Seed and Germination Characteristics of 20 Amazonian Liana Species

Abstract
:1. Introduction
2. Results and Discussion
2.1. Species with Sufficient Replicates from Various Mother Plants
2.2. Seed Biometrics and Germination at Constant Temperature (25°C)—All Species

{kind=link}
{kind=link}
{kind=link}
| Species with replicates | INPA herbarium number | family- subfamily | Treatment | Replicates x seed number per treatment | mother plants used for seed sampling | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| fresh seed | dried seed | |||||||||
| 25 °C L | 20/30 °C L | 15/35 °C L | 25 °C D | 25 °C L | ||||||
| Anemopaegma oligoneuron (Sprague & Sandw.) Gentry | 231927 | Bignoniaceae | 96 ± 8a | 98 ± 5a | 92 ± 10a | 71 ± 24 | 66 ± 18 | 25 × 10 | 5 | |
| Arrbidaea trailii Sprague | 231926 | Bignoniaceae | 96 ± 6a | 100 ± 0a | 94 ± 9a | 92 ± 9 | 86 ± 8 | 4 × 16 | 3 | |
| Mimosa guilandinae var. spruceana (Benth.) Barneby | 231932 | Fabaceae-Mimosoidae | 32 ± 19a | 33 ± 21a | 35 ± 22a | 23 ± 12 | 36 ± 23 | 10 × 20 | 5 | |
| Passiflora nitida HBK | 231937 | Passifloraceae | 20 ± 22a *** | 0 ± 0b | 0 ± 0b | 14 ± 8 | 6 ± 7 | 8 × 25 | 4 | |
| Senna sp.2 | 231922 | Fabaceae-Caesalpinioideae | 100 ± 0a | 100 ± 0a | 98 ± 4a | 94 ± 8 | 89 ± 6 | 9 × 12 | 3 | |
| Species with pseudoreplicates or small replicates | ||||||||||
| Acacia multipinnata Ducke | Fabaceae-Mimosoidae | 80 | 80 | 90 | 70 | 40 | 1 × 10 | 1 | ||
| Anemopaegma floridum Mart. ex DC | 231931 | Bignoniaceae | 94 ± 9 | 94 ± 9 | 100 ± 0 | 38 ± 0 | 0 ± 0 | 2 × 10 | 1 | |
| Anomospermum solimoesanum (Moldenke) Krukoff& Barneby | 231935 | Menispermaceae | 90 ± 14 | 40 ± 30 | 95 ± 11 | 44 ± 11 | 0 ± 0 | 5 × 4 | 3 | |
| Aristolochia silvatica Barb. Rodr. | 231928 | Aristolochiaceae | 90 ± 10a | 88 ± 16a | 0 ± 0b ** | 0 ± 0 | 72 ± 8 | 3 × 20 | 1 | |
| Cissus sicyoides L. | 231923 | Vitaceae | 93 ± 2a | 100 ± 0a | 95 ± 6a | 25 ± 8 | 67 ± 30 | 4 × 25 | 1 | |
| Coccoloba sp. | 231936 | Polygonaceae | 46 ± 13a | 44 ± 20a | 5 ± 4b * | 41 ± 8.5 | 0 ± 0 | 4 × 20 | 1 | |
| Gnetum c.f. nodiflorum Brongn. | 231933 | Gnetaceae | 7 ± 12 | 13 ± 11 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 3 × 5 | 1 | |
| Matelea badilloi Morillo | 231924 | Apocynaceae-Asclepiadoideae | 61 ± 20a | 98 ± 3b | 0 ± 0c *** | 0.6 ± 1.6 | 63 ± 34 | 6 × 25 | 1 | |
| Mucuna sp. | 231940 | Fabaceae- Fabaoidae | 67 ± 0 | na | na | na | 0 ± 0 | 2 × 3 | 1 | |
| Passiflora c.f. acuminata DC. | Passifloraceae | 43 ± 61 | na | na | 43 ± 33 | 20 ± 19 | 2 × 15 | 1 | ||
| Paullinia c.f. capreolata (Aubl.) Radk | 231939 | Sapindaceae | 100 ± 0 | 100 ± 0 | 100 ± 0 | 95 ± 1 | 0 ± 0 | 2 × 20 | 1 | |
| Paullinia rugosa Benth. ex Radk | 231938 | Sapindaceae | 100 ± 0 | na | na | 95 ± 7 | 0 ± 0 | 2 × 10 | 1 | |
| Smilax sp. | 231930 | Smilacaceae | 60 | na | na | 60 | 0 | 1 × 10 | 1 | |
| Strychnos c.f. amazonica Krukoff | 231925 | Loganiaceae | 50 | na | na | 70 | 0 | 1 × 10 | 1 | |
| Strychnos glabra Sagot ex Progl. | 231929 | Loganiaceae | 65 ± 7 | 75 ± 7 | 25 ± 0 | 65 ± 7 | 0 ± 0 | 2 × 10 | 1 | |
| Species with replicates | fresh seed mass (g) | n | seed moisture (%) | n | longest axis (mm) | n | seed shape | P (D-S) | n | G (DT-DS) | RLG | germination time (d) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 50% | ||||||||||||||
| M. guilandinae | 0.016 ± 0.005 | 30 | 13.2 ± 5.2 | 30 | 7.0 ± 0.9 | 60 | flat | 0.055 | 20 | DT | 0.58 | 5 | 9 | ||
| Senna sp. 2 | 0.031 ± 0.008 | 50 | 34.6 ± 3.3 | 50 | 6.4 ± 1.0 | 50 | oval | 0.002 | 20 | DT | 0.52 | 2 | 2.5 | ||
| P. nitida | 0.033 ± 0.006 | 50 | 32.0 ± 3.4 | 50 | 7.0 ± 0.7 | 50 | flat | 0.001 | 20 | DT | 0.58 | 108 | 156 | ||
| A. trailii | 0.035 ± 0.008 | 41 | 29.9 ± 8.8 | 41 | 29.2 ± 4.4 | 41 | winged & flat | 0.037 | 20 | DT | 0.51 | 11 | 23 | ||
| A. oligoneuron | 0.191 ± 0.042 | 60 | 65.2 ± 3.6 | 60 | 38.7 ± 4.3 | 60 | winged & flat | 0.026 | 23 | DT | 0.57 | 14 | 16 | ||
| Species with pseudoreplicates or small replicates | |||||||||||||||
| M. badilloi | 0.016 ± 0.002 | 50 | 48.4 ± 2.9 | 50 | 8.9 ± 0.4 | 50 | flat | 0.001 | 20 | DT | 0.99 | 5 | 15 | ||
| P. acuminata | 0.021 ± 0.005 | 15 | 47.6 ± 11.6 | 14 | 7.3 ± 0.3 | 30 | flat | 0.017 | 18 | DT | 0.68 | 92 | 105 | ||
| C. sicyoides | 0.031 ± 0.002 | 30 | 31.1 ± 3.2 | 30 | 5.1 ± 0.3 | 30 | drop like | 0.010 | 20 | DT | 0.79 | 11 | 24 | ||
| A. multipinnata | 0.037 ± 0.008 | 30 | 24.2 ± 3.5 | 14 | 6.4 ± 0.7 | 30 | flat | 0.068 | 21 | DT | 0.53 | 3 | 3 | ||
| A. silvatica | 0.035 ± 0.004 | 30 | 30.4 ± 3.2 | 30 | 18.6 ± 1.9 | 30 | winged & flat | 0.022 | 21 | DT | 1.00 | 10 | 84 | ||
| A .floridum | 0.396 ± 0.037 | 30 | 65.0 ± 3.7 | 10 | 52.9 ± 4.8 | 30 | winged & flat | 0.060 | 15 | DS | 0.71 | 21 | 31 | ||
| Coccoloba sp. | 0.238 ± 0.051 | 51 | 40.5 ± 3.8 | 51 | 7.7 ± 0.6 | 51 | round | 0.222 | 20 | DS | 0.53 | 31 | 70 | ||
| P. rugosa | 0.464 ± 0.097 | 30 | 42.3 ± 3.9 | 10 | 10.0 ± 0.8 | 30 | round | 0.780 | 7 | DS | 0.51 | 9 | 12 | ||
| S. amazonica | 0.691 ± 0.221 | 30 | 52.1 ± 3.1 | 11 | 14.9 ± 1.8 | 30 | triangular | 0.523 | 9 | DS | 0.51 | 20 | 44 | ||
| S. glabra | 0.776 ± 0.101 | 30 | 56.2 ± 3.1 | 10 | 12.4 ± 0.7 | 30 | triangular | 0.814 | 9 | DS | 0.42 | 20 | 27 | ||
| Smilax sp. | 0.584 ± 0.123 | 30 | 41.6 ± 3.1 | 10 | 11.3 ± 1.2 | 30 | triangular | 0.770 | 7 | DS | 0.50 | 20 | 29 | ||
| P. capreolata | 0.477 ± 0.051 | 10 | 28.9 ± 3.1 | 10 | 13.3 ± 0.5 | 30 | oval | 0.814 | 10 | DS | 0.50 | 4 | 6 | ||
| G. nodiflorum | 12.19 ± 2.346 | 46 | 50.2 ± 3.4 | 19 | 43.2 ± 3.1 | 46 | oval | 0.977 | 12 | DS | 1.00 n = 1 | 155 n = 1 | 155 n = 1 | ||
| A. solimoesanum | 13.97 ± 2.603 | 84 | 32.0 ± 3.2 | 22 | 36.1 ± 2.6 | 84 | oval | 0.543 | 20 | DS | 0.67 | 91 | 157 | ||
| Mucuna sp. | 34.16 ± 12.002 | 25 | 67.8 ± 3.2 | 9 | 38.8 ± 5.7 | 25 | round | 0.984 | 10 | DS | na | 53 n = 4 | 60 n = 4 | ||
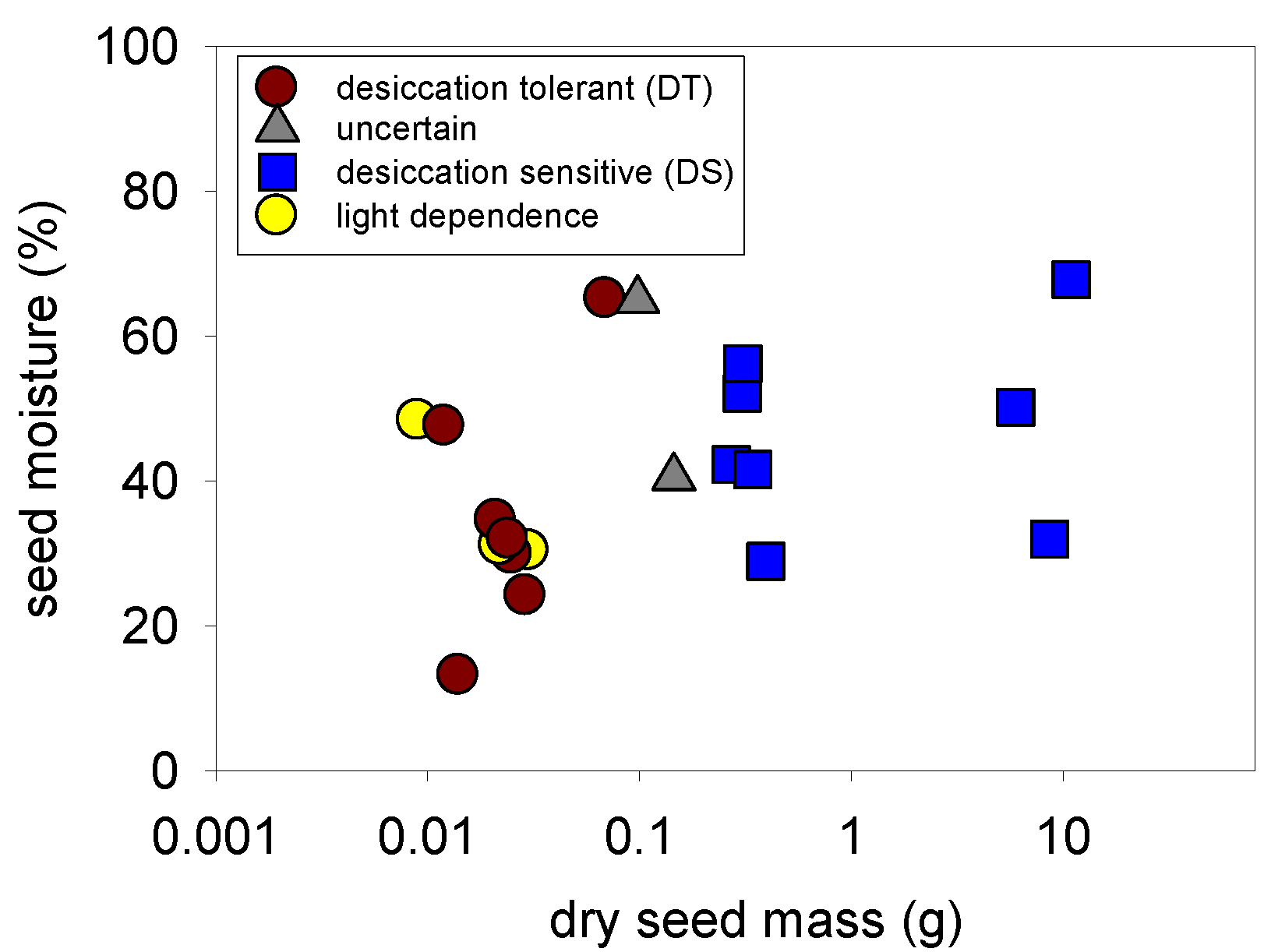
2.3. Desiccation Tolerance—All Species
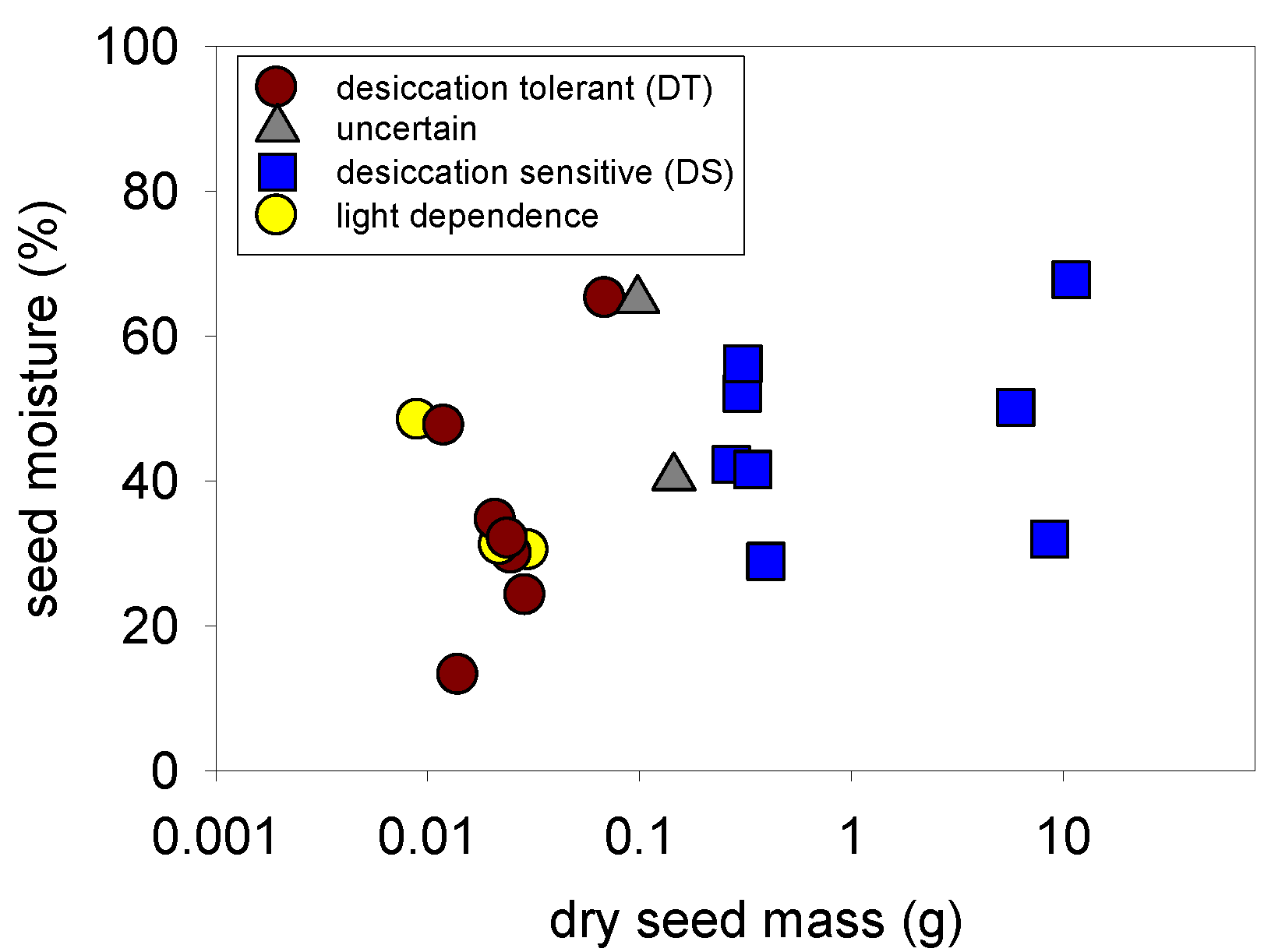
2.4. Light Germination—All Species
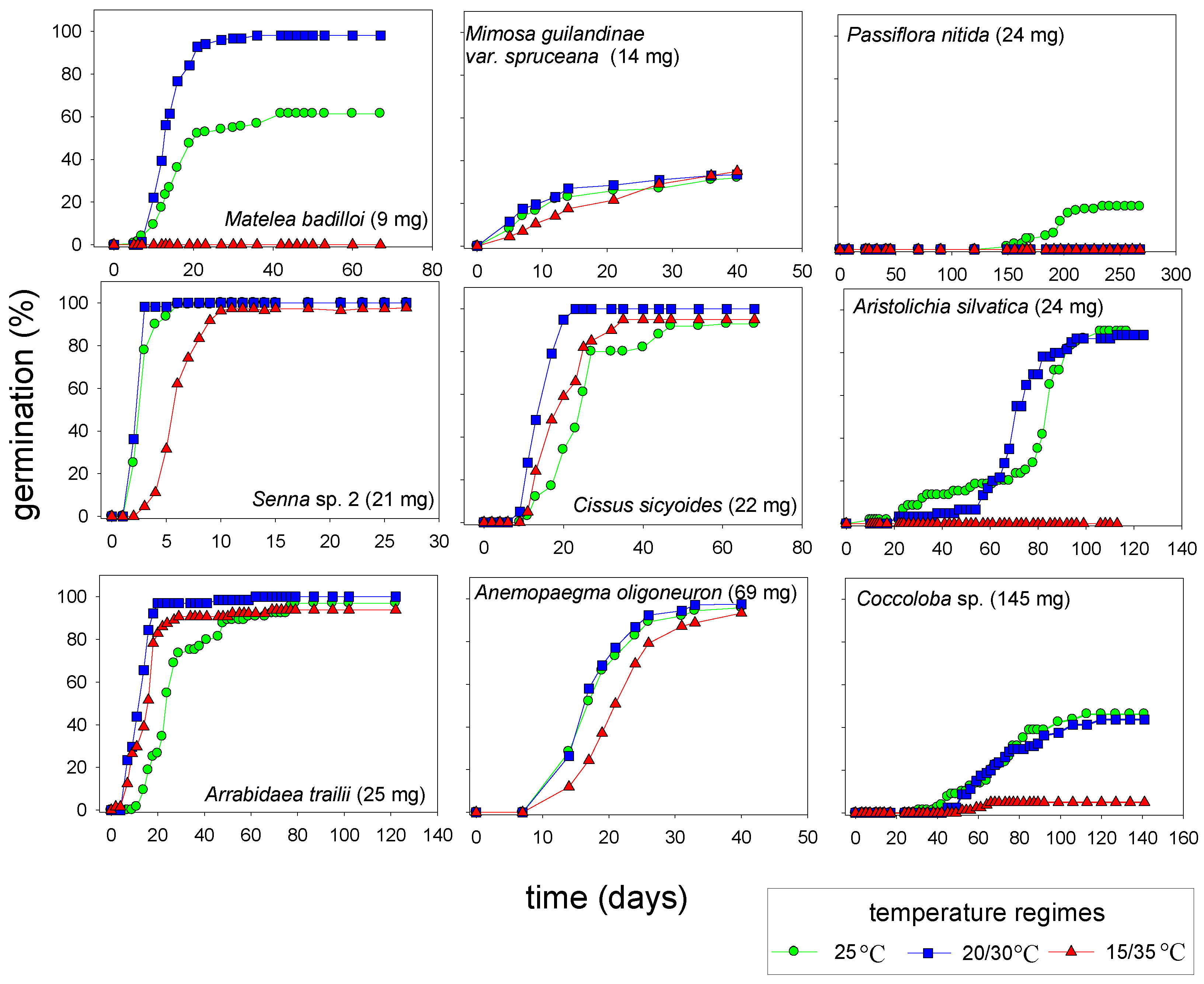
2.5. Germination at Alternating Temperatures
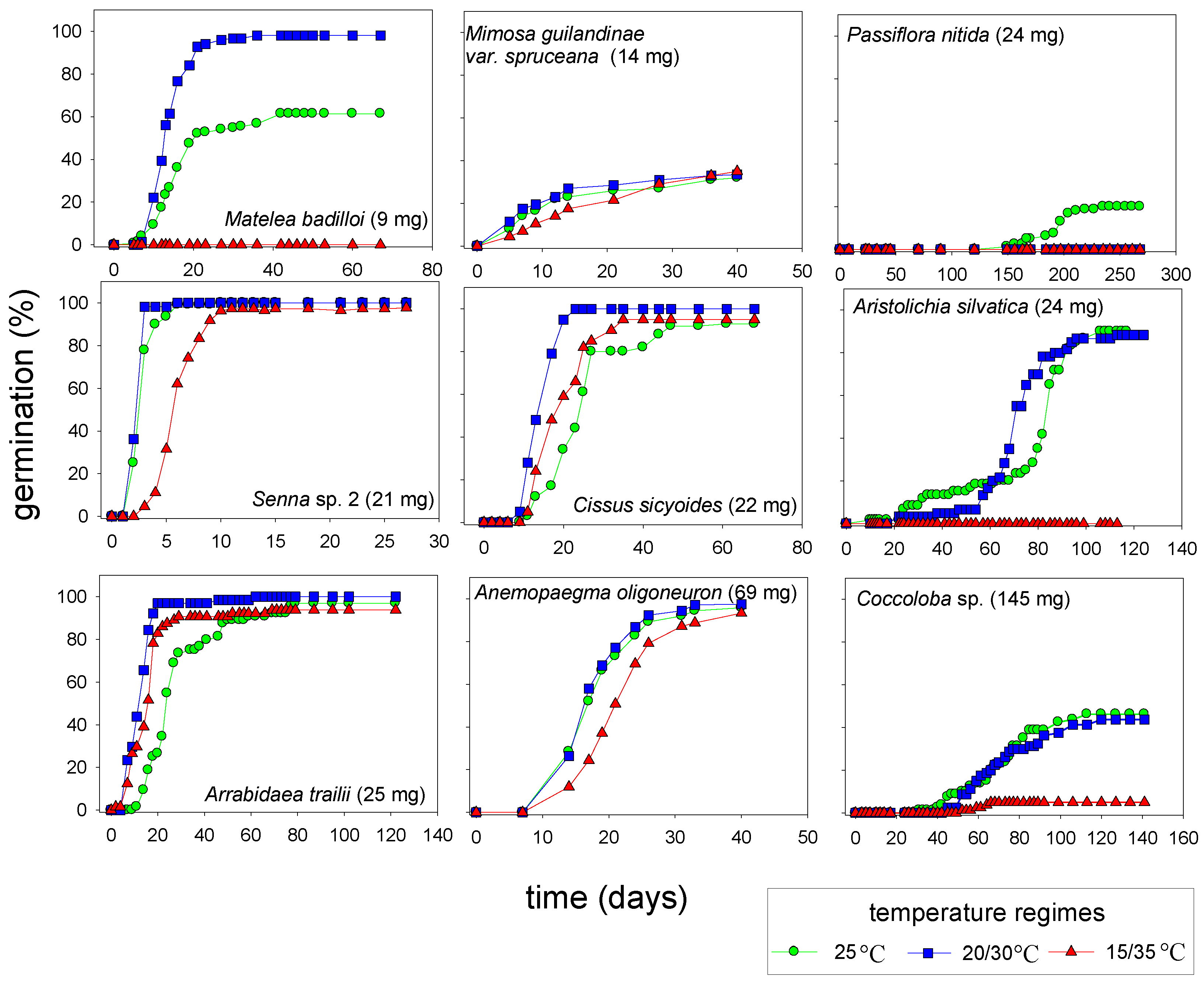
3. Experimental Section
3.1. Seed Collection and Biometry
3.2. Desiccation Tolerance
3.3. Germination Test
3.4. Data Analysis
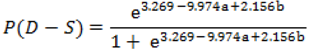
4. Conclusions
Acknowledgments

References
- Putz, F.E. The Biology of Vines; Putz, F.E., Mooney, H.A., Eds.; Cambridge University Press: Cambridge, UK, 1991; p. 501. [Google Scholar]
- Emmons, L.H.; Gentry, A.H. Tropical forest structure and the distribution of gliding and prehensile-tailed vertebrates. Am. Nat. 1983, 121, 513–524. [Google Scholar]
- Odegaard, F. The relative importance of trees versus lianas as hosts for phytophagous beetles (Coleoptera) in tropical forests. J. Biogeogr. 2000, 27, 283–296. [Google Scholar] [CrossRef]
- Putz, F.E. The natural history of lianas on Barro Colorado Island, Panama. Ecology 1984, 65, 1713–1724. [Google Scholar] [CrossRef]
- Kainer, K.A.; Wadt, L.H.O.; Gomes-Silva, D.A.P.; Capanu, M. Liana loads and their association with Bertholletia excelsa fruit and nut production, diameter growth and crown attributes. J. Trop. Ecol. 2006, 22, 147–154. [Google Scholar]
- Schnitzer, S.A.; Kuzee, M.E.; Bongers, F. Disentangling above- and below-ground competition between lianas and trees in a tropical forest. J. Ecol. 2005, 93, 1115–1125. [Google Scholar] [CrossRef]
- Laurance, W.F.; Perez-Salicrup, D.; Delamonica, P.; Fearnside, P.M.; D’Angelo, S.; Jerozolinski, A.; Pohl, L.; Lovejoy, T.E. Rain forest fragmentation and the structure of Amazonian liana communities. Ecology 2001, 82, 105–116. [Google Scholar]
- Phillips, O. The ethnobotany and econmic botany of tropical vines. In The biology of Vines; Putz, F.E., Mooney, H.A., Eds.; Cambridge University Press: Cambridge, UK, 1991; pp. 427–475. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds; Academic Press: San Diego, CA, USA, 1998; p. 271. [Google Scholar]
- Sanches, M.C.; Válio, F.M. Seed and seedling survival of some climber species in a southeast Brazilian tropical forest. Biotropica 2002, 34, 323–327. [Google Scholar]
- Gerwing, J.J. Life history diversity among six species of canopy lianas in an old-growth forest of the eastern Brazilian Amazon. For. Ecol. Manage. 2004, 190, 57–72. [Google Scholar] [CrossRef]
- Dupuy, J.M.; Chazdon, R.L. Effects of vegetation cover on seedling and sapling dynamics in secondary tropical wet forests in Costa Rica. J. Trop. Ecol. 2006, 22, 65–76. [Google Scholar]
- Hegarty, E.E.; Caballé, G. Distribution and abundance of vines in forest communities. In The Biology of Vines; Putz, F.E., Mooney, H.A., Eds.; Cambridge University Press: Cambridge, UK, 1991; pp. 313–335. [Google Scholar]
- DeWalt, S.J.; Schnitzer, S.A.; Denslow, J.S. Density and diversity of lianas along a chronosequence in a central Panamanian lowland forest. J. Trop. Ecol. 2000, 16, 1–19. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Carson, W.P. Treefall gaps and the maintenance of species diversity in a tropical forest. Ecology 2001, 82, 913–919. [Google Scholar] [CrossRef]
- Roeder, M.; Hölscher, D.; Ferraz, I.D.K. Liana regeneration in secondary and primary forest of Central Amazonia. Plant Ecol. Div. 2010, 3, 165–174. [Google Scholar] [CrossRef]
- Pearson, T.R.H.; Burslem, D.F.R.P.; Mullins, C.E.; Dalling, J.W. Germination ecology of neotropical pioneers: Interacting effects of environmental conditions and seed size. Ecology 2002, 83, 2798–2807. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, A.P.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Letcher, S.G.; Chazdon, R.L. Life history traits of lianas during tropical forest succession. Biotropica 2012, 44, 720–727. [Google Scholar] [CrossRef]
- Liu, K.; Eastwood, R.J.; Flynn, S.; Turner, R.M.; Stuppy, W.H. Seed Information Database (release 7.1, May 2008) 2008. Available online: http://data.kew.org/sid/ (accessed on 24 October 2012).
- Pritchard, H.W.; Wood, C.B.; Hodges, S.S.; Vautier, H.J. 100-seed test for desiccation tolerance and germination: A case study on eight tropical palm species. Seed Sci. Technol. 2004, 32, 393–403. [Google Scholar]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Prediction of desiccation sensitivity in seeds of woody species: A probabilistic model based on two seed traits of 104 species. Ann. Bot. 2006, 97, 667–674. [Google Scholar] [CrossRef]
- Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers, C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Martins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.; et al. Flora da Reserva Ducke- Guia de identificação das plantas vasculares de uma floresta de terra-firme na Amazônia Central; INPA-DFID: Manaus, Brazil, 1999; p. 799. [Google Scholar]
- Milberg, P.; Andersson, L.; Thompson, K. Large-seeded species are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef]
- Missouri Botanical Garden. TROPICOS. Botanical information system at the Missouri Botanical Garden. Available online: http://www.tropicos.org/ (accessed on 25 October 2012).
- Passos, I.R.S.; Matos, G.V.C.; Meletti, L.M.M.; Scott, M.D.S.; Bernacci, L.C.; Vieira, M.A.R. Utilização do ácido giberélico para a quebra de dormência de sementes de Passiflora nitida Kunth germinadas in vitro. Rev. Bras. Frutic. 2004, 26, 380–381. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An intermediate category of seed storage behaviour? I. Coffee. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Black, M.; Bewley, J.D.; Halmer, P. The Encyclopedia of Seeds-Science, Technology and Uses; CABI: Wallingford, UK, 2006; p. 354. [Google Scholar]
- Hong, T.D.; Ellis, R.H. A Protocol to Determine Seed Storage Behaviour; IPGRI Technical Bulletin 1; International Plant Genetic Resource Institute: Rome, Italy, 1996; p. 64. [Google Scholar]
- Lleras, E. Neglected crops: 1492 from a different perspective. In FAO, Plant Production and Protection Series, No.26; Hernándo Bermejo, J.E., León, J., Eds.; FAO: Rome, Italy, 1994; pp. 223–228. [Google Scholar]
- Sivakumar, V.; Anandalakshmi, R.; Warrier, R.R.; Tigabu, M.; Odén, P.C.; Vijayachandran, S.N.; Geetha, S.; Singhet, B.G. Effects of presowing treatments, desiccation and storage conditions on germination of Strychnos nux-vomica seeds, a valuable medicinal plant. New Forest 2006, 32, 121–131. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Berjak, P. Evolutionary and ecological aspects of recalcitrant seed biology. Seed Sci. Res. 2000, 10, 301–306. [Google Scholar]
- Hong, T.D.; Ellis, R.H. Contrasting seed storage behaviour among different species of Meliaceae. Seed Sci. Technol. 1998, 26, 77–95. [Google Scholar]
- Recalcitrant seeds and intermediates. Agroforest Seed Circ. 1993, 3, 22–26.
- Hong, T.D.; Ellis, R.H. The effect of the initial rate of drying on the subsequent ability of immature seeds of Norway maple (Acer plantanoides) to survive rapid desiccation. Seed Sci. Res. 1997, 7, 41–45. [Google Scholar]
- Janzen, D.H. Reduction of Mucuna andreana (Leguminosae) seedling fitness by artifical seed damage. Ecology 1976, 57, 826–828. [Google Scholar] [CrossRef]
- Jankowska-Blaszczuk, M.; Daws, M.I. Impact of red: far red ratios on germination of temperate forest herbs in relation to shade tolerance, seed mass and persistence in the soil. Funct. Ecol. 2007, 21, 1055–1062. [Google Scholar] [CrossRef]
- Cuzzol, G.R.F.; Lucas, M.N.C. Germinação de sementes de Matelea maritima (Jack.) Woods (Asclepiadaceae). Rev. Bras. Bot. 1999, 22, 1–7. [Google Scholar] [CrossRef]
- Probert, R.J. The role of temperature in germination ecophysiology. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; pp. 261–292. [Google Scholar]
- INMET-Instituto Nacional de Meteorologia. Ministério da Agricultura, Pecuária e Abastecimento. Available online: http://www.inmet.gov.br/portal/ (accessed on 18 March 2010).
- Gerwing, J.J.; Schnitzer, S.A.; Burnham, R.J.; Bongers, F.; Chave, J.; DeWalt, S.J.; Ewango, C.E.N.; Foster, R.; Kenfack, D.; Martínez-Ramos, M.; et al. A standard protocol for liana censuses. Biotropica 2006, 38, 256–261. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Roeder, M.; Ferraz, I.D.K.; Hölscher, D. Seed and Germination Characteristics of 20 Amazonian Liana Species. Plants 2013, 2, 1-15. https://doi.org/10.3390/plants2010001
Roeder M, Ferraz IDK, Hölscher D. Seed and Germination Characteristics of 20 Amazonian Liana Species. Plants. 2013; 2(1):1-15. https://doi.org/10.3390/plants2010001
Chicago/Turabian StyleRoeder, Mareike, Isolde D. K. Ferraz, and Dirk Hölscher. 2013. "Seed and Germination Characteristics of 20 Amazonian Liana Species" Plants 2, no. 1: 1-15. https://doi.org/10.3390/plants2010001
APA StyleRoeder, M., Ferraz, I. D. K., & Hölscher, D. (2013). Seed and Germination Characteristics of 20 Amazonian Liana Species. Plants, 2(1), 1-15. https://doi.org/10.3390/plants2010001