Oxidative Damage and Antioxidant Defense in Sesamum indicum after Different Waterlogging Durations

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
2.2. Relative Water Content Measurement
2.3. Photosynthetic Pigments Measurement
2.4. Measurement of Proline Content
2.5. Measurement of Lipid Peroxidation
2.6. Determination of Hydrogen Peroxide Content
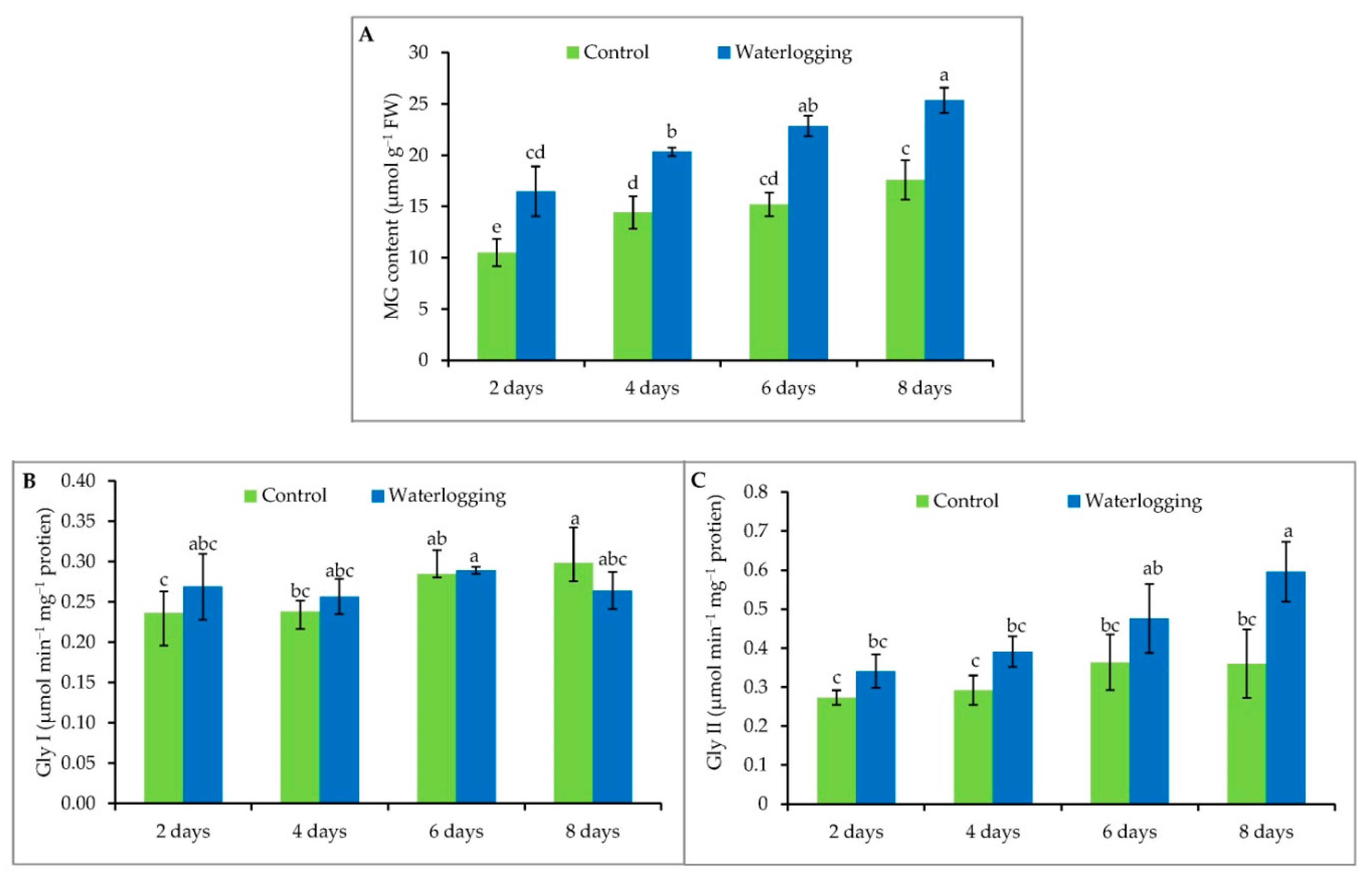
2.7. Measurement of Methylglyoxal Level
2.8. Extraction and Measurement of Ascorbate and Glutathione
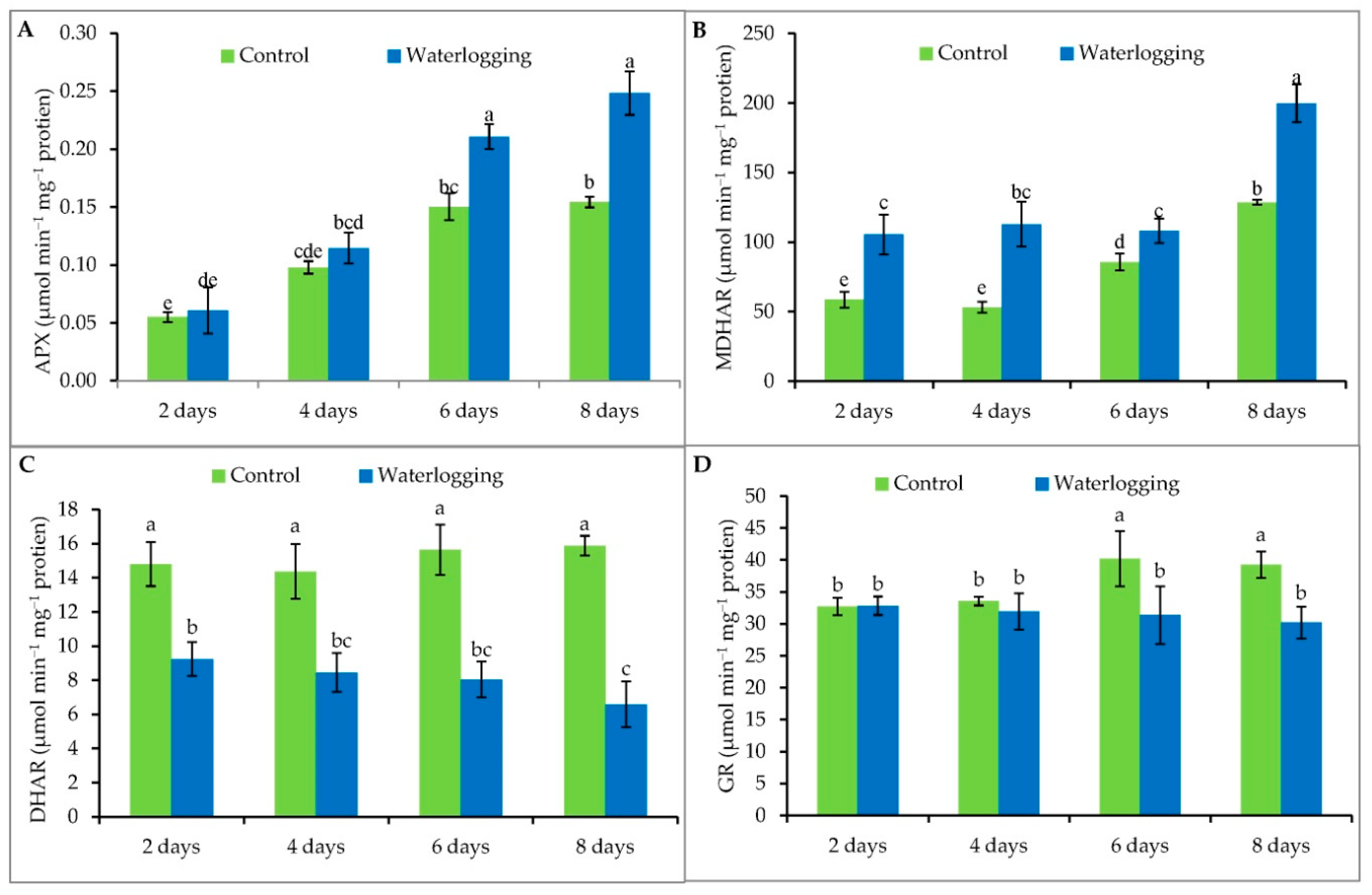
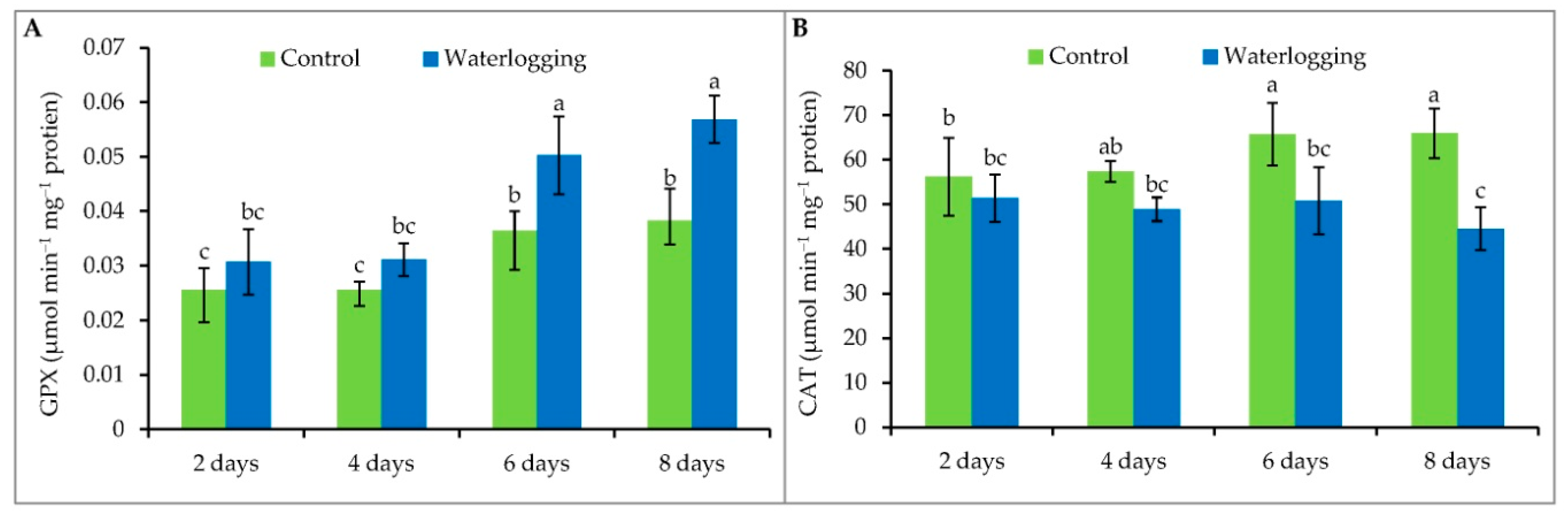
2.9. Enzyme Extraction and Assays
2.10. Statistical Analysis
3. Results
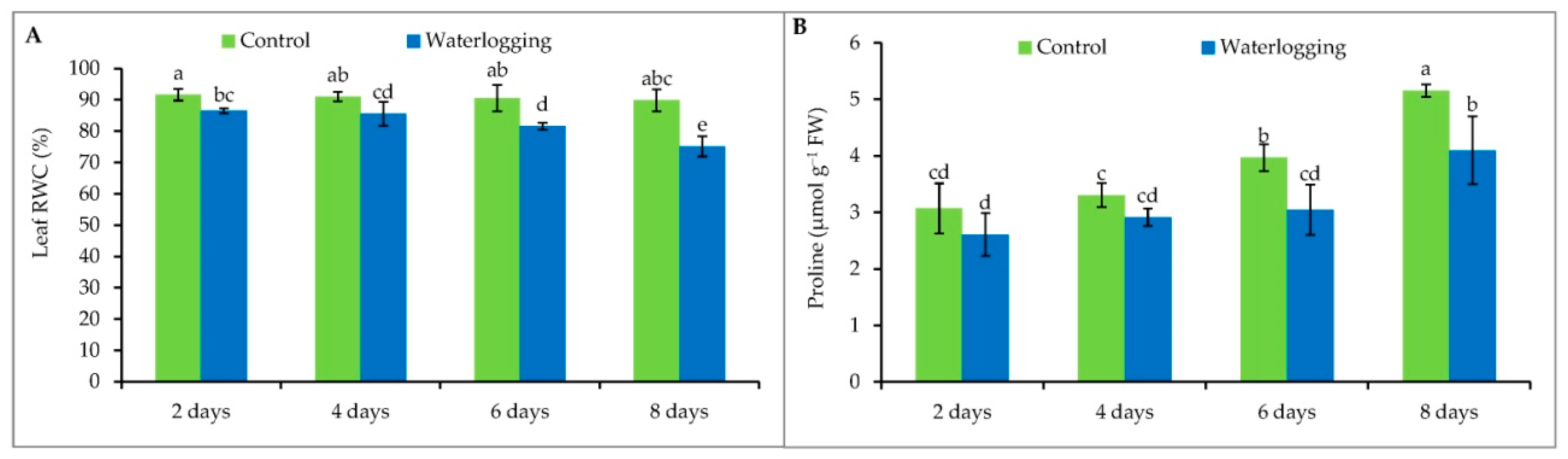
3.1. Leaf Relative Water Content
3.2. Proline Content
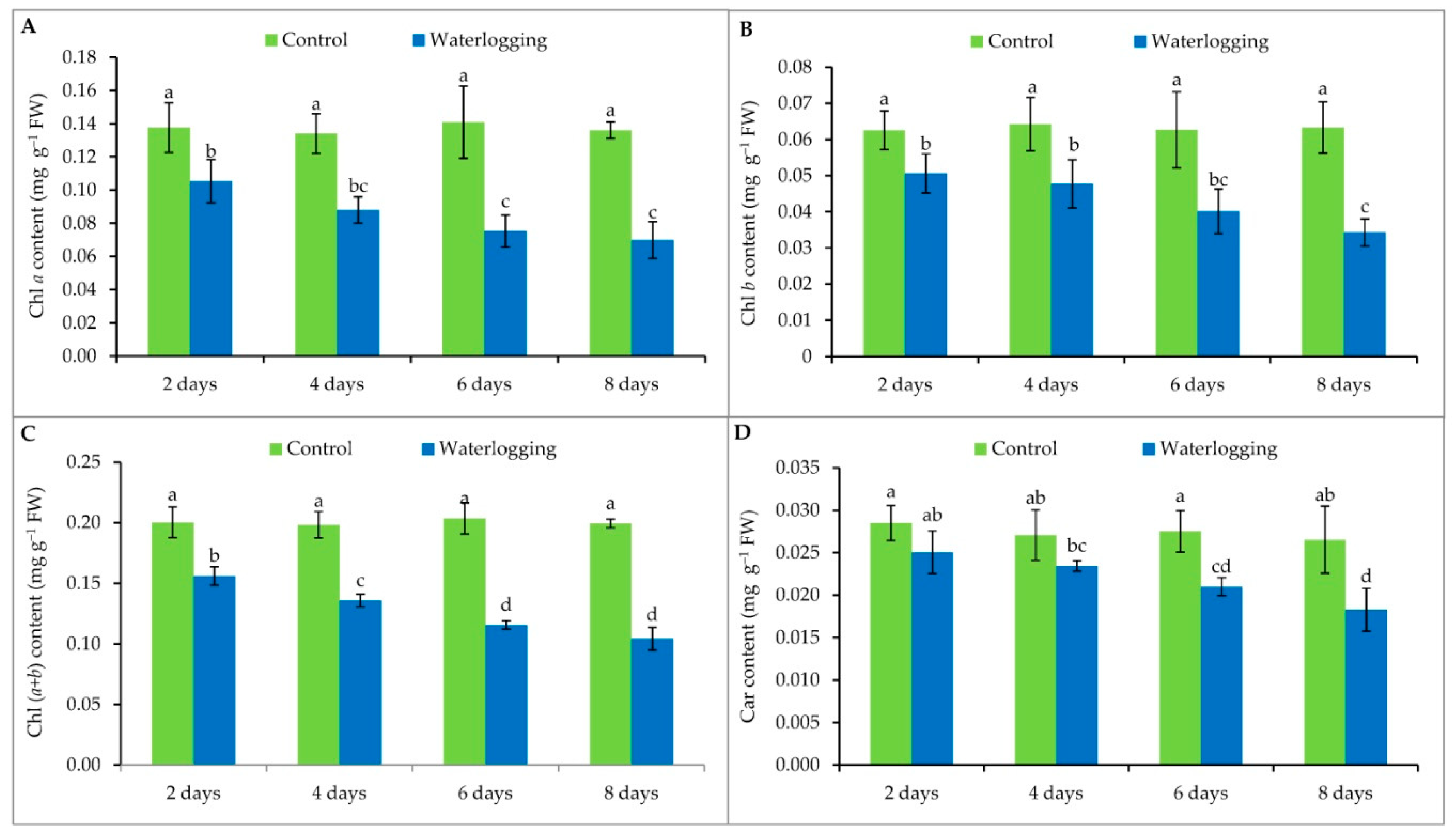
3.3. Chlorophyll Content
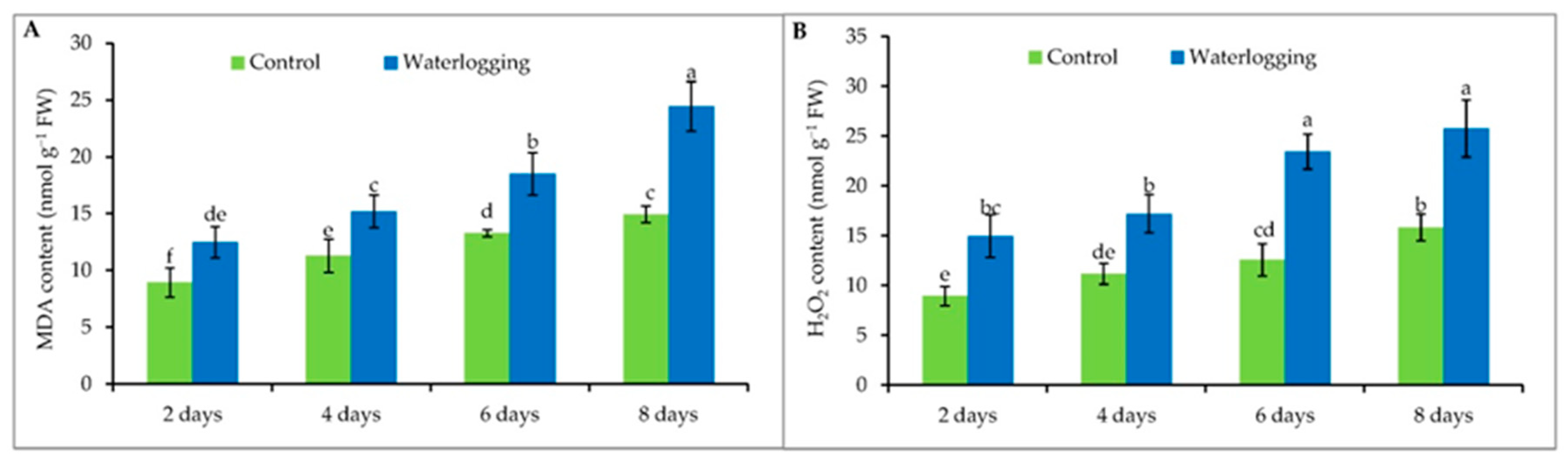
3.4. Lipid Peroxidation and Hydrogen Peroxide Content
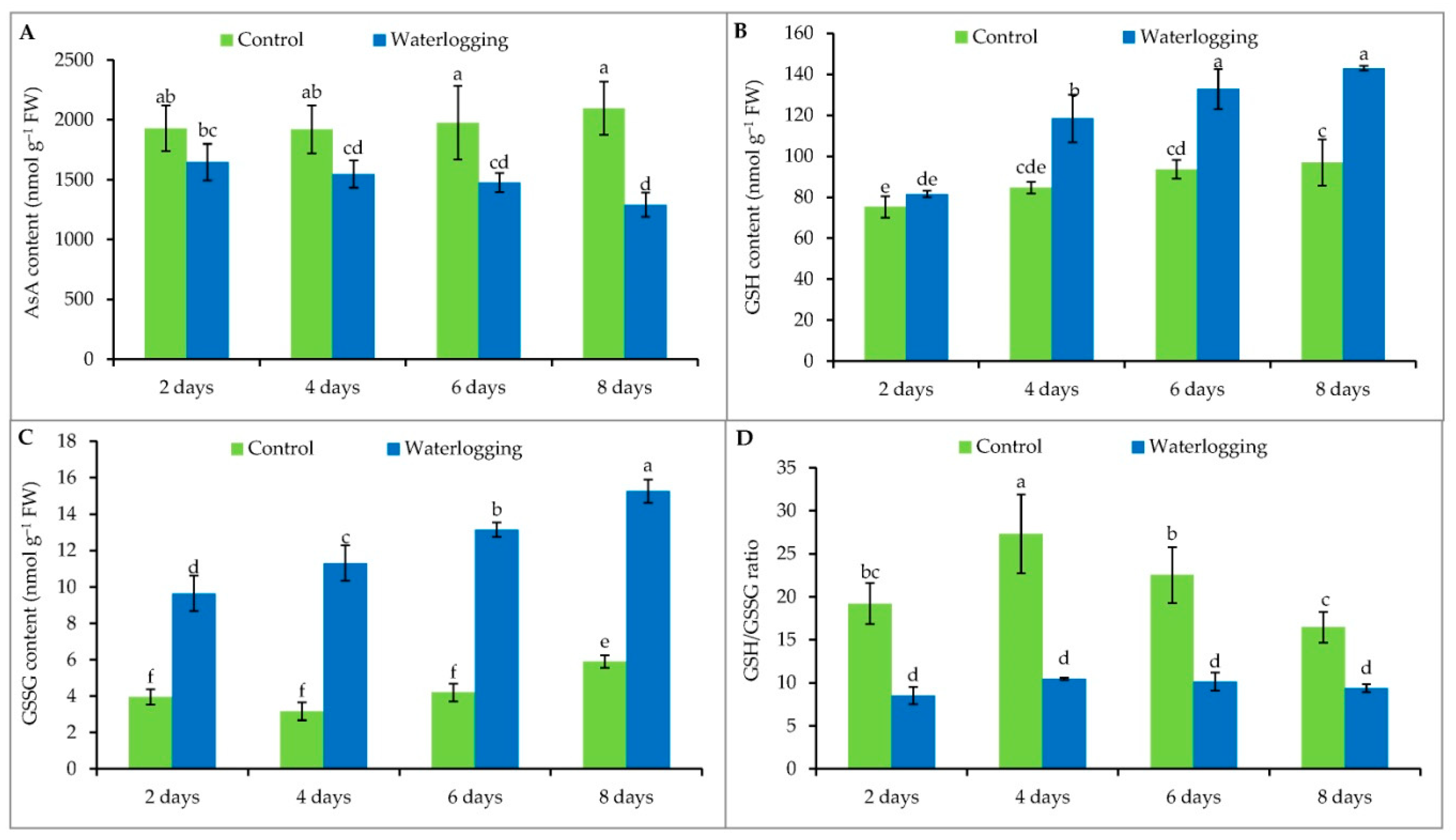
3.5. Ascorbate and Glutathione Contents
3.6. Antioxidant Enzyme Activity
3.7. Methylglyoxal Detoxification
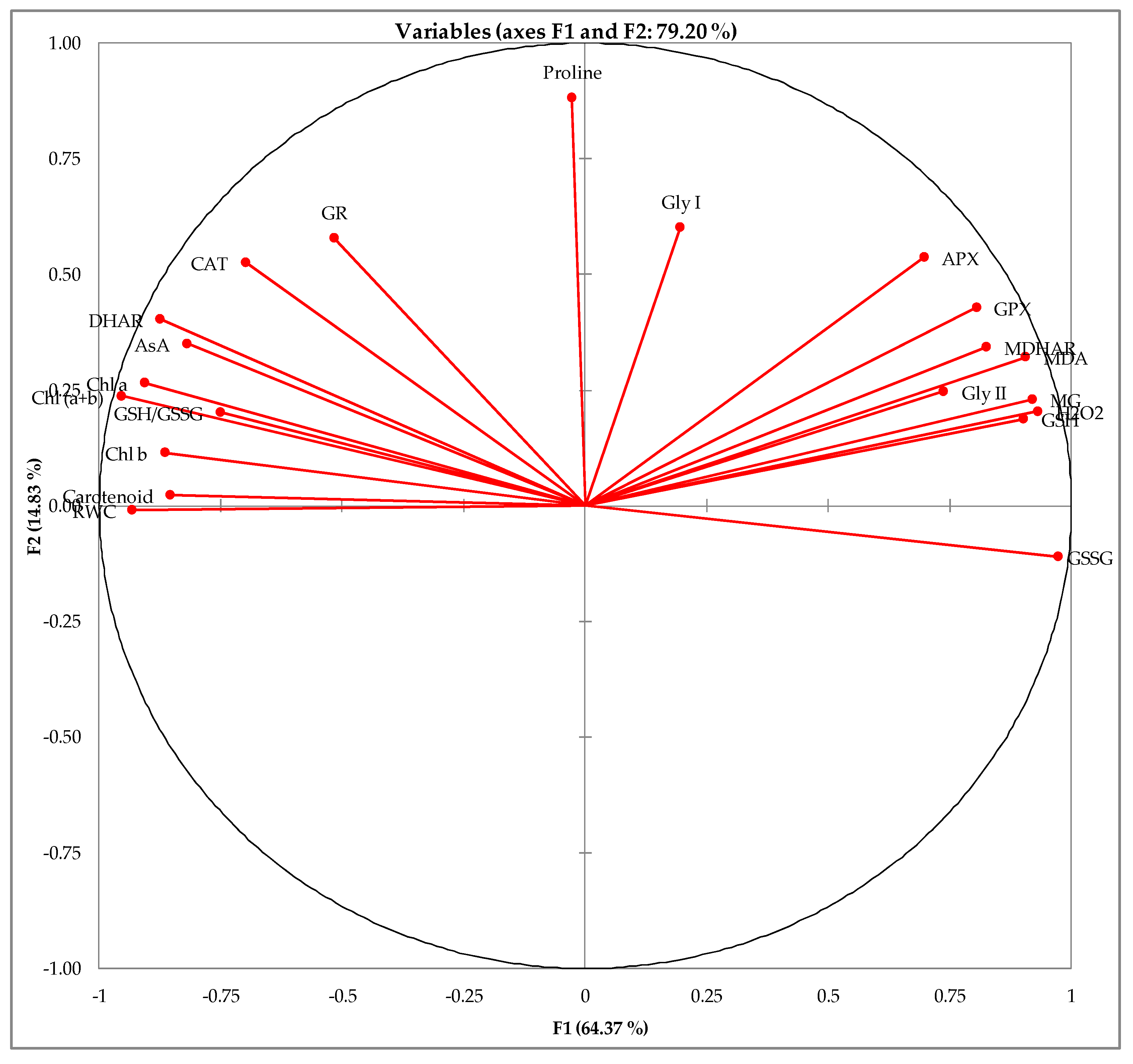
3.8. Correlation among the Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Hasanuzzaman, M.; Hossain, M.A.; Teixeira da Silva, J.A.; Fujita, M. Plant responses and tolerance to abiotic oxidative stress: Antioxidant defense is a key factor. In Crop Stress and its Management: Perspectives and Strategies; Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, M., Eds.; Springer: Berlin, Germany, 2012; pp. 261–316. [Google Scholar]
- NDA. 2016. Available online: http://www.n-d-a.org/flooding.php (accessed on 22 August 2016).
- Setter, T.L.; Waters, I.; Sharma, S.K.; Singh, K.N.; Kulshreshtha, N.; Yaduvanshi, N.P.S.; Ram, P.C.; Singh, B.N.; Rane, J.; McDonald, G.; et al. Review of wheat improvement for waterlogging tolerance in Australia and India: the importance of anaerobiosis and element toxicities associated with different soils. Ann. Bot. 2009, 103, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Arru, L.; Fornaciari, S. Root oxygen deprivation and leaf biochemistry in trees. In Waterlogging Signalling and Tolerance in Plants; Mancuso, S., Shabala, S., Eds.; Springer: Berlin, Germany, 2010; pp. 197–213. [Google Scholar]
- Hasanuzzaman, M.; Mahmud, J.A.; Nahar, K.; Anee, T.I.; Inafuku, M.; Oku, H.; Fujita, M. Responses, Adaptation, and ROS Metabolism in Plants Exposed to Waterlogging Stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation Under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: New York, NY, USA, 2017; pp. 267–281. [Google Scholar]
- Capon, S.J.; James, C.S.; Williams, L.; Quinn, G.P. Responses to flooding and drying in seedlings of a common Australian desert floodplain shrub: Muehlenbeckia florulenta Meisn. Environ. Exp. Bot. 2009, 66, 178–185. [Google Scholar] [CrossRef]
- Gambrell, R.P.; Patrick, W.H. Chemical and microbiological properties of anaerobic soils and sediments. In Plant Life in Anaerobic Environments; Hook, D.D., Crawford, R.M.M., Eds.; CRC Press: Boca Raton, FL, USA, 1978; pp. 375–423. [Google Scholar]
- Else, M.A.; Coupland, D.; Dutton, L.; Jackson, M.B. Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure and slows leaf expansion in flooded plants of castor oil (Riccinus communis) despite diminished delivery of ABA from the roots to shoots in the xylem sap. Physiol. Plant. 2001, 111, 46–54. [Google Scholar] [CrossRef]
- Chang, W.P.; Huang, L.; Shen, M.; Webster, C.; Burlingame, A.L.; Roberts, J.K. Protein synthesis and tolerance of anoxia in root tips of maize seedlings acclimated to a low oxygen environment and identification of protein by mass spectrometry. Plant. Physiol. 2000, 122, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Ahmad, M.S.A.; Ashraf, M.; Al-Qurainy, F.; Ashraf, M.Y. Alleviation of waterlogging stress in upland cotton (Gossypium hirsutum L.) by exogenous application of potassium in soil and as a foliar spray. Crop Pasture Sci. 2011, 62, 25–38. [Google Scholar] [CrossRef]
- Ashraf, M.A. Waterlogging stress in plants: A review. Afr. J. Agril. Res. 2012, 7, 1976–1981. [Google Scholar]
- Malik, A.I.; Colmer, T.D.; Lamber, H.; Schortemeyer, M. Changes in physiological and morphological traits of roots and shoots of wheat in response to different depths of waterlogging. Aust. J. Plant. Physiol. 2001, 28, 1121–1131. [Google Scholar] [CrossRef]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotech. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Sairam, R.K.; Srivastava, G.C. Changes in antioxidant activity in sub-cellular fraction of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Ezin, V.; Pena, R.D.L.; Ahanchede, A. Flooding tolerance of tomato genotypes during vegetative and reproductive stages. Braz. J. Plant Physiol. 2010, 22, 131–142. [Google Scholar] [CrossRef]
- Paltaa, J.A.; Ganjealic, A.; Turnerb, N.C.; Siddique, K.H.M. Effects of transient subsurface waterlogging on root growth, plant biomass and yield of chickpea. Agric. Water Manag. 2010, 97, 1469–1476. [Google Scholar] [CrossRef]
- Rasaei, A.; Ghobadi, M.E.; Jalali-Honarmand, S.; Ghobadi, M.; Saeidi, M. Waterlogging and its effects on nitrogen of soil and plant. Ann. Biol. Res. 2012, 3, 119–124. [Google Scholar]
- Kumar, P.; Pal, M.; Joshi, R.; Sairam, R.K. Yield, growth and physiological responses of mung bean [Vigna radiata (L.) Wilczek] genotypes to waterlogging at vegetative stage. Physiol. Mol. Biol. Plants 2013, 19, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism oxidative stress and signal transduction. Ann. Rev. Plant Mol. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Hossain, M.S.; Mahmud, J.A.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 200. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, L.; Zhang, Y.; Qi, X.; Li, D.; Wei, W. Global gene expression responses to waterlogging in roots of sesame (Sesamum indicum L.). Acta Physiol. Plant. 2012, 34, 2241–2249. [Google Scholar]
- Snowden, R.E.D.; Wheeler, B.D. Iron toxicity to fern plant species. J. Ecol. 1993, 81, 35–46. [Google Scholar]
- Wei, W.; Li, D.; Wang, L.; Ding, X.; Zhang, Y.; Gao, Y.; Zhang, X. Morpho-anatomical and physiological responses to waterlogging of sesame (Sesamum indicum L.). Plant Sci. 2013, 208, 102–111. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity. Technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Arnon, D.T. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teari, D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Exogenous nitric oxide alleviates high temperature induced oxidative stress in wheat (Triticum aestivum L.) seedlings by modulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci. 2012, 6, 1314–1323. [Google Scholar]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Wild, R.; Ooi, L.; Srikanth, V.; Münch, G. A quick, convenient and economical method for the reliable determination of methylglyoxal in millimolar concentrations: The N-acetyl-L-cysteine assay. Anal. Bioanal. Chem. 2012, 403, 2577–2581. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; He, W.; Guo, J.; Chang, X.; Su, P.; Zhang, L. Increased sensitivity to salt stress in an ascorbate-deficient Arabidopsis mutant. J. Exp. Bot. 2005, 56, 3041–3049. [Google Scholar] [CrossRef]
- Paradiso, A.; Berardino, R.; de Pinto, M.; di Toppi, L.; Storelli, S.; de Gara, F.T. Increase in ascorbate-glutathione metabolism as local and precocious systemic responses induced by cadmium in durum wheat plants. Plant Cell Physiol. 2008, 49, 362–374. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in the regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dorr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Principato, G.B.; Rosi, G.; Talesa, V.; Govannini, E.; Uolila, L. Purification and characterization of two forms of glyoxalase II from rat liver and brain of Wistar rats. Biochem. Biophys. Acta 1987, 911, 349–355. [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M.; Karzel, E.M. Osmotic adjustment of sugar beets in response to soil salinity and its influence on stomatal conductance, growth and yield. Agric. Water Manag. 1997, 34, 57–69. [Google Scholar] [CrossRef]
- Min, X.J.; Bartholomew, D.P. Effects of flooding and drought on ehtylene metabolism, titratable acidity and fruiting of pineapple. Acta. Hortic. 2005, 666, 135–148. [Google Scholar] [CrossRef]
- Barickman, T.C.; Simpson, C.R.; Sams, C.E. Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants. Plants 2019, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Arias, C.C.; Gómez-Caro, S.; Restrepo-Díaz, H. Physiological, biochemical and chlorophyll fluorescence parameters of Physalis peruviana L. seedlings exposed to different short-term waterlogging periods and Fusarium wilt infection. Agronomy 2019, 9, 213. [Google Scholar] [CrossRef]
- Aloni, B.; Rosenshtein, G. Effect of flooding on tomato cultivars: The relationship between proline accumulation and other morphological and physiological changes. Physiol. Plant. 1982, 56, 513–517. [Google Scholar] [CrossRef]
- Olgun, M.; Kumlay, A.M.; Adiguzel, M.C.; Caglar, A. Effects of waterlogging in wheat (T. aestivum L.). Acta Agric. Scand. B Plant Soil Sci. 2008, 58, 193–198. [Google Scholar]
- Bajpai, S.; Chandra, R. Effect of waterlogging stress on growth characteristics and SOD gene expression in sugarcane. Int. J. Sci. Res. Pub. 2015, 5, 1–8. [Google Scholar]
- Bacanamwo, M.; Purcell, L.C. Soybean dry matter and N accumulation responses to flooding stress, N sources and hypoxia. J. Exp. Bot. 1999, 50, 689–696. [Google Scholar] [CrossRef]
- Collaku, A.; Harrison, S.A. Loses in wheat due to waterlogging. Crop Sci. 2002, 42, 444–450. [Google Scholar] [CrossRef]
- Prasad, S.; Ram, P.C.; Uma, S. Effect of waterlogging duration on chlorophyll content, nitrate reductase activity, soluble sugars and grain yield of maize. Ann. Plant Physiol. 2004, 18, 1–5. [Google Scholar]
- Mensah, J.K.; Obadoni, B.O.; Eruotor, P.G.; Onome-Irieguna, F. Simulated flooding and drought effects on germination, growth, and yield parameters of sesame (Sesamum indicum L.). Afr. J. Biotechnol. 2006, 5, 1249–1253. [Google Scholar]
- Yiu, J.C.; Liu, C.W.; Kuo, C.T.; Tseng, M.J.; Lai, Y.S.; Lai, W.J. Changes in antioxidant properties and their relationship to paclobutrazol indued flooding tolerance in Welsh Onion. J. Sci. Food Agric. 2008, 88, 1222–1230. [Google Scholar] [CrossRef]
- Ullah, I.; Waqas, M.; Khan, M.A.; Lee, I.-J.; Kim, W.-C. Exogenous ascorbic acid mitigates flood stress damages of Vigna angularis. Appl. Biol. Chem. 2017, 60, 603–614. [Google Scholar] [CrossRef]
- Zheng, X.; Zhou, J.; Tan, D.-X.; Wang, N.; Wang, L.; Shan, D.; Kong, J. Melatonin improves waterlogging tolerance of Malus baccata (Linn.) borkh. seedlings by maintaining aerobic respiration, photosynthesis and ROS migration. Front. Plant Sci. 2017, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Alegre, L. Plant aging increases oxidative stress in chloroplast. Planta 2002, 214, 608–615. [Google Scholar]
- Dalton, D.A.; Russell, S.A.; Hanus, F.J.; Pascoe, G.A.; Evans, H.J. Enzymatic reactions of ascorbate and glutathione that prevent peroxide damage in soybean root nodules. Proc. Nat. Acad. Sci. USA 1986, 83, 3811–3815. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-induced upregulation of the antioxidant defense and methylglyoxal detoxification system reduces salinity-induced damage in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol. Rep. 2011, 5, 353–365. [Google Scholar] [CrossRef]
- Lin, K.R.; Weng, C.; Lo, H.; Chen, J. Study of the root antioxidative system of tomatoes and eggplants under waterlogged conditions. Plant Sci. 2004, 167, 355–365. [Google Scholar] [CrossRef]
- May, M.J.; Vernoux, T.; Leaver, C.; Van Montagu, M.; Inze, D. Glutathione homeostasis in plants: Implications for environmental sensing and plant development. J. Exp. Bot. 1998, 49, 649–667. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Fujita, M. Modulation of antioxidant machinery and the methylglyoxal detoxification system in selenium-supplemented Brassica napus seedlings confers tolerance to high temperature stress. Biol. Trace Elem. Res. 2014, 161, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Fujita, M. Glutathione-induced drought stress tolerance in mung bean: Coordinated roles of the antioxidant defense and methylglyoxal detoxification systems. AoB Plants 2015, 7, plv069. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.R.; Ahmed, F.; Mokarroma, N.; Rohman, M.M.; Golder, P.C. Physiological and biochemical changes in waterlog tolerant sesame genotypes. SAARC J. Agric. 2016, 14, 31–45. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhang, J. Advances in the research on the AsA–GSH cycle in horticultural crops. Front. Agric. China 2010, 4, 84–90. [Google Scholar] [CrossRef]
- Pang, C.H.; Wang, B.S. Role of ascorbate peroxidase and glutathione reductase in ascorbate–glutathione cycle and stress tolerance in plants. In Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Anjum, N.A., Chan, M.T., Umar, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 91–112. [Google Scholar]
- Apostolova, P.; Szalai, G.; Kocsy, G.; Janda, T.; Popova, L.P. Environmental factors affecting components of ascorbate-glutathione pathway in crop plants. In Oxidative Stress in Plants: Causes, Consequences and Tolerance; Anjum, N.A., Umar, S., Ahmad, A., Eds.; I. K. International: New Delhi, India, 2011; pp. 52–70. [Google Scholar]
- Wang, Y.; Wisniewski, M.; Meilan, R.; Cui, M.; Fuchigami, L. Transgenic tomato (Lycopersicon esculentum) overexpressing cAPX exhibits enhanced tolerance to UV-B and heat stress. J. Appl. Hortic. 2006, 8, 87–90. [Google Scholar]
- Xu, W.F.; Shi, W.M.; Ueda, A.; Takabe, T. Mechanisms of salt tolerance in transgenic Arabidopsis thaliana carrying a peroxisomal ascorbate peroxidase gene from barley. Pedosphere 2008, 18, 486–495. [Google Scholar] [CrossRef]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plant. 2000, 110, 42–51. [Google Scholar] [CrossRef]
- Martínez, J.P.; Araya, H. Ascorbate–glutathione cycle: Enzymatic and non-enzymatic integrated mechanisms and its biomolecular regulation. In Ascorbate Glutathione Pathway and Stress Tolerance in Plants; Anjum, N.A., Chan, M.T., Umar, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 303–322. [Google Scholar]
- Chalapathi Rao, A.S.V.; Reddy, A.R. Glutathione reductase: A putative redox regulatory system in plant cells. In Sulfur Assimilation and Abiotic Stresses in Plants; Khan, N.A., Singh, S., Umar, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 111–147. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
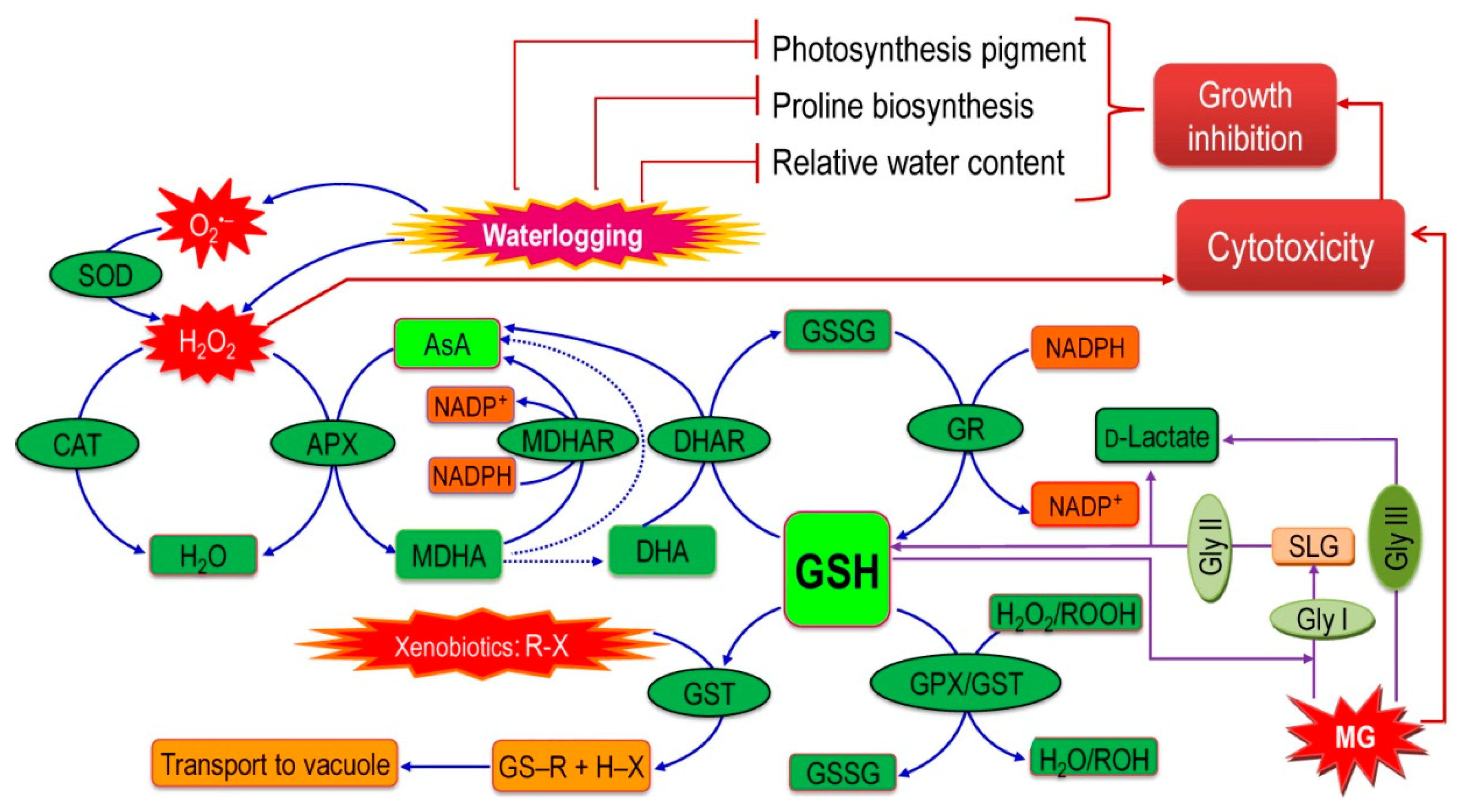{kind=link}
| Variables | RWC | Pro | Chl a | Chl b | Chl (a + b) | Car | MDA | H2O2 | AsA | GSH | GSSG | GSH/GSSG | APX | MDHAR | DHAR | GR | GPX | CAT | MG | Gly I | Gly II |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RWC | 1 | −0.035 | 0.808 | 0.808 | 0.862 | 0.790 | −0.844 | −0.816 | 0.811 | −0.832 | −0.876 | 0.631 | −0.656 | −0.804 | 0.781 | 0.496 | −0.793 | 0.708 | −0.833 | −0.138 | −0.665 |
| Proline | 1 | 0.258 | 0.181 | 0.252 | 0.012 | 0.253 | 0.168 | 0.298 | 0.129 | −0.147 | 0.210 | 0.452 | 0.373 | 0.395 | 0.408 | 0.325 | 0.443 | 0.128 | 0.349 | 0.140 | |
| Chl a | 1 | 0.705 | 0.975 | 0.847 | −0.710 | −0.813 | 0.862 | −0.805 | −0.918 | 0.742 | −0.490 | −0.618 | 0.897 | 0.573 | −0.621 | 0.733 | −0.752 | −0.052 | −0.529 | ||
| Chl b | 1 | 0.844 | 0.585 | −0.741 | −0.784 | 0.677 | −0.721 | −0.862 | 0.688 | −0.570 | −0.671 | 0.849 | 0.513 | −0.669 | 0.624 | −0.803 | −0.085 | −0.632 | |||
| Chl (a + b) | 1 | 0.823 | −0.767 | −0.859 | 0.863 | −0.833 | −0.962 | 0.775 | −0.548 | −0.676 | 0.943 | 0.593 | −0.678 | 0.749 | −0.818 | −0.066 | −0.596 | ||||
| Car | 1 | −0.741 | −0.794 | 0.777 | −0.790 | −0.843 | 0.638 | −0.607 | −0.709 | 0.710 | 0.425 | −0.629 | 0.535 | −0.725 | −0.092 | −0.566 | |||||
| MDA | 1 | 0.900 | −0.597 | 0.871 | 0.822 | −0.556 | 0.808 | 0.879 | −0.662 | −0.307 | 0.859 | −0.501 | 0.912 | 0.291 | 0.837 | ||||||
| H2O2 | 1 | −0.634 | 0.877 | 0.889 | −0.679 | 0.757 | 0.801 | −0.759 | −0.364 | 0.817 | −0.512 | 0.897 | 0.279 | 0.752 | |||||||
| AsA | 1 | −0.732 | −0.814 | 0.562 | −0.418 | −0.545 | 0.830 | 0.627 | −0.543 | 0.736 | −0.632 | 0.034 | −0.422 | ||||||||
| GSH | 1 | 0.840 | −0.519 | 0.765 | 0.740 | −0.691 | −0.291 | 0.817 | −0.538 | 0.891 | 0.213 | 0.666 | |||||||||
| GSSG | 1 | −0.861 | 0.600 | 0.780 | −0.915 | −0.530 | 0.706 | −0.685 | 0.878 | 0.194 | 0.646 | ||||||||||
| GSH/GSSG | 1 | −0.300 | −0.650 | 0.793 | 0.414 | −0.461 | 0.537 | −0.639 | −0.182 | −0.438 | |||||||||||
| APX | 1 | 0.658 | −0.377 | −0.195 | 0.835 | −0.102 | 0.778 | 0.359 | 0.511 | ||||||||||||
| MDHAR | 1 | −0.586 | −0.194 | 0.748 | −0.434 | 0.808 | 0.285 | 0.709 | |||||||||||||
| DHAR | 1 | 0.630 | −0.509 | 0.767 | −0.720 | 0.054 | −0.531 | ||||||||||||||
| GR | 1 | −0.153 | 0.674 | −0.345 | 0.226 | −0.221 | |||||||||||||||
| GPX | 1 | −0.351 | 0.817 | 0.359 | 0.661 | ||||||||||||||||
| CAT | 1 | −0.539 | 0.190 | −0.565 | |||||||||||||||||
| MG | 1 | 0.369 | 0.718 | ||||||||||||||||||
| Gly I | 1 | 0.320 | |||||||||||||||||||
| Gly II | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anee, T.I.; Nahar, K.; Rahman, A.; Mahmud, J.A.; Bhuiyan, T.F.; Alam, M.U.; Fujita, M.; Hasanuzzaman, M. Oxidative Damage and Antioxidant Defense in Sesamum indicum after Different Waterlogging Durations. Plants 2019, 8, 196. https://doi.org/10.3390/plants8070196
Anee TI, Nahar K, Rahman A, Mahmud JA, Bhuiyan TF, Alam MU, Fujita M, Hasanuzzaman M. Oxidative Damage and Antioxidant Defense in Sesamum indicum after Different Waterlogging Durations. Plants. 2019; 8(7):196. https://doi.org/10.3390/plants8070196
Chicago/Turabian StyleAnee, Taufika Islam, Kamrun Nahar, Anisur Rahman, Jubayer Al Mahmud, Tasnim Farha Bhuiyan, Mazhar Ul Alam, Masayuki Fujita, and Mirza Hasanuzzaman. 2019. "Oxidative Damage and Antioxidant Defense in Sesamum indicum after Different Waterlogging Durations" Plants 8, no. 7: 196. https://doi.org/10.3390/plants8070196
APA StyleAnee, T. I., Nahar, K., Rahman, A., Mahmud, J. A., Bhuiyan, T. F., Alam, M. U., Fujita, M., & Hasanuzzaman, M. (2019). Oxidative Damage and Antioxidant Defense in Sesamum indicum after Different Waterlogging Durations. Plants, 8(7), 196. https://doi.org/10.3390/plants8070196