


Foliar Application of Oil Palm Wood Vinegar Enhances Pandanus amaryllifolius Tolerance under Drought Stress

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
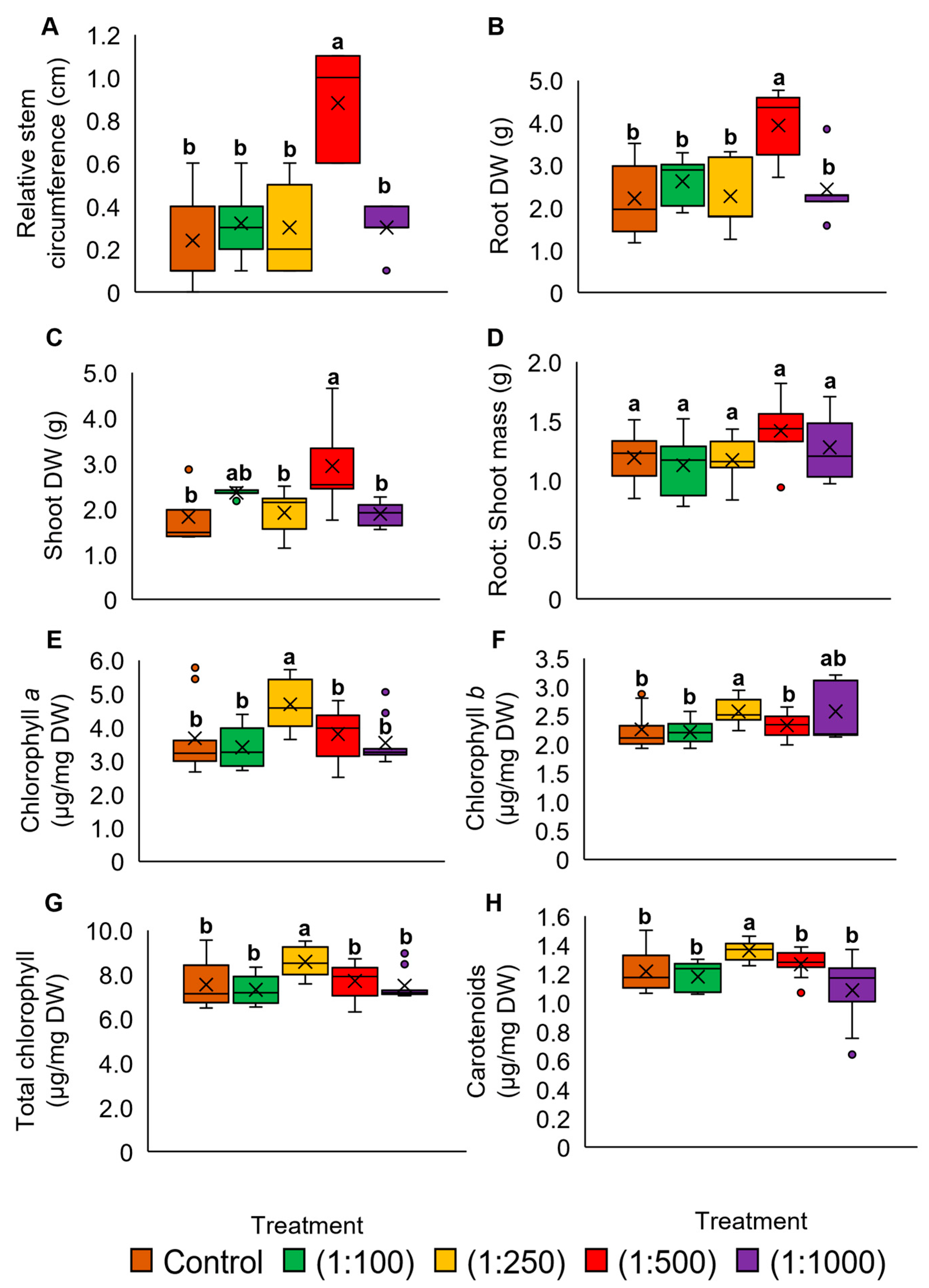
2.1. Different Oil Palm Wood Vinegar (OPWV) Dilution Factors and Application Frequencies on Pandanus amaryllifolius Growth
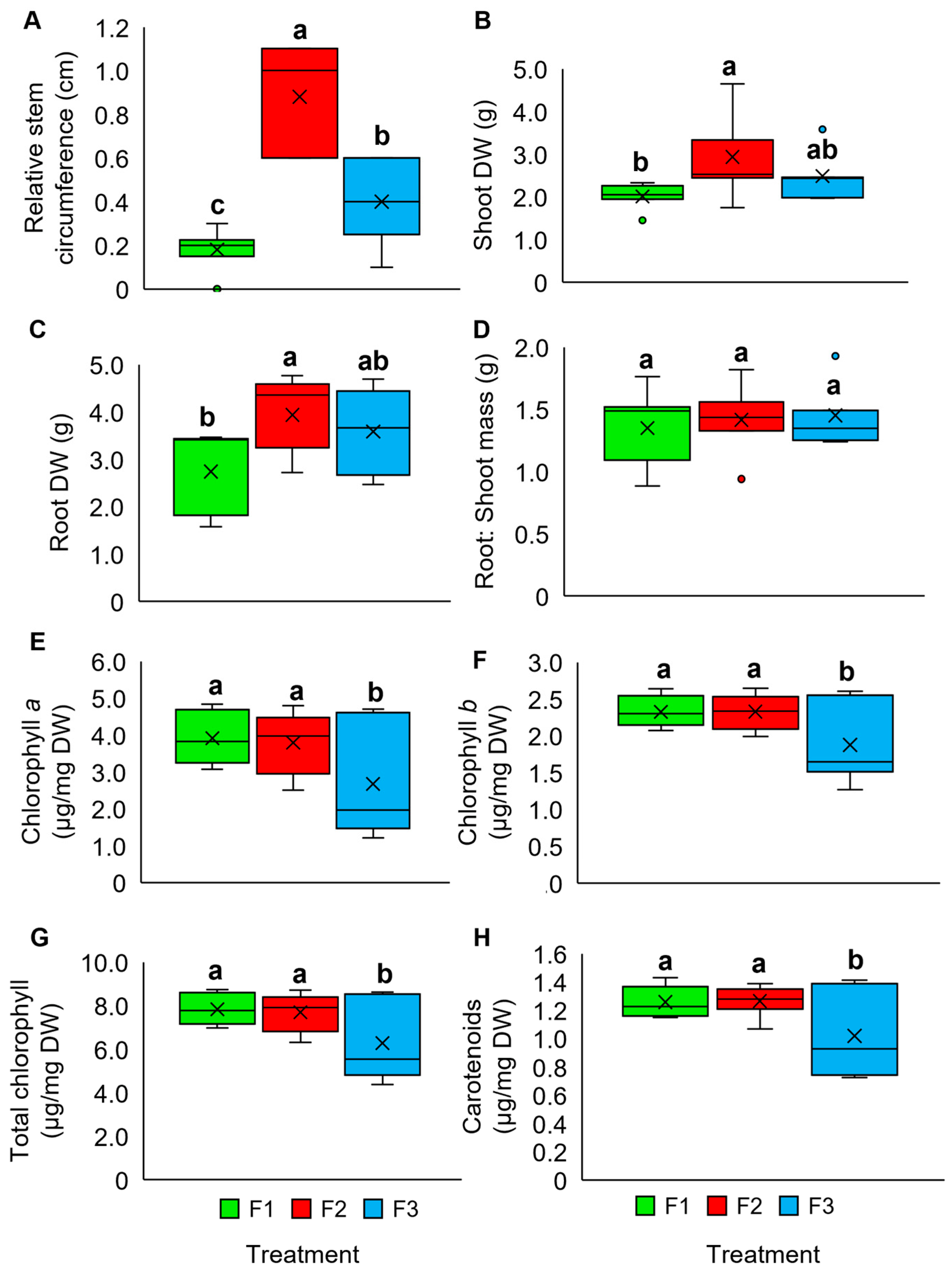
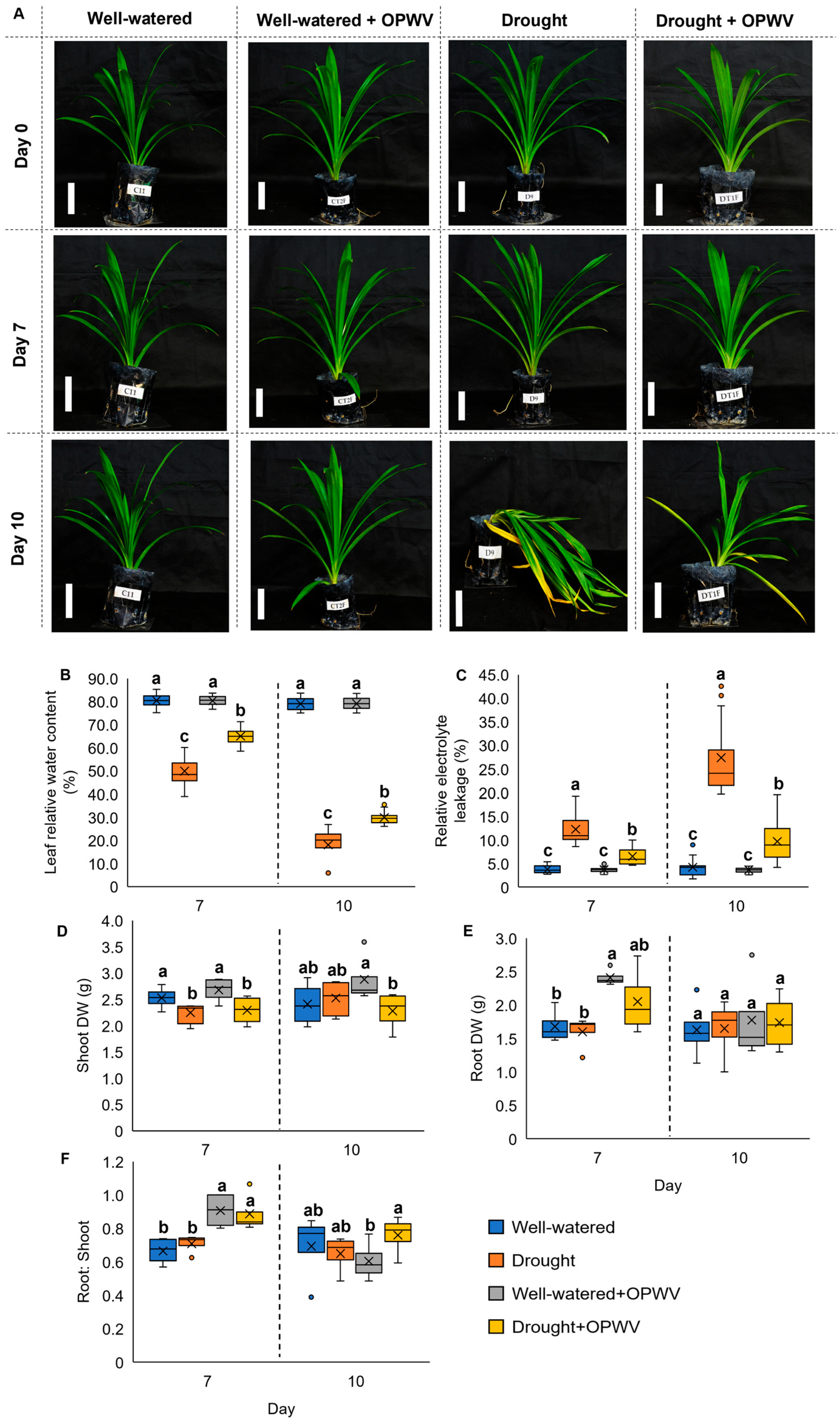
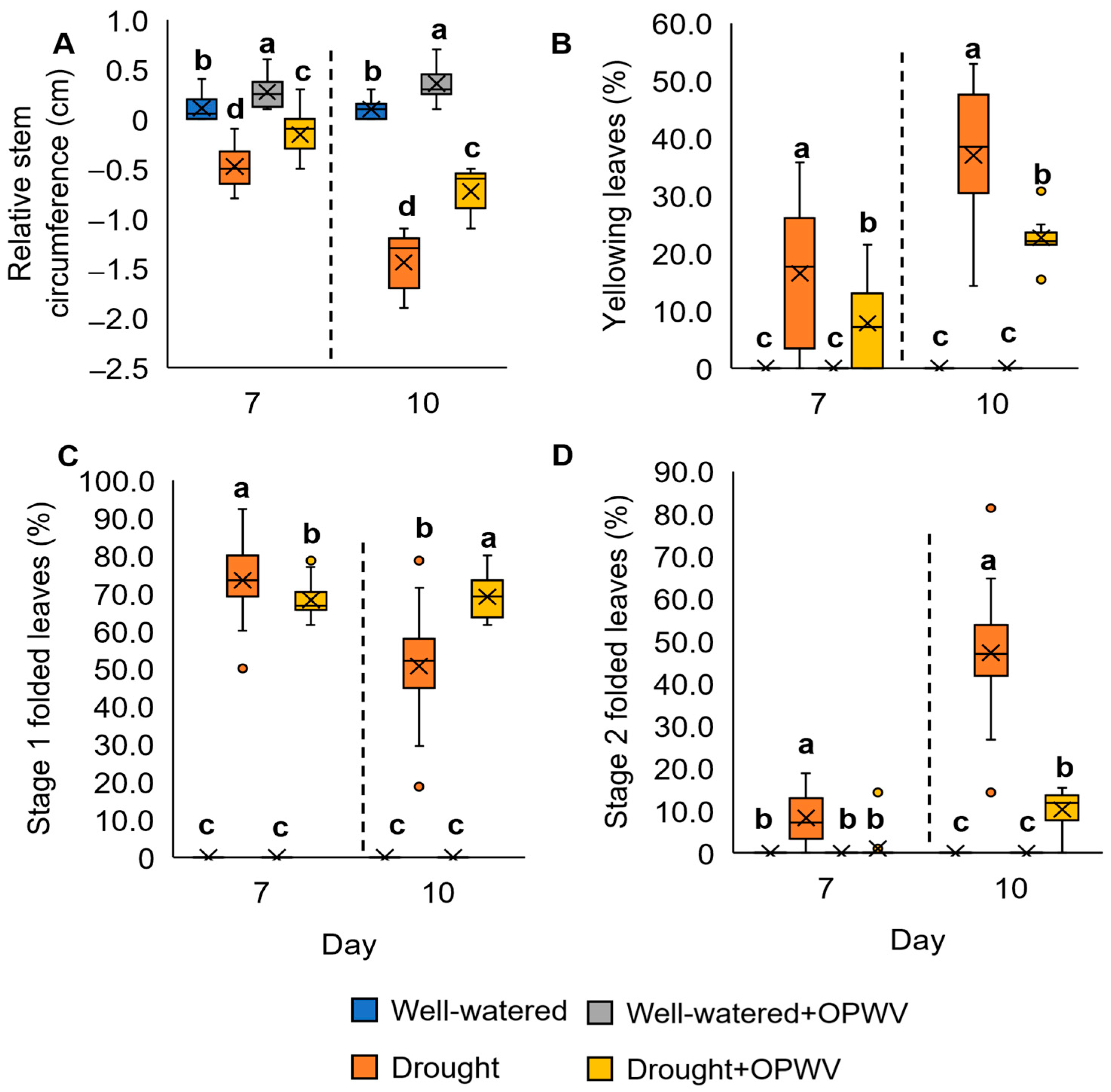
2.2. Oil Palm Wood Vinegar (OPWV) Improved Drought-Stressed Pandanus amaryllifolius
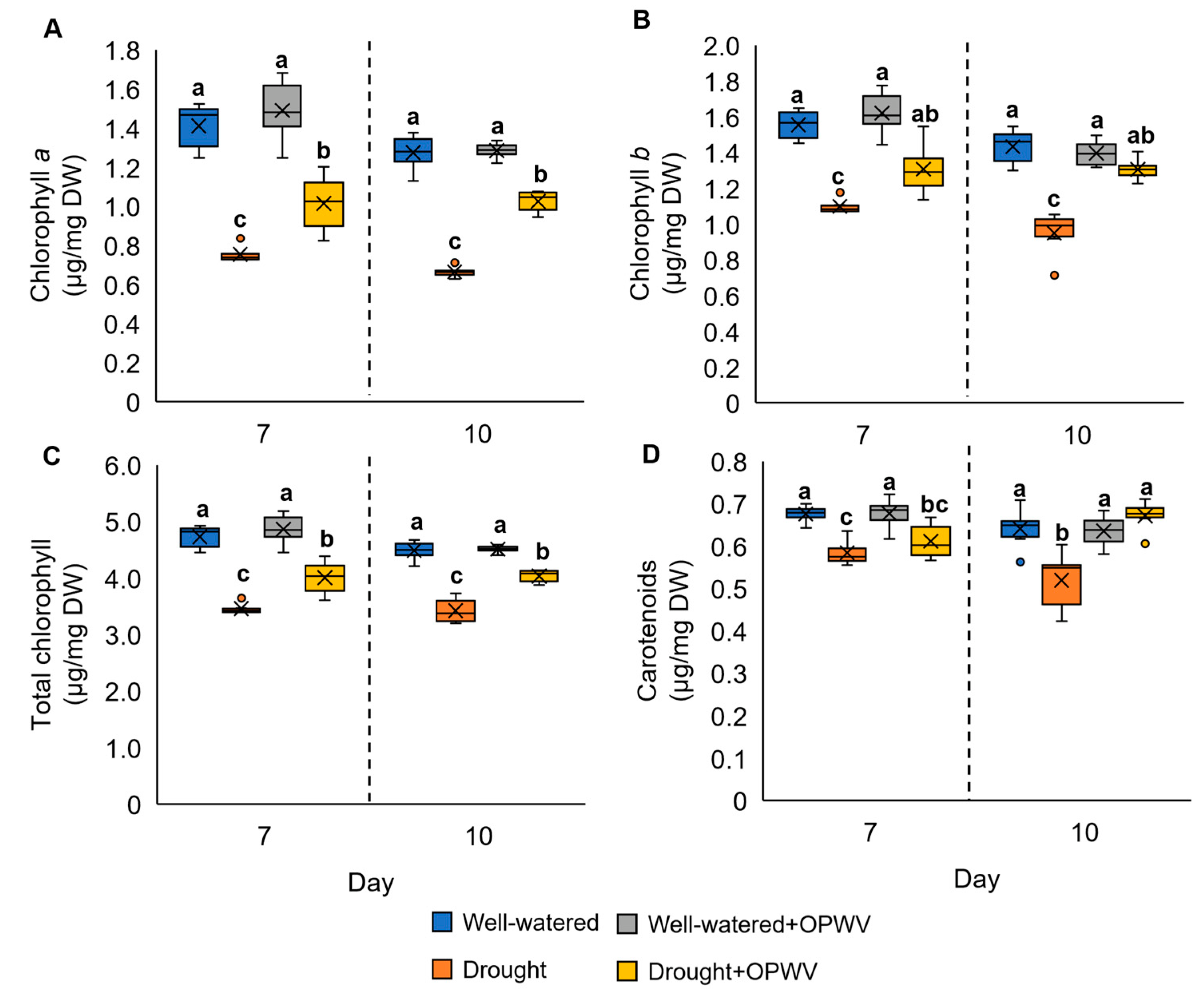
2.3. Leaf Pigment Constituents in Pandanus amaryllifolius in Response to OPWV
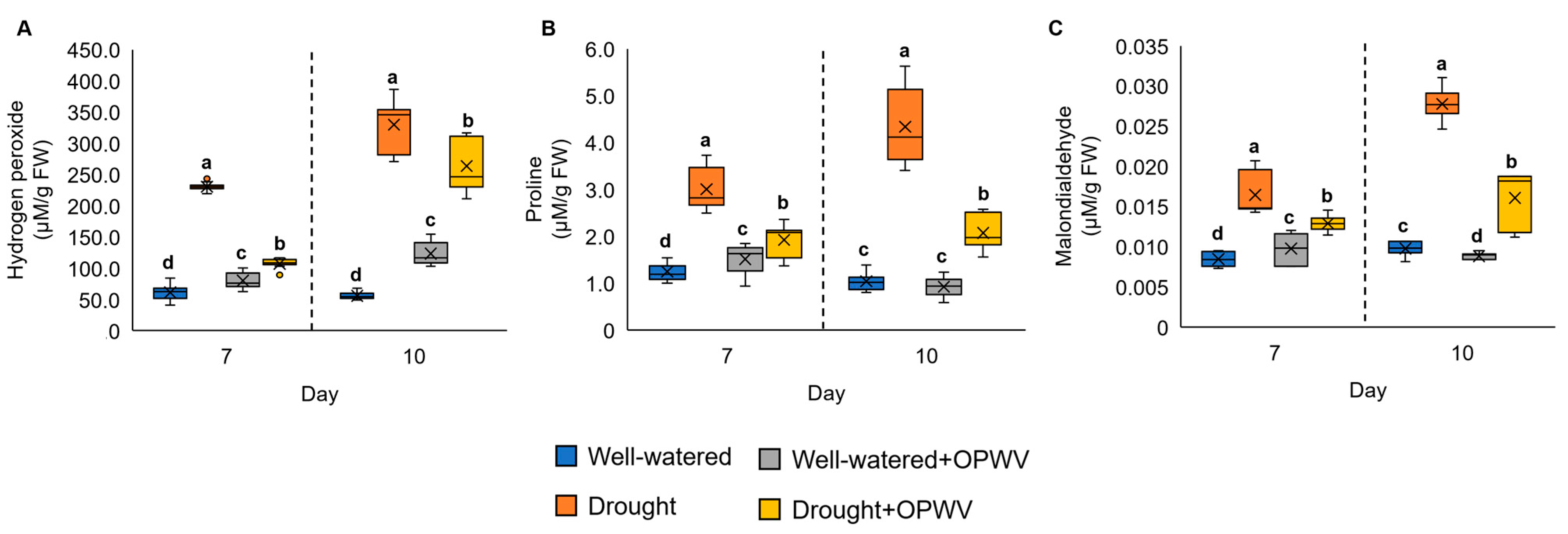
2.4. Hydrogen Peroxide, Osmolyte and Lipid Peroxidation of OPWV-Treated Pandanus amaryllifolius
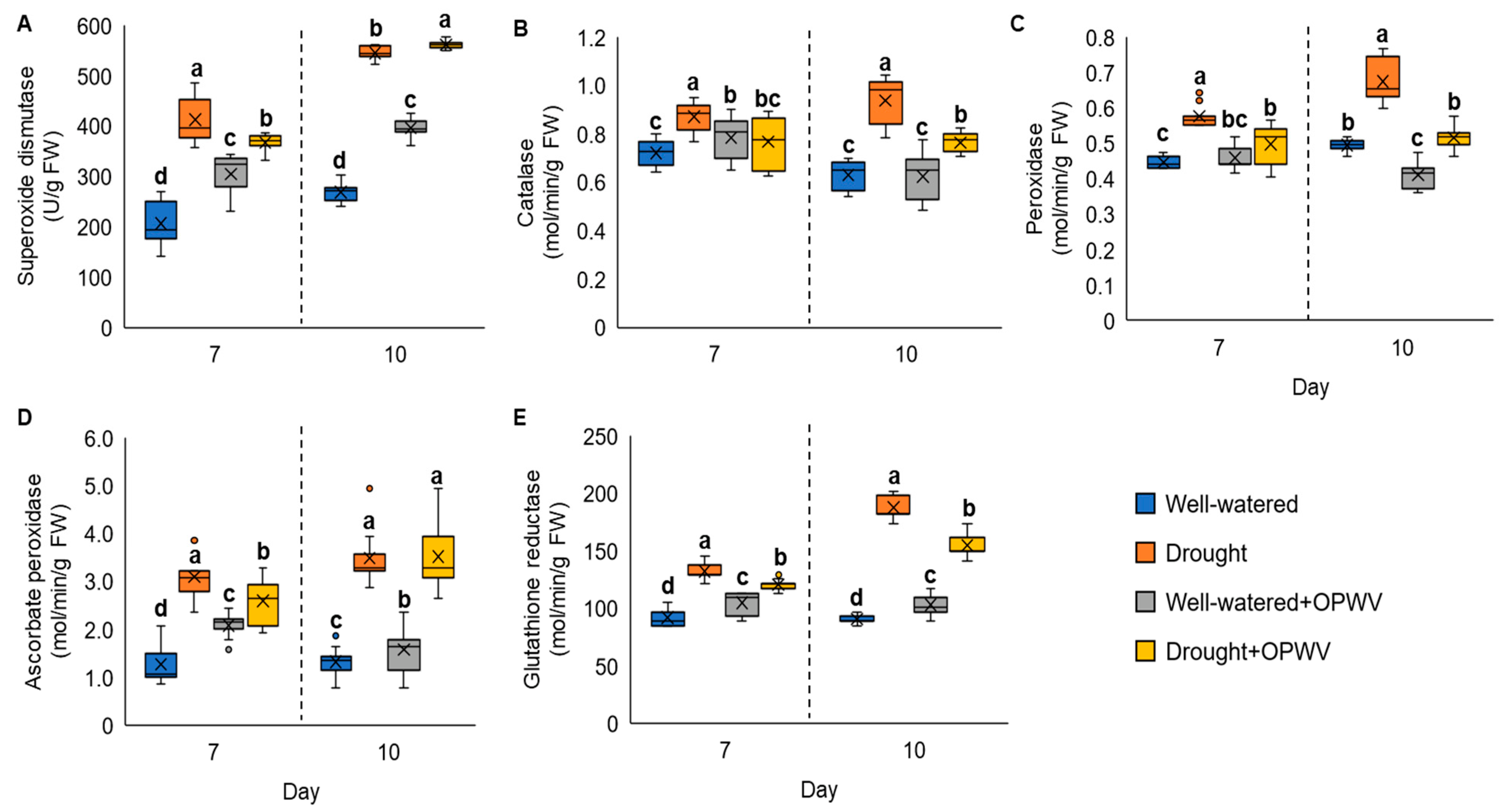
2.5. Antioxidant Enzymes Altered in Pandanus amaryllifolius in Response to OPWV and Drought Stress
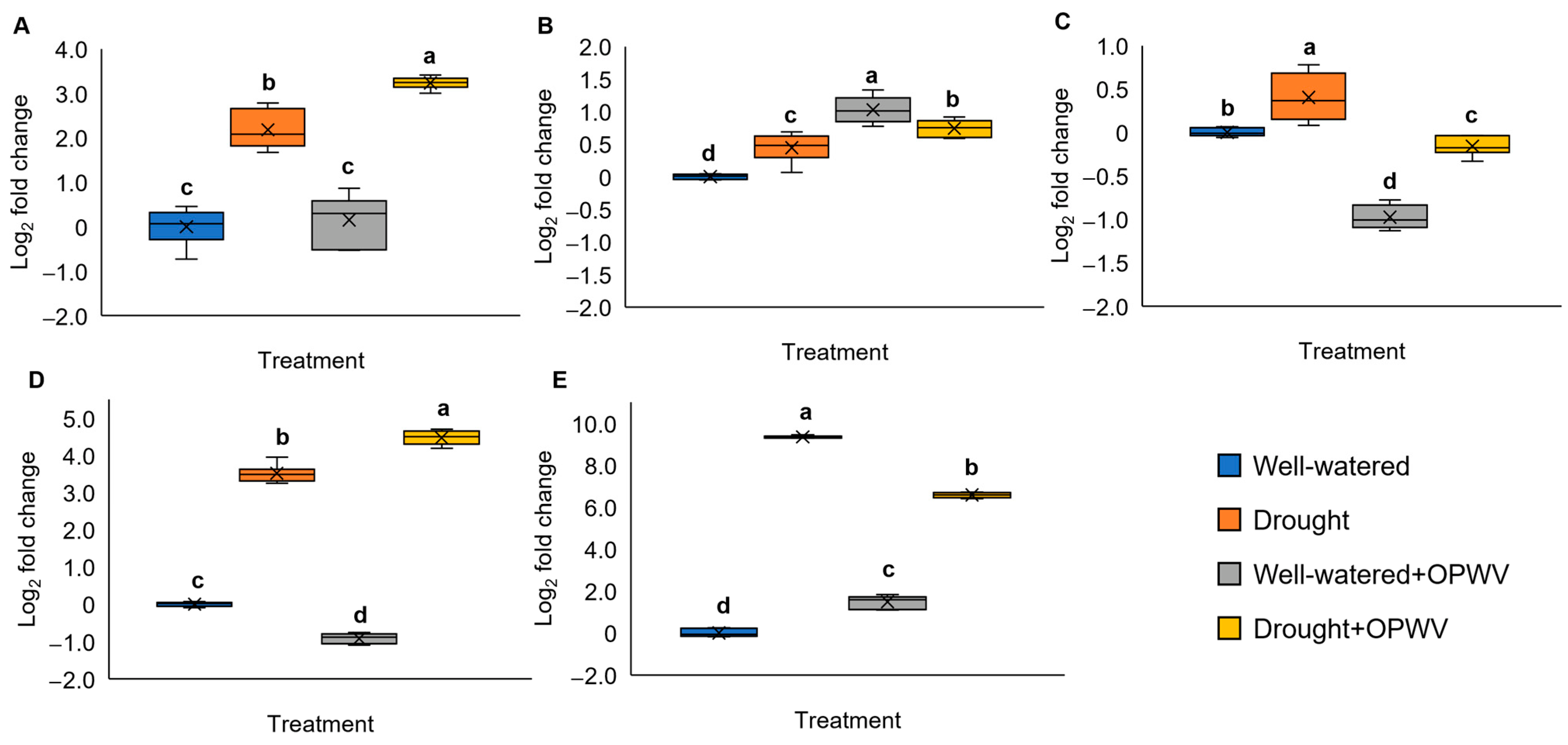
2.6. Pandanus amaryllifolius Drought-Stressed Responsive Genes Triggered with OPWV Application
2.7. OPWV GC-MS Profiling
3. Discussion
3.1. OPWV Induces Morphological Alteration and Improves Drought Tolerance
3.2. OPWV Ameliorated Photosynthetic Pigments
3.3. OPWV Mitigated Drought Stress Indicators
3.4. Enhanced Antioxidant Responses with OPWV Application
3.5. Carbohydrate-Related and Drought-Responsive Genes Altered after OPWV Treatment
3.6. OPWV Compound Profile Elucidated
4. Materials and Methods
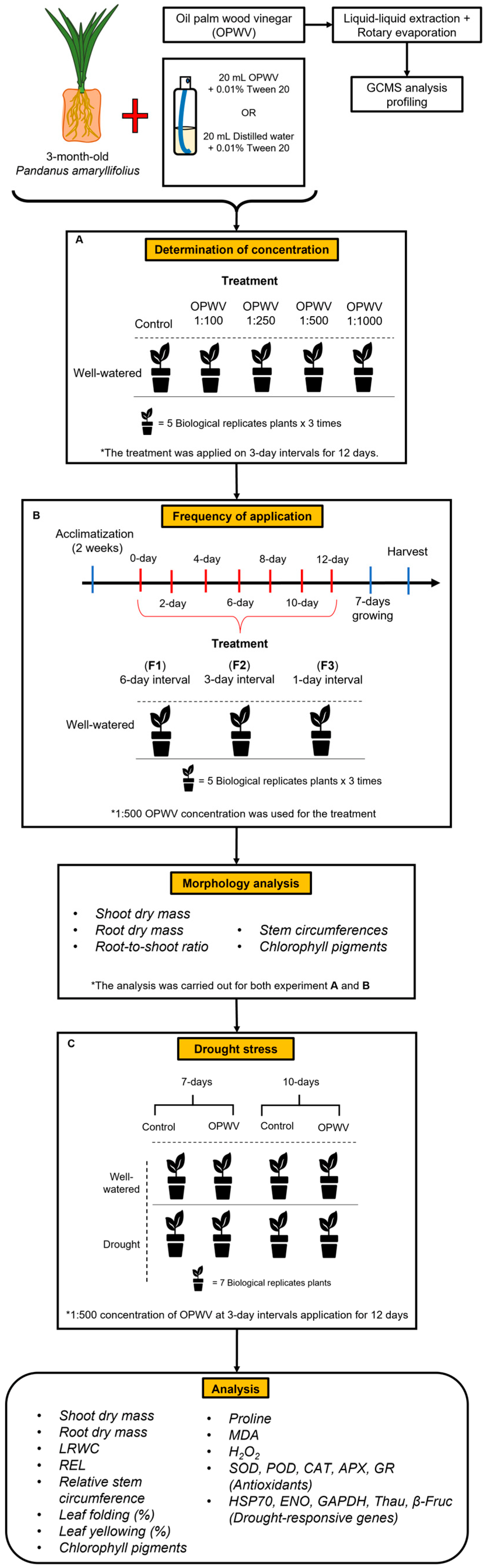
4.1. Plant Material
4.2. Experimental Design and Foliar Application
4.3. Determination of Photosynthetic Pigment Contents
- Ca = Chlorophyll a
- Cb = Chlorophyll b
4.4. Drought Treatment
4.5. Leaf Relative Water Content
4.6. Relative Electrolyte Leakage
4.7. Plant Weight and Relative Stem Circumference
4.8. Examination of Folded and Yellowing Leaves
4.9. Malondialdehyde Content
- A532–A600 = Absorbance of MDA–TBA
- VTr = Volume of reaction (mL)
- Ve = Volume of enzyme extract (mL)
4.10. Proline Content
4.11. Antioxidant Enzyme Assays
- ∆A = Difference in absorbance
- VTr = Volume of reaction (mL)
- Ve = Volume of enzyme extract (mL)
- ∆t = Difference in time of absorbance (min)
- For CAT, ε(Hydrogen peroxide) = 36.0 mol−1 cm−1
- For APX, ε(Ascorbic acid) = 2.8 mmol−1 cm−1
- For POD, ε(Tetraguaiacol) = 26.6 mol−1 cm−1
- For GR, ε(NADPH) = 6220 mol−1 cm−1
4.11.1. Catalase
4.11.2. Ascorbate Peroxidase
4.11.3. Peroxidase
4.11.4. Glutathione Reductase
4.11.5. Superoxide Dismutase
4.11.6. Hydrogen Peroxide
4.12. RNA Extraction and cDNA Synthesis
4.13. Quantitative Real-Time PCR
4.14. Oil Palm Wood Vinegar (OPWV) Gas Chromatography-Mass Spectrometry (GC-MS) Profiling
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Teoh, E.Y.; Teo, C.H.; Baharum, N.A.; Pua, T.-L.; Tan, B.C. Waterlogging Stress Induces Antioxidant Defense Responses, Aerenchyma Formation and Alters Metabolisms of Banana Plants. Plants 2022, 11, 2052. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Fadiji, A.E.; Santoyo, G.; Yadav, A.N.; Babalola, O.O. Efforts towards Overcoming Drought Stress in Crops: Revisiting the Mechanisms Employed by Plant Growth-Promoting Bacteria. Front. Microbiol. 2022, 13, 962427. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Lau, S.-E.; Teo, W.F.A.; Teoh, E.Y.; Tan, B.C. Microbiome Engineering and Plant Biostimulants for Sustainable Crop Improvement and Mitigation of Biotic and Abiotic Stresses. Discov. Food 2022, 2, 9. [Google Scholar] [CrossRef]
- Frioni, T.; VanderWeide, J.; Palliotti, A.; Tombesi, S.; Poni, S.; Sabbatini, P. Foliar vs. Soil Application of Ascophyllum nodosum Extracts to Improve Grapevine Water Stress Tolerance. Sci. Hortic. 2021, 277, 109807. [Google Scholar] [CrossRef]
- Ali, E.F.; El-Shehawi, A.M.; Ibrahim, O.H.M.; Abdul-Hafeez, E.Y.; Moussa, M.M.; Hassan, F.A.S. A Vital Role of Chitosan Nanoparticles in Improvisation the Drought Stress Tolerance in Catharanthus roseus (L.) through Biochemical and Gene Expression Modulation. Plant Physiol. Biochem. 2021, 161, 166–175. [Google Scholar] [CrossRef]
- Zhang, K.; Khan, Z.; Liu, J.; Luo, T.; Zhu, K.; Hu, L.; Bi, J.; Luo, L. Germination and Growth Performance of Water-Saving and Drought-Resistant Rice Enhanced by Seed Treatment with Wood Vinegar and Biochar under Dry Direct-Seeded System. Agronomy 2022, 12, 1223. [Google Scholar] [CrossRef]
- Voko, M.P.; Kulkarni, M.G.; Ngoroyemoto, N.; Gupta, S.; Finnie, J.F.; Van Staden, J. Vermicompost Leachate, Seaweed Extract and Smoke-Water Alleviate Drought Stress in Cowpea by Influencing Phytochemicals, Compatible Solutes and Photosynthetic Pigments. Plant Growth Regul. 2022, 97, 327–342. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, X.; Dong, J. Preparation, Chemical Constituents and Antimicrobial Activity of Pyroligneous Acids from Walnut Tree Branches. J. Anal. Appl. Pyrolysis 2010, 87, 24–28. [Google Scholar] [CrossRef]
- Yang, J.-F.; Yang, C.-H.; Liang, M.-T.; Gao, Z.-J.; Wu, Y.-W.; Chuang, L.-Y. Chemical Composition, Antioxidant, and Antibacterial Activity of Wood Vinegar from Litchi chinensis. Molecules 2016, 21, 1150. [Google Scholar] [CrossRef]
- Loo, A.Y.; Jain, K.; Darah, I. Antioxidant Activity of Compounds Isolated from the Pyroligneous Acid, Rhizophora apiculata. Food Chem. 2008, 107, 1151–1160. [Google Scholar] [CrossRef]
- Azadfar, M.; Gao, A.H.; Bule, M.V.; Chen, S. Structural Characterization of Lignin: A Potential Source of Antioxidants Guaiacol and 4-Vinylguaiacol. Int. J. Biol. Macromol. 2015, 75, 58–66. [Google Scholar] [CrossRef]
- Ozturk Sarikaya, S.B. Acethylcholinesterase Inhibitory Potential and Antioxidant Properties of Pyrogallol. J. Enzyme Inhib. Med. Chem. 2015, 30, 761–766. [Google Scholar] [CrossRef]
- Zhu, K.; Gu, S.; Liu, J.; Luo, T.; Khan, Z.; Zhang, K.; Hu, L. Wood Vinegar as a Complex Growth Regulator Promotes the Growth, Yield, and Quality of Rapeseed. Agronomy 2021, 11, 510. [Google Scholar] [CrossRef]
- Vannini, A.; Moratelli, F.; Monaci, F.; Loppi, S. Effects of Wood Distillate and Soy Lecithin on the Photosynthetic Performance and Growth of Lettuce (Lactuca sativa L.). SN Appl. Sci. 2021, 3, 113. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, L.; Song, Q.; Wang, S.; Wang, Y.; Ge, Y. Root Proteomics Reveals the Effects of Wood Vinegar on Wheat Growth and Subsequent Tolerance to Drought Stress. Int. J. Mol. Sci. 2019, 20, 943. [Google Scholar] [CrossRef]
- Amnan, M.A.M.; Aizat, W.M.; Khaidizar, F.D.; Tan, B.C. Drought Stress Induces Morpho-Physiological and Proteome Changes of Pandanus amaryllifolius. Plants 2022, 11, 221. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z. Profiling of phenolic compounds and their antioxidant and anticancer activities in pandan (Pandanus amaryllifolius Roxb.) extracts from different locations of Malaysia. BMC Complement. Altern. Med. 2013, 13, 341. [Google Scholar] [CrossRef]
- Lau, S.-E.; Hamdan, M.F.; Pua, T.-L.; Saidi, N.B.; Tan, B.C. Plant Nitric Oxide Signaling under Drought Stress. Plants 2021, 10, 360. [Google Scholar] [CrossRef]
- Ye, H.; Song, L.; Schapaugh, W.T.; Ali, M.L.; Sinclair, T.R.; Riar, M.K.; Mutava, R.N.; Li, Y.; Vuong, T.; Valliyodan, B.; et al. The Importance of Slow Canopy Wilting in Drought Tolerance in Soybean. J. Exp. Bot. 2020, 71, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Amnan, M.A.M.; Pua, T.-L.; Lau, S.-E.; Tan, B.C.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Komatsu, S. Osmotic Stress in Banana Is Relieved by Exogenous Nitric Oxide. PeerJ 2021, 9, e10879. [Google Scholar] [CrossRef] [PubMed]
- Rasul, F.; Gupta, S.; Olas, J.J.; Gechev, T.; Sujeeth, N.; Mueller-Roeber, B. Priming with a Seaweed Extract Strongly Improves Drought Tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 1469. [Google Scholar] [CrossRef]
- Adiba, A.; Hssaini, L.; Haddioui, A.; Hamdani, A.; Charafi, J.; El Iraqui, S.; Razouk, R. Pomegranate Plasticity to Water Stress: Attempt to Understand Interactions between Cultivar, Year and Stress Level. Heliyon 2021, 7, e07403. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Hosseini, M.S.; Daneshvar Hakimi Meybodi, N.; Abadía, J.; Germ, M.; Gholami, R.; Abdelrahman, M. Evaluation of Drought Tolerance in Three Commercial Pomegranate Cultivars Using Photosynthetic Pigments, Yield Parameters and Biochemical Traits as Biomarkers. Agric. Water Manag. 2022, 261, 107357. [Google Scholar] [CrossRef]
- Ma, J.; Islam, F.; Ayyaz, A.; Fang, R.; Hannan, F.; Farooq, M.A.; Ali, B.; Huang, Q.; Sun, R.; Zhou, W. Wood Vinegar Induces Salinity Tolerance by Alleviating Oxidative Damages and Protecting Photosystem II in Rapeseed Cultivars. Ind. Crops Prod. 2022, 189, 115763. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and Biochemistry of 4-Hydroxynonenal, Malonaldehyde and Related Aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Habib, N.; Ali, Q.; Ali, S.; Javed, M.T.; Zulqurnain Haider, M.; Perveen, R.; Shahid, M.R.; Rizwan, M.; Abdel-Daim, M.M.; Elkelish, A.; et al. Use of Nitric Oxide and Hydrogen Peroxide for Better Yield of Wheat (Triticum aestivum L.) under Water Deficit Conditions: Growth, Osmoregulation, and Antioxidative Defense Mechanism. Plants 2020, 9, 285. [Google Scholar] [CrossRef]
- Hameed, A.; Akram, N.A.; Saleem, M.H.; Ashraf, M.; Ahmed, S.; Ali, S.; Abdullah Alsahli, A.; Alyemeni, M.N. Seed Treatment with α-Tocopherol Regulates Growth and Key Physio-Biochemical Attributes in Carrot (Daucus carota L.) Plants under Water Limited Regimes. Agronomy 2021, 11, 469. [Google Scholar] [CrossRef]
- Gohari, G.; Alavi, Z.; Esfandiari, E.; Panahirad, S.; Hajihoseinlou, S.; Fotopoulos, V. Interaction between Hydrogen Peroxide and Sodium Nitroprusside Following Chemical Priming of Ocimum basilicum L. against Salt Stress. Physiol. Plant. 2020, 168, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in Modulating Proline Metabolism in Plants for Salt and Drought Stress Tolerance: Phytohormones, Mineral Nutrients and Transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Dien, D.C.; Mochizuki, T.; Yamakawa, T. Effect of Various Drought Stresses and Subsequent Recovery on Proline, Total Soluble Sugar and Starch Metabolisms in Rice (Oryza sativa L.) Varieties. Plant Prod. Sci. 2019, 22, 530–545. [Google Scholar] [CrossRef]
- Furlan, A.L.; Bianucci, E.; Giordano, W.; Castro, S.; Becker, D.F. Proline Metabolic Dynamics and Implications in Drought Tolerance of Peanut Plants. Plant Physiol. Biochem. 2020, 151, 566–578. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.-S.E.; M. Abu-Elsaoud, A.; Hafez, Y.M. Exogenous Application of Proline and Salicylic Acid Can Mitigate the Injurious Impacts of Drought Stress on Barley Plants Associated with Physiological and Histological Characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- AlKahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of Silicon and Proline Application on the Oxidative Machinery in Drought-Stressed Sugar Beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Rady, M.M.; Boriek, S.H.K.; Abd El-Mageed, T.A.; Seif El-Yazal, M.A.; Ali, E.F.; Hassan, F.A.S.; Abdelkhalik, A. Exogenous Gibberellic Acid or Dilute Bee Honey Boosts Drought Stress Tolerance in Vicia Faba by Rebalancing Osmoprotectants, Antioxidants, Nutrients, and Phytohormones. Plants 2021, 10, 748. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, Y.; Xu, Z.; Chen, Z.; Duan, L. Physiological and Transcriptome Profiling Analyses Reveal Important Roles of Coronatine in Improving Drought Tolerance of Tobacco. J. Plant Growth Regul. 2020, 39, 1346–1358. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, F.; Feng, Z.; Li, Y.; Ma, M.; Wang, G.; Zhao, H. A Vacuolar Invertase CsVI2 Regulates Sucrose Metabolism and Increases Drought Tolerance in Cucumis sativus L. Int. J. Mol. Sci. 2022, 23, 176. [Google Scholar] [CrossRef]
- Augustine, S.M.; Cherian, A.V.; Syamaladevi, D.P.; Subramonian, N. Erianthus arundinaceus HSP70 (EaHSP70) Acts as a Key Regulator in the Formation of Anisotropic Interdigitation in Sugarcane (Saccharum spp. Hybrid) in Response to Drought Stress. Plant Cell Physiol. 2015, 56, 2368–2380. [Google Scholar] [CrossRef]
- Djemal, R.; Bahloul, O.; Khoudi, H. A novel Thaumatin-Like Protein From Durum Wheat, TdPR-5, is Homologous To Known Plant Allergens, Responsive to Stress Exposure, and Confers Multiple-Abiotic Stress Tolerances to Transgenic Yeast. Plant Gene 2022, 31, 100360. [Google Scholar] [CrossRef]
- Kappachery, S.; Baniekal-Hiremath, G.; Yu, J.W.; Park, S.W. Effect of Over-and under-Expression of Glyceraldehyde 3-Phosphate Dehydrogenase on Tolerance of Plants to Water-Deficit Stress. Plant Cell Tissue Organ Cult. 2015, 121, 97–107. [Google Scholar] [CrossRef]
- Misra, R.C.; Sandeep; Kamthan, M.; Kumar, S.; Ghosh, S. A Thaumatin-like Protein of Ocimum basilicum Confers Tolerance to Fungal Pathogen and Abiotic Stress in Transgenic Arabidopsis. Sci. Rep. 2016, 6, 25340. [Google Scholar] [CrossRef] [PubMed]
- Odeny Ojola, P.; Nyaboga, E.N.; Njiru, P.N.; Orinda, G. Overexpression of Rice Thaumatin-like Protein (Ostlp) Gene in Transgenic Cassava Results in Enhanced Tolerance to Colletotrichum gloeosporioides f. sp. Manihotis. J. Genet. Eng. Biotechnol. 2018, 16, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, H.; Rajput, R.; Pandey, A.; Upadhyay, S.K. Molecular Characterization Revealed the Role of Thaumatin-Like Proteins of Bread Wheat in Stress Response. Front. Plant Sci. 2022, 12, 807448. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Feng, X.; Yu, J. Wood Vinegar Resulting From the Pyrolysis of Apple Tree Branches for Annual Bluegrass Control. Ind. Crops Prod. 2021, 147, 114193. [Google Scholar] [CrossRef]
- Oramahi, H.A.; Yoshimura, T.; Diba, F.; Setyawati, D. Nurhaida Antifungal and Antitermitic Activities of Wood Vinegar from Oil Palm Trunk. J. Wood Sci. 2018, 64, 311–317. [Google Scholar] [CrossRef]
- Xue, R.; Cui, E.-L.; Hu, G.-Q.; Zhu, M.-Q. The Composition, Physicochemical Properties, Antimicrobial and Antioxidant Activity of Wood Vinegar Prepared by Pyrolysis of Eucommia ulmoides Oliver Branches under Different Refining Methods and Storage Conditions. Ind. Crops Prod. 2022, 178, 114586. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Huang, S.; Li, W.; Ashraf, U.; Ma, L.; Mo, Z. Exogenous Melatonin and Catechol Application Modulate Physio-Biochemical Attributes and Early Growth of Fragrant Rice Under Cd Toxicity. J. Soil Sci. Plant Nutr. 2021, 21, 2285–2296. [Google Scholar] [CrossRef]
- Singh, N.; Mansoori, A.; Jiwani, G.; Solanke, A.U.; Thakur, T.K.; Kumar, R.; Chaurasiya, M.; Kumar, A. Antioxidant and Antimicrobial Study of Schefflera vinosa Leaves Crude Extracts against Rice Pathogens. Arab. J. Chem. 2021, 14, 103243. [Google Scholar] [CrossRef]
- Doyle, S.M.; Rigal, A.; Grones, P.; Karady, M.; Barange, D.K.; Majda, M.; Pařízková, B.; Karampelias, M.; Zwiewka, M.; Pěnčík, A.; et al. A Role for the Auxin Precursor Anthranilic Acid in Root Gravitropism via Regulation of PIN-FORMED Protein Polarity and Relocalisation in Arabidopsis. New Phytol. 2019, 223, 1420–1432. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Plant Cell Membranes; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Turner, N.C. Techniques and Experimental Approaches for the Measurement of Plant Water Status. Plant Soil 1981, 58, 339–366. [Google Scholar] [CrossRef]
- Quan, W.; Hu, Y.; Mu, Z.; Shi, H.; Chan, Z. Overexpression of AtPYL5 under the Control of Guard Cell Specific Promoter Improves Drought Stress Tolerance in Arabidopsis. Plant Physiol. Biochem. 2018, 129, 150–157. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis, 2nd ed.; Bergmeyer, H.U., Ed.; Academic Press: Cambridge, MA, USA, 1974; pp. 673–684. ISBN 978-0-12-091302-2. [Google Scholar]
- Chen, G.-X.; Asada, K. Inactivation of Ascorbate Peroxidase by Thiols Requires Hydrogen Peroxide. Plant Cell Physiol. 1992, 33, 117–123. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of Catalases and Peroxidases: In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1955; Volume 2, pp. 764–775. [Google Scholar]
- Mannervik, B. Measurement of Glutathione Reductase Activity. Curr. Protoc. Toxicol. 1999, 7.2.1–7.2.4. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants: Protective Role of Exogenous Polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Asif, M.H.; Dhawan, P.; Nath, P. A Simple Procedure for the Isolation of High Quality RNA from Ripening Banana Fruit. Plant Mol. Biol. Report. 2000, 18, 109–115. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.; Sultana, M.; Kumar, V.; Gangopadhyay, G. Isolation and Characterization of BADH2 Gene from in Vitro Propagated Pandanus amaryllifolius Roxb. Plant Cell Tissue Organ Cult. 2017, 130, 131–140. [Google Scholar] [CrossRef]
- Mahmud, K.N.; Yahayu, M.; Siti Hajar, M.S.; Rizan, N.H.; ChaiBing, M.; Mustafa, N.F.; Ngadiran, S.; Ujang, S.; Zakaria, Z.A. Evaluation on Efficiency of Pyroligneous Acid from Palm Kernel Shell as Antifungal and Solid Pineapple Biomass as Antibacterial and Plant Growth Promoter. Sains Malays. 2016, 45, 1423–1434. [Google Scholar]
- Mohd Hamzah, M.A.A.; Hasham, R.; Nik Malek, N.A.N.; Raja Sulong, R.S.; Yahayu, M.; Abdul Razak, F.I.; Zakaria, Z.A. Structural-Based Analysis of Antibacterial Activities of Acid Condensate from Palm Kernel Shell. Biomass Convers. Biorefin. 2022. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identified Compound | Molecular Formula | Retention Time (RT) | m/z | Area under Peak | Concentration (ppm/µL) | Concentration (ppm/mL) |
|---|---|---|---|---|---|---|---|
| 1 | Phenyl carbamate | C7H7NO2 | 7.111 | 94.15 | 48,765,158 | 340.55 | 340,550 |
| 2 | Phenol | C6H6O | 6.868 | 94.05 | 12,407,245 | 86.65 | 86,650 |
| 3 * | Pyridine, 2,4,6-trimethyl- | C8H11N | 7.191 | 121.05 | 7,159,676 | 50.00 | 50,000 |
| 4 | Guaiacol | C7H8O2 | 9.079 | 109.05 | 3,369,657 | 23.53 | 23,530 |
| 5 | Syringol | C8H10O3 | 13.856 | 154.05 | 2,215,362 | 15.47 | 15,470 |
| 6 | 2-(2′,4′,4′,6′,6′,8′,8′-Heptamethyltetrasiloxan-2′-yloxy)-2,4,4,6,6,8,8,10,10-nonamethylcyclopentasiloxane | C16H48O10Si9 | 19.103 | 73.1 | 1,836,709 | 12.83 | 12,830 |
| 7 | 3-Isopropoxy-1,1,1,7,7,7-hexamethyl-3,5,5-tris(trimethylsiloxy) tetrasiloxane | C18H52O7Si7 | 23.366 | 73.1 | 1,741,464 | 12.16 | 12,160 |
| 8 | Catechol | C6H6O2 | 11.458 | 110.05 | 1,096,505 | 7.66 | 7660 |
| 9 | 3-Oxabicyclo[3.3.0]oct-7-en-2-one,4-methoxy- | C9H12O3 | 8.946 | 109.05 | 961,798 | 6.72 | 6720 |
| 10 | o-Creosol | C7H8O | 8.271 | 108.05 | 803,567 | 5.61 | 5610 |
| 11 | 1,2-Cyclopentanedione, 3-methyl- | C6H8O2 | 8.011 | 112.05 | 753,407 | 5.26 | 5260 |
| 12 | 1,2,4-Trimethoxybenzene | C9H12O3 | 15.875 | 168.1 | 546,464 | 3.82 | 3820 |
| 13 | 4-Ethylguaiacol | C9H12O2 | 12.918 | 137.05 | 513,484 | 3.59 | 3590 |
| 14 | 3-Methoxyycatechol | C7H8O3 | 12.644 | 140.05 | 508,764 | 3.55 | 3550 |
| 15 | Methoxyacetylene | C10H14O3 | 17.18 | 167.1 | 242,644 | 1.69 | 1690 |
| 16 | Creosol | C8H10O2 | 10.938 | 123 | 190,595 | 1.33 | 1330 |
| 17 | Methyl palmitate | C17H34O2 | 22.761 | 74.1 | 116,444 | 0.81 | 810 |
| Gene | Sequences, 5′–3′ |
|---|---|
| HSP70_Pandan (PanHSP70) | F-ACCTACAAGGGTGAGGAGAAG R-GAAATAGGCAGGGACAGTGATG |
| GAPDH_Pandan (PanGAPDH) [65] | F-AGGGTGGTGCCAAGAAGGT R-CCACCTCTCCAGTCCTT |
| Enolase_Pandan (PanENO) | F-TGAGTGATGGCACTTACGCC R-ACGTTCTCCACAGCCTTGAG |
| Thaumatin_Pandan (PanThau) | F-TCGCTGTCCTTCTCCTTTGG R-CACCTTGTGAGGAATGCAGC |
| β-fructofuranosidase_Pandan (Panβ-Fruc) | F-GAACCCTGGATGGTATCGGG R-CCGGCAAATGCTCCTAAGTG |
| * Actin_Pandan | F-GAGGCTATTCCTTCACCACTAC R-GTCTCAAGCTCCTCCTCATAATC |
| * Elongation factor-1_Pandan | F-TCTTCACAAAGCCAGCATCTC R-GACTGCCACACCTCTCATATTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Amnan, M.A.; Teo, W.F.A.; Aizat, W.M.; Khaidizar, F.D.; Tan, B.C. Foliar Application of Oil Palm Wood Vinegar Enhances Pandanus amaryllifolius Tolerance under Drought Stress. Plants 2023, 12, 785. https://doi.org/10.3390/plants12040785
Mohd Amnan MA, Teo WFA, Aizat WM, Khaidizar FD, Tan BC. Foliar Application of Oil Palm Wood Vinegar Enhances Pandanus amaryllifolius Tolerance under Drought Stress. Plants. 2023; 12(4):785. https://doi.org/10.3390/plants12040785
Chicago/Turabian StyleMohd Amnan, Muhammad Asyraf, Wee Fei Aaron Teo, Wan Mohd Aizat, Fiqri Dizar Khaidizar, and Boon Chin Tan. 2023. "Foliar Application of Oil Palm Wood Vinegar Enhances Pandanus amaryllifolius Tolerance under Drought Stress" Plants 12, no. 4: 785. https://doi.org/10.3390/plants12040785
APA StyleMohd Amnan, M. A., Teo, W. F. A., Aizat, W. M., Khaidizar, F. D., & Tan, B. C. (2023). Foliar Application of Oil Palm Wood Vinegar Enhances Pandanus amaryllifolius Tolerance under Drought Stress. Plants, 12(4), 785. https://doi.org/10.3390/plants12040785