

Isolation and Characterization of Plant-Growth-Promoting, Drought-Tolerant Rhizobacteria for Improved Maize Productivity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Collection of Soil Samples
2.2. Isolation of Culturable Rhizobacteria
2.3. Morphological, Microscopic, and Biochemical Characterization of Rhizobacteria
2.4. Drought Tolerance Screening of the Rhizobacteria
2.5. Screening of Rhizobacteria for Plant-Growth-Promoting Traits
2.5.1. Ammonia Production
2.5.2. Phosphate Solubilization
2.5.3. Siderophore Production
2.5.4. Nitrogen Fixation
2.5.5. Indole-Acetic Acid (IAA) Production
2.5.6. Hydrogen Cyanide (HCN) Determination
2.5.7. 1-Aminocyclopropane-1-carboxylate (ACC) Deaminase Activity
2.6. Tolerance of Bacterial Isolates to pH, Temperatures, Salt Concentrations, and Heavy Metals
2.6.1. Bacterial Tolerance to Different pH
2.6.2. Tolerance of Bacterial Isolates to Different Temperatures
2.6.3. Bacterial Growth Response to Different Salt Concentrations
2.6.4. Bacterial Tolerance to Different Heavy Metals
2.7. Extraction of DNA and Genotypic Identification
2.7.1. Polymerase Chain Reaction (PCR) Amplification
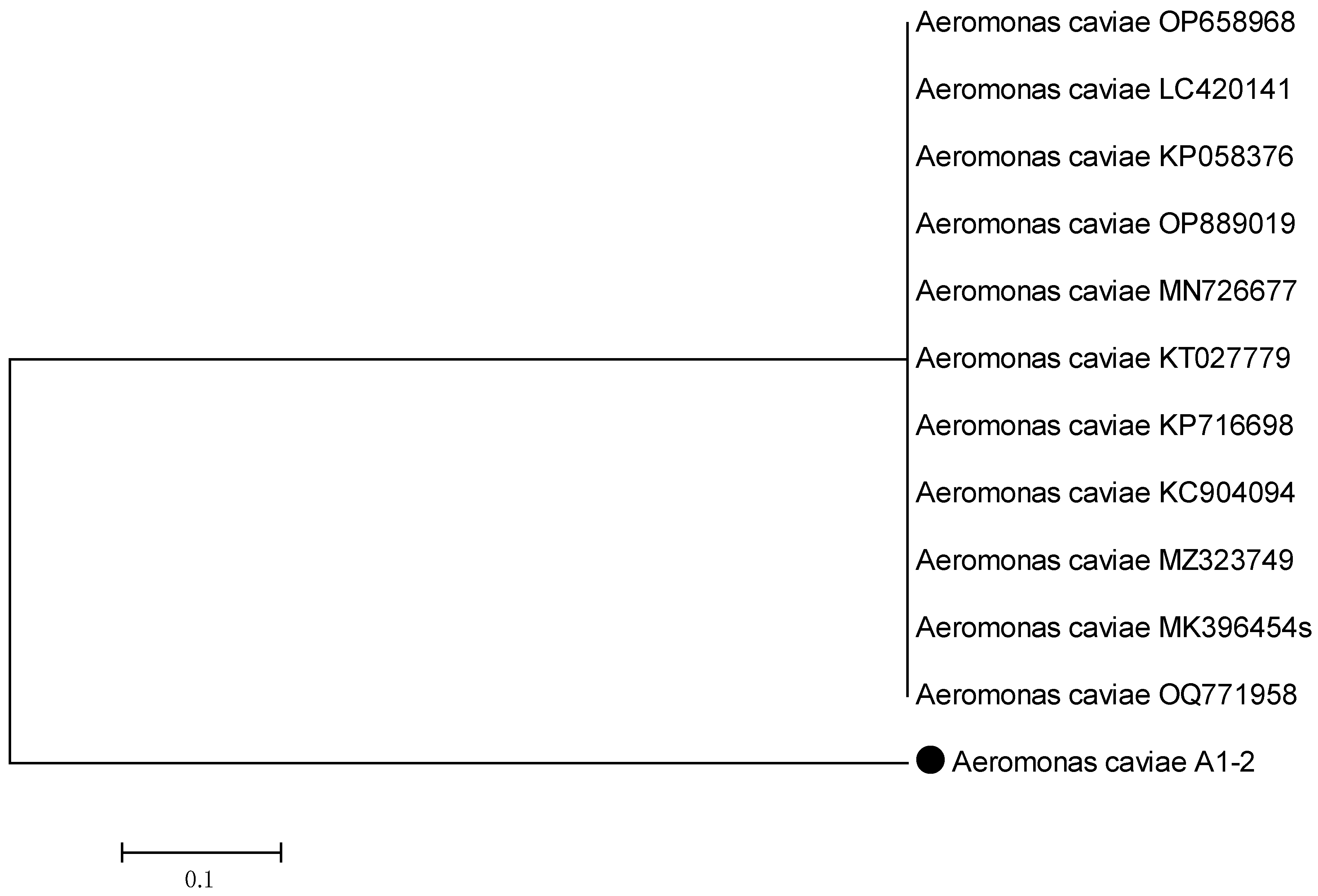
2.7.2. Alignment Sequence and Construction of the Phylogenetic Tree
2.8. Experimental Set-up and Maize Growth Conditions
Below-Ground–Above-Ground/Morphological Parameters
2.9. Parameters Measured
2.9.1. Leaf Relative Water Content
2.9.2. Electrolyte Leakage
2.9.3. Soluble Sugar Content in Maize Leaves
2.9.4. Proline Content in Maize Leaves
2.10. Statistical Analyses
2.11. Data Availability Statement
3. Results
3.1. Characterization of the Rhizobacteria Strains
3.2. Selection of Drought-Tolerant Rhizobacteria Strains
3.3. Plant-Growth-Promoting Abilities of Rhizobacterial Strains and Phylogeny
3.4. Rhizobacteria Tolerance to Drought Stress under Different Growth Conditions
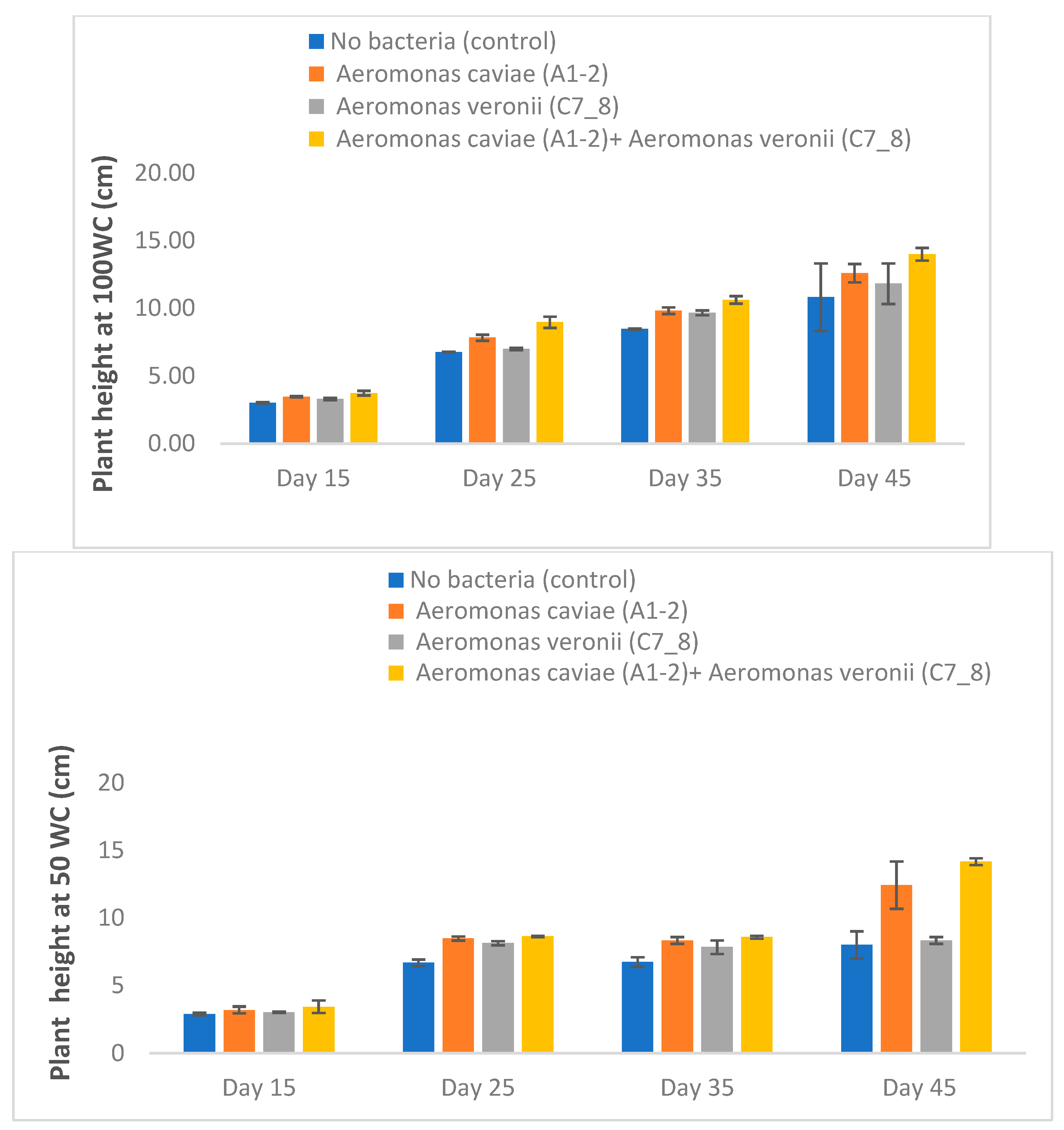
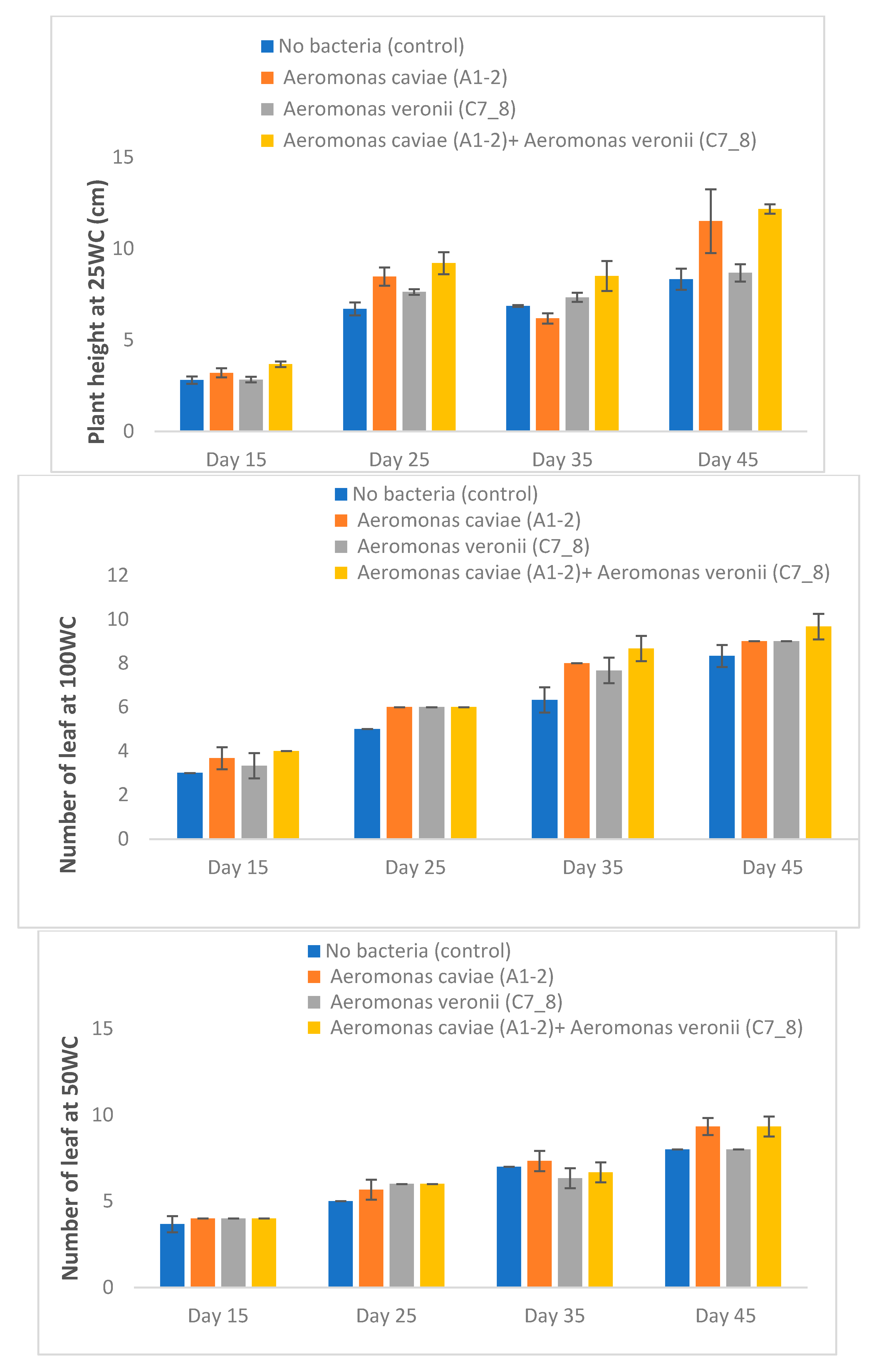
3.5. Rhizobacteria Improve the Growth of Maize Plants Exposed to Varying Levels of Drought Stress in a Greenhouse
3.6. Rhizobacteria Improving the Biomass of Maize Exposed to Drought Stress in the Greenhouse
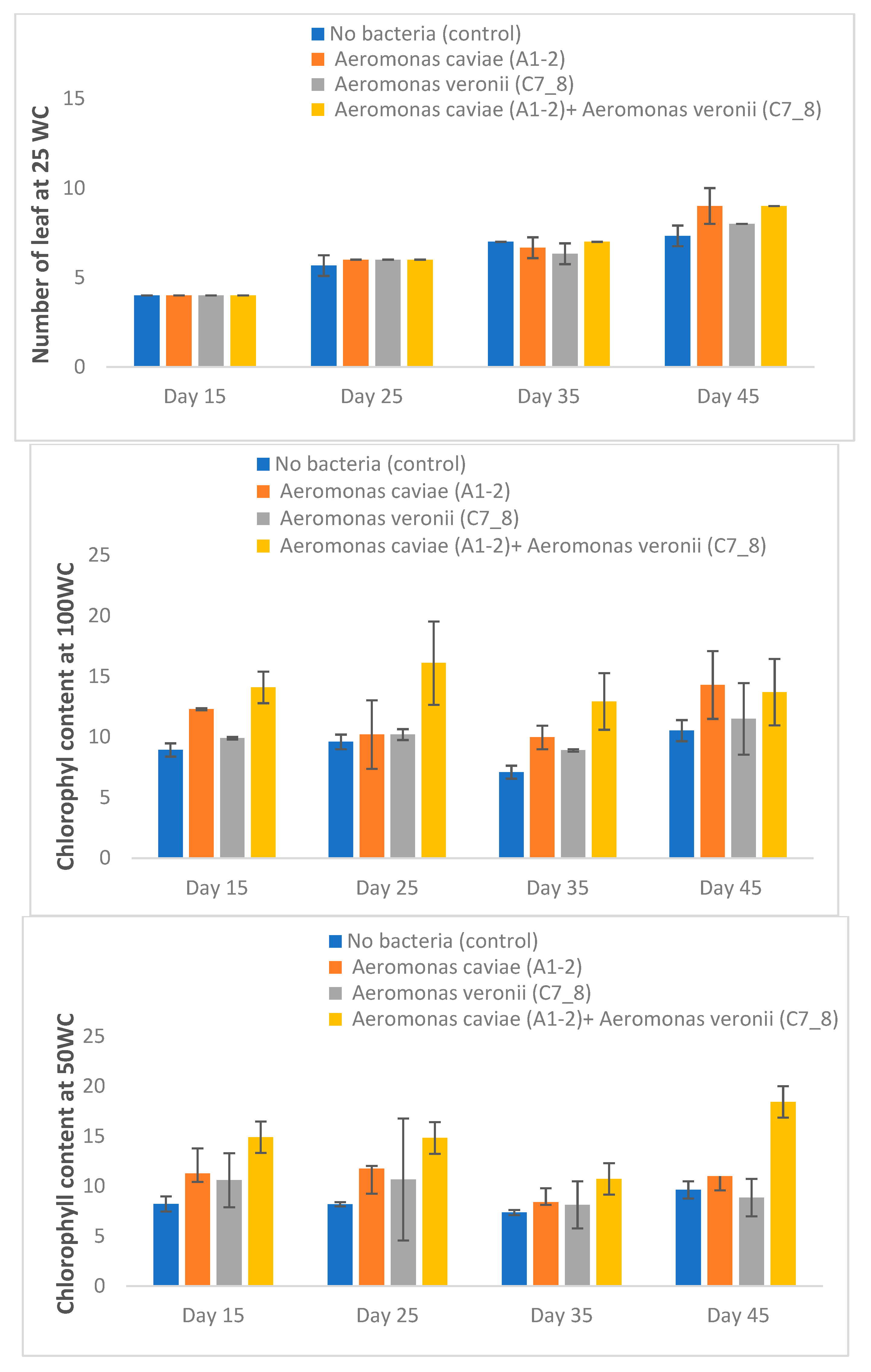
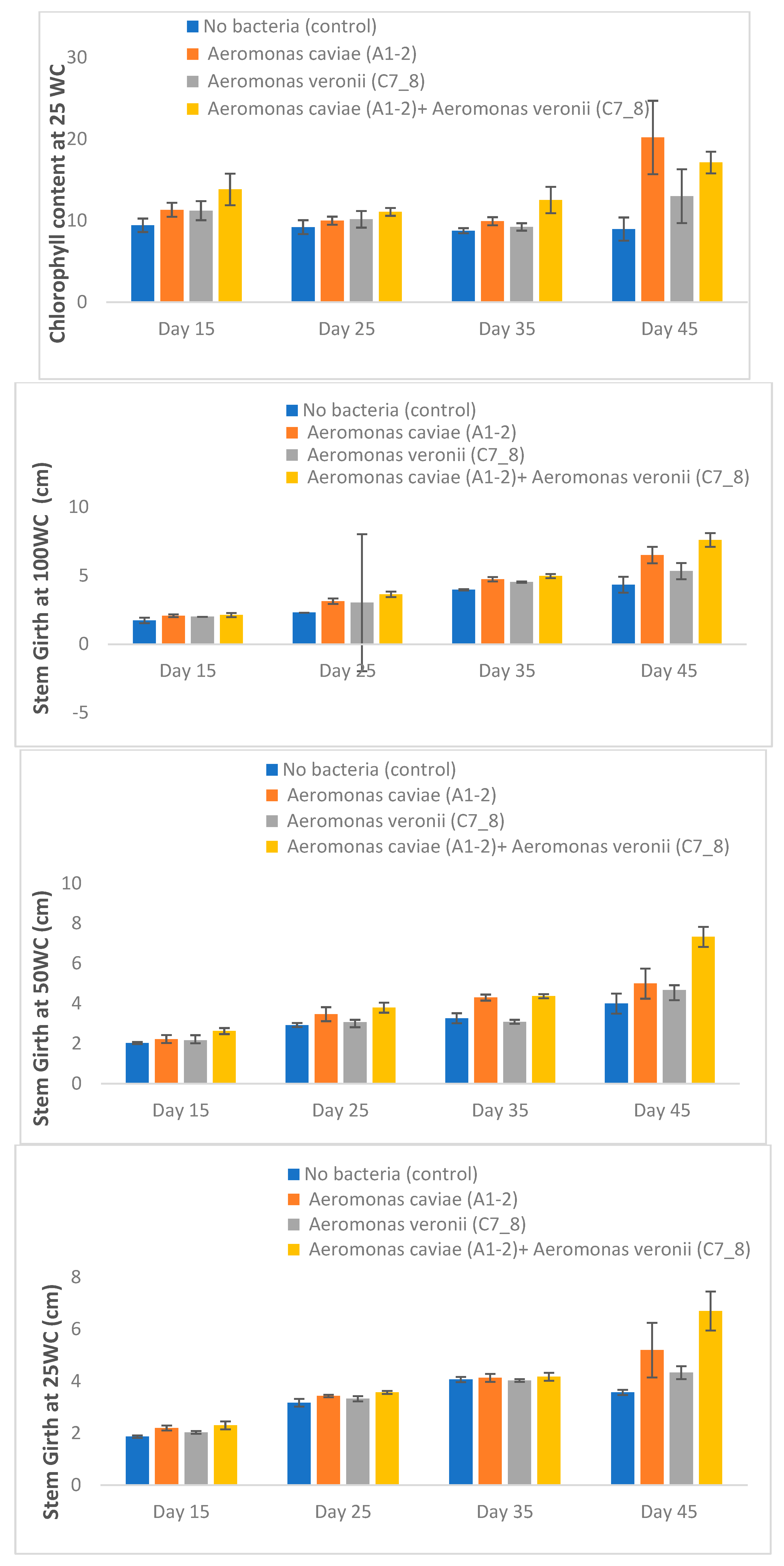
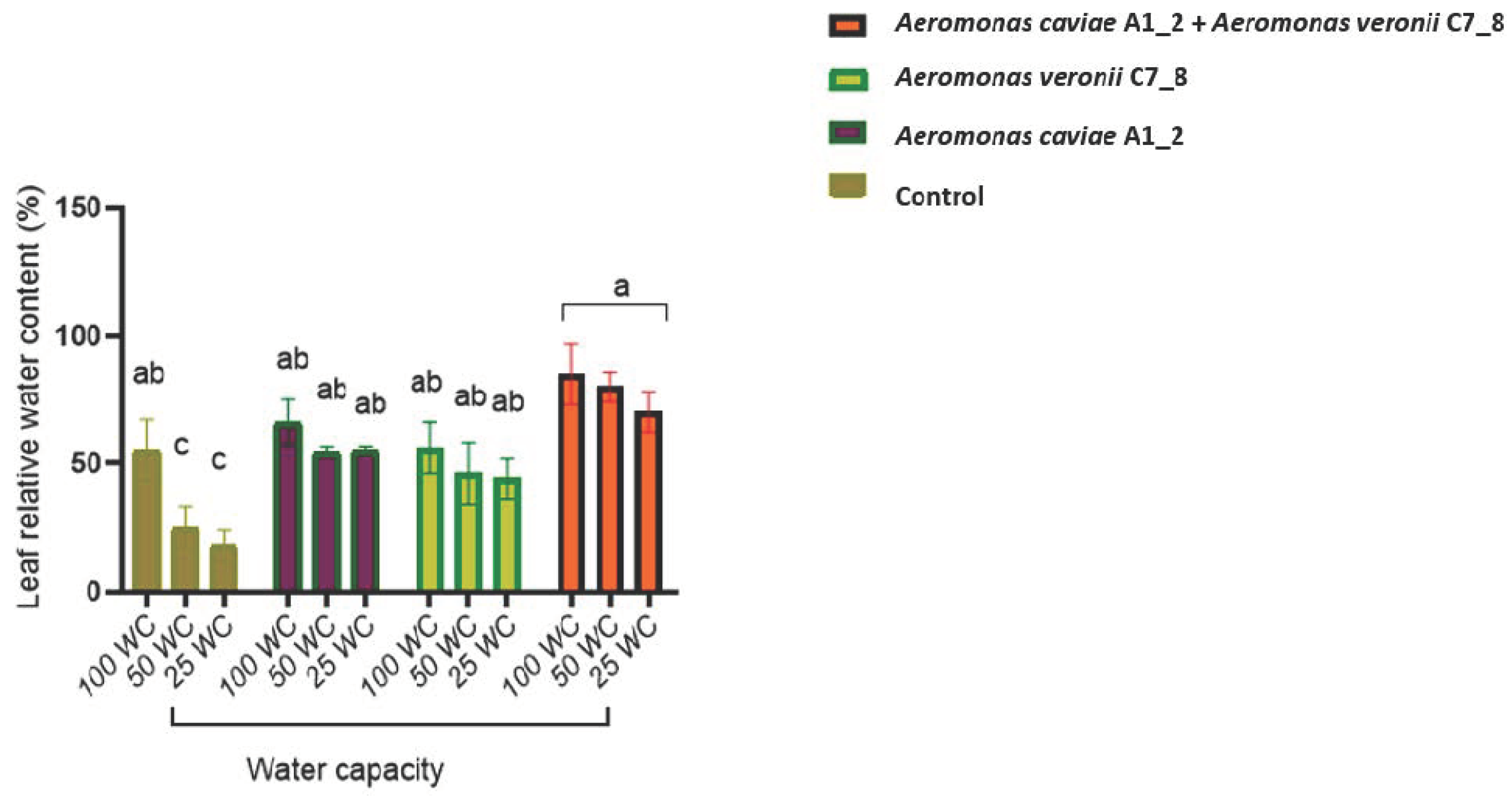
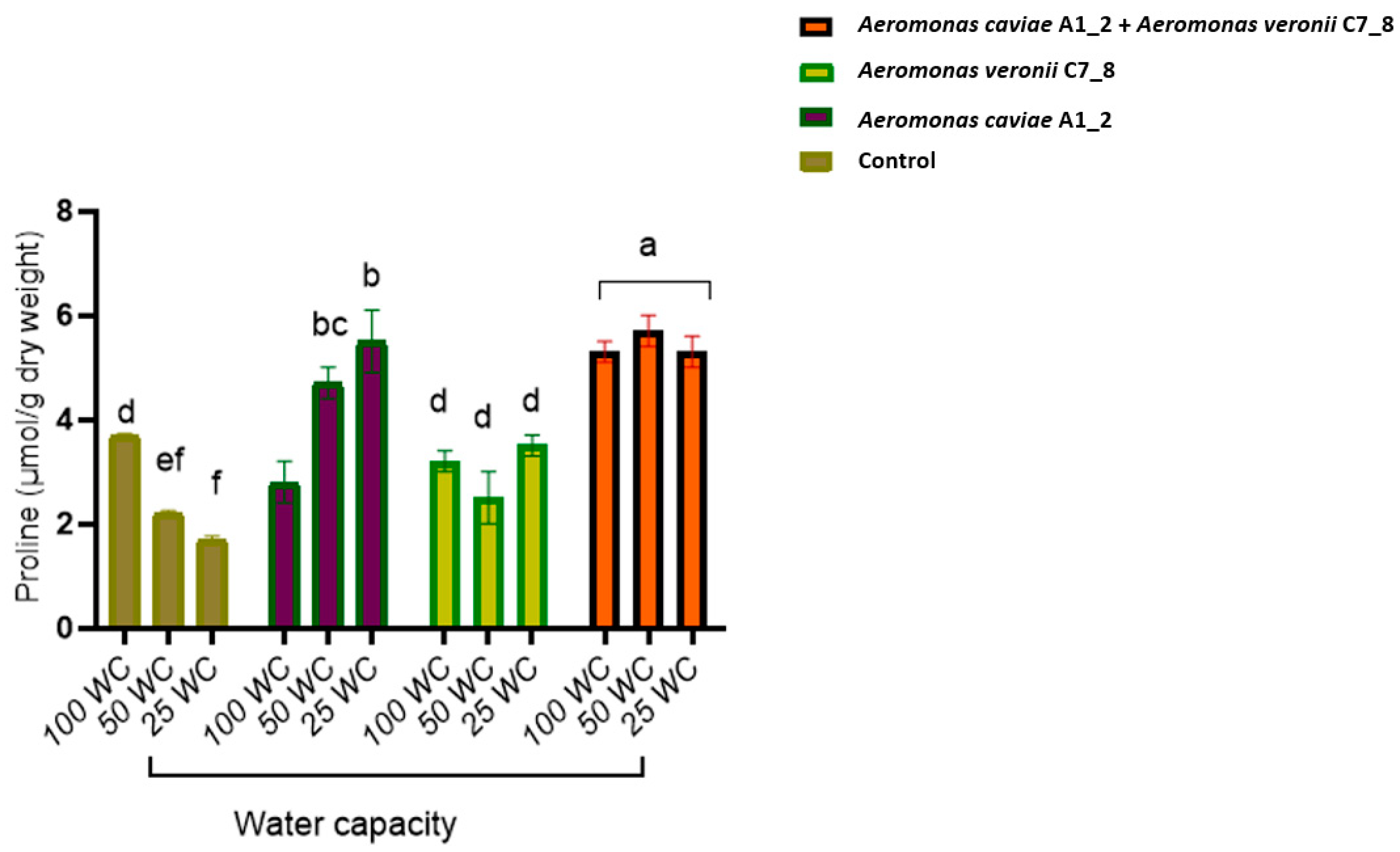
3.7. Rhizobacteria Improve the Morphophysiological Attributes of Maize Plants Exposed to Drought Stress in the Greenhouse
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adeleke, B.S.; Babalola, O.O. Meta-omics of endophytic microbes in agricultural biotechnology. Biocatal. Agric. Biotechnol. 2022, 42, 102332. [Google Scholar] [CrossRef]
- Sandhya, V.; SK, A.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Robert, G.A.; Rajasekar, M.; Manivannan, P. Triazole-induced drought stress amelioration on growth, yield, and pigments composition of Helianthus annuus L.(sunflower). Int. Multidiscip. Res. J. 2015, 5, 6–15. [Google Scholar]
- Aroca, R.; Irigoyen, J.J.; Sánchez-Díaz, M. Drought enhances maize chilling tolerance. II. Photosynthetic traits and protective mechanisms against oxidative stress. Physiol. Plant. 2003, 117, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef] [PubMed]
- Eke, P.; Kumar, A.; Sahu, K.P.; Wakam, L.N.; Sheoran, N.; Ashajyothi, M.; Patel, A.; Fekam, F.B. Endophytic bacteria of desert cactus (Euphorbia trigonas Mill) confer drought tolerance and induce growth promotion in tomato (Solanum lycopersicum L.). Microbiol. Res. 2019, 228, 126302. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hussain, M.B.; Zahir, Z.A.; Mitter, B.; Sessitsch, A. Drought stress amelioration in wheat through inoculation with Burkholderia phytofirmans strain PsJN. Plant Growth Reg. 2014, 73, 121–131. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Drogue, B.; Doré, H.; Borland, S.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Which specificity in cooperation between phytostimulating rhizobacteria and plants? Res. Microbiol. 2012, 163, 500–510. [Google Scholar] [CrossRef]
- Gepstein, S.; Glick, B.R. Strategies to ameliorate abiotic stress-induced plant senescence. Plant Mol. Biol. 2013, 82, 623–633. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Pablo, C.H.; Mavrodi, O.V.; Weller, D.M.; Thomashow, L.S.; Mavrodi, D.V. Rhizosphere plant-microbe interactions under water stress. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 115, pp. 65–113. [Google Scholar]
- Kushwaha, P.; Kashyap, P.L.; Bhardwaj, A.K.; Kuppusamy, P.; Srivastava, A.K.; Tiwari, R.K. Bacterial endophyte mediated plant tolerance to salinity: Growth responses and mechanisms of action. World J. Microbiol. Biotechnol. 2020, 36, 26. [Google Scholar] [CrossRef] [PubMed]
- Ngoma, L.; Mogatlanyane, K.; Babalola, O.O. Screening of endophytic bacteria towards the development of cottage industry: An in vitro study. J. Hum. Ecol. 2014, 47, 45–63. [Google Scholar] [CrossRef]
- Babalola, O.O.; Sanni, A.I.; Odhiambo, G.D.; Torto, B. Plant growth-promoting rhizobacteria do not pose any deleterious effect on cowpea and detectable amounts of ethylene are produced. World J. Microbiol. Biotechnol. 2007, 23, 747–752. [Google Scholar] [CrossRef]
- Gupta, A.; Gopal, M.; Thomas, G.V.; Manikandan, V.; Gajewski, J.; Thomas, G.; Seshagiri, S.; Schuster, S.C.; Rajesh, P.; Gupta, R. Whole genome sequencing and analysis of plant growth promoting bacteria isolated from the rhizosphere of plantation crops coconut, cocoa and arecanut. PLoS ONE 2014, 9, e104259. [Google Scholar] [CrossRef] [PubMed]
- Mathur, P.; Roy, S. Insights into the plant responses to drought and decoding the potential of root associated microbiome for inducing drought tolerance. Physiol. Plant. 2021, 172, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Naing, A.H.; Maung, T.T.; Kim, C.K. The ACC deaminase-producing plant growth-promoting bacteria: Influences of bacterial strains and ACC deaminase activities in plant tolerance to abiotic stress. Physiol. Plant. 2021, 173, 1992–2012. [Google Scholar] [CrossRef]
- Prischl, M.; Hackl, E.; Pastar, M.; Pfeiffer, S.; Sessitsch, A. Genetically modified Bt maize lines containing cry3Bb1, cry1A105 or cry1Ab2 do not affect the structure and functioning of root-associated endophyte communities. Appl. Soil Ecol. 2012, 54, 39–48. [Google Scholar] [CrossRef]
- Ali, S.Z.; Sandhya, V.; Venkateswar Rao, L. Isolation and characterization of drought-tolerant ACC deaminase and exopolysaccharide-producing fluorescent Pseudomonas sp. Ann. Microbiol. 2014, 64, 493–502. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.i.; Kolapo, K. Drought resistance in rice from conventional to molecular breeding: A review. Int. J. Mol. Sci. 2019, 20, 3519. [Google Scholar] [CrossRef] [PubMed]
- Qaim, M. Role of new plant breeding technologies for food security and sustainable agricultural development. Appl. Econ. Perspect. Policy 2020, 42, 129–150. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O.; Aremu, B.R. Genomic insights into plant growth promoting rhizobia capable of enhancing soybean germination under drought stress. BMC Microbiol. 2019, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, W.; Li, Q.; Cui, R.; Wang, Z.; Wang, Y.; Zhang, Y.-Z.; Ding, W.; Shen, X. Deciphering the root endosphere microbiome of the desert plant Alhagi sparsifolia for drought resistance-promoting bacteria. Appl. Environ. Microbiol. 2020, 86, 02819–02863. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Alex, T.; Oelmüller, R. Piriformospora indica: The versatile and multifunctional root endophytic fungus for enhanced yield and tolerance to biotic and abiotic stress in crop plants. J. Tropical Agric. 2014, 52, 103–122. [Google Scholar]
- Viscardi, S.; Ventorino, V.; Duran, P.; Maggio, A.; De Pascale, S.; Mora, M.; Pepe, O. Assessment of plant growth promoting activities and abiotic stress tolerance of Azotobacter chroococcum strains for a potential use in sustainable agriculture. J. Soil Sci. Plant Nutr. 2016, 16, 848–863. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Clarke, P.H.; Cowan, S. Biochemical methods for bacteriology. Microbiology 1952, 6, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Cappuccino, J.; Sherman, N. Microbiology: A Laboratory Manual; Pearson: New York, NY, USA, 1992; pp. 125–179. [Google Scholar]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- AlAli, H.A.; Khalifa, A.; Al-Malki, M. Plant growth-promoting rhizobacteria from Ocimum basilicum improve growth of Phaseolus vulgaris and Abelmoschus esculentus. S. Afr. J. Bot. 2021, 139, 200–209. [Google Scholar] [CrossRef]
- Loper, J.; Schroth, M. Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology 1986, 76, 386–389. [Google Scholar] [CrossRef]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Duan, J.; Müller, K.M.; Charles, T.C.; Vesely, S.; Glick, B.R. 1-aminocyclopropane-1-carboxylate (ACC) deaminase genes in rhizobia from southern Saskatchewan. Microb. Ecol. 2009, 57, 423–436. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef]
- Devereux, R.; Willis, S.G. Amplification of ribosomal RNA sequences. In Molecular Microbial Ecology Manual; Springer: Berlin/Heidelberg, Germany, 1995; pp. 277–287. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Prakamhang, J.; Tittabutr, P.; Boonkerd, N.; Teamtisong, K.; Uchiumi, T.; Abe, M.; Teaumroong, N. Proposed some interactions at molecular level of PGPR coinoculated with Bradyrhizobium diazoefficiens USDA110 and B. japonicum THA6 on soybean symbiosis and its potential of field application. Appl. Soil Ecol. 2015, 85, 38–49. [Google Scholar] [CrossRef]
- Yallou, C.; Aïhou, K.; Adjanohoun, A.; Toukourou, M.; Sanni, O.; Ali, D. Itinéraires techniques de production de maïs au Bénin. Fiche Technique. Dépôt Légal 2010, 4992, 18. [Google Scholar]
- Ortiz, N.; Armada, E.; Duque, E.; Roldán, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2015, 174, 87–96. [Google Scholar] [CrossRef]
- Zarik, L.; Meddich, A.; Hijri, M.; Hafidi, M.; Ouhammou, A.; Ouahmane, L.; Duponnois, R.; Boumezzough, A. Use of Arbuscular mycorrhizal fungi to improve the drought tolerance of Cupressus atlantica G. Comptes Rendus Biol. 2016, 339, 185–196. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.A.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Ding, L.-J.; Cui, H.-L.; Nie, S.-A.; Long, X.-E.; Duan, G.-L.; Zhu, Y.-G. Microbiomes inhabiting rice roots and rhizosphere. FEMS Microbiol. Ecol. 2019, 95, fiz040. [Google Scholar] [CrossRef]
- Mommer, L.; Kirkegaard, J.; van Ruijven, J. Root–root interactions: Towards a rhizosphere framework. Trends Plant Sci. 2016, 21, 209–217. [Google Scholar] [CrossRef]
- Perez-Perez, R.; Oudot, M.; Hernandez, I.; Napoles, M.C.; Perez-Martinez, S.; Sosa-Del Castillo, D.; Postal, G.; San José de las Lajas, M. Isolation and characterization of Stenotrophomonas asociated to maize (Zea mays L.) rhizosphere. Cultiv. Trop. 2020, 41, e03. [Google Scholar]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Ann. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Davey, M.E.; O’toole, G.A. Microbial biofilms: From ecology to molecular genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef]
- Nawaz, M.S.; Arshad, A.; Rajput, L.; Fatima, K.; Ullah, S.; Ahmad, M.; Imran, A. Growth-stimulatory effect of quorum sensing signal molecule N-acyl-homoserine lactone-producing multi-trait Aeromonas spp. on wheat genotypes under salt stress. Front. Microbiol. 2020, 11, 2164. [Google Scholar] [CrossRef]
- Noori, F.; Etesami, H.; Noori, S.; Forouzan, E.; Jouzani, G.S.; Malboobi, M.A. Whole genome sequence of Pantoea agglomerans ANP8, a salinity and drought stress–resistant bacterium isolated from alfalfa (Medicago sativa L.) root nodules. Biotechnol. Rep. 2021, 29, e00600. [Google Scholar] [CrossRef]
- Filgueiras, L.; Silva, R.; Almeida, I.; Vidal, M.; Baldani, J.I.; Meneses, C.H.S.G. Gluconacetobacter diazotrophicus mitigates drought stress in Oryza sativa L. Plant Soil 2020, 451, 57–73. [Google Scholar] [CrossRef]
- Annapurna, K.; Govindasamy, V.; Sharma, M.; Ghosh, A.; Chikara, S.K. Whole genome shotgun sequence of Bacillus paralicheniformis strain KMS 80, a rhizobacterial endophyte isolated from rice (Oryza sativa L.). 3 Biotech 2018, 8, 223. [Google Scholar] [CrossRef]
- Rani, R.; Kumar, V.; Usmani, Z.; Gupta, P.; Chandra, A. Influence of plant growth promoting rhizobacterial strains Paenibacillus sp. IITISM08, Bacillus sp. PRB77 and Bacillus sp. PRB101 using Helianthus annuus on degradation of endosulfan from contaminated soil. Chemosphere 2019, 225, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Hol, W.G.; Bezemer, T.M.; Biere, A. Getting the ecology into interactions between plants and the plant growth-promoting bacterium Pseudomonas fluorescens. Front. Plant Sci. 2013, 4, 81. [Google Scholar] [CrossRef]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Saleem, M.; Hussain, S. Perspectives of bacterial ACC deaminase in phytoremediation. Trends Biotechnol. 2007, 25, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Cruz, J.A.; Tubana, B.S.; Fultz, L.M.; Dalen, M.S.; Ham, J.H. Identification and profiling of silicate-solubilizing bacteria for plant growth-promoting traits and rhizosphere competence. Rhizosphere 2022, 23, 100566. [Google Scholar] [CrossRef]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G. Azospirillum–plant relationships: Environmental and physiological advances (1990–1996). Canad. J. Microbiol. 1997, 43, 103–121. [Google Scholar] [CrossRef]
- Zaveri, S.H.; Dasgupta, S. The Interaction between Plant and Endophytes for Conferring Tolerance to Biotic and Abiotic Stress in Presence of ACC (1-Aminocyclopropane-1-Carboxylate) Deaminase: A Stress Weapon. Acta Sci. Biotechnol. 2023, 4, 9–14. [Google Scholar]
- Liu, Y.; Gao, J.; Bai, Z.; Wu, S.; Li, X.; Wang, N.; Du, X.; Fan, H.; Zhuang, G.; Bohu, T. Unraveling mechanisms and impact of microbial recruitment on oilseed rape (Brassica napus L.) and the rhizosphere mediated by plant growth-promoting rhizobacteria. Microorganisms 2021, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Raj, K.; Kumar, P.; Wati, L. Antagonistic and growth-promoting potential of multifarious bacterial endophytes against Fusarium wilt of chickpea. Egypt. J. Biol. Pest Contr. 2022, 32, 17. [Google Scholar] [CrossRef]
- Chaudhary, P.; Xu, M.; Ahamad, L.; Chaudhary, A.; Kumar, G.; Adeleke, B.S.; Verma, K.K.; Hu, D.-M.; Širić, I.; Kumar, P. Application of synthetic consortia for improvement of soil fertility, pollution remediation, and agricultural productivity: A review. Agronomy 2023, 13, 643. [Google Scholar] [CrossRef]
- Abedinzadeh, M.; Etesami, H.; Alikhani, H.A. Characterization of rhizosphere and endophytic bacteria from roots of maize (Zea mays L.) plant irrigated with wastewater with biotechnological potential in agriculture. Biotechnol. Rep. 2019, 21, e00305. [Google Scholar] [CrossRef] [PubMed]
- Fasusi, O.A.; Amoo, A.E.; Babalola, O.O. Characterization of plant growth-promoting rhizobacterial isolates associated with food plants in South Africa. Antonie Van Leeuwenhoek 2021, 114, 1683–1708. [Google Scholar] [CrossRef] [PubMed]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.-I.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [PubMed]
- Nehra, M.; Dilbaghi, N.; Marrazza, G.; Kaushik, A.; Sonne, C.; Kim, K.-H.; Kumar, S. Emerging nanobiotechnology in agriculture for the management of pesticide residues. J. Hazard. Mater. 2021, 401, 123369. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Dikilitas, M.; Abdel-Azeem, A.M.; Ahluwalia, A.S.; Saxena, A.K. Biodiversity, and biotechnological contribution of beneficial soil microbiomes for nutrient cycling, plant growth improvement and nutrient uptake. Biocatal. Agric. Biotechnol. 2021, 33, 102009. [Google Scholar] [CrossRef]
- Asari, S.; Tarkowská, D.; Rolčík, J.; Novák, O.; Palmero, D.V.; Bejai, S.; Meijer, J. Analysis of plant growth-promoting properties of Bacillus amyloliquefaciens UCMB5113 using Arabidopsis thaliana as host plant. Planta 2017, 245, 15–30. [Google Scholar] [CrossRef]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Majeed, A.; Abbasi, M.K.; Hameed, S.; Imran, A.; Rahim, N. Isolation and characterization of plant growth-promoting rhizobacteria from wheat rhizosphere and their effect on plant growth promotion. Front. Microbiol. 2015, 6, 198. [Google Scholar] [CrossRef]
- Kunwar, V.S.; Lamichhane, J.; Gauchan, D.P. Strategies to improve phosphorus availability in a sustainable agricultural system. Int. J. Innov. Sci. Res. Technol. 2018, 3, 323–331. [Google Scholar]
- Albareda, M.; Rodríguez-Navarro, D.N.; Camacho, M.; Temprano, F.J. Alternatives to peat as a carrier for rhizobia inoculants: Solid and liquid formulations. Soil Biol. Biochem. 2008, 40, 2771–2779. [Google Scholar] [CrossRef]
- Siddiq, S.; Saleem, U.; Ahmad, K.; Anayat, A.; Affan, Q.; Anwar, M.; Nazir, H.; Asghar, N. Comparison of conventional and non-conventional carriers for bacterial survival and plant growth. Int. J. Agric. Innovations Res. 2018, 6, 126–129. [Google Scholar]
- Scott, S.; Housh, A.; Powell, G.; Anstaett, A.; Gerheart, A.; Benoit, M.; Wilder, S.; Schueller, M.; Ferrieri, R. Crop yield, ferritin and Fe (II) boosted by Azospirillum brasilense (HM053) in corn. Agronomy 2020, 10, 394. [Google Scholar] [CrossRef]
- Vacheron, J.; Renoud, S.; Muller, D.; Babalola, O.O.; Prigent-Combaret, C. Alleviation of abiotic and biotic stresses in plants by Azospirillum. In Handbook for Azospirillum: Technical Issues Protocols; Springer: Berlin/Heidelberg, Germany, 2015; pp. 333–365. [Google Scholar]
- Diarrassouba, N.; Dago, D.N.; Soro, S.; Fofana, I.J.; Silué, S.; Coulibaly, A. Multi-variant statistical analysis evaluating the impact of rhizobacteria (Pseudomonas fluorescens) on growth and yield parameters of two varieties of maize (Zea mays. L). Int. J. Contemp. Appl. Sci. 2015, 2, 206–224. [Google Scholar]
- Noumavo, P.A.; Agbodjato, N.A.; Gachomo, E.W.; Salami, H.A.; Baba-Moussa, F.; Adjanohoun, A.; Kotchoni, S.O.; Baba-Moussa, L. Metabolic and biofungicidal properties of maize rhizobacteria for growth promotion and plant disease resistance. Afr. J. Biotechnol. 2015, 14, 811–819. [Google Scholar]
- Amogou, O.; Dagbénonbakin, G.; Agbodjato, N.A.; Noumavo, P.A.; Salako, K.V.; Adoko, M.Y.; Kakaï, R.G.; Adjanohoun, A.; Baba-Moussa, L. Applying rhizobacteria on maize cultivation in Northern Benin: Effect on growth and yield. Agric. Sci. 2019, 10, 763–782. [Google Scholar] [CrossRef]
- Mejri, D.; Gamalero, E.; Souissi, T. Formulation development of the deleterious rhizobacterium Pseudomonas trivialis X33d for biocontrol of brome (Bromus diandrus) in durum wheat. J. Appl. Microbiol. 2013, 114, 219–228. [Google Scholar] [CrossRef]
- Agbodjato, N.A.; Noumavo, P.A.; Adjanohoun, A.; Agbessi, L.; Baba-Moussa, L. Synergistic effects of plant growth promoting rhizobacteria and chitosan on in vitro seeds germination, greenhouse growth, and nutrient uptake of maize (Zea mays L.). Biotechnol. Res. Int. 2016, 2016, 7830182. [Google Scholar] [CrossRef] [PubMed]
- Janardan, Y.; Verma, J.P.; Tiwari, K.N. Effect of plant growth promoting rhizobacteria on seed germination and plant growth chickpea (Cicer arietinum L.) under in vitro conditions. Biol. Forum 2010, 2, 15–18. [Google Scholar]
- Kumar, A.; Maurya, B.; Raghuwanshi, R. Isolation and characterization of PGPR and their effect on growth, yield and nutrient content in wheat (Triticum aestivum L.). Biocatal. Agric. Biotechnol. 2014, 3, 121–128. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Fathi Abd_Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Control | 5% | 10% | 15% | 20% | 25% | 30% |
|---|---|---|---|---|---|---|---|
| A5-1 | 0.35 ± 0.17fg | 0.18 ± 0.01c | 0.16 ± 0.01d | 0.16 ± 0.00f | 0.16 ± 0.00d | 0.13 ± 0.00f | 0.12 ± 0.00c |
| A1-2 | 1.01 ± 0.07b | 0.67 ± 0.23a | 0.80 ± 0.15a | 0.89 ± 0.03a | 0.87 ± 0.03a | 0.81 ± 0.01a | 0.71 ± 0.01a |
| B8-3 | 0.73 ± 0.03bc | 0.38 ± 0.11ab | 0.41 ± 0.02bc | 0.39 ± 0.00c | 0.37 ± 0.02b | 0.30 ± 0.00c | 0.26 ± 0.03bc |
| B12-4 | 0.65 ± 0.08cde | 0.40 ± 0.11abc | 0.41 ± 0.01bc | 0.41 ± 0.01c | 0.41 ± 0.00b | 0.35 ± 0.01b | 0.30 ± 0.00bc |
| B9-5 | 0.21 ± 0.03g | 0.24 ± 0.05c | 0.21 ± 0.02cd | 0.20 ± 0.00ef | 0.20 ± 0.00d | 0.18 ± 0.01e | 0.15 ± 0.01c |
| B15-6 | 0.43 ± 0.08egf | 0.27 ± 0.05c | 0.31 ± 0.02cbd | 0.31 ± 0.00d | 0.28 ± 0.00c | 0.22 ± 0.00d | 0.20 ± 0.00c |
| C6-7 | 0.84 ± 0.06bc | 0.40 ± 0.11abc | 0.45 ± 0.04b | 0.41 ± 0.00c | 0.40 ± 0.0.00b | 0.32 ± 0.01bc | 0.29 ± 0.00bc |
| C7_8 | 1.31 ± 0.13a | 0.53 ± 0.20ab | 0.73 ± 0.15a | 0.84 ± 0.03b | 0.84 ± 0.03a | 0.81 ± 0.04a | 0.77 ± 0.03a |
| C1-9 | 0.35 ± 0.01fg | 0.33 ± 0.08bc | 0.31 ± 0.02bcd | 0.21 ± 0.00e | 0.20 ± 0.00d | 0.19 ± 0.00de | 0.14 ± 0.01c |
| A9-10 | 0.23 ± 0.03g | 0.24 ± 0.04c | 0.17 ± 0.00d | 0.17 ± 0.01ef | 0.16 ± 0.00d | 0.17 ± 0.01e | 0.16 ± 0.01c |
| A10-11 | 0.55 ± 0.05edf | 0.25 ± 0.04c | 0.20 ± 0.01cd | 0.18 ± 0.01ef | 0.17 ± 0.01d | 0.17 ± 0.01e | 0.15 ± 0.01c |
| LSD | 0.22 | 0.23 | 0.20 | 0.04 | 0.04 | 0.04 | 0.24 |
| Rhizobacterial Isolates | Plant-Growth-Promoting Traits | ||||||
|---|---|---|---|---|---|---|---|
| Nitrogen Fixation | HCN Production | Siderophore Production | Ammonia Production | IAA Production | Phosphate Solubilization | ACC Deaminase | |
| B. licheniformis A5-1 | + | – | ++ | +++ | + | + | + |
| A. caviae A1-2 | + | – | ++ | ++++ | ++ | + | + |
| B. cereus B8-3 | + | – | + | +++ | + | + | + |
| P. flexa B12-4 | + | – | + | ++ | + | + | + |
| B. licheniformis B9-5 | + | – | + | ++ | + | + | + |
| B. simplex B15-6 | + | – | + | ++ | + | + | + |
| P. flexa C6-7 | + | – | ++ | +++ | ++ | + | + |
| A. veronii C7_8 | + | – | ++ | ++++ | ++ | + | + |
| P. aryabhattai C1-9 | + | – | + | ++ | + | + | + |
| B. halotolerans A9-10 | + | – | + | +++ | + | + | + |
| P.endophytica A10-11 | + | – | + | +++ | + | + | + |
| Rhizobacteria | Media | 0% Salt | 1% Salt | 3% Salt | 5% Salt |
|---|---|---|---|---|---|
| Aeromonas caviae (A1-2) | NB | 1.64 ± 0.18 a | 1.42 ± 0.56 a | 1.37 ± 0.60 a | 0.55 ± 0.02 b |
| LB | 1.62 ± 0.18 a | 1.34 ± 0.56 ab | 0.96 ± 0.56 b | 0.50 ± 0.02 c | |
| TSB | 1.47 ± 0.12 a | 1.28 ± 0.56 ab | 0.90 ± 0.56 b | 0.45 ± 0.02 c | |
| R2AB | 1.37 ± 0.12 a | 1.22 ± 0.51 a | 1.18 ± 0.56 a | 0.40 ± 0.02 b | |
| Aeromonas veronii (C7_8) | NB | 1.61 ± 0.18 a | 1.42 ± 0.54 a | 0.70 ± 0.543 bc | 0.45 ± 0.12 c |
| LB | 1.60 ± 0.13 a | 1.35 ± 0.61 ab | 1.00 ± 0.61 abc | 0.41 ± 0.12 c | |
| TSB | 1.52 ± 0.25 a | 1.28 ± 0.55 ab | 0.96 ± 0.56 abc | 0.37 ± 0.12 c | |
| R2AB | 1.4224 ± 0.3154 a | 1.22 ± 0.60 ab | 0.93 ± 0.57 abc | 0.38 ± 0.18 c |
| Rhizobacteria | Media | pH4 | pH7 | pH10 |
|---|---|---|---|---|
| Aeromonas caviae (A1-2) | NB | 0.97 ± 0.05 ab | 1.11 ± 0.02 a | 0.56 ± 0.32 c |
| LB | 0.88 ± 0.05 b | 1.03 ± 0.27 a | 0.50 ± 0.29 c | |
| TSB | 0.79 ± 0.04 b | 0.96 ± 0.30 a | 0.45 ± 0.26 c | |
| R2AB | 0.71 ± 0.04 b | 1.03 ± 0.56 a | 0.27 ± 0.01 c | |
| Aeromonas veronii (C7_8) | NB | 0.91 ± 0.09 ab | 1.53 ± 0.60 a | 0.40 ± 0.01 c |
| LB | 0.82 ± 0.07 ab | 1.11 ± 0.56 a | 0.36 ± 0.01 c | |
| TSB | 0.74 ± 0.07 ab | 1.36 ± 0.59 a | 0.32 ± 0.01 c | |
| R2AB | 0.66 ± 0.06 ab | 1.29 ± 0.60 a | 0.30 ± 0.02 c |
| Rhizobacteria | Media | 25 °C | 30 °C | 35 °C | 40 °C |
|---|---|---|---|---|---|
| Aeromonas caviae (A1-2) | NB | 0.93 ± 0.03 a | 1.20 ± 0.01 a | 1.72 ± 0.01 a | 0.83 ± 0.03 a |
| LB | 0.87 ± 0.07 a | 1.08 ± 0.01 a | 1.59 ± 0.17 a | 0.75 ± 0.032 b | |
| TSB | 0.99 ± 0.42 a | 0.97 ± 0.01 a | 1.21 ± 0.48 a | 0.67 ± 0.02 b | |
| R2AB | 0.88 ± 0.01 b | 1.01 ± 0.59 a | 1.32 ± 0.58 a | 0.61 ± 0.02 b | |
| Aeromonas veronii (C7_8) | NB | 0.97 ± 0.01 a | 1.09 ± 0.01 a | 1.64 ± 0.05 a | 0.82 ± 0.04 a |
| LB | 0.87 ± 0.01 b | 0.98 ± 0.01 b | 1.55 ± 0.01 a | 0.74 ± 0.04 c | |
| TSB | 0.88 ± 0.01 b | 1.02 ± 0.40 a | 1.40 ± 0.01 a | 0.66 ± 0.03 c | |
| R2AB | 1.04 ± 0.57 a | 1.26 ± 0.01 a | 0.79 ± 0.01 b | 0.60 ± 0.03 b |
| Rhizobacteria | Media | 0 (Control) | Pb2+ | Cr2O72- | Cd2+ |
|---|---|---|---|---|---|
| Aeromonas caviae (A1-2) | NB | 1.28 ± 0.54 a | 0.76 ± 0.08 ab | 0.63 ± 0.10 b | 0.50 ± 0.09 b |
| LB | 1.56 ± 0.22 a | 0.68 ± 0.08 b | 0.56 ± 0.09 bc | 0.44 ± 0.08 c | |
| TSB | 1.51 ± 0.13 a | 0.61 ± 0.07 b | 0.51 ± 0.08 c | 0.40 ± 0.07 c | |
| R2AB | 1.08 ± 0.36 a | 0.55 ± 0.06 b | 0.46 ± 0.07 b | 0.36 ± 0.06 c | |
| Aeromonas veronii (C7_8) | NB | 1.26 ± 0.53 a | 0.74 ± 0.08 b | 0.61 ± 0.10 b | 0.49 ± 0.09 c |
| LB | 1.26 ± 0.35 a | 0.67 ± 0.01 b | 0.55 ± 0.09 b | 0.44 ± 0.08 c | |
| TSB | 1.15 ± 0.34 a | 0.60 ± 0.07 b | 0.50 ± 0.08 b | 0.40 ± 0.07 c | |
| R2AB | 1.0787 ± 0.3217 a | 0.54 ± 0.06 b | 0.45 ± 0.07 c | 0.36 ± 0.06 c |
| Water Regime | Growth Parameters | Control | A1-2 | C7_8 | A1-2+C7_8 |
|---|---|---|---|---|---|
| 100 WC | Plant height (cm) | 7.27 ± 0.95 c | 8.43 ± 0.43 ab | 7.95 ± 0.50 b | 9.33 ± 0.34 a |
| Number of leaves | 5.67 ± 0.29 d | 6.67 ± 0.14 b | 6.50 ± 0.29 c | 7.08 ± 0.29 a | |
| Stem girth (cm) | 3.08 ± 0.21 c | 4.19 ± 0.31 b | 3.73 ± 0.18 bc | 4.58 ± 0.27 a | |
| Chlorophyll content | 9.12 ± 0.71 bcd | 11.78 ± 2.22 b | 10.13 ± 0.92 bc | 14.13 ± 2.74 a | |
| Leaf area (cm2) | 52.60 ± 5.35 c | 79.67 ± 4.30 ab | 63.81 ± 2.89 abc | 85.13 ± 14.13 a | |
| 50 WC | Plant height (cm) | 6.08 ± 0.45 d | 8.11 ± 0.63 ab | 6.83 ± 0.27 c | 8.71 ± 0.23 a |
| Number of leaves | 5.92 ± 0.14 a | 6.58 ± 0.43 a | 6.08 ± 0.14 a | 6.50 ± 0.29 a | |
| Stem girth (cm) | 3.06 ± 0.23 b | 3.75 ± 0.41 ab | 3.45 ± 0.20 ab | 4.53 ± 0.32 a | |
| Chlorophyll content | 8.44 ± 0.52 c | 10.69 ± 1.49 b | 9.57 ± 0.89 b | 14.64 ± 3.56 a | |
| Leaf area (cm2) | 46.88 ± 1.76 c | 66.57 ± 8.72 ab | 44.44 ± 0.52 abc | 72.37 ± 0.73 a | |
| 25 WC | Plant height (cm) | 6.10 ± 0.30 d | 7.68 ± 0.75 ab | 6.62 ± 0.29 c | 8.38 ± 0.53 a |
| Number of leaves | 6.00 ± 0.29 a | 6.42 ± 0.39 a | 6.08 ± 0.14 a | 6.50 ± 0.00 a | |
| Stem girth (cm) | 3.17 ± 0.11 b | 3.73 ± 0.36 ab | 3.43 ± 0.13 b | 4.18 ± 0.34 a | |
| Chlorophyll content | 9.09 ± 0.92 c | 12.93 ± 1.98 ab | 10.92 ± 1.57 abc | 13.64 ± 1.50 a | |
| Leaf area (cm2) | 42.67 ± 11.19 c | 55.94 ± 2.03 a | 47.78 ± 1.29 bc | 73.80 ± 0.82 a |
| Water Capacity | Parameters (g) | Control | A1-2 | C7_8 | A1-2+C7_8 |
|---|---|---|---|---|---|
| 100 | Fresh Total Plant Mass | 10.32 ± 0.22 c | 18.70 ± 4.36 ab | 11.85 ± 1.01 b | 19.41 ± 4.01 a |
| Fresh Root Mass | 3.59 ± 1.69 d | 6.28 ± 1.75 ab | 3.52 ± 0.45 c | 8.42 ± 2.31 a | |
| Fresh Shoot Mass | 6.49 ± 1.58 d | 10.06 ± 2.14 b | 8.28 ± 0.98 c | 13.19 ± 2.53 a | |
| Dry Total Plant Mass | 1.45 ± 0.03 d | 3.22 ± 0.68 ab | 1.81 ± 0.23 cd | 3.45 ± 0.62 a | |
| Dry Root Mass | 0.53 ± 0.17 cd | 1.24 ± 0.35 ab | 0.58 ± 0.06 cd | 1.35 ± 0.34 a | |
| Dry Shoot Mass | 1.03 ± 0.09 bcd | 1.87 ± 0.34 ab | 1.21 ± 0.19 abc | 2.12 ± 0.28 a | |
| 50 | Fresh Total Plant Mass | 5.75 ± 2.12 cd | 13.45 ± 3.58 b | 7.53 ± 1.31 bc | 16.68 ± 0.90 a |
| Fresh Root Mass | 1.70 ± 0.85 c | 4.59 ± 0.98 a | 3.93 ± 0.58 b | 4.35 ± 0.48 a | |
| Fresh Shoot Mass | 4.03 ± 1.27 cd | 8.72 ± 2.52 ab | 3.58 ± 1.86 d | 12.31 ± 1.36 a | |
| Dry Total Plant Mass | 1.07 ± 0.06 bcd | 2.15 ± 0.71 ab | 1.33 ± 0.14 bc | 2.80 ± 0.24 a | |
| Dry Root Mass | 0.37 ± 0.05 d | 0.91 ± 0.36 ab | 0.47 ± 0.08 bc | 0.96 ± 0.03 a | |
| Dry Shoot Mass | 0.64 ± 0.03 d | 1.43 ± 0.49 ab | 0.78 ± 0.08 abc | 1.77 ± 0.22 a | |
| 25 | Fresh Total Plant Mass | 5.12 ± 0.44 cd | 10.52 ± 3.86 ab | 6.69 ± 1.20 bc | 10.29 ± 2.85 a |
| Fresh Root Mass | 1.49 ± 0.21 c | 2.03 ± 0.33 ab | 1.75 ± 1.08 abc | 3.91 ± 0.70 a | |
| Fresh Shoot Mass | 2.94 ± 0.07 bcd | 6.61 ± 3.04 b | 5.14 ± 1.36 bc | 8.54 ± 1.78 a | |
| Dry Total Plant Mass | 0.91 ± 0.07 d | 1.72 ± 0.49 ab | 1.21 ± 0.13 c | 1.86 ± 0.56 a | |
| Dry Root Mass | 0.39 ± 0.05 bcd | 0.50 ± 0.13 b | 0.43 ± 0.03 ac | 0.74 ± 0.21 a | |
| Dry Shoot Mass | 0.51 ± 0.01 cd | 0.98 ± 0.27 ab | 0.78 ± 0.13 c | 1.27 ± 0.34 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agunbiade, V.F.; Fadiji, A.E.; Agbodjato, N.A.; Babalola, O.O. Isolation and Characterization of Plant-Growth-Promoting, Drought-Tolerant Rhizobacteria for Improved Maize Productivity. Plants 2024, 13, 1298. https://doi.org/10.3390/plants13101298
Agunbiade VF, Fadiji AE, Agbodjato NA, Babalola OO. Isolation and Characterization of Plant-Growth-Promoting, Drought-Tolerant Rhizobacteria for Improved Maize Productivity. Plants. 2024; 13(10):1298. https://doi.org/10.3390/plants13101298
Chicago/Turabian StyleAgunbiade, Victor Funso, Ayomide Emmanuel Fadiji, Nadège Adoukè Agbodjato, and Olubukola Oluranti Babalola. 2024. "Isolation and Characterization of Plant-Growth-Promoting, Drought-Tolerant Rhizobacteria for Improved Maize Productivity" Plants 13, no. 10: 1298. https://doi.org/10.3390/plants13101298