
SmRAV1, an AP2 and B3 Transcription Factor, Positively Regulates Eggplant’s Response to Salt Stress

College of Horticulture and Landscape Architecture, Yangzhou University, Yangzhou 225009, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2023, 12(24), 4174; https://doi.org/10.3390/plants12244174
Submission received: 14 November 2023
/
Revised: 9 December 2023
/
Accepted: 12 December 2023
/
Published: 15 December 2023
(This article belongs to the Special Issue Growth, Development, and Stress Response of Horticulture Plants)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Salt stress is a lethal abiotic stress threatening global food security on a consistent basis. In this study, we identified an AP2 and B3 domain-containing transcription factor (TF) named SmRAV1, and its expression levels were significantly up-regulated by NaCl, abscisic acid (ABA), and hydrogen peroxide (H2O2) treatment. High expression of SmRAV1 was observed in the roots and sepal of mature plants. The transient expression assay in Nicotiana benthamiana leaves revealed that SmRAV1 was localized in the nucleus. Silencing of SmRAV1 via virus-induced gene silencing (VIGS) decreased the tolerance of eggplant to salt stress. Significant down-regulation of salt stress marker genes, including SmGSTU10 and SmNCED1, was observed. Additionally, increased H2O2 content and decreased catalase (CAT) enzyme activity were recorded in the SmRAV1-silenced plants compared to the TRV:00 plants. Our findings elucidate the functions of SmRAV1 and provide opportunities for generating salt-tolerant lines of eggplant.
1. Introduction
Salinity/salt stress is the second-biggest abiotic factor affecting agricultural productivity worldwide by damaging numerous physiological, biochemical, and molecular processes [1,2,3,4,5,6,7]. Breeding salt-tolerant varieties would certainly minimize the damaging effects of salinity [8]. To breed new verities, it is imminently important to under the intricate signaling pathway regulating plant response to salt stress. Despite the plethora of literature available, research gaps are still at large in the field of salt stress biology. Understanding the transcription factors mediating the underlying mechanism of salt stress would add another cornerstone to the scientific research.
Transcription factors play vital roles in plant growth and plant responses to abiotic stresses [9,10]. The RAV transcription factors contain a B3 DNA-binding domain and/or an APETALA2 (AP2) DNA-binding domain [11,12,13]. RAV transcription factors regulate the transcript expression of downstream target genes by binding to core sequences of 5′-CAACA-3′ and 5′-CACCTG-3′ [14] and function in regulating vegetative growth [15,16,17], flowering [11,18,19], phytohormone signal transduction [20,21], biotic stresses response [22,23,24,25], and abiotic stresses response such as salt [21,26,27,28], and drought [21,29]. To date, RAV gene family members have been identified in multiple plant species, such as six RAV genes in Arabidopsis thaliana [13], 26 in wheat [30], 33 in Gossypium hirsutum [31], 11 in pear [32], 15 in rice [33], 13 in soybean [34], and five in peach [35]. RAV proteins have been reported in Arabidopsis thaliana, garnering seed germination [17], seedling development [17], salt stress response [21,28], drought stress response [21], and flowering [36]. For example, Arabidopsis RAV1 interacts with SnRK2 kinases to repress the transcript expression of ABI3, ABI4, and ABI5, promoting seed germination and seedling development in an abscisic acid (ABA)-dependent manner [17]. The overexpression of Arabidopsis RAVs (RAV1, RAV1L, RAV2) decreased the tolerance of Arabidopsis plants to drought and salt stresses in an ABA-independent manner [21]. The CmTEM1 overexpression delayed chrysanthemum flowering by directly binding to the flowering integrator CmAFL1 promoter, thus repressing this gene’s expression [11]. In addition, RAV transcription factors also played an important role against pathogenic microorganism attacks in plants. The RNA-silencing suppressor (RSS) 24-kDa protein (p24) of grapevine leafroll-associated virus 2 repressed the direct activation of RAV transcription factor VvRAV1 in Vitis vinifera to grapevine pathogenesis-related protein 1 (PR1) expression by interacting with VvRAV1, leading to a weakened resistance of grape to pathogens [37]. Although some reports indicated that RAV transcription factors played a vital role in plant against salt stress, the functions and mechanisms of RAV transcription factors against salt stress remain largely unclear.
Eggplant (Solanum melongena) is a popular Solanaceae vegetable that is sensitive to salinity stress [38]. Soil salinity significantly affects eggplant growth, development, and overall yield. The topics of salinity and seeking salt-tolerant cultivars are of great interest to agronomists and growers. Therefore, it is helpful to breed new eggplant cultivars with high salinity tolerance. In this study, we identified a RAV transcription factor family member named SmRAV1, which was significantly up-regulated by salt stress treatment, by analyzing mRNA sequencing data of eggplant roots treated with salt stress according to our previous study [39]. We further analyzed SmRAV1 expression and the function in eggplant against salt stress, and demonstrated that SmRAV1 positively regulates the tolerance of eggplant to salt stress.
2. Results
2.1. Sequence Analysis of SmRAV1
From our previous sequencing data, we observed that the SmRAV1 (Smechr1102045) increased significantly in response to salt stress [39] (Figure S1). The SmRAV1 has an ORF (open reading frame) of 1101 bp (base pair) and encoding a 366 bp amino acid. By utilizing the SMART (http://smart.embl-heidelberg.de/, accessed on 11 August 2023) website, we found that SmRAV1 possesses one AP2 and one B3 conserved domain (Figure 1a,d). We further analyzed the secondary and tertiary structures of SmRAV1 via the PRABI-GERLAND (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 11 August 2023) and SWISS-MODEL (https://swissmodel.expasy.org/interactive, accessed on 11 August 2023) websites, respectively. The results showed that the secondary structure of SmRAV1 consisted of four types, including alpha helix (Hh), beta turn (Tt), extended strand (Ee), and random coil (Cc) (Figure 1b, Table S2). The results of tertiary structures of SmRAV1 showed that it shared 82.45% similarities with the E1U2K4.1.A tertiary structure model (Figure 1c). We next performed the multiple alignment of amino acid sequences and found that the SmRAV1 amino acid sequence shared 82, 78, 78, 78%, 76, 69, 64, 61, 61, 53, 58, and 52% sequence similarities with its homologs from the other plant species, including Solanum dulcamara SdRAV1-like (XP_055801403.1), Solanum tuberosum StRAV1 (XP_006366067.1), Solanum stenotomum StRAV1-like (XP_049391936.1), Solanum verrucosum SvRAV1-like (XP_049363435.1), Solanum lycopersicum SlRAV1 (XP_004236999.1), Nicotiana tabacum NtRAV1-like (XP_016445426.1), Vitis vinifera VvRAV1 (UYF10651.1), Solanum lycopersicum SlRAV2 (ABY57635.1), Capsicum annuum CaRAV1 (XP_016548054.2), Zea mays ZmRAV1 (PWZ33711.1), Arabidopsis thaliana AtRAV1 (OAP16099.1), and Oryza sativa Os01g0693400 (NP_001388398.1), respectively (Figure 1d). Moreover, we constructed evolutionary trees by using their amino sequences to analyze the phylogenetic relationships between the above RAV homologs. SmRAV1 had the closest relationship with SdRAV1-like (Figure 1e). In addition, we predicted the cis-elements within the 2000 bp promoter of SmRAV1 via the PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 12 August 2023) website. The SmRAV1 contained multiple cis-elements related to phytohormone response (two ethylene response elements (EREs) and one gibberellin response element (GARE-motif), transcription factor-binding elements (five MYB transcription factor-binding elements (MYB), four MYC transcription factor-binding elements (MYC), two WRKY transcription factor-binding elements (W-box), one bZIP transcription factor-binding element (A-box), and one HD-zip transcription factor binding element (HD-Zip), and stress response element STRE, apart from the elements related to light response (Figure 1f).
2.2. Expression Profiles of SmRAV1 under Different Stresses
The expression of SmRAV1 in the roots of salt-stressed eggplant was analyzed. We found that the expression level of SmRAV1 was significantly up-regulated by salt stress treatment, and reached its peak at 24 h. We also investigated the expression profiles of SmRAV1 under different treatments, including high temperature (HT), low temperature (LT), dehydration stress, ABA, and hydrogen peroxide (H2O2), in eggplant. The result showed that apart from the NaCl treatment, both the ABA and H2O2 treatments could significantly induce the up-regulation of SmRAV1 expression. On the other hand, low expression was recorded in response to HT, LT, and dehydration stress, respectively (Figure 2), implying that SmRAV1 may function in eggplant response to salt stress or be involved in ABA- and H2O2-mediated signal transduction pathways.
2.3. Analysis of Tissue Specific Expression of SmRAV1
The tissue-specific expression was observed in order to understand the role of SmRAV1 in developmental biology. A significantly higher expression was observed in the roots compared to young leaves (YL) and stems (ST). Also, high expression was recorded in the sepals (SE) and flowers (FL) than other tissues (Figure 3).
2.4. Subcellular Localization of SmRAV1
The conserved domain analysis revealed that SmRAV1 was composed of AP2 and B3 domains (Figure 1a). SmRAV1 proteins may localize in the nucleus. For validation, we firstly predicted SmRAV1 subcellular localization by searching the Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/, accessed on 12 August 2023) website using an SmRAV1 amino acid sequence [40]. The result showed that SmRAV1 proteins localized in the nucleus (Figure S2a). We searched the INSP (http://www.csbio.sjtu.edu.cn/bioinf/INSP/, accessed on 12 August 2023) website using the amino acid sequence, which further confirmed the nuclear signals of SmRAV1 [41]. SmRAV1 has an NLS (201VGKLNRLV208) in its amino acid sequence (Figure 1d, Figure S2b). These results suggest that SmRAV1 is probably a nuclear-localized protein. Subsequently, we performed a subcellular localization assay. We constructed the recombinant vector 35S:GFP-SmRAV1 by cloning the SmRAV1 coding sequence (CDS) into plant overexpression vector pBinGFP2. The CDS of AtH2B was fused with a red fluorescence protein (RFP) tag to generate 35S:AtH2B-RFP structure. The AtH2B is generally used as a nuclear localization marker [42,43] (Figure 4a). The Agrobacterium GV3101 cells harboring the 35S:AtH2B-RFP structure were mixed with the GV3101 cells containing the 35S:GFP-SmRAV1 or 35S:GFP (empty vector) structures at a 1:1 ratio, respectively. The Nicotiana benthamiana leaves infiltrated with above Agrobacterium mixtures were used to observe the fluorescence signals using a laser scanning confocal microscope. We found that the green fluorescence signals of SmRAV1-GFP proteins occurred in the nucleus of the epidermic cells in Nicotiana benthamiana leaves, and overlapped with the red nuclear fluorescence signals of localization marker AtH2B-RFP proteins. Meanwhile, the green fluorescence signals of GFP proteins expressed by 35S:GFP vector were distributed in entire cells (Figure 4b), suggesting that SmRAV1 localizes in the nucleus.
2.5. Silencing of SmRAV1 Enhances Susceptibility of Eggplant against Salt Stress
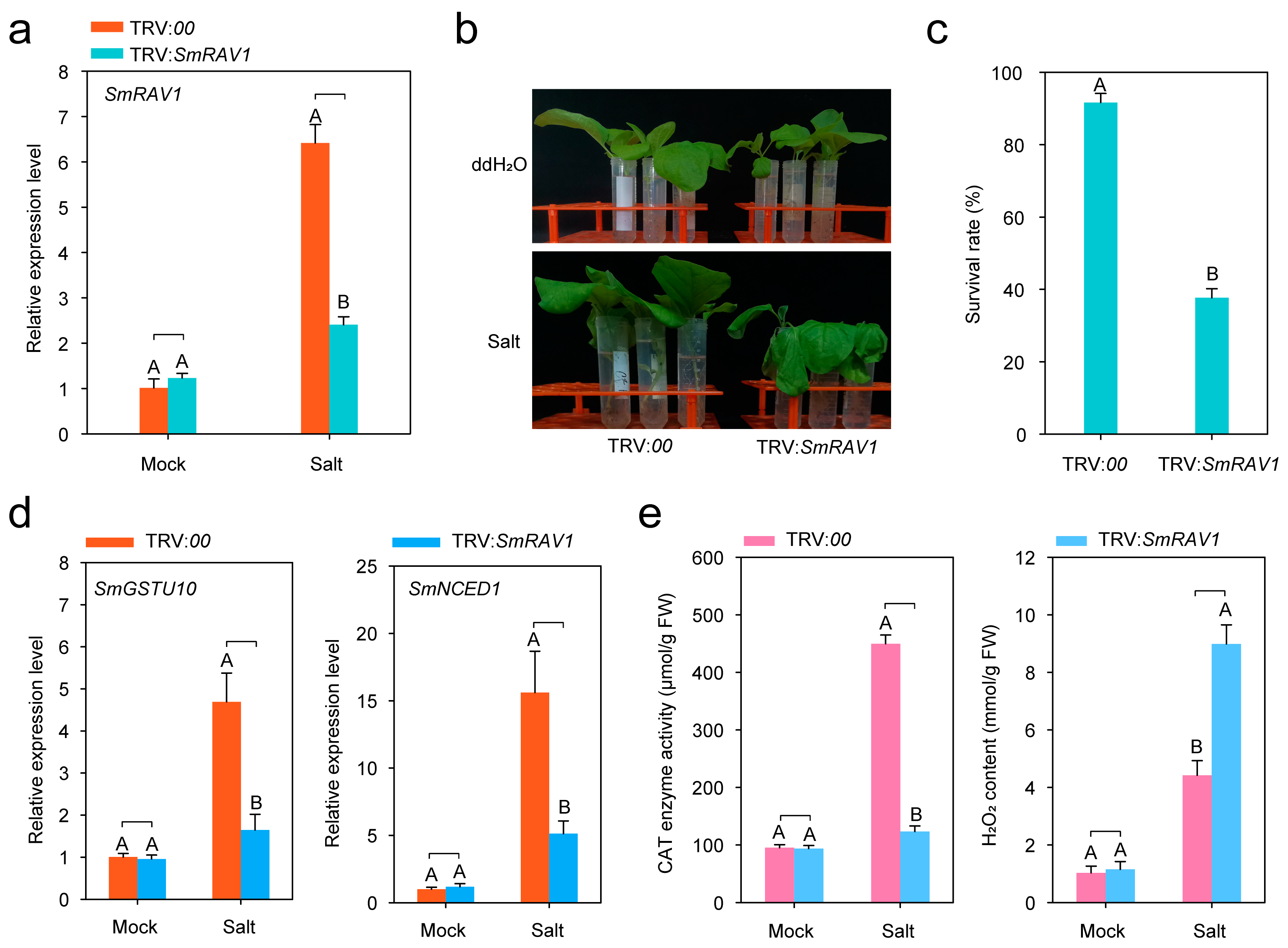
Previous studies have revealed that RAV transcription factors play vital roles against salt stress in plants [21,27,30,44]. In this study, we also found that salt stress could significantly induce SmRAV1 expression in eggplant roots (Figure 2). Therefore, we speculated that SmRAV1 may play an essential role in eggplant’s tolerance against salt stress. To confirm this possibility, we performed a virus-induced gene silencing (VIGS) assay to silence SmRAV1 expression. Firstly, we detected the silencing efficiency of SmRAV1 by RT-qPCR assay. The transcription level of SmRAV1 in the roots of SmRAV1-silenced (TRV:SmRAV1) eggplant plants was significantly reduced (approximately 63%) compared to the control plants (TRV:00) under salt stress treatment (Figure 5a). At 36 h post-treatment, the SmRAV1-silenced eggplant exhibited more serious wilting symptoms than the control plants under 200 mM NaCl solution treatment (Figure 5b), accompanied by a lower survival rate (Figure 5c). In addition, reduced expression of SmGSTU10 and SmNCED1 (Figure 5d), markedly decreased reactive-oxygen-scavenging enzymes catalase (CAT) activity, and increased H2O2 content under salt stress treatment were also observed (Figure 5e). These results suggest that SmRAV1 positively modulates the response of eggplant to salinity stress.
3. Discussion
Transcription factors are proteins that help to turn specific genes “on” or “off” by binding to nearby DNA [45,46,47,48,49]. Accumulating evidence has revealed the various functions of RAV transcription factors in the processes of seed germination [17,21,29], seedling growth and development [16,17,18,50], flowering [11,19], and abiotic and biotic stresses [22,23,24,29,34,51]—salt stress in particular [26,27,28,30,50]. Among the multiple abiotic stresses, salt stress is one of the most important factors threating plants during their entire growth processes. Therefore, it is important to improve the salt tolerance through molecular and biochemical approaches. Although previous studies have reported the functions and regulatory mechanisms of RAV transcription factors, the underlying functions and regulatory mechanisms of RAV transcription factors in plants’ responses to salt stress remain elusive. Herein, we identified a RAV transcription factor family member named SmRAV1 and examined its expression profiles under HT, LT, salt, dehydration, ABA, and H2O2 treatment; subcellular localization; and the function of SmRAV1 in eggplant’s response to salt stress.
By analyzing the mRNA sequencing data of eggplant roots treated with salt stress [39], we obtained a candidate gene named SmRAV1, which had the highest sequence similarity with its homolog RAV1 in Arabidopsis thaliana, and its expression was up-regulated by salt stress (Figure S1). To confirm this result, an induced expression profile of SmRAV1 in eggplant roots treated with NaCl solution was observed (Figure 2). Arabidopsis RAV1 negatively modulates the tolerance of Arabidopsis to drought and salt stress [21]. We analyzed SmRAV1 sequences and found that SmRAV1 harbored one conserved AP2 and B3 domain (Figure 1a), and exhibited the highest sequence similarity with SdRAV1-like in Solanum dulcamara (Figure 1d,e). However, no reports exist of SdRAV1-like involvement in Solanum dulcamara’s response to salt stress. In addition, we found some cis-elements related to phytohormone response and transcription factor binding within the SmRAV1 promoter. These cis-elements were related to the physiological and biochemical processes, as well as the stress signal response [52,53,54]. Therefore, we presumed that SmRAV1 may function in eggplant’s response to salt stress. SmRAV1 expression was not only induced by salt stress, but also up-regulated by ABA and H2O2 treatment, although it was significantly down-regulated by HT, LT, and dehydration treatment (Figure 2), indicating that SmRAV1 may play a negative role in eggplant’s response to HT, LT, and dehydration stresses. These results are similar to those of the previous reports. For example, NtRAV4 played a negative role in Nicotiana tabacum against drought stress by enhancing its antioxidant capacity and defense system [55]. In Arabidopsis, RAV1 expression was reduced by dryness, and its overexpression enhanced transpiration in drought conditions, thus enhancing sensitivity [21]. Moreover, we detected the expression levels of SmRAV1 in diverse tissues from eggplant plants in different developmental stages. High expression was found in the roots of seedlings as well as in the SE and FL of mature plants (Figure 3), suggesting that SmRAV1 may be involved in the process of flower development or the growth and defense of roots. SmRAV1 proteins may localize in the nucleus by predicting its subcellular localization and NLS sequences (Figure S2). We further verified that SmRAV1 proteins localize in the nuclei of epidermic cells of Nicotiana benthamiana leaves by subcellular localization assay (Figure 4b).
To investigate the function of SmRAV1 in eggplant’s response to salt stress, we carried out a VIGS assay to assess the effect of SmRAV1 silencing and the tolerance of eggplant against salt stress. Our data demonstrated that the silencing of SmRAV1 decreased the tolerance of eggplant to salt stress (Figure 5b) and markedly down-regulated the expression level of salt stress defense related marker genes (SmGSTU10 and SmNCED1) (Figure 5d), which was accompanied by a significant decrease in CAT enzyme activity and an increase in H2O2 content (Figure 5e). It can be suggested that SmRAV1 positively functions in eggplant against salt stress. Similar to our findings, overexpression of MtRAV3 was found to enhance the tolerance of Medicago truncatula against salt and osmotic stresses [50]. Wheat RAV1 overexpression enhanced the tolerance of Arabidopsis to salt stress [30]. In Betula platyphylla, BpRAV1 could activate the expression of the superoxide dismutase (SOD) and peroxidase (POD) genes by directly binding to the RAV1A and RBS1 elements to enhance the reactive-oxygen-species-scavenging abilities of SOD and POD, thus improving salt and osmotic stress tolerance [26]. A recent study showed that CsRAV1 overexpression enhanced the tolerance of transgenic Arabidopsis and cucumber seedlings to salt stress and ABA [44]. Interestingly, few reports have indicated that RAV transcription factors acted as negative regulators of salt stress. For example, Arabidopsis RAV1, RAV1L and RAV2/TEM2 negatively regulated its tolerance to salt and drought stress [21]. The overexpression of A-TsRAVs, RAV proteins divided into group A according to their sequence similarity in Thellungiella salsuginea, showed weak growth retardation under salt stress conditions, suggesting that all A-TsRAVs play a negative role in Arabidopsis against salt stress [56]. It is worth mentioning that, apart from the function of RAV transcription factors in the fine-tuning of plant responses to abiotic stress, RAV proteins seem to be negative regulators in plant growth and development [18,21,56,57].
In a word, our data indicate that SmRAV1 positively functions in eggplant’s response to salt stress and lays a foundation for the genetic enhancement of the salinity tolerance of eggplant.
4. Materials and Methods
4.1. Plant Material Growth and Conditions
The growth conditions of eggplant and Nicotiana benthamiana plants followed our previous studies [39,58]. Briefly, seeds of the inbred eggplant line ML41 were packaged with clean gauze, then soaked in a 55 °C water bath for 15 min and left in tap water at room temperature overnight. The seeds were placed in an illumination incubator under the condition of 25 °C for germination. The seedlings with two cotyledons were planted in small plastic pots with nutrient soil, then placed into an illumination incubator under the conditions of 25 °C, a 16 h light/8 h dark photoperiod, and 60% relative humidity. Seeds of Nicotiana benthamiana were sowed on wet filter paper and placed in the illumination incubator at 25 °C for germination. The seedlings of Nicotiana benthamiana were planted in small plastic pots with nutrient soil, then grown under the same conditions as eggplant.
4.2. Abiotic Stresses Treatment
For the treatment of HT and LT, the 4–6-leaf-stage eggplant plants were placed in the illumination incubator under the conditions of 43 °C and 4 °C, respectively. The treated leaves were harvested under the liquid nitrogen condition at the time points of 0, 0.5, 1, 3, 6, and 12 h. The eggplant plants were gently pulled from the nutrient soil, and then the roots were washed with tap water. The washed plants were cultivated in Hoagland nutrient solution for two days. For NaCl, ABA, and H2O2 treatment, the roots were soaked in 200 mM NaCl, 100 μM ABA, and 1 mM H2O2 solution, respectively. The roots were harvested under the condition of liquid nitrogen at the time points of 0, 2, 6, 12, 24, and 48 h. For dehydration stress treatment, the surfaces of the eggplant roots were wiped and placed on the experiment table for continuous dehydration. The roots were harvested under liquid nitrogen at the time points of 0, 0.5, 1, 3, 6, and 9 h.
4.3. Plant Total RNA Extraction, cDNA Synthesis, and RT-qPCR Analysis
The samples were ground into powder under liquid nitrogen conditions, and the total RNA of the eggplant leaves or roots was extracted using the FastPure Plant Total RNA Isolation Kit (polysaccharides- and polyphenolics-rich) kit (RC401-01, Vazyme, Nanjing, China). The purity and concentration of total RNA were measured by an ultra-microspectrophotometer, and the integrity of the total RNA was assessed by an agarose gel electrophoresis assay. For cDNA synthesis, the mRNA from the total RNA was synthesized to cDNA using a HiScript III RT SuperMix for qPCR (+gDNA wiper) kit (R323-01, Vazyme). The transcript expression levels of the target genes were detected by RT-qPCR assay. The RT-qPCR assay was carried out using the ChamQ Universal SYBR qPCR Master Mix kit (Q711-02, Vazyme), according to the specifications. The specific primer pair sequences used to detect the target genes’ expression are listed in Table S1. SmActin (Smechr1100649) was used as a reference gene to standardize the target genes’ expression. Three biological replications and the 2−∆∆CT method were used to analyze the expression of the target genes [59].
4.4. Agrobacterium tumefaciens Cultivation and Infiltration
Agrobacterium tumefaciens strain GV3101 cells, harboring 35S: GFP-SmRAV1 or 35S: GFP structures, were cultivated in liquid Luria–Bertani medium containing 50 μg/mL kanamycin and 50 μg/mL rifampicin in a shaker at 200 rpm and 28 °C overnight. The GV3101 cells were harvested by centrifuging at 28 °C and 6000 rpm for 5 min. The cells were resuspended in infiltration buffer (10 mM MES, 10 mM MgCl2, 200 mM acetosyringone, pH = 5.4) and the OD600 was adjusted to 0.8. The bacterial solution was incubated in the shaker at 28 °C and 60 rpm for 1~3 h, and then injected into the leaves using a disposable sterile syringe without a needle.
4.5. Subcellular Localization Assay
The full-length CDS of SmRAV1 was cloned into the restriction endonuclease Sma Ⅰ of plant overexpression vector pBinGFP2 using the ClonExpress II One Step Cloning Kit (C112-01, Vazyme). Agrobacterium tumefaciens strain GV3101 cells harboring 35S:AtH2B-RFP vector were mixed with the GV3101 cells carrying 35S:GFP-SmRAV1 or 35S:GFP structures at a 1:1 ratio, and then gently incubated in the shaker at 28 °C and 60 rpm for 1 h. The mixtures were infiltrated into the leaves of Nicotiana benthamiana. After 48 h, fluorescence signals were observed in the epidermic cells of Nicotiana benthamiana leaves via laser scanning confocal microscope with an excitation wavelength of 488 nm for GFP and an excitation wavelength of 532 nm for RFP.
4.6. VIGS Assay
The VIGS assay was performed following our previous studies [39,58]. The 300 bp specific DNA fragment of SmRAV1 was amplified using specific primer pairs and cloned into the entry vector pDONR207 by a BP reaction, then transferred into the destination vector pTRV2:00 by an LR reaction. The Agrobacterium GV3101 cells containing TRV1 vector were mixed with the cells harboring TRV2:00, TRV2:SmRAV1, or TRV2:SmPDS vectors at a 1:1 ratio, and then slowly incubated under the conditions of 28 °C and 60 rpm for 1 h. The mixtures were injected into the cotyledons of 2–3-week-old eggplant seedlings, and the infiltrated seedlings were placed into the illumination incubator at 20 °C without light for 48 h. After treatment, seedlings grew under the conditions of 25 °C, a 16 h light/8 h dark photoperiod, and a relative humidity of 60 for 3 weeks.
4.7. Measurement of Physiological Indices
5. Conclusions
In this study, we selected a RAV transcription factor family member called SmRAV1 by analyzing the mRNA sequencing data of eggplant roots treated with salt stress, and it was up-regulated by the salt stress. The results of further analysis showed that the transcript expression levels of SmRAV1 were significantly up-regulated by salt, ABA, and H2O2 treatment, but down-regulated under HT, LT, and dehydration stress treatment. By means of a subcellular localization assay, the SmRAV1 protein localized in the nucleus. According to the results of the VIGS assay, we found that silencing of SmRAV1 decreased the tolerance of eggplant to salt stress; significantly down-regulated the transcript expression levels of salt stress defense-related marker genes, including SmGSTU10 and SmNCED1; markedly decreased CAT enzyme-activity; and increased H2O2 content compared to control plants. These results indicate that SmRAV1 positively functions in eggplant’s response to salt stress, provide new insight into the function of RAV transcription factors in plants against salt stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12244174/s1, Figure S1: Analysis of FPKM value of SmRAV1 from mRNA sequencing data of eggplant roots at 0 and 24 h post-salt-stress treatment. Three biological repeats were used to calculate the mean ± standard deviation. Different upper letters indicate significant differences, as determined by Student’s t-test (p < 0.01). Figure S2: Prediction of SmRAV1 subcellular localization and its NLS sequence. (a) Prediction of SmRAV1 subcellular localization by searching the Plant-mPLoc website. (b) Prediction of NLS sequences of SmRAV1 by searching INSP website. Table S1: The primer pairs sequences used in this study. Table S2: Secondary structure of the SmRAV1 protein.
Author Contributions
L.S. and X.Y. designed the research. J.D., B.Y., and L.S. performed the experiments. L.S. and X.Y. analyzed the data. L.S. wrote the manuscript. L.S. and X.Y. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32102385), and the National Key Research and Development Program of China (2019YFD1000300).
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
We are thankful to Rahat Sharif (College of Horticulture and Landscape architecture, Yangzhou University) for revising the language of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hu, Y.; Fricke, W.; Schmidhalter, U. Salinity and the growth of non-halophytic grass leaves: The role of mineral nutrient distribution. Funct. Plant Biol. 2005, 32, 973–985. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrao, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, Physiology, and Molecular Breeding Approaches for Improving Salt Tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef]
- Negrao, S.; Schmockel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Involvement of auxin in growth and stress response of cucumber. Veg. Res. 2022, 2, 13. [Google Scholar] [CrossRef]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Hormonal interactions underlying parthenocarpic fruit formation in horticultural crops. Hortic. Res. 2022, 9, uhab024. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhou, H.; Fan, Y.; Guo, Y.; Zhang, M.; Shabala, S.; Zhao, C.; Lv, C.; Guo, B.; Wang, F.; et al. HvNCX, a prime candidate gene for the novel qualitative locus qS7.1 associated with salinity tolerance in barley. Theor. Appl. Genet. 2023, 136, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Jeridi, M.; Siddiqui, S.; Shah, A.Z.; Ali, S. Genome-wide identification, characterization, and expression analysis of the Chalcone Synthase gene family in Oryza sativa under Abiotic Stresses. Plant Stress 2023, 9, 100201. [Google Scholar] [CrossRef]
- Ahmad, S.; Ali, S.; Shah, A.Z.; Khan, A.; Faria, S. Chalcone synthase (CHS) family genes regulate the growth and response of cucumber (Cucumis sativus L.) to Botrytis cinerea and abiotic stresses. Plant Stress 2023, 8, 100159. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, Q.; Zhang, Z.; Cheng, P.; Song, A.; Zhou, L.; Wang, L.; Chen, S.; Chen, F.; Jiang, J. The RAV transcription factor TEMPRANILLO1 involved in ethylene-mediated delay of chrysanthemum flowering. Plant J. 2023. [Google Scholar] [CrossRef]
- Swaminathan, K.; Peterson, K.; Jack, T. The plant B3 superfamily. Trends Plant Sci. 2008, 13, 647–655. [Google Scholar] [CrossRef]
- Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Marin-Gonzalez, E.; Suarez-Lopez, P.; Pelaz, S. RAV genes: Regulation of floral induction and beyond. Ann. Bot. 2014, 114, 1459–1470. [Google Scholar] [CrossRef]
- Kagaya, Y.; Ohmiya, K.; Hattori, T. RAV1, a novel DNA-binding protein, binds to bipartite recognition sequence through two distinct DNA-binding domains uniquely found in higher plants. Nucleic Acids Res. 1999, 27, 470–478. [Google Scholar] [CrossRef]
- Tavares, E.Q.P.; De Souza, A.P.; Romim, G.H.; Grandis, A.; Plasencia, A.; Gaiarsa, J.W.; Grima-Pettenati, J.; de Setta, N.; Van Sluys, M.A.; Buckeridge, M.S. The control of endopolygalacturonase expression by the sugarcane RAV transcription factor during aerenchyma formation. J. Exp. Bot. 2019, 70, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hao, D.; Chen, L.; Lu, Q.; Zhang, Y.; Li, Y.; Duan, Y.; Li, W. Roles for a soybean RAV-like orthologue in shoot regeneration and photoperiodicity inferred from transgenic plants. J. Exp. Bot. 2012, 63, 3257–3270. [Google Scholar] [CrossRef]
- Feng, C.Z.; Chen, Y.; Wang, C.; Kong, Y.H.; Wu, W.H.; Chen, Y.F. Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development. Plant J. 2014, 80, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zhao, L.; Li, D.; Hao, D.; Zhan, Y.; Li, W. A GmRAV ortholog is involved in photoperiod and sucrose control of flowering time in soybean. PLoS ONE 2014, 9, e89145. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, M.; Zhang, L.; Jiang, Y.; Zhao, C.; Shahid, M.Q.; Bai, Y.; Hao, J.; Peng, J.; Gao, Y.; et al. EjRAV1/2 Delay Flowering through Transcriptional Repression of EjFTs and EjSOC1s in Loquat. Front. Plant Sci. 2021, 12, 816086. [Google Scholar] [CrossRef]
- Zhang, K.; Zhao, L.; Yang, X.; Li, M.; Sun, J.; Wang, K.; Li, Y.; Zheng, Y.; Yao, Y.; Li, W. GmRAV1 regulates regeneration of roots and adventitious buds by the cytokinin signaling pathway in Arabidopsis and soybean. Physiol. Plant. 2019, 165, 814–829. [Google Scholar] [CrossRef]
- Fu, M.; Kang, H.K.; Son, S.H.; Kim, S.K.; Nam, K.H. A subset of Arabidopsis RAV transcription factors modulates drought and salt stress responses independent of ABA. Plant Cell Physiol. 2014, 55, 1892–1904. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yan, Y.; Lu, Y.; Liu, G.; Liu, J.; Shi, H. The co-modulation of RAV transcription factors in ROS burst and extensive transcriptional reprogramming underlies disease resistance in cassava. Plant Cell Rep. 2022, 41, 1261–1272. [Google Scholar] [CrossRef]
- Wei, Y.; Chang, Y.; Zeng, H.; Liu, G.; He, C.; Shi, H. RAV transcription factors are essential for disease resistance against cassava bacterial blight via activation of melatonin biosynthesis genes. J. Pineal Res. 2018, 64, e12454. [Google Scholar] [CrossRef] [PubMed]
- Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya; You, S.J.; Hsieh, T.H.; Chao, T.C.; Chan, M.T. Tomato RAV transcription factor is a pivotal modulator involved in the AP2/EREBP-mediated defense pathway. Plant Physiol. 2011, 156, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Sohn, K.H.; Lee, S.C.; Jung, H.W.; Hong, J.K.; Hwang, B.K. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol. Biol. 2006, 61, 897–915. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhang, K.; Yang, C. Functional roles of the birch BpRAV1 transcription factor in salt and osmotic stress response. Plant Sci. 2022, 315, 111131. [Google Scholar] [CrossRef]
- Karami, M.; Fatahi, N.; Lohrasebi, T.; Razavi, K. RAV transcription factor regulatory function in response to salt stress in two Iranian wheat landraces. J. Plant Res. 2022, 135, 121–136. [Google Scholar] [CrossRef]
- Osnato, M.; Cereijo, U.; Sala, J.; Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Rodriguez-Goberna, M.R.; Riechmann, J.L.; Rodriguez-Concepcion, M.; Pelaz, S. The floral repressors TEMPRANILLO1 and 2 modulate salt tolerance by regulating hormonal components and photo-protection in Arabidopsis. Plant J. 2021, 105, 7–21. [Google Scholar] [CrossRef]
- Gao, Y.; Han, D.; Jia, W.; Ma, X.; Yang, Y.; Xu, Z. Molecular characterization and systematic analysis of NtAP2/ERF in tobacco and functional determination of NtRAV-4 under drought stress. Plant Physiol. Biochem. 2020, 156, 420–435. [Google Scholar] [CrossRef]
- Luo, Y.X.; Chen, S.K.; Wang, P.D.; Peng, D.; Zhang, X.; Li, H.F.; Feng, C.Z. Genome-Wide Analysis of the RAV Gene Family in Wheat and Functional Identification of TaRAV1 in Salt Stress. Int. J. Mol. Sci. 2022, 23, 8834. [Google Scholar] [CrossRef]
- Kabir, N.; Lin, H.; Kong, X.; Liu, L.; Qanmber, G.; Wang, Y.; Zhang, L.; Sun, Z.; Yang, Z.; Yu, Y.; et al. Identification, evolutionary analysis and functional diversification of RAV gene family in cotton (G. hirsutum L.). Planta 2021, 255, 14. [Google Scholar] [CrossRef]
- Liu, J.; Deng, Z.; Liang, C.; Sun, H.; Li, D.; Song, J.; Zhang, S.; Wang, R. Genome-Wide Analysis of RAV Transcription Factors and Functional Characterization of Anthocyanin-Biosynthesis-Related RAV Genes in Pear. Int. J. Mol. Sci. 2021, 22, 5567. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, Y.; Zhang, H.; Ma, Q.; Wei, Z.; Chen, J.; Sun, Z. Genome-Wide Analysis of the RAV Transcription Factor Genes in Rice Reveals Their Response Patterns to Hormones and Virus Infection. Viruses 2021, 13, 752. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.P.; Xu, Z.S.; Zheng, W.J.; Zhao, W.; Wang, Y.X.; Yu, T.F.; Chen, M.; Zhou, Y.B.; Min, D.H.; Ma, Y.Z.; et al. Genome-Wide Analysis of the RAV Family in Soybean and Functional Identification of GmRAV-03 Involvement in Salt and Drought Stresses and Exogenous ABA Treatment. Front. Plant Sci. 2017, 8, 905. [Google Scholar] [CrossRef]
- Zhang, C.H.; Shangguan, L.F.; Ma, R.J.; Sun, X.; Tao, R.; Guo, L.; Korir, N.K.; Yu, M.L. Genome-wide analysis of the AP2/ERF superfamily in peach (Prunus persica). Genet. Mol. Res. 2012, 11, 4789–4809. [Google Scholar] [CrossRef] [PubMed]
- Castillejo, C.; Pelaz, S. The balance between CONSTANS and TEMPRANILLO activities determines FT expression to trigger flowering. Curr. Biol. 2008, 18, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Li, H.; Wang, J.; Zeng, Q.; Huang, W.; Huang, H.; Xie, Y.; Yu, S.; Kan, Q.; et al. GLRaV-2 protein p24 suppresses host defenses by interaction with a RAV transcription factor from grapevine. Plant Physiol. 2022, 189, 1848–1865. [Google Scholar] [CrossRef] [PubMed]
- Yarra, R.; Kirti, P.B. Expressing class I wheat NHX (TaNHX2) gene in eggplant (Solanum melongena L.) improves plant performance under saline condition. Funct. Integr. Genom. 2019, 19, 541–554. [Google Scholar] [CrossRef]
- Shen, L.; Zhao, E.; Liu, R.; Yang, X. Transcriptome Analysis of Eggplant under Salt Stress: AP2/ERF Transcription Factor SmERF1 Acts as a Positive Regulator of Salt Stress. Plants 2022, 11, 2205. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Guo, Y.; Yang, Y.; Huang, Y.; Shen, H.B. Discovering nuclear targeting signal sequence through protein language learning and multivariate analysis. Anal. Biochem. 2020, 591, 113565. [Google Scholar] [CrossRef] [PubMed]
- Launholt, D.; Merkle, T.; Houben, A.; Schulz, A.; Grasser, K.D. Arabidopsis chromatin-associated HMGA and HMGB use different nuclear targeting signals and display highly dynamic localization within the nucleus. Plant Cell 2006, 18, 2904–2918. [Google Scholar] [CrossRef]
- Boisnard-Lorig, C.; Colon-Carmona, A.; Bauch, M.; Hodge, S.; Doerner, P.; Bancharel, E.; Dumas, C.; Haseloff, J.; Berger, F. Dynamic analyses of the expression of the HISTONE::YFP fusion protein in arabidopsis show that syncytial endosperm is divided in mitotic domains. Plant Cell 2001, 13, 495–509. [Google Scholar] [CrossRef]
- Li, J.; Song, C.; Li, H.; Wang, S.; Hu, L.; Yin, Y.; Wang, Z.; He, W. Comprehensive analysis of cucumber RAV family genes and functional characterization of CsRAV1 in salt and ABA tolerance in cucumber. Front. Plant Sci. 2023, 14, 1115874. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, U.; Doll, J. Arabidopsis WRKY53, a Node of Multi-Layer Regulation in the Network of Senescence. Plants 2019, 8, 578. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, U.; Laun, T.; Miao, Y. The complex regulation of WRKY53 during leaf senescence of Arabidopsis thaliana. Eur. J. Cell Biol. 2010, 89, 133–137. [Google Scholar] [CrossRef]
- Wang, X.; Chen, K.; Zhou, M.; Gao, Y.; Huang, H.; Liu, C.; Fan, Y.; Fan, Z.; Wang, Y.; Li, X. GmNAC181 promotes symbiotic nodulation and salt tolerance of nodulation by directly regulating GmNINa expression in soybean. New Phytol. 2022, 236, 656–670. [Google Scholar] [CrossRef]
- Segarra, G.; Van der Ent, S.; Trillas, I.; Pieterse, C.M. MYB72, a node of convergence in induced systemic resistance triggered by a fungal and a bacterial beneficial microbe. Plant Biol. 2009, 11, 90–96. [Google Scholar] [CrossRef]
- Son, G.H.; Wan, J.; Kim, H.J.; Nguyen, X.C.; Chung, W.S.; Hong, J.C.; Stacey, G. Ethylene-responsive element-binding factor 5, ERF5, is involved in chitin-induced innate immunity response. Mol. Plant Microbe Interact. 2012, 25, 48–60. [Google Scholar] [CrossRef]
- Wang, S.; Guo, T.; Shen, Y.; Wang, Z.; Kang, J.; Zhang, J.; Yi, F.; Yang, Q.; Long, R. Overexpression of MtRAV3 enhances osmotic and salt tolerance and inhibits growth of Medicago truncatula. Plant Physiol. Biochem. 2021, 163, 154–165. [Google Scholar] [CrossRef]
- Li, X.J.; Li, M.; Zhou, Y.; Hu, S.; Hu, R.; Chen, Y.; Li, X.B. Overexpression of cotton RAV1 gene in Arabidopsis confers transgenic plants high salinity and drought sensitivity. PLoS ONE 2015, 10, e0118056. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Fu, J.; Shen, Q.; Wang, Q. OsWRKY10 extensively activates multiple rice diterpenoid phytoalexin biosynthesis genes to enhance rice blast resistance. Plant J. 2023, 115, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Sun, B.; Cai, W.; Zhou, X.; Mao, Y.; Chen, C.; Wei, J.; Cao, B.; Chen, C.; Chen, G.; et al. Natural variations in the MYB transcription factor MYB31 determine the evolution of extremely pungent peppers. New Phytol. 2019, 223, 922–938. [Google Scholar] [CrossRef]
- Guo, Q.; Major, I.T.; Kapali, G.; Howe, G.A. MYC transcription factors coordinate tryptophan-dependent defence responses and compromise seed yield in Arabidopsis. New Phytol. 2022, 236, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, J.; Duan, W.; Ma, X.; Qu, L.; Xu, Z.; Yang, Y.; Xu, J. NtRAV4 negatively regulates drought tolerance in Nicotiana tabacum by enhancing antioxidant capacity and defence system. Plant Cell Rep. 2022, 41, 1775–1788. [Google Scholar] [CrossRef]
- Yang, S.; Luo, C.; Song, Y.; Wang, J. Two Groups of Thellungiella salsuginea RAVs Exhibit Distinct Responses and Sensitivity to Salt and ABA in Transgenic Arabidopsis. PLoS ONE 2016, 11, e0153517. [Google Scholar] [CrossRef]
- Zhao, L.; Luo, Q.; Yang, C.; Han, Y.; Li, W. A RAV-like transcription factor controls photosynthesis and senescence in soybean. Planta 2008, 227, 1389–1399. [Google Scholar] [CrossRef]
- Shen, L.; He, J.; Yang, X. Genome-wide identification of calmodulin-binding protein 60 gene family and function of SmCBP60A1 in eggplant response to salt stress. Sci. Hortic. 2023, 322, 112448. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Figure 1.
Sequence analysis of SmRAV1. (a) Schematic diagram of conserved domains of SmRAV1. (b) Analysis of secondary structure of SmRAV1. The blue, red, green, and azalein lines, respectively, represent secondary structure types of helix, sheet, turn, and coil. (c) Prediction of tertiary structure model of SmRAV1 protein. (d) Analysis of multiple alignment analysis of RAV proteins’ amino acid sequences from Solanum melongena, Solanum dulcamara, Solanum tuberosum, Solanum stenotomum, Solanum verrucosum, Solanum lycopersicum, Nicotiana tabacum, Vitis vinifera, Capsicum annuum, Zea mays, Arabidopsis thaliana, and Oryza sativa. The black, azalein, and blue shadows, respectively, represent 100%, 75~100%, and 50~75% sequence similarity. (e) Phylogenetic relationship analysis of SmRAV1 with its homologs from selected plant species. (f) Prediction of cis-elements within the assumed promoter sequences of SmRAV1.
Figure 1.
Sequence analysis of SmRAV1. (a) Schematic diagram of conserved domains of SmRAV1. (b) Analysis of secondary structure of SmRAV1. The blue, red, green, and azalein lines, respectively, represent secondary structure types of helix, sheet, turn, and coil. (c) Prediction of tertiary structure model of SmRAV1 protein. (d) Analysis of multiple alignment analysis of RAV proteins’ amino acid sequences from Solanum melongena, Solanum dulcamara, Solanum tuberosum, Solanum stenotomum, Solanum verrucosum, Solanum lycopersicum, Nicotiana tabacum, Vitis vinifera, Capsicum annuum, Zea mays, Arabidopsis thaliana, and Oryza sativa. The black, azalein, and blue shadows, respectively, represent 100%, 75~100%, and 50~75% sequence similarity. (e) Phylogenetic relationship analysis of SmRAV1 with its homologs from selected plant species. (f) Prediction of cis-elements within the assumed promoter sequences of SmRAV1.

Figure 2.
Analysis of transcript expression profiles of SmRAV1 in eggplant leaves or roots under HT, LT, dehydration, NaCl, H2O2, and ABA treatment. Three biological repeats were used to calculate the mean ± standard deviation. Different upper letters indicate significant differences, as analyzed by Fisher’s protected LSD test (p < 0.01).
Figure 2.
Analysis of transcript expression profiles of SmRAV1 in eggplant leaves or roots under HT, LT, dehydration, NaCl, H2O2, and ABA treatment. Three biological repeats were used to calculate the mean ± standard deviation. Different upper letters indicate significant differences, as analyzed by Fisher’s protected LSD test (p < 0.01).

Figure 3.
Analysis of tissue-specific expression of SmRAV1. Three biological repeats were used to calculate the mean ± standard deviation.
Figure 3.
Analysis of tissue-specific expression of SmRAV1. Three biological repeats were used to calculate the mean ± standard deviation.
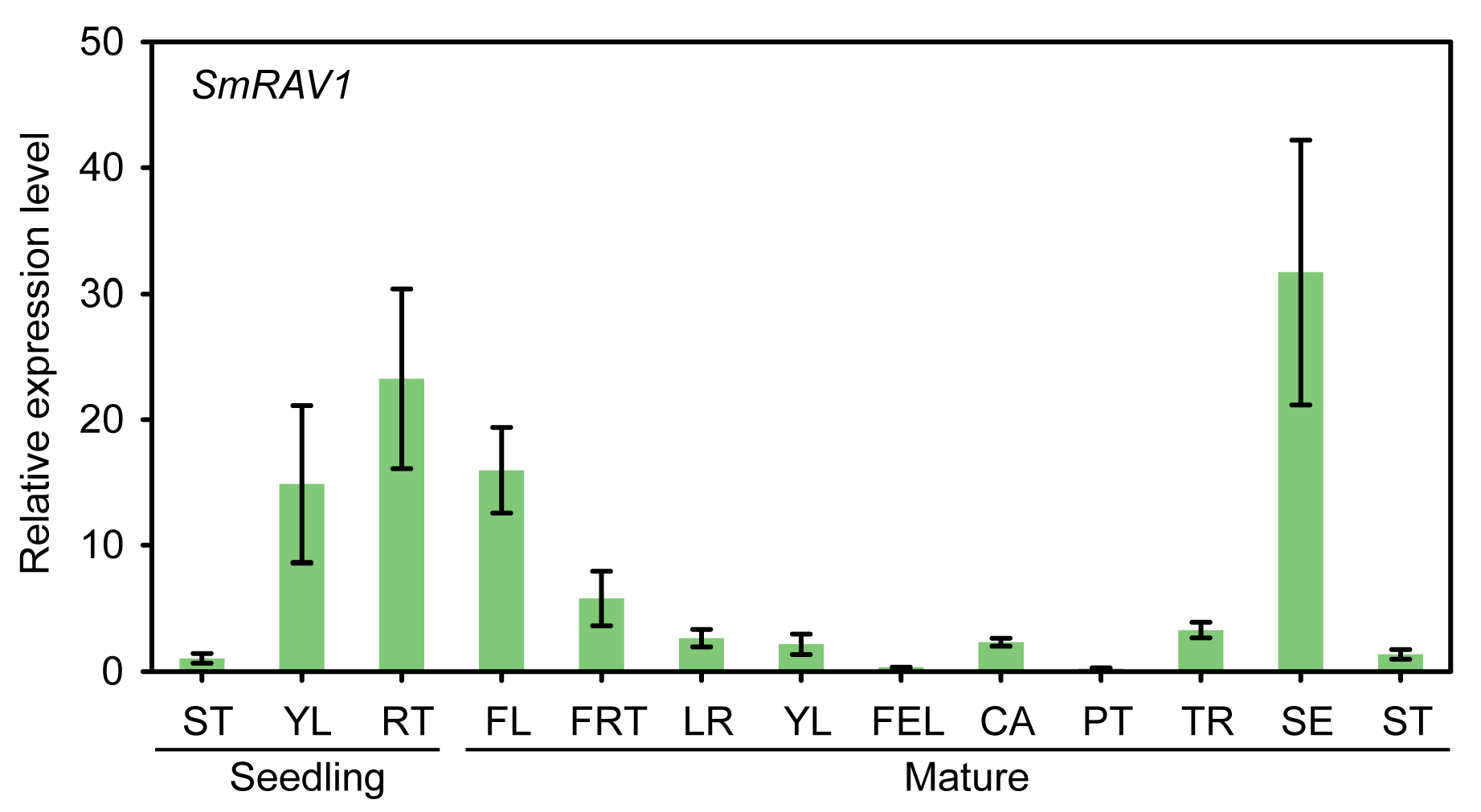
Figure 4.
Subcellular localization of SmRAV1. (a) Schematic diagram of 35S:GFP, 35S:GFP-SmRAV1, and 35S:AtH2B-RFP structures. (b) Analysis of subcellular localization of SmRAV1 in the epidermic cells of Nicotiana benthamiana leaves. Bar = 25 μm.
Figure 4.
Subcellular localization of SmRAV1. (a) Schematic diagram of 35S:GFP, 35S:GFP-SmRAV1, and 35S:AtH2B-RFP structures. (b) Analysis of subcellular localization of SmRAV1 in the epidermic cells of Nicotiana benthamiana leaves. Bar = 25 μm.

Figure 5.
Silencing of SmRAV1 enhances susceptibility of eggplant’s response to salt stress. (a) Detection of silencing efficiency of SmRAV1. (b) SmRAV1-silenced eggplant plants exhibited more sensitivity under salt stress treatment at 36 h post-treatment. (c) Analysis of survival rate of SmRAV1-silenced and control plants under salt stress treatment at 36 h post treatment. (d) Analysis of transcript expression levels of salt-stress-defense-related marker genes, including SmGSTU10 and SmNCED1, in the roots of SmRAV1-silenced and control eggplants at 24 h post-salt-stress treatment. (e) Measurement of CAT enzyme activity and H2O2 content in the roots of SmRAV1-silenced and control eggplants at 48 h post-salt-stress treatment. In (a,c–e), three biological repeats were used to calculate the mean ± standard deviation. Different upper letters indicate significant differences, as determined by Student’s t-test (p < 0.01).
Figure 5.
Silencing of SmRAV1 enhances susceptibility of eggplant’s response to salt stress. (a) Detection of silencing efficiency of SmRAV1. (b) SmRAV1-silenced eggplant plants exhibited more sensitivity under salt stress treatment at 36 h post-treatment. (c) Analysis of survival rate of SmRAV1-silenced and control plants under salt stress treatment at 36 h post treatment. (d) Analysis of transcript expression levels of salt-stress-defense-related marker genes, including SmGSTU10 and SmNCED1, in the roots of SmRAV1-silenced and control eggplants at 24 h post-salt-stress treatment. (e) Measurement of CAT enzyme activity and H2O2 content in the roots of SmRAV1-silenced and control eggplants at 48 h post-salt-stress treatment. In (a,c–e), three biological repeats were used to calculate the mean ± standard deviation. Different upper letters indicate significant differences, as determined by Student’s t-test (p < 0.01).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ding, J.; Yao, B.; Yang, X.; Shen, L. SmRAV1, an AP2 and B3 Transcription Factor, Positively Regulates Eggplant’s Response to Salt Stress. Plants 2023, 12, 4174. https://doi.org/10.3390/plants12244174
AMA Style
Ding J, Yao B, Yang X, Shen L. SmRAV1, an AP2 and B3 Transcription Factor, Positively Regulates Eggplant’s Response to Salt Stress. Plants. 2023; 12(24):4174. https://doi.org/10.3390/plants12244174
Chicago/Turabian StyleDing, Junjie, Bowen Yao, Xu Yang, and Lei Shen. 2023. "SmRAV1, an AP2 and B3 Transcription Factor, Positively Regulates Eggplant’s Response to Salt Stress" Plants 12, no. 24: 4174. https://doi.org/10.3390/plants12244174
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.