed]
- Anna-Marie, S.; Sandra, K.B.; Unte, U.S.; Peter, H.; Koen, D.; Heinz, S. The Arabidopsis Aborted MicrosporeS (AMS) gene encodes a MYC class transcription factor. Plant J. Cell Mol. Biol. 2010, 33, 413–423. [Google Scholar]
- Na, L.; Da-Sheng, Z.; Hai-Sheng, L.; Chang-Song, Y.; Xiao-Xing, L.; Wan-Qi, L.; Zheng, Y.; Ben, X.; Huang-Wei, C.; Jia, W. The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development. Plant Cell 2006, 18, 2999–3014. [Google Scholar]
- Cai, Q.; Yuan, Z.; Chen, M.; Yin, C.; Luo, Z.; Zhao, X.; Liang, W.; Hu, J.; Zhang, D. Jasmonic acid regulates spikelet development in rice. Nat. Commun. 2014, 5, 3476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uji, Y.; Taniguchi, S.; Tamaoki, D.; Shishido, H.; Akimitsu, K.; Gomi, K. Overexpression of OsMYC2 Results in the Up-Regulation of Early JA-Rresponsive Genes and Bacterial Blight Resistance in Rice. Plant Cell Physiol. 2016, 57, 1814–1827. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Figueroa, P.; Browse, J. Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis. J. Exp. Bot. 2011, 62, 2143–2154. [Google Scholar] [CrossRef] [Green Version]
- Fabian, S.; Patricia, F.C.; Mark, Z.; Monica, D.D.; Sandra, F.; Gaétan, G.; Lewsey, M.G.; Ecker, J.R.; Roberto, S.; Philippe, R. Arabidopsis basic helix-loop-helix transcription factors MYC2, MYC3, and MYC4 regulate glucosinolate biosynthesis, insect performance, and feeding behavior. Plant Cell 2013, 25, 3117–3132. [Google Scholar]
- Zong, Y.; Xi, X.; Li, S.; Chen, W.; Zhang, B.; Liu, D.; Liu, B.; Wang, D.; Zhang, H. Allelic Variation and Transcriptional Isoforms of WheatTaMYC1Gene Regulating Anthocyanin Synthesis in Pericarp. Front. Plant Sci. 2017, 8, 1645. [Google Scholar] [CrossRef]
- Shoeva, O.Y.; Mock, H.P.; Kukoeva, T.V.; Börner, A.; Khlestkina, E.K. Regulation of the Flavonoid Biosynthesis Pathway Genes in Purple and Black Grains of Hordeum vulgare. PLoS ONE 2016, 11, e0163782. [Google Scholar] [CrossRef]
- Todd, A.T.; Enwu, L.; Polvi, S.L.; Pammett, R.T.; Page, J.E. A functional genomics screen identifies diverse transcription factors that regulate alkaloid biosynthesis in Nicotiana benthamiana. Plant J. 2010, 62, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhou, W.; Su, J.; Wang, X.; Li, L.; Wang, L.; Cao, X.; Wang, Z. Overexpression of SmMYC2 Increases the Production of Phenolic Acids in Salvia miltiorrhiza. Front. Plant Sci. 2017, 8, 1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenka, S.K.; Nims, N.E.; Vongpaseuth, K.; Boshar, R.A.; Roberts, S.C.; Walker, E.L. Jasmonate-responsive expression of paclitaxel biosynthesis genes in Taxus cuspidata cultured cells is negatively regulated by the bHLH transcription factors TcJAMYC1, TcJAMYC2, and TcJAMYC4. Front. Plant Sci. 2015, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp.Bot. 2007, 58, 221. [Google Scholar] [CrossRef]
- Ohta, M.; Sato, A.; Na, R. MYC-type transcription factors, MYC67 and MYC70, interact with ICE1 and negatively regulate cold tolerance in Arabidopsis. Sci. Rep. 2018, 8, 11622. [Google Scholar] [CrossRef]
- Lorenzo, O.; Chico JMSanchez-Serrano, J.J.; Solano, R. Jasmonate-insensitive1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Figueroa, P.; Browse, J.; Figueroa, P.; Browse, J. Male sterility in Arabidopsis induced by overexpression of a MYC5-SRDX chimeric repressor. Plant J. 2015, 81, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Calvo, P.; Chini, A.; Fernandez-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef]
- Ogawa, S.; Kawahara-Miki, R.; Miyamoto, K.; Yamane, H.; Nojiri, H.; Tsujii, Y.; Okada, K. OsMYC2 mediates numerous defence-related transcriptional changes via jasmonic acid signalling in rice. Biochem. Biophys. Res. Commun. 2017, 486, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Wusirika, R.; Jorge, D.; Yong-Jin, P.; Carlos, B.; John, E.; Phillip, S.M.; Bennetzen, J.L. Different types and rates of genome evolution detected by comparative sequence analysis of orthologous segments from four cereal genomes. Genetics 2002, 162, 1389. [Google Scholar]
- Yu, C.S.; Chen, Y.C.; Hwang, J.K. Prediction of protein subcellular localization. Proteins-Struct. Funct. Bioinform. 2010, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Simionato, E.; Ledent, V.; Richards, G.; Thomas-Chollier, M.; Kerner, P.; Coornaert, D.; Degnan, B.M.; Vervoort, M. Origin and diversification of the basic helix-loop-helix gene family in metazoans: Insights from comparative genomics. BMC Evol. Biol. 2007, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Ledent, V.; Paquet, O.; Vervoort, M. Phylogenetic analysis of the human basic helix-loop-helix proteins. Genome Biol. 2002, 3, 1–18. [Google Scholar] [CrossRef]
- Zhang, D.; Li, G.; Wang, Y. A genome-wide identification and analysis of basic helix-loop-helix transcription factors in cattle. Gene 2017, 626, 241. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Song, S.; Huang, H.; Wang, J.; Qi, T.; Xie, D. MYC5 is Involved in Jasmonate-Regulated Plant Growth, Leaf Senescence and Defense Responses. Plant Cell Physiol. 2017, 58, 1752–1763. [Google Scholar] [Green Version]
- Urao, T.; Yamaguchishinozaki, K.; Mitsukawa, N.; Shibata, D.; Shinozaki, K. Molecular cloning and characterization of a gene that encodes a MYC-related protein in Arabidopsis. Plant Mol. Biol. 1996, 32, 571–576. [Google Scholar] [CrossRef]
- Benson, A.; GenBank, D. GenBank: update. Nucleic Acids Res. 2004, 32, D23–D26. [Google Scholar] [CrossRef]
- Bouchard, C.; Staller, P.; Eilers, M. Control of Cell Proliferation by Myc. Trends Cell Biol. 1998, 8, 202–206. [Google Scholar] [CrossRef]
- Xiaoxing, L.; Xuepeng, D.; Haixiong, J.; Yujin, S.; Yuanping, T.; Zheng, Y.; Jingkang, G.; Wanqi, L.; Liang, C.; Jingyuan, Y. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol. 2006, 141, 1167. [Google Scholar]
- Blackwood, E.; Eisenman, R. Max: A helix-loop-helix zipper protein that forms a sequence-specific DNA-binding complex with Myc. Science 1991, 251, 1211. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Guan, Y.; Chen, S.; Li, H. Genome-wide analysis of basic helix-loop-helix (bHLH) transcription factors in Brachypodium distachyon. BMC Genom. 2017, 18, 619. [Google Scholar] [CrossRef] [PubMed]
- Yupeng, W.; Haibao, T.; Debarry, J.D.; Xu, T.; Jingping, L.; Xiyin, W.; Tae-Ho, L.; Huizhe, J.; Barry, M.; Hui, G. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar]
- Magali, L.; Patrice, D.; Gert, T.; Kathleen, M.; Yves, M.; Yves, V.D.P.; Pierre, R.; Stephane, R. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar]
- Giuliano, G.; Pichersky, E.; Malik, V.S.; Timko, M.P.; Scolnik, P.A.; Cashmore, A.R. An evolutionarily conserved protein binding sequence upstream of a plant light-regulated gene. Proc. Natl. Acad. Sci. USA 1988, 85, 7089–7093. [Google Scholar] [CrossRef] [PubMed]
- Haidar, M.A.; Henning, D.; Busch, H. Sp1 is essential and its position is important for p120 gene transcription: A 35 bp juxtaposed positive regulatory element enhances transcription 2.5 fold. Nucleic Acids Res. 1991, 19, 6559–6563. [Google Scholar] [CrossRef]
- Carlini, L.E.; Ketudat, M.; Parsons, R.L.; Prabhakar, S.; Schmidt, R.J.; Guiltinan, M.J. The maize EmBP-1 orthologue differentially regulates Opaque2-dependent gene expression in yeast and cultured maize endosperm cells. Plant Mol. Biol. 1999, 41, 339–349. [Google Scholar] [CrossRef]
- Zhang, L.F.; Li, W.F.; Han, S.Y.; Yang, W.H.; Qi, L.W. cDNA cloning, genomic organization and expression analysis during somatic embryogenesis of the translationally controlled tumor protein (TCTP) gene from Japanese larch (Larix leptolepis). Gene 2013, 529, 150–158. [Google Scholar] [CrossRef]
- He, Y.; Gan, S. Identical promoter elements are involved in regulation of the OPR1 gene by senescence and jasmonic acid in Arabidopsis. Plant Mol. Biol. 2001, 47, 595–605. [Google Scholar] [CrossRef]
- Zheng, Z.; Xiaoming, Y.; Yaping, F.; Longfei, Z.; Hantian, W.; Xinchun, L. Overexpression of PvPin1, a Bamboo Homolog of PIN1-Type Parvulin 1, Delays Flowering Time in Transgenic Arabidopsis and Rice. Front. Plant Sci. 2017, 8, 1526. [Google Scholar] [CrossRef] [PubMed]
- Ezcurra, I.; Ellerström, M.; Wycliffe, P.; Stålberg, K.; Rask, L. Interaction between composite elements in the napA promoter: Both the B-box ABA-responsive complex and the RY/G complex are necessary for seed-specific expression. Plant Mol. Biol. 1999, 40, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Sutoh, K.; Yamauchi, D. Two cis-acting elements necessary and sufficient for gibberellin-upregulated proteinase expression in rice seeds. Plant J. 2010, 34, 635–645. [Google Scholar] [CrossRef]
- Liu, Z.-B. Soybean GH3 promoter contains multiple auxin-inducible elements. Plant Cell 1994, 6, 645–657. [Google Scholar] [PubMed]
- Ballas, N.; Wong, L.-M.; Theologis, A. Identification of the Auxin-responsive Element, AuxRE, in the Primary indoleacetic Acid-inducible Gene, PS-IAA4/5, of Pea (Pisum sativum). J. Mol. Biol. 1993, 233, 580–596. [Google Scholar] [CrossRef] [PubMed]
- Goldsbrough, A.P.; Albrecht, H.; Stratford, R. Salicylic acid-inducible binding of a tobacco nuclear protein to a 10 bp sequence which is highly conserved amongst stress-inducible genes. Plant J. Cell Mol. Biol. 2010, 3, 563–571. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Zhao, J.; Hu, Y.; Zhang, T.Z. Isolation of GhMYB9 gene promoter and characterization of its activity in transgenic cotton. Biol. Plant. 2015, 59, 629–636. [Google Scholar] [CrossRef]
- Dunn, M.A.; White, A.J.; Vural, S.; Hughes, M.A. Identification of promoter elements in a low-temperature-responsive gene (blt4.9) from barley (Hordeum vulgare L.). Plant Mol. Biol. 1998, 38, 551–564. [Google Scholar] [CrossRef]
- Geffers, R.; Sell, S.; Cerff, R.; Hehl, R. The TATA box and a Myb binding site are essential for anaerobic expression of a maize GapC4 minimal promoter in tobacco. Biochim. Biophys. Acta 2001, 1521, 120–125. [Google Scholar] [CrossRef]
- Estes, K.S.; Anderson, D.G.; Stoler, D. Anoxic induction of a sarcoma virus-related VL30 retrotransposon is mediated by a cis-acting element which binds hypoxia-inducible factor 1 and an anoxia-inducible factor. J. Virol. 1995, 69, 6335–6341. [Google Scholar] [Green Version]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. Cell Mol. Biol. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Amoutzias, G.D.; Robertson, D.L.; Peer, Y.V.; Oliver, S.G. Choose your partners: Dimerization in eukaryotic transcription factors. Trends Biochem. Sci. 2008, 33, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Jiang, H.; Li, L.; Zhai, Q.; Qi, L.; Zhou, W.; Liu, X.; Li, H.; Zheng, W.; Sun, J.; et al. The Arabidopsis Mediator Subunit MED25 Differentially Regulates Jasmonate and Abscisic Acid Signaling through Interacting with the MYC2 and ABI5 Transcription Factors. Plant Cell. 2012, 24, 2898–2916. [Google Scholar] [CrossRef] [PubMed]
- Baxevanis, A.D.; Vinson, C.R. Interactions of coiled coils in transcription factors: Where is the specificity? Curr. Opin. Genet. Dev. 1993, 3, 278–285. [Google Scholar] [CrossRef]
- Wei, K.; Chen, H. Comparative functional genomics analysis of bHLH gene family in rice, maize and wheat. BMC Plant Biol. 2018, 18, 309. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Ping, X.G.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Nasser, S.; Schenk, P.M.; Manners, J.M. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar]
- Hongtao, Z.; Sabah, H.; Grégory, M.; Yanxia, Z.; Guillaume, C.; Martial, P.; Pascal, G.; Johan, M. The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 2011, 67, 61–71. [Google Scholar]
- Dan, M.B.; Staines, D.M.; Perry, E.; Kersey, P.J. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomic Data. Methods Mol. Biol. 2017, 1533, 1. [Google Scholar]
- Finn, R.D.; John, T.; Jaina, M.; Coggill, P.C.; Stephen John, S.; Hans-Rudolf, H.; Goran, C.; Kristoffer, F.; Eddy, S.R.; Sonnhammer, E.L.L. The Pfam protein families database. Nucleic Acids Res. 2008, 36, 281–288. [Google Scholar] [CrossRef]
- Panu, A.; Manohar, J.; Konstantin, A.; Delphine, B.; Gabor, C.; Edouard, D.C.; Séverine, D.; Volker, F.; Arnaud, F.; Elisabeth, G. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597. [Google Scholar]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; Mcgettigan, P.; Mcwilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lopez, R.; et al. Clustal W and clustal X version 2.0. Bioinformatics 2007, 3, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Procter, J.; Martin, D.A.; Barton, G.J. Jalview: Visualization and Analysis of Molecular Sequences, Alignments, and Structures. BMC Bioinform. 2005, 6, 1. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296. [Google Scholar] [CrossRef]
- Bailey, T.L.; Nadya, W.; Chris, M.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, 369–373. [Google Scholar] [CrossRef]
- Liu, C.; Xie, T.; Chen, C.; Luan, A.; Long, J.; Li, C.; Ding, Y.; He, Y. Genome-wide organization and expression profiling of the R2R3-MYB transcription factor family in pineapple (Ananas comosus). BMC Genom. 2017, 18, 503. [Google Scholar] [CrossRef]
- Hong, S.Y.; Seo, P.J.; Yang, M.-S.; Xiang, F.; Park, C.-M. Exploring valid reference genes for gene expression studies inBrachypodium distachyonby real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Kamran, M.; Sullivan, W.; Chirkova, L.; Okamoto, M.; Degryse, F.; McLauchlin, M.; Gilliham, M.; Tyerman, S.D. Aluminium-Activated Malate Transporters Can Facilitate GABA Transport. Plant Cell 2018, 30, 1147–1164. [Google Scholar] [CrossRef]
- Kim, B.R.; Nam, H.-Y.; Kim, S.-U.; Kim, S.-I.; Chang, Y.-J. Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice. Biotechnol. Lett. 2003, 25, 1869–1872. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔ C T Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chromosome location, homologous gene pairs of TaMYC, OsMYC, and BdMYC genes. Gray lines in the background indicate the collinear blocks within wheat, rice, and B. distachyon genomes, while the red lines highlight the homologous gene pairs.
Figure 1.
Chromosome location, homologous gene pairs of TaMYC, OsMYC, and BdMYC genes. Gray lines in the background indicate the collinear blocks within wheat, rice, and B. distachyon genomes, while the red lines highlight the homologous gene pairs.

Figure 2.
Alignment of the 61 different myelocytomatosis oncogenes (MYC) proteins from plants and animals. Including 40 MYCs in this study, 8 Arabidopsis MYCs, and 13 animal MYCs proteins from previous studies as described above.
Figure 2.
Alignment of the 61 different myelocytomatosis oncogenes (MYC) proteins from plants and animals. Including 40 MYCs in this study, 8 Arabidopsis MYCs, and 13 animal MYCs proteins from previous studies as described above.

Figure 3.
Phylogenetic tree of 61 MYC transcription factors (TFs). There are 48 plant MYCs and 13 known animal MYCs. The Ta, Os, Bd, and At represent wheat, rice, B. distachyon, and Arabidopsis, respectively.
Figure 3.
Phylogenetic tree of 61 MYC transcription factors (TFs). There are 48 plant MYCs and 13 known animal MYCs. The Ta, Os, Bd, and At represent wheat, rice, B. distachyon, and Arabidopsis, respectively.

Figure 4.
(A) Phylogenetic relationships, (B) gene structures, (C) motifs, and (D) conserved regions of wheat, rice, and B. distachyon MYCs. The tree was constructed with 1000 bootstrap replications using MEGA7 based on the full-length protein sequence. The exon–intron structure of these genes was graphically displayed by the Gene Structure Display Server using the coding sequences (CDS) and DNA sequences of MYC genes. The protein sequence of MYC proteins was used to predict the conserved motifs/region by using the MEME Suite web server.
Figure 4.
(A) Phylogenetic relationships, (B) gene structures, (C) motifs, and (D) conserved regions of wheat, rice, and B. distachyon MYCs. The tree was constructed with 1000 bootstrap replications using MEGA7 based on the full-length protein sequence. The exon–intron structure of these genes was graphically displayed by the Gene Structure Display Server using the coding sequences (CDS) and DNA sequences of MYC genes. The protein sequence of MYC proteins was used to predict the conserved motifs/region by using the MEME Suite web server.

Figure 5.
Main cis-elements in TaMYC, OsMYC, and BdMYC gene promoters.

Figure 6.
Expression patterns of TaMYC, OsMYC, and BdMYC genes in different tissues. R, S, L, F indicate roots, stems, leaves, and inflorescences. The horizontal and vertical coordinates stand for four different tissues and the relative expression, respectively.
Figure 6.
Expression patterns of TaMYC, OsMYC, and BdMYC genes in different tissues. R, S, L, F indicate roots, stems, leaves, and inflorescences. The horizontal and vertical coordinates stand for four different tissues and the relative expression, respectively.

Figure 7.
Expression patterns of TaMYC, OsMYC, and BdMYC genes under abiotic stresses; CK (H2O), Heat, Cold, PEG, Salt, ABA, GA indicate different treatments. The horizontal and vertical axes stand for different abiotic stresses and the relative expression, respectively.
Figure 7.
Expression patterns of TaMYC, OsMYC, and BdMYC genes under abiotic stresses; CK (H2O), Heat, Cold, PEG, Salt, ABA, GA indicate different treatments. The horizontal and vertical axes stand for different abiotic stresses and the relative expression, respectively.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, S.; Zhao, H.; Luo, T.; Liu, Y.; Nie, X.; Li, H. Characteristics and Expression Pattern of MYC Genes in Triticum aestivum, Oryza sativa, and Brachypodium distachyon. Plants 2019, 8, 274. https://doi.org/10.3390/plants8080274
AMA Style
Chen S, Zhao H, Luo T, Liu Y, Nie X, Li H. Characteristics and Expression Pattern of MYC Genes in Triticum aestivum, Oryza sativa, and Brachypodium distachyon. Plants. 2019; 8(8):274. https://doi.org/10.3390/plants8080274
Chicago/Turabian StyleChen, Shoukun, Hongyan Zhao, Tengli Luo, Yue Liu, Xiaojun Nie, and Haifeng Li. 2019. "Characteristics and Expression Pattern of MYC Genes in Triticum aestivum, Oryza sativa, and Brachypodium distachyon" Plants 8, no. 8: 274. https://doi.org/10.3390/plants8080274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.