Superhost Plants Alter the Structure of Plant–Galling Insect Networks in Neotropical Savannas

 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
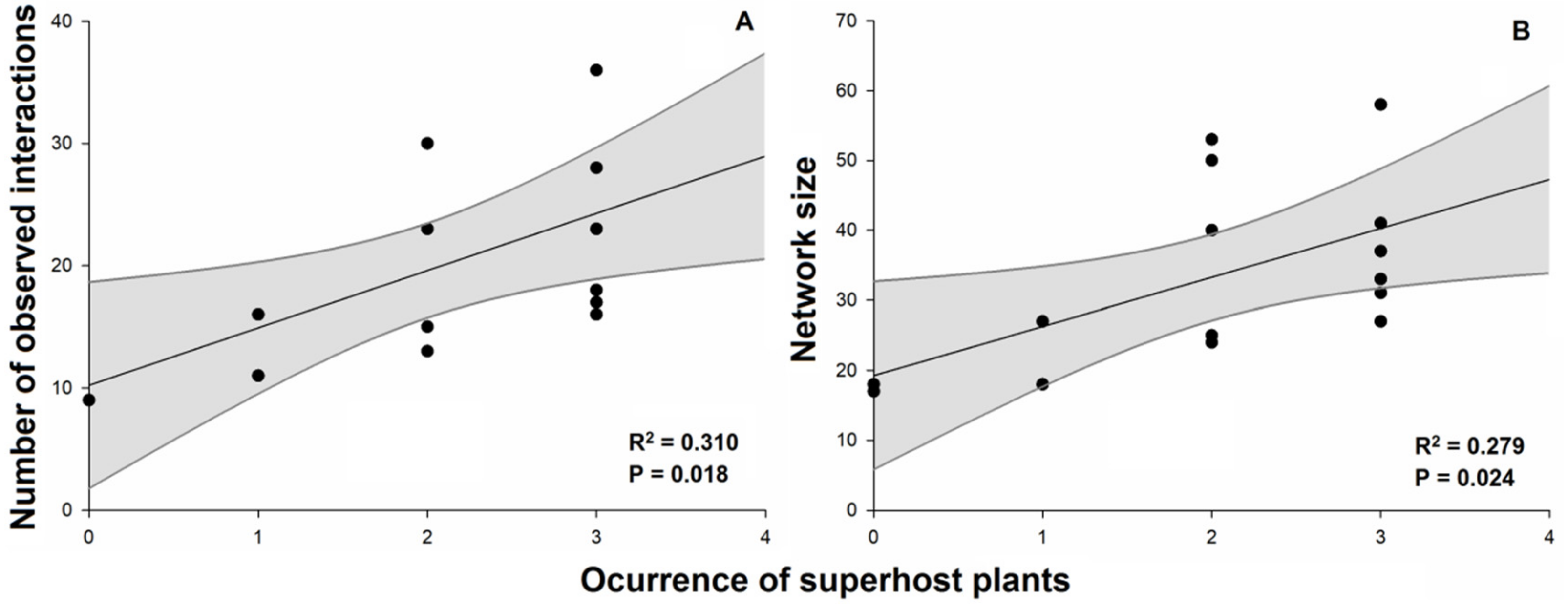
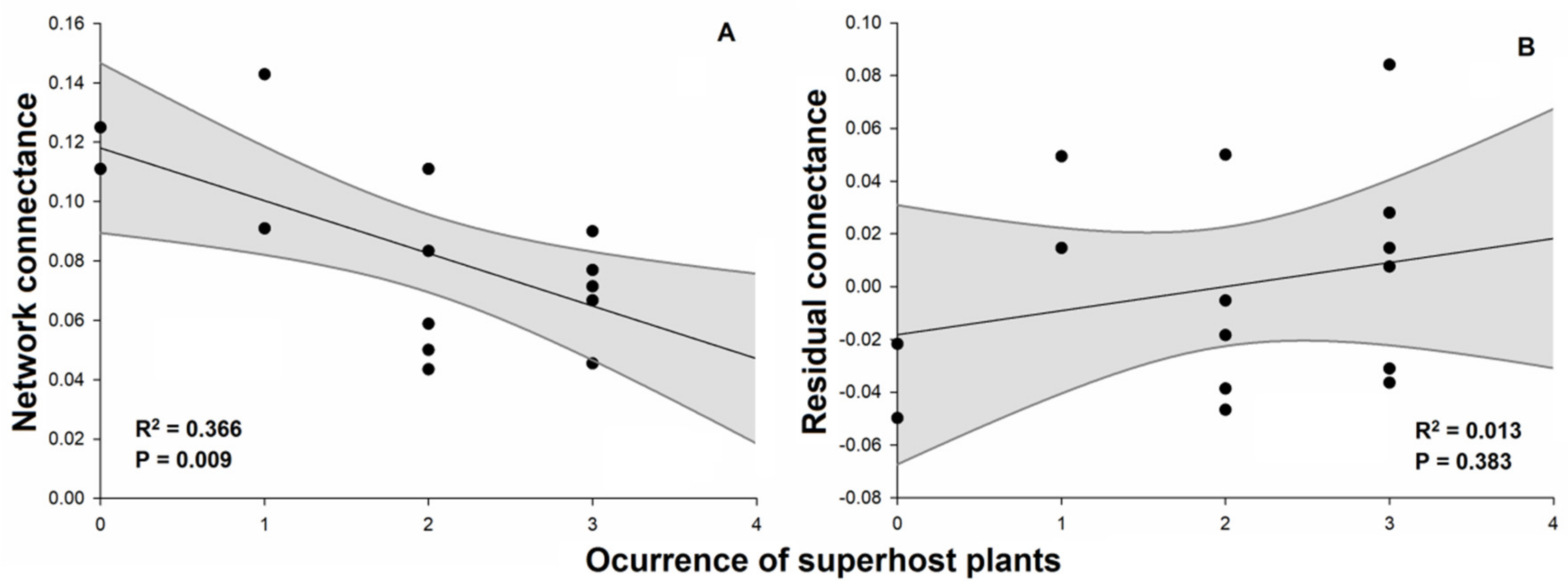
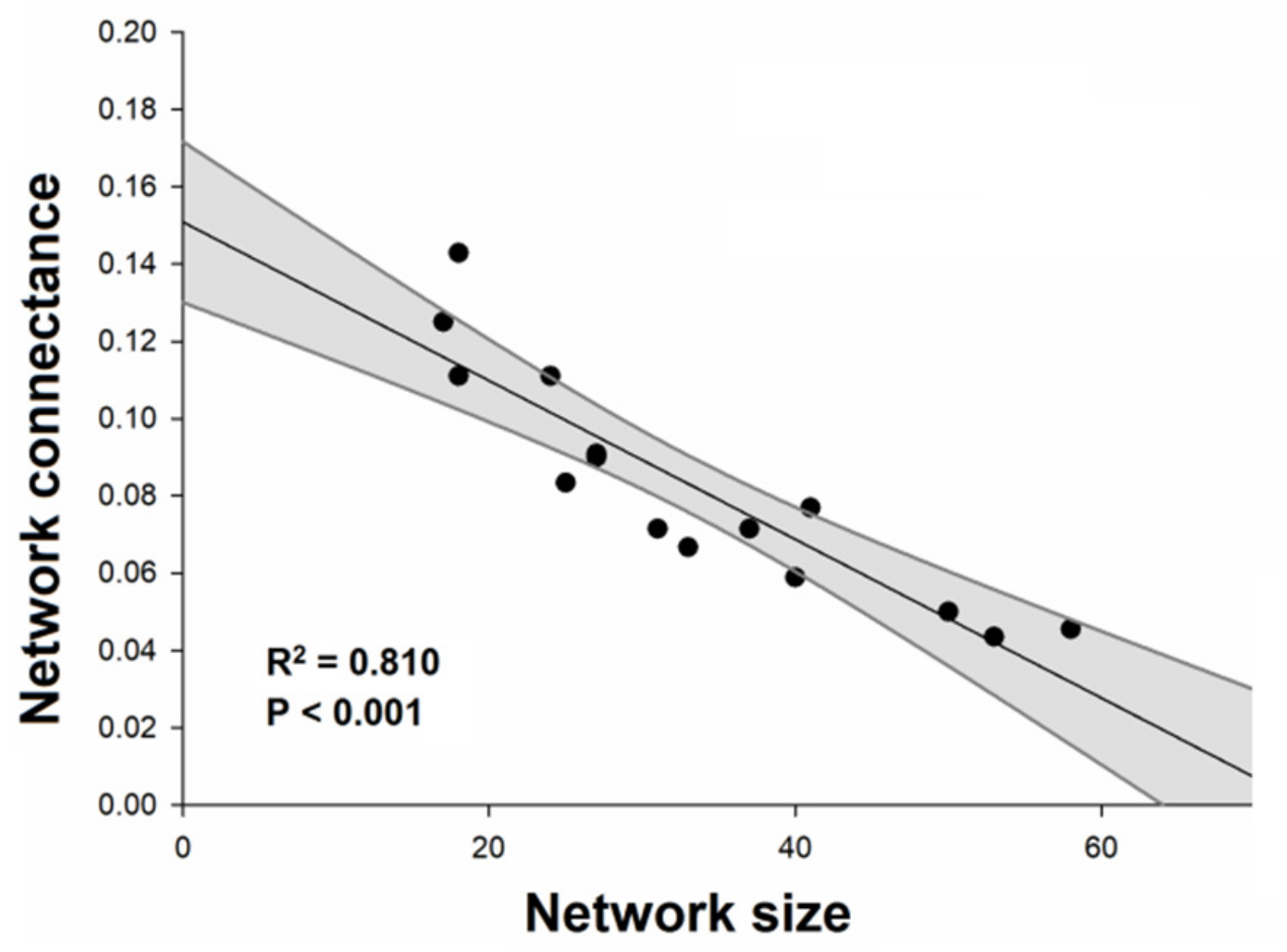
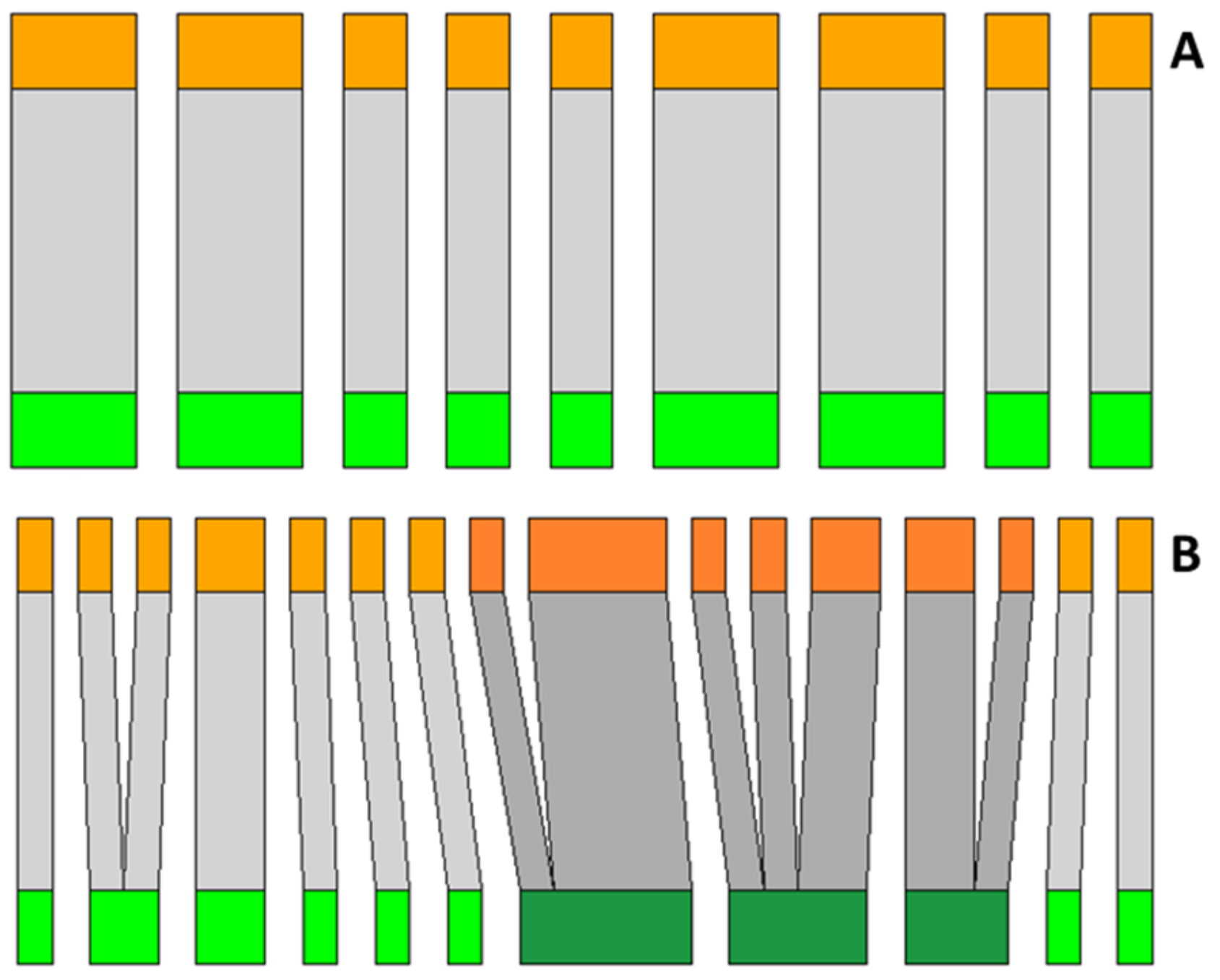
2. Results
3. Discussion
4. Materials and Methods
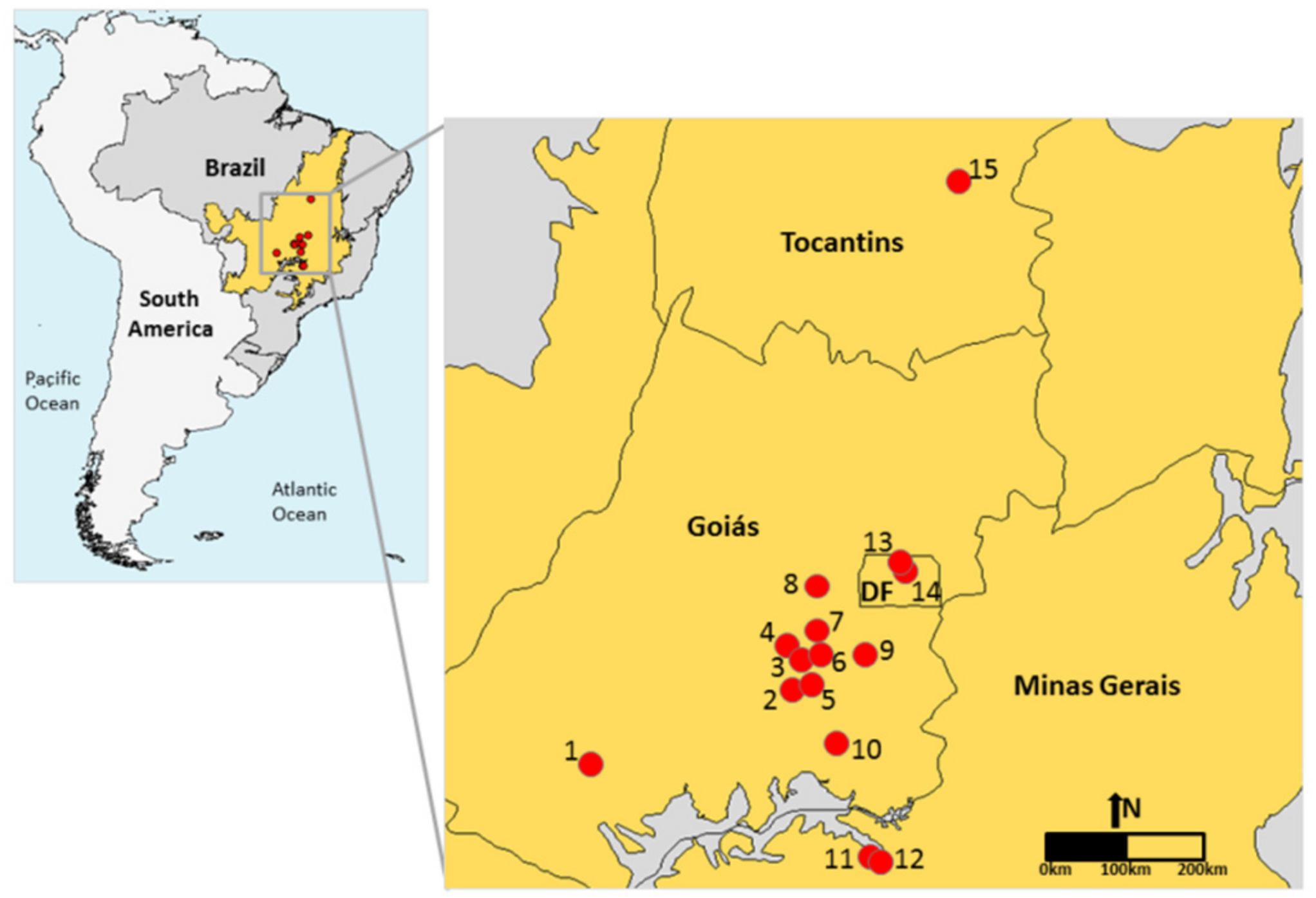
4.1. Study Area
4.2. Sampling of Plant–Galling Insect Interactions
4.3. Determination of Superhost Plants
4.4. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haddad, N.M.; Tilman, D.; Haarstad, J.; Ritchie, M.; Knops, J.M. Contrasting effects of plant richness and composition on insect communities: A field experiment. Am. Nat. 2001, 158, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, T.M.; Novotny, V.; Basset, Y. Insects on plants: Diversity of herbivore assemblages revisited. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 597–620. [Google Scholar] [CrossRef]
- Araújo, W.S. Different relationships between galling and non-galling herbivore richness and plant species richness: A meta-analysis. Arthropod Plant. Interact. 2013, 7, 373–377. [Google Scholar] [CrossRef]
- Kuchenbecker, J.; Fagundes, M. Diversity of insects associated with two common plants of the Brazilian Cerrado: Responses of two guilds of herbivores to bottom-up and top-down forces. Eur. J. Entomol. 2018, 115, 354–363. [Google Scholar] [CrossRef]
- Cornell, H.V. Endophage-ectophage ratios and plant defense. Evol. Ecol. 1989, 3, 64–76. [Google Scholar] [CrossRef]
- Shorthouse, J.D.; Wool, D.; Raman, A. Gall-inducing insects–Nature’s most sophisticated herbivores. Basic Appl. Ecol. 2005, 6, 407–411. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 10, 512–522. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Carneiro, M.A.; Isaias, R.M. Gall-inducing insects: From anatomy to biodiversity. In Insect Bioecology and Nutrition for Integrated Pest Management; Panizzi, A.R., Parra, J.R.P., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 369–395. [Google Scholar]
- Carneiro, M.A.A.; Branco, C.S.; Braga, C.E.; Almada, E.D.; Costa, M.; Maia, V.C.; Fernandes, G.W. Are gall midge species (Diptera, Cecidomyiidae) host-plant specialists? Rev. Bras. Entomol. 2009, 3, 365–378. [Google Scholar] [CrossRef]
- Hardy, N.B.; Cook, L.G. Gall-induction in insects: Evolutionary dead-end or speciation driver? BMC Evol. Biol. 2010, 10, 257. [Google Scholar] [CrossRef]
- Araújo, W.S.; Grandez-Riosllar, J. Exotic species and the structure of a plant-galling network. Netw. Biol. 2017, 7, 21–32. [Google Scholar]
- Araújo, W.S.; Scareli-Santos, C.; Guilherme, F.A.G.; Cuevas-Reyes, P. Comparing galling insect richness among Neotropical savannas: Effects of plant richness, vegetation structure and super-host presence. Biodivers. Conserv. 2013, 22, 1083–1094. [Google Scholar]
- Veldtman, R.; McGeoch, M.A. Gall-forming insect species richness along a non-scleromorphic vegetation rainfall gradient in South Africa: The importance of plant community composition. Austral Ecol. 2003, 28, 1–13. [Google Scholar] [CrossRef]
- Fagundes, M.; Xavier, R.C.F.; Faria, M.L.; Cuevas-Reyes, P.; Lopes, L.G.; Reis-Junior, R. Plant phenological asynchrony and community structure of gall-inducing insects in a super-host tropical tree species. Ecol. Evol. 2018, 8, 10687–10697. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.S.; Cuevas-Reyes, P.; Guilherme, F.A.G. Local and regional determinants of galling-insect richness in Neotropical savanna. J. Trop. Ecol. 2014, 30, 269–272. [Google Scholar]
- Coutinho, R.D.; Cuevas-Reyes, P.; Fernandes, G.W.; Fagundes, M. Community structure of gall-inducing insects associated with a tropical shrub: Regional, local and individual patterns. Trop. Ecol. 2019, 60, 74–82. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Carneiro, M.A.A.; Lara, A.C.F.; Allain, L.R.; Andrade, G.I.; Julião, G.R.; Reis, T.R.; Silva, I.M. Galling insects on neotropical species of Baccharis (Asteraceae). Trop. Zool. 1996, 9, 315–332. [Google Scholar] [CrossRef]
- Costa, F.V.D.; Fagundes, M.; Neves, F.S. Arquitetura da planta e diversidade de galhas associadas à Copaifera langsdorffii (Fabaceae). Ecol. Austral 2010, 20, 9–17. [Google Scholar]
- Blanche, K.R.; Westoby, M. Gall-forming insect diversity is linked to soil fertility via host plant taxon. Ecology 1995, 76, 2334–2337. [Google Scholar] [CrossRef]
- Oyama, K.; Scareli-Santos, C.; Mondragón-Sánchez, M.L.; Tovar-Sánchez, E.; Cuevas-Reyes, C. Morphological variations of gall-forming insects on different species of oaks (Quercus) in Mexico. In Ecology and Conservation of Neotropical Montane Oak Forests; Kappelle, M., Ed.; Springer: Berlin, Gemany, 2006; pp. 259–269. [Google Scholar]
- Poisot, T.; Stouffer, D.B.; Kéfi, S. Describe, understand and predict: Why do we need networks in ecology? Funct. Ecol. 2016, 30, 1878–1882. [Google Scholar] [CrossRef]
- Dormann, C.F.; Fründ, J.; Blüthgen, N.; Gruber, B. Indices, graphs and null models: Analyzing bipartite ecological networks. Open Ecol. J. 2009, 2, 7–24. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 1–9. [Google Scholar]
- Araújo, W.S.; Vieira, M.C.; Lewinsohn, T.M.; Almeida-Neto, M. Contrasting effects of land use intensity and exotic host plants on the specialization of interactions in plant-herbivore networks. PLoS ONE 2015, 10, e0115606. [Google Scholar]
- Ribeiro, J.F.; Walter, B.M.T. As principais fitofisionomias do bioma Cerrado. In Cerrado: Ecologia e Flora, 1st ed.; Sano, S.M., Almeida, S.P., Ribeiro, J.F., Eds.; EMBRAPA: Brasília, Brazil, 2008; Volume 1, pp. 152–212. [Google Scholar]
- Oliveira-Filho, A.D.; Martins, F.R. A comparative study of five cerrado areas in southern Mato Grosso, Brazil. Edinb. J. Bot. 1991, 48, 307–332. [Google Scholar] [CrossRef]
- Gonçalves-Alvim, S.; Fernandes, G.W. Biodiversity of galling insects: Historical, community and habitat effects in four neotropical savannas. Biodivers. Conserv. 2001, 10, 79–98. [Google Scholar]
- Silva-Júnior, M.C.; Santos, G.C. 100 árvores do Cerrado: Guia de Campo, 1st ed.; Rede de sementes do Cerrado: Brasília, Brazil, 2005. [Google Scholar]
- Ratter, J.A.; Bridgewater, S.; Ribeiro, J.F. Analysis of the floristic composition on the Brazilian cerrado vegetation III: Comparison of the woody vegetation of 376 areas. Edinb. J. Bot. 2003, 60, 57–109. [Google Scholar] [CrossRef]
- Felfili, J.M.; Silva-Júnior, M.C.; Sevilha, A.C.; Fagg, C.W.; Walter, B.M.T.; Nogueira, P.E.; Rezende, A.Z. Diversity, floristic and structural patterns of cerrado vegetation in Central Brazil. Plant Ecol. 2004, 175, 37–46. [Google Scholar] [CrossRef]
- Araújo, W.S.; Kóllar, J. First characterization of a highly specialized ecological network composed by gall-inducing mites and their host plants. Int. J. Acarol. 2019, 45, 1–4. [Google Scholar]
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. Food-web structure and network theory: The role of connectance and size. Proc. Natl. Acad. Sci. USA 2002, 20, 12917–12922. [Google Scholar] [CrossRef]
- Landi, P.; Minoarivelo, H.O.; Brännström, Å.; Hui, C.; Dieckmann, U. Complexity and stability of ecological networks: A review of the theory. Pop. Ecol. 2018, 60, 319–345. [Google Scholar] [CrossRef]
- Delmas, E.; Besson, M.; Brice, M.H.; Burkle, L.A.; Dalla Riva, G.V.; Fortin, M.J.; Gravel, D.; Guimaraes, P.R., Jr.; Hembry, D.H.; Newman, E.A.; et al. Analysing ecological networks of species interactions. Biol. Rev. 2019, 94, 16–36. [Google Scholar] [CrossRef]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; pp. 1–2673. [Google Scholar]
- Staab, M.; Blüthgen, N.; Klein, A.M. Tree diversity alters the structure of a tri-trophic network in a biodiversity experiment. Oikos 2015, 124, 827–834. [Google Scholar] [CrossRef]
- Welti, E.; Helzer, C.; Joern, A. Impacts of plant diversity on arthropod communities and plant–herbivore network architecture. Ecosphere 2017, 8, e01983. [Google Scholar] [CrossRef]
- Saiz, H.; Gómez-Gardeñes, J.; Borda, J.P.; Maestre, F.T. The structure of plant spatial association networks is linked to plant diversity in global drylands. J. Ecol. 2018, 106, 1443–1453. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Network Measure | Deviance Resid. | Df | Resid. Dev | F-Value | P-Value |
|---|---|---|---|---|---|
| Linkage density of plant species | 8.2298 | 11 | 8.1250 | 3.714 | 0.045 |
| Number of observed interactions | 351.56 | 13 | 626.04 | 7.300 | 0.018 |
| Network size | 784.00 | 13 | 1584.9 | 6.430 | 0.025 |
| Network connectance | 0.0050 | 13 | 0.0072 | 9.090 | 0.009 |
| Area | Locality | Coordinates | Occurrence of Superhost Plants |
|---|---|---|---|
| 1 | Lajeado, GO | 17°53′ S, 51°38′ W | Q. grandiflora and Q. multiflora |
| 2 | Banana Menina, GO | 16°59′ S, 49°14′ W | Q. grandiflora and Q. multiflora |
| 3 | Senador Canedo, GO | 16°43′ S, 49°06′ W | Q. grandiflora |
| 4 | Itanhangá, GO | 16°33′ S, 49°17′ W | Q. grandiflora and Q. parviflora |
| 5 | Bela Vista, GO | 15°57′ S, 48°56′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 6 | Bom Sucesso, GO | 16°42′ S, 49°02′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 7 | UEG, GO | 16°22′ S, 48°56′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 8 | Pedreira, GO | 15°50′ S, 48°55′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 9 | Fazenda Geraldo, GO | 16°40′ S, 48°18′ W | Q. grandiflora and Q. parviflora |
| 10 | Caldas Novas, GO | 17°42′ S, 48°38′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 11 | Caça e Pesca, MG | 19°00′ S, 48°18′ W | Q. grandiflora, Q. multiflora and Q. parviflora |
| 12 | Floresta do Lobo, MG | 19°05′ S, 48°09′ W | Q. grandiflora and Q. multiflora |
| 13 | APA Cafuringa, DF | 15°31′ S, 47°57′ W | No Qualea |
| 14 | REBio Contagem, DF | 15°37′ S, 47°52′ W | No Qualea |
| 15 | Porto Real, TO | 11°00′ S, 47°13′ W | Q. parviflora |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo, W.S.d.; Moreira, L.T.; Falcão, L.A.D.; Borges, M.A.Z.; Fagundes, M.; Faria, M.L.d.; Guilherme, F.A.G. Superhost Plants Alter the Structure of Plant–Galling Insect Networks in Neotropical Savannas. Plants 2019, 8, 369. https://doi.org/10.3390/plants8100369
Araújo WSd, Moreira LT, Falcão LAD, Borges MAZ, Fagundes M, Faria MLd, Guilherme FAG. Superhost Plants Alter the Structure of Plant–Galling Insect Networks in Neotropical Savannas. Plants. 2019; 8(10):369. https://doi.org/10.3390/plants8100369
Chicago/Turabian StyleAraújo, Walter Santos de, Leuzeny Teixeira Moreira, Luiz Alberto Dolabela Falcão, Magno Augusto Zazá Borges, Marcílio Fagundes, Maurício Lopes de Faria, and Frederico Augusto Guimarães Guilherme. 2019. "Superhost Plants Alter the Structure of Plant–Galling Insect Networks in Neotropical Savannas" Plants 8, no. 10: 369. https://doi.org/10.3390/plants8100369