10,000-Times Diluted Doses of ACCase-Inhibiting Herbicides Can Permanently Change the Metabolomic Fingerprint of Susceptible Avena fatua L. Plants


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
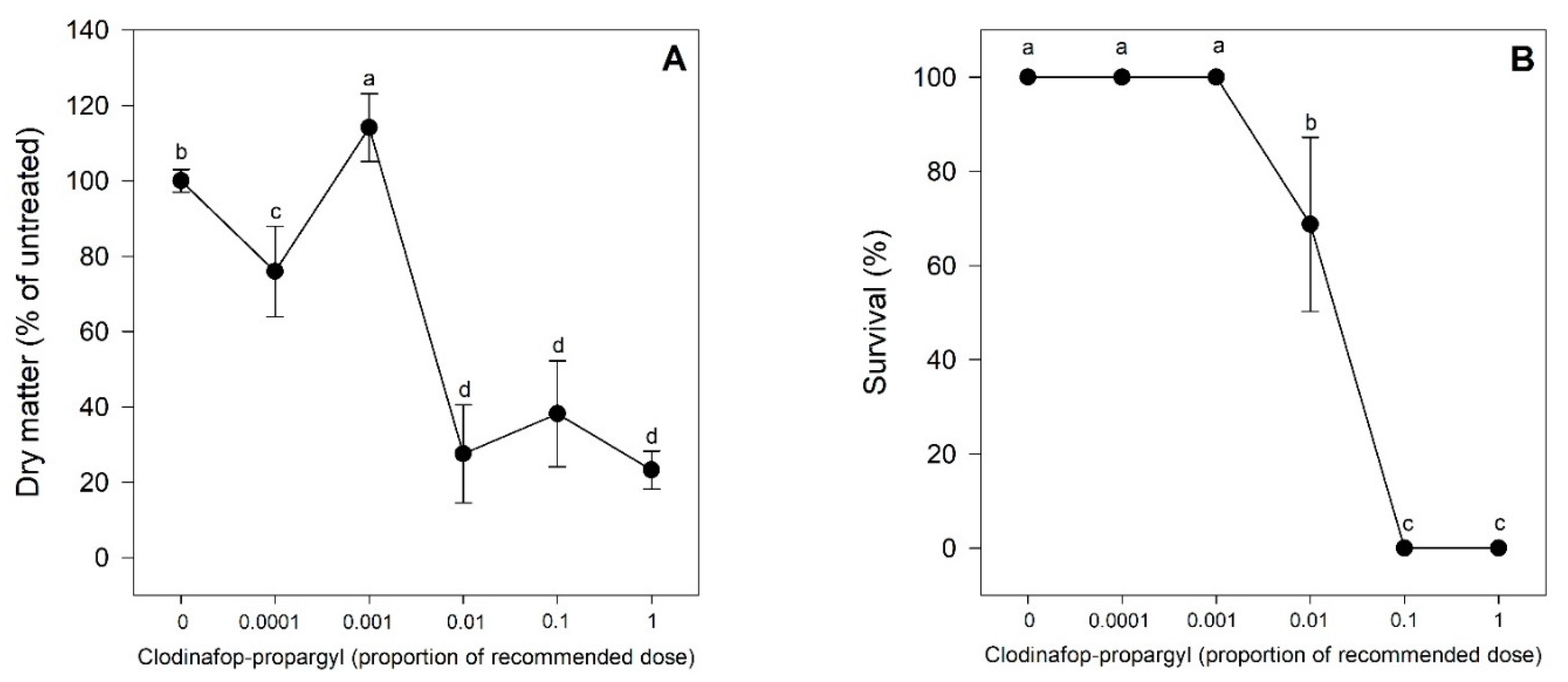
2.1. Experiment 1. Biomass Accumulation and Survival
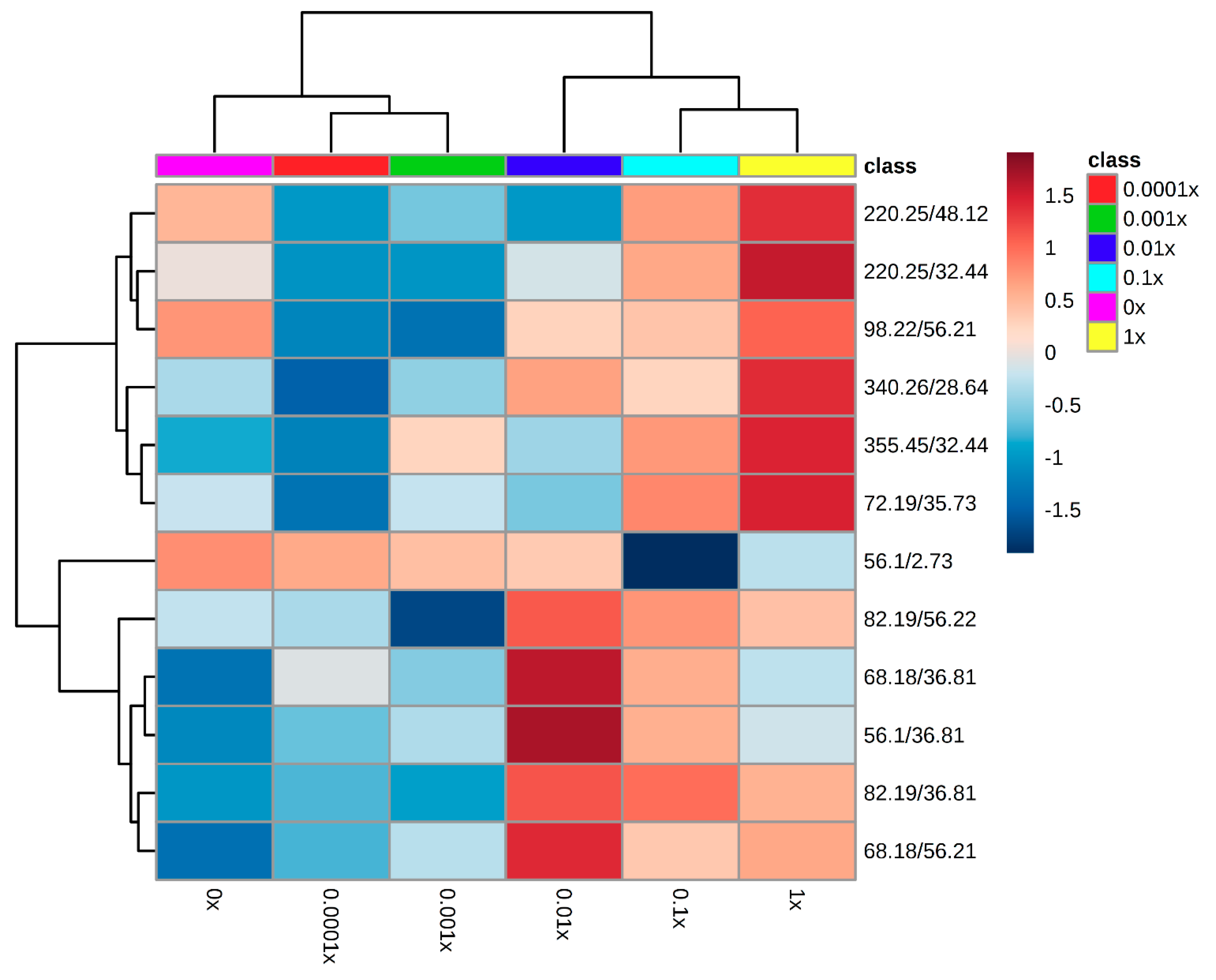
Metabolomic Fingerprinting of the First Experiment
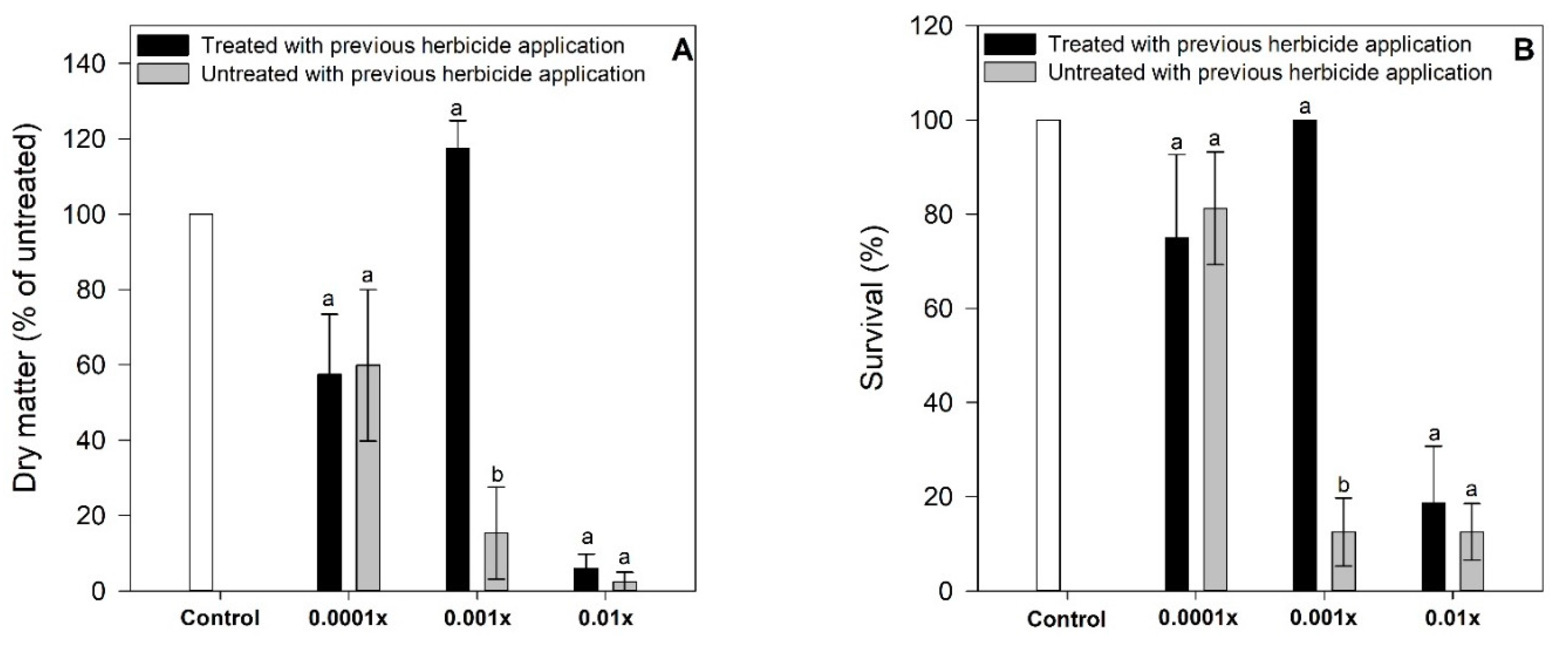
2.2. Experiment 2. Biomass Accumulation and Survival
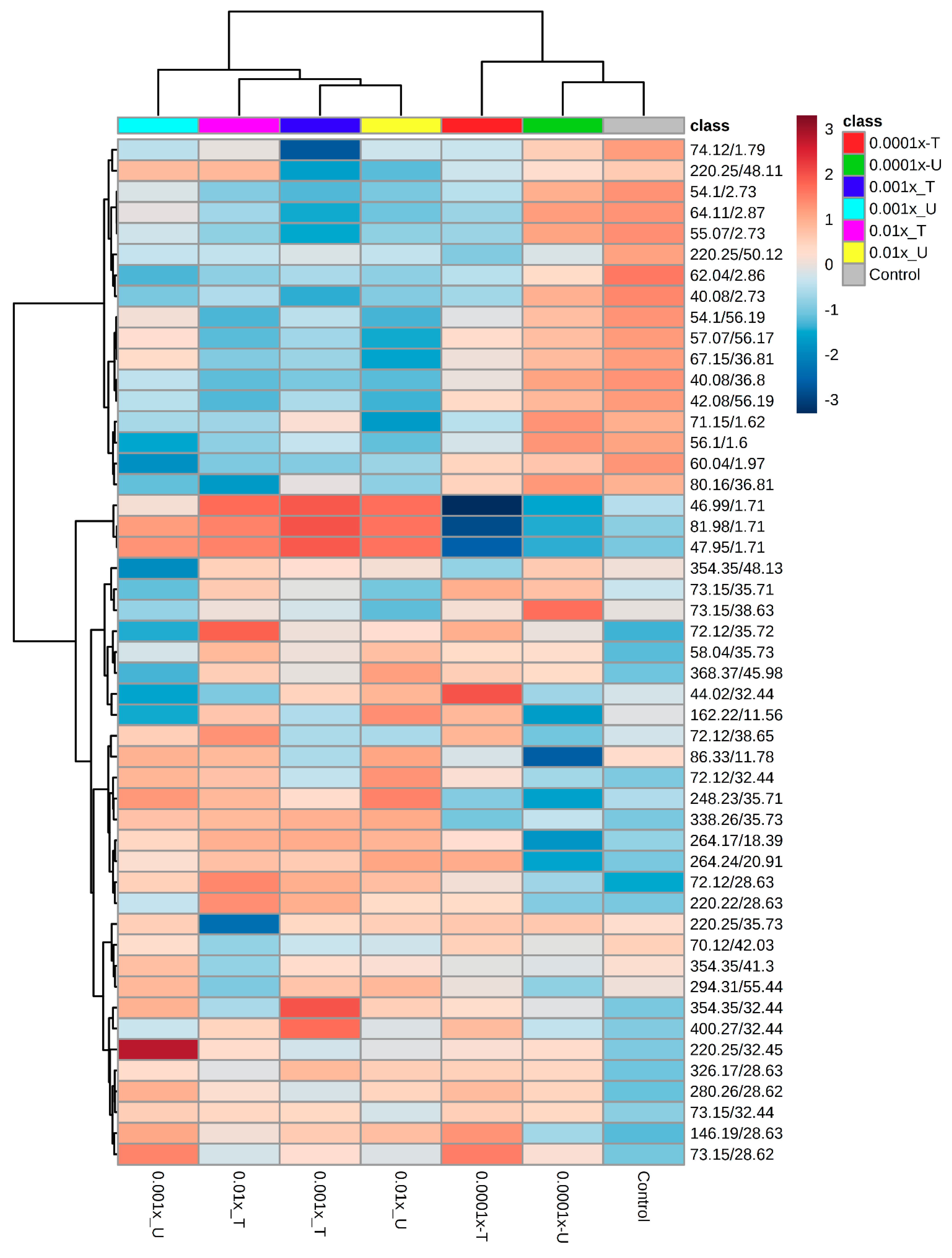
Metabolomic Fingerprint the Second Experiment
3. Discussion
4. Methods
4.1. Study System
4.2. Experiment 1. Determination of the Minimum Dose of Herbicide That Produces Changes to Plants’ Metabolism
4.2.1. Survival and Biomass Reduction
4.2.2. Metabolic Fingerprinting Using GC-MS
4.2.3. Data Analyses
4.3. Experiment 2. Responses of Weeds Treated with Non-Lethal Dose to Posterior Herbicide Application
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed]
- Neve, P. Challenges for herbicide resistance evolution and management: 50 Years after Harper. Weed Res. 2007, 47, 365–369. [Google Scholar] [CrossRef]
- Jang, S.; Marjanovic, J.; Gornicki, P. Resistance to herbicides caused by single amino acid mutations in acetyl-CoA carboxylase in resistant populations of grassy weeds. New Phytol. 2013, 197, 1110–1116. [Google Scholar] [CrossRef]
- Maxwell, B.D.; Roush, M.L.; Radosevich, S.R. Predicting the Evolution and Dynamics of Herbicide Resistance in Weed Populations Resistance in Weed Populations. Weed Technol. 1990, 4, 2–13. [Google Scholar] [CrossRef]
- Manalil, S. Evolution of herbicide resistance in Lolium rigidum under low herbicide rates: An Australian experience. Crop Sci. 2014, 54, 461–474. [Google Scholar] [CrossRef]
- Yu, Q.; Powles, S. Metabolism-Based Herbicide Resistance and Cross-Resistance in Crop Weeds: A Threat to Herbicide Sustainability and Global Crop Production. Plant Physiol. 2014, 166, 1106–1118. [Google Scholar] [CrossRef]
- Manalil, S.; Busi, R.; Renton, M.; Powles, S.B. Rapid Evolution of Herbicide Resistance by Low Herbicide Dosages. Weed Sci. 2011, 59, 210–217. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S. Recurrent selection with reduced herbicide rates results in the rapid evolution of herbicide resistance in Lolium rigidum. Theor. Appl. Genet. 2005, 110, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Neve, P.; Powles, S. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 2005, 95, 485–492. [Google Scholar] [CrossRef]
- Conrath, U. Molecular aspects of defence priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flores, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef]
- Belz, R.G.; Duke, S.O. Herbicides and plant hormesis. Pest Manag. Sci. 2014, 70, 698–707. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Bachmann, K.A.; Bailer, A.J.; Bolger, P.M.; Borak, J.; Cai, L.; Cedergreen, N.; Cherian, M.G.; Chiueh, C.C.; Clarkson, T.W.; et al. Biological stress response terminology: Integrating the concepts of adaptive response and preconditioning stress within a hormetic dose-response framework. Toxicol. Appl. Pharmacol. 2007, 222, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Gressel, J. Low pesticide rates may hasten the evolution of resistance by increasing mutation frequencies. Pest Manag. Sci. 2011, 67, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Brito, I.P.F.S.; Tropaldi, L.; Carbonari, C.A.; Velini, E.D. Hormetic effects of glyphosate on plants. Pest Manag. Sci. 2018, 74, 1064–1070. [Google Scholar] [CrossRef]
- Cedergreen, N.; Streibig, J.C.; Kudsk, P.; Mathiassen, S.K.; Duke, S.O. The Occurrence of Hormesis in Plants and Algae. Dose-Response 2007, 5, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Cedergreen, N. Herbicides can stimulate plant growth. Weed Res. 2008, 48, 429–438. [Google Scholar] [CrossRef]
- Gressel, J.; Dodds, J. Commentary: Hormesis can be used in enhancing plant productivity and health; but not as previously envisaged. Plant Sci. 2013, 213, 123–127. [Google Scholar] [CrossRef]
- Belz, R.G.; Cedergreen, N.; Duke, S.O. Herbicide hormesis—can it be useful in crop production? Weed Res. 2011, 51, 321–332. [Google Scholar] [CrossRef]
- Vieira, B.C.; Luck, J.D.; Amundsen, K.L.; Gaines, T.A.; Werle, R.; Kruger, G.R. Response of Amaranthus spp. following exposure to sublethal herbicide rates via spray particle drift. PLoS ONE 2019, 14, e0220014. [Google Scholar] [CrossRef]
- Saito, K.; Matsuda, F. Metabolomics for Functional Genomics, Systems Biology, and Biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef]
- Viant, M.R. Applications of metabolomics to the environmental sciences. Metabolomics. 2009, 5, 1–2. [Google Scholar] [CrossRef]
- Brunetti, C.; George, R.M.; Tattini, M.; Field, K.; Davey, M.P. Metabolomics in plant environmental physiology. J. Exp. Bot. 2013, 64, 4011–4020. [Google Scholar] [CrossRef]
- Trenkamp, S.; Eckes, P.; Busch, M.; Fernie, A.R. Temporally resolved GC-MS-based metabolic profiling of herbicide treated plants treated reveals that changes in polar primary metabolites alone can distinguish herbicides of differing mode of action. Metabolomics 2009, 5, 277–291. [Google Scholar] [CrossRef]
- García-Flores, M.; Juárez-Colunga, S.; García-Casarrubias, A.; Trachsel, S.; Winkler, R.; Tiessen, A. Metabolic profiling of plant extracts using direct-injection electrospray ionization mass spectrometry allows for high-throughput phenotypic characterization according to genetic and environmental effects. J. Agric. Food Chem. 2015, 63, 1042–1052. [Google Scholar] [CrossRef]
- Torres-García, J.R.; Tafoya-Razo, J.A.; Velázquez-Márquez, S.; Tiessen, A. Double herbicide-resistant biotypes of wild oat (Avena fatua) display characteristic metabolic fingerprints before and after applying ACCase- and ALS-inhibitors. Acta Physiol. Plant. 2018, 40, 119. [Google Scholar] [CrossRef]
- Delye, C. Weed Resistance to Acetyl Coenzyme A Carboxylase Inhibitors: An Update. Weed Sci. 2005, 53, 728–746. [Google Scholar] [CrossRef]
- Somody, C.N.; Nalewaja, J.D.; Miller, S. Wild Oat (Avena fatua) and Avena sterilis Morphological Characteristics and Response to Herbicides. Weed Sci. 1984, 32, 353–359. [Google Scholar] [CrossRef]
- Fukusaki, E.; Kobayashi, A. Plant Metabolomics: Potential for Practical Operation. J. Biosci. Bioeng. 2005, 100, 347–354. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Chrysayi-Tokousbalides, M. Metabolomics in pesticide research and development: Review and future perspectives. Metabolomics 2011, 7, 35–53. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Cawthray, G.R.; Wang, S.F.; Powles, S.B. Enhanced rates of herbicide metabolism in low herbicide-dose selected resistant Lolium rigidum. Plant Cell Environ. 2013, 36, 818–827. [Google Scholar] [CrossRef]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 23 July 2019).
- Tafoya-Razo, J.A.; Núñez-Farfán, J.; Torres-García, J.R. Migration by seed dispersal of ACCase-inhibitor-resistant Avena fatua in north-western Mexico. Pest Manag. Sci. 2017, 73, 167–173. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tafoya-Razo, J.A.; Oregel-Zamudio, E.; Velázquez-Márquez, S.; Torres-García, J.R. 10,000-Times Diluted Doses of ACCase-Inhibiting Herbicides Can Permanently Change the Metabolomic Fingerprint of Susceptible Avena fatua L. Plants. Plants 2019, 8, 368. https://doi.org/10.3390/plants8100368
Tafoya-Razo JA, Oregel-Zamudio E, Velázquez-Márquez S, Torres-García JR. 10,000-Times Diluted Doses of ACCase-Inhibiting Herbicides Can Permanently Change the Metabolomic Fingerprint of Susceptible Avena fatua L. Plants. Plants. 2019; 8(10):368. https://doi.org/10.3390/plants8100368
Chicago/Turabian StyleTafoya-Razo, J António, Ernesto Oregel-Zamudio, Sabina Velázquez-Márquez, and Jesús R. Torres-García. 2019. "10,000-Times Diluted Doses of ACCase-Inhibiting Herbicides Can Permanently Change the Metabolomic Fingerprint of Susceptible Avena fatua L. Plants" Plants 8, no. 10: 368. https://doi.org/10.3390/plants8100368
APA StyleTafoya-Razo, J. A., Oregel-Zamudio, E., Velázquez-Márquez, S., & Torres-García, J. R. (2019). 10,000-Times Diluted Doses of ACCase-Inhibiting Herbicides Can Permanently Change the Metabolomic Fingerprint of Susceptible Avena fatua L. Plants. Plants, 8(10), 368. https://doi.org/10.3390/plants8100368