Tungsten Toxicity in Plants


Abstract
:Abbreviations
| AO | aldehyde oxidase |
| ER | endoplasmic reticulum |
| mARC | mitochondrial amidoxime reducing component |
| MoCo | molybdenum cofactor |
| Mo-enzymes | molybdoenzymes |
| MT | microtubule |
| NR | nitrate reductase |
| PCD | programmed cell death |
| ROS | reactive oxygen species |
| SO | sulphite oxidase |
| UPR | unfolded protein response |
| W-PCD | tungsten-induced programmed cell death |
| XDH | xanthine dehydrogenase |
1. Introduction
2. W Effects on Plants
2.1. W Uptake and Accumulation
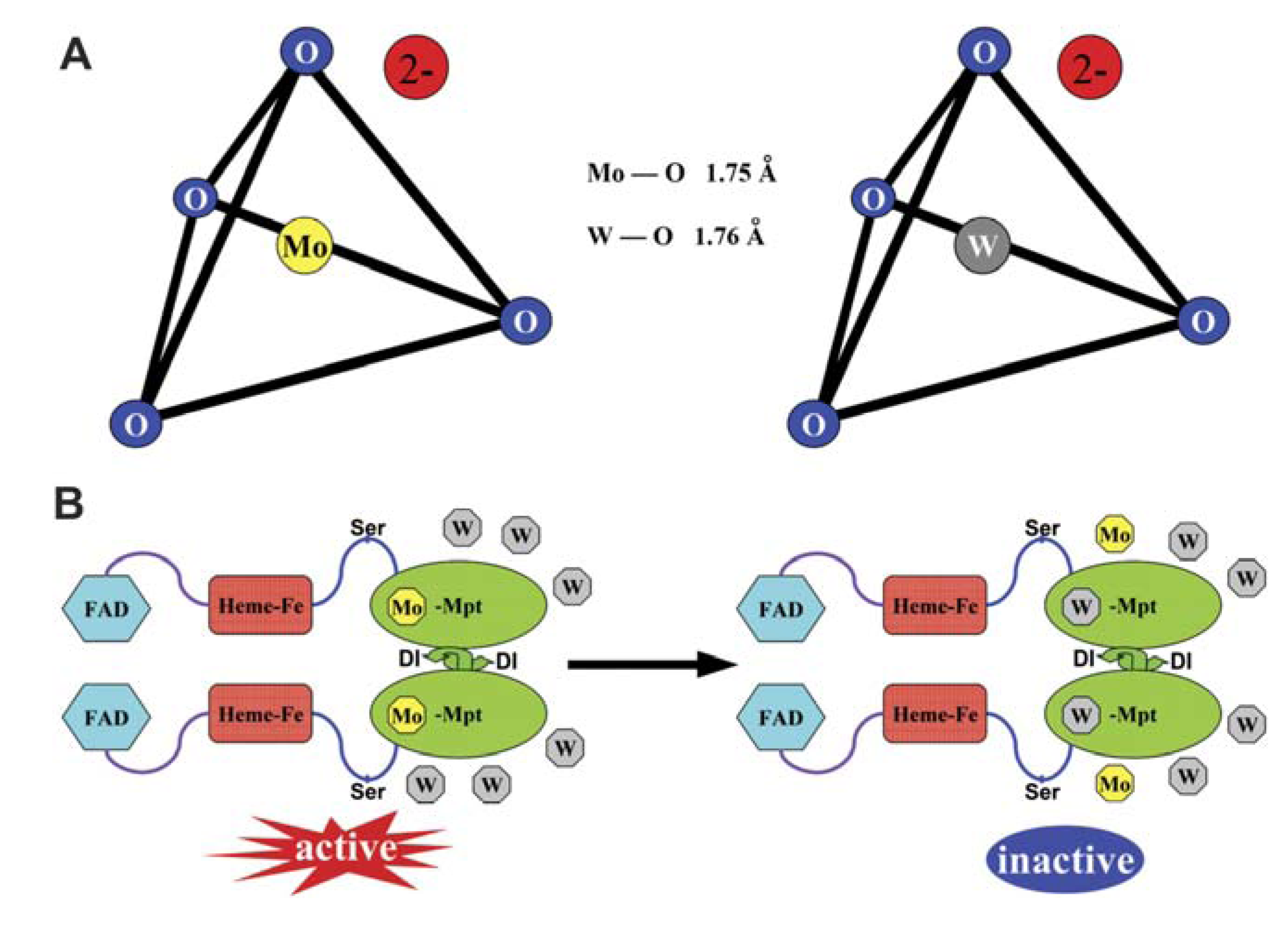
2.2. W-Mo Antagonism: W as an Inhibitor of Mo-Enzyme Activity
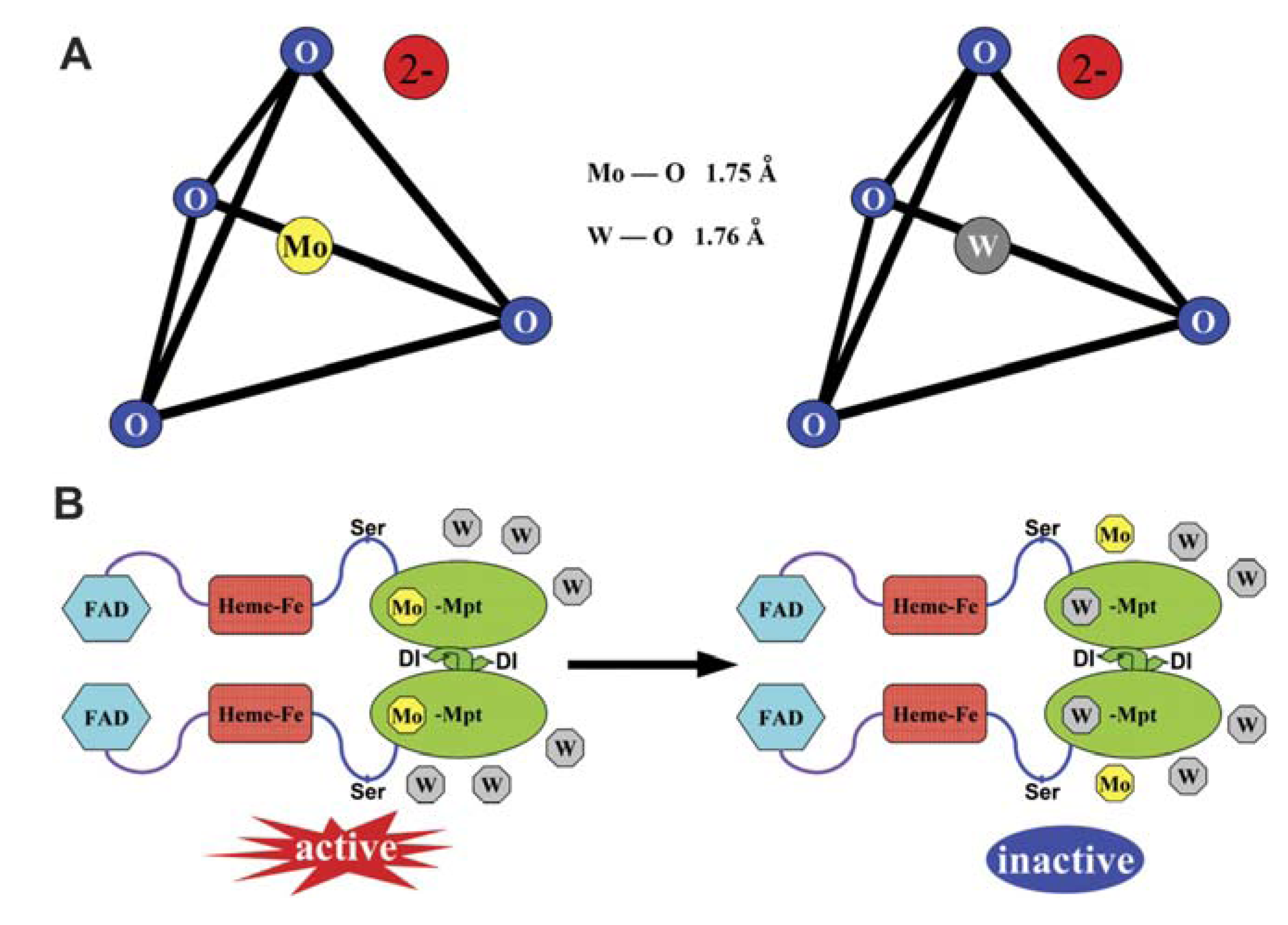
2.3. W Is More Than a Mo-Enzyme Inhibitor
2.3.1. W Inhibits Plant Growth

{kind=link}
{kind=link}
| Plant species | Treatments | Effects/Results | Reference |
|---|---|---|---|
| Brassica rapa | Seeds sown in Magenta boxes on ½ MS medium with 50-150 mg L−1 W as Na2WO4. | Increased production of a blue substance correlated with anthocyanin accumulation. | [36] |
| B. juncea | |||
| B. oleracea | |||
| Hordeum vulgare L. cv. Steptoe (barley) | 0.25, 0.5, 1, 10, 100, 500 μM Na2WO4, 9 days. Hydroponics. | Significant reduction in root and shoot biomass at high concentrations (≥10 μM). | [44] |
| Lolium perenne (ryegrass) | Urban and forest soil amended with ammunition grade W powder (average particle size 5 μM): 1, 10, 100, 1,000, 10,000 mg kg−1. Up to 9 months. | Serious reduction of plant growth and death after two months. | [19] |
| Pisum sativum cv. Onmard (pea) | 200, 500 mg L−1 Na2WO4, up to 8 days. Hydroponics. | Inhibition of root elongation and lateral root formation, retardment of seedling growth rate and new leaf emergence. | [25] |
| Gossypium hisutum cv. | |||
| Campo (cotton) | |||
| Helianthus annuus L. (sunflower) | Processed field soil, spiked with 6,500 mg kg−1 metallic W powder, aged for six months. Diluted soil samples contained from 0 to 6,500 mg W kg−1. Plants grown for two or four weeks. | Plant total weight significantly reduced at W concentration ≥2,600 mg kg−1, root and shoot length reduced at ≥3,900 mg kg−1. | [34] |
| Avena sativa cv. Ogle (oat) | Natural soil containing 0 (control), 0.803, 2.41, 7.21, 21.7, 65.0, 195.1, and 586 mg W kg−1 dry soil. | Toxicity was recorded at ~58.6-293 mg W kg−1 dry soil, with lettuce being more sensitive than radish, followed by oat. | [53] |
| Raphanus sativus cv. Crimson Giant | |||
| (radish) | |||
| Lactuca sativa cv. Grand Rapids (lettuce) | |||
| Triticum aestivum L. var Raj4037. | Potted soil watered with 3, 9, 27, 81, 243 mg kg−1 Na2WO4.2H2O solutions plus control. Samples harvested after 60 days. | Lower concentrations (3, 9 mg kg−1) of W had promotive effects in growth, biomass, chlorophyll, carbohydrate and Mo contents, higher ones decreased them. | [35] |
| Vigna unguiculata L. Walp. var. Sephali Shikha-313 | Potted soil watered with 5, 10, 15, 20, 25 μg g−1 Na2WO4.2H2O solutions. Samples harvested after fruiting. | Lower applied doses (5, 10, 15 μg g−1) of W promoted root-shoot length. Higher doses retarded root-shoot length. | [54] |
| Brassica oleracea (cabbage) | Grown in aged W powder-spiked soil containing monomeric and polymeric tungstates provided as Na2WO4·2H2O. | Cabbage growth was impaired at 436 mg W kg−1 W soil. | [23] |
| Lactuca sativa (lettuce) |
2.3.2. W-Induced Programmed Cell Death (PCD)
| Metal | Plant species | Conditions and responses | Reference |
|---|---|---|---|
| Cd | Nicotiana tabacum Mill. | Chronic exposure of suspension cells to 50-100 mM CdSO4 induced apoptotic-like PCD, including DNA fragmentation into oligonucleosomal units (50-200 kb fragments). | [72] |
| BY-2 cell line | |||
| Al | Hordeum vulgare | In root-tip cells 0.1-1.0 mM Al treatments for 8 h induced PCD, possibly via a ROS-modified signal transduction pathway, whereas 10-50 mM Al treatments caused necrosis. | [73] |
| Cd | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | Cadmium (CdSO4) induced apoptotic-like PCD that required increased H2O2 production and activation of phospholipase C and D and ethylene signalling pathways. | [74] |
| Al | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | Suspension cells treated with 100 μM AlCl3 showed typical features of PCD (nuclear and cytoplasmic condensation), executed by caspase-like proteases. | [75] |
| Cd | Lycopersicon esculentum Mill. (tomato). Cell line MsK8 | 0.1 mM CdSO4 in cell suspension culture induced cell death after 24 h, involving caspase-like proteases, indicating that PCD took place. | [76] |
| Cd | Nicotiana tabacum L. (tobacco) BY-2 cell line | 50 μM CdSO4 induced internucleosomal DNA fragmentation connected with the action of cysteine proteases and the loss of membrane integrity, in particular of tonoplast. | [70] |
| W | Pisum sativum L. cv. Onmard (pea) | Root tip cells of young seedlings exposed to 200 mg L−1 Na2WO4 for 12-72 h executed PCD through ER stress-UPR. The expression of the PCD-related genes DAD-1 and HSR203J was altered. | [28] |
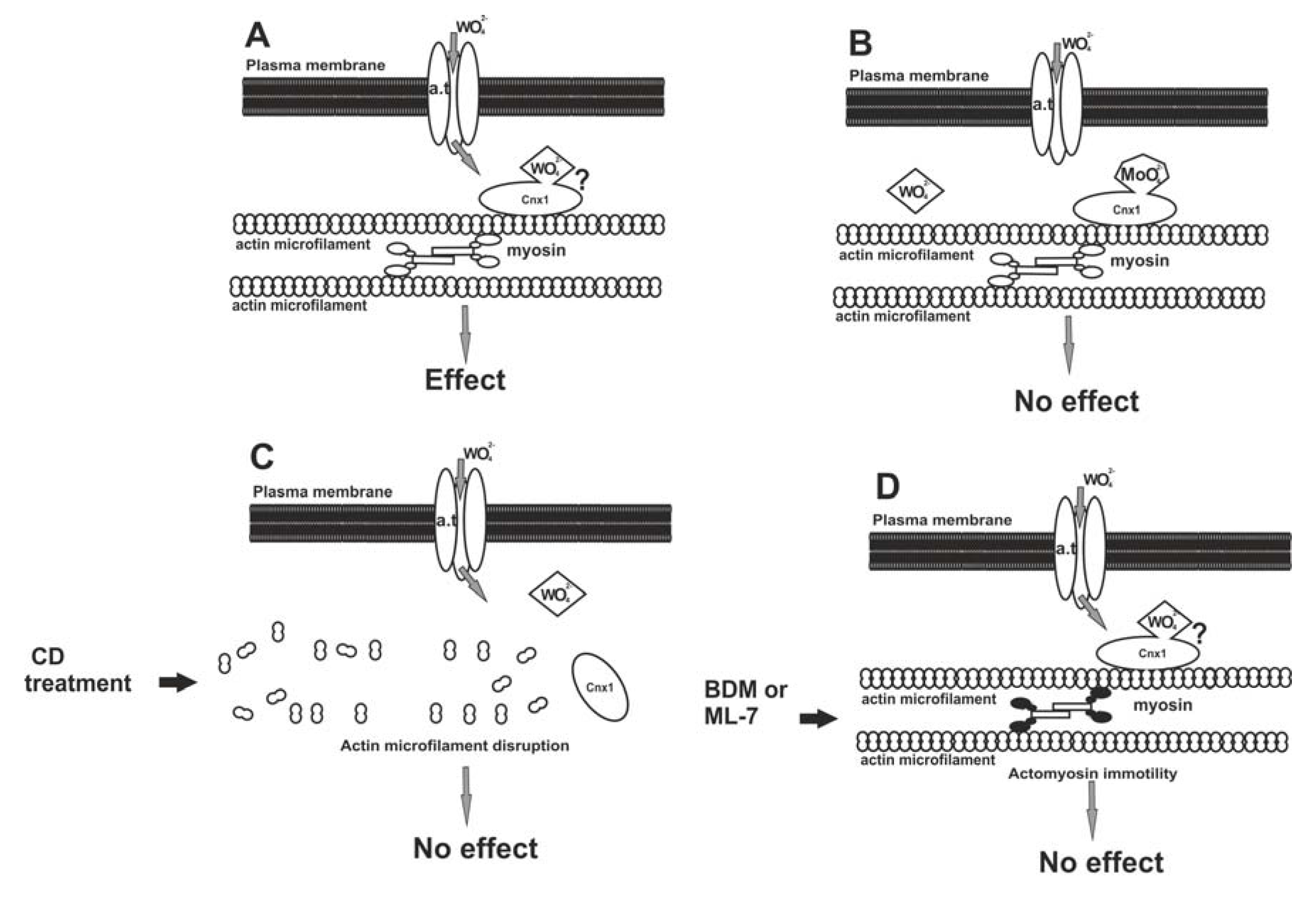
2.4. A Model for W Entrance in Plant Cells

3. Conclusions and Perspectives
Acknowledgements
References
- Koutsospyros, A.; Braida, W.; Christodoulatos, C.; Dermatas, D.; Strigul, N. A review of tungsten: From environmental obscurity to scrutiny. J. Hazard. Mater. 2006, 136, 1–19. [Google Scholar] [CrossRef]
- Pyatt, F.B.; Pyatt, A.J. The bioaccumulation of tungsten and copper by organisms inhabiting metalliferous areas in North Queensland, Australia: An evaluation of potential health implications. J. Environ. Health Res. 2004, 3, 13–18. [Google Scholar]
- Pratas, J.; Prasad, M.N.V.; Freitas, H.; Conde, L. Plants growing in abandoned mines of Portugal are useful for biogeochemical exploration of arsenic, antimony, tungsten and mine reclamation. J. Geochem. Explor. 2005, 85, 99–107. [Google Scholar] [CrossRef]
- Wilson, B.; Pyatt, F.B. Bio-availability of tungsten in the vicinity of an abandoned mine in the English Lake District and some potential health implications. Sci. Total Environ. 2006, 370, 401–408. [Google Scholar] [CrossRef]
- Senesi, N.; Padovaro, G.; Brunetti, G. Scandium, titanium, tungsten and zirconium content in commercial inorganic fertilizers and their contribution to soil. Environ. Techn. Lett. 1988, 9, 1011–1020. [Google Scholar] [CrossRef]
- Clausen, J.L.; Korte, N. Environmental fate of tungsten from military use. Sci. Total Environ. 2009, 407, 2887–2893. [Google Scholar]
- Karachalios, A.; Wazne, M.; Betancur, J.N.; Christodoulatos, C.; Braida, W.; O’Connor, G. Immobilization of copper, lead, and tungsten in mixed munitions firing rang. Contaminated soils by various amendments. J. Haz. Toxic Radioact. Waste 2011, 15, 151–159. [Google Scholar] [CrossRef]
- Sadiq, M.; Mian, A.A.; Althagafi, K.M. Inter-city comparison of metals in scalp hair collected after the Gulf War 1991. J. Environ. Sci. Health A 1992, 27, 1415–1431. [Google Scholar]
- Harper, E.M.; Graedel, T.E. Illuminating tungsten’s life cycle in the United States: 1975–2000. Environ. Sci. Technol. 2008, 42, 3835–3842. [Google Scholar]
- Clemente, R.; Nicholas, W.L. Tungsten. In Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability; Alloway, B.J., Ed.; Springer Science and Business Media: Dordrecht, Germany, 2013. [Google Scholar] [CrossRef]
- Thomas, V.G.; Roberts, M.J.; Harrison, P.T. Assessment of the environmental toxicity and carcinogenicity of tungsten-based shot. Ecotoxicol. Environ. Saf. 2009, 72, 1031–1037. [Google Scholar] [CrossRef] [Green Version]
- Kelly, A.D.R.; Lemaire, M.; Young, Y.K.; Eustache, J.H.; Guilbert, C.; Molina, M.F.; Mann, K.K. In vivo tungsten exposure alters B cell development and increases DNA damage in murine bone marrow. Toxicol. Sci. 2012. [Google Scholar] [CrossRef]
- Strigul, N.S.; Koutsospyros, A.; Christodoulatos, C. Tungsten in the former Soviet Union: Review of environmental regulations and related research. Land Contam. Reclam. 2009, 17, 189–215. [Google Scholar] [CrossRef]
- Sheppard, P.R.; Bierman, B.J.; Rhodes, K.; Ridenour, G.; Witten, M.L. Comparison of size and geography of airborne tungsten particles in Fallon, Nevada, and Sweet Home, Oregon, with implications for public health. J. Environ. Public Health 2012. [Google Scholar] [CrossRef]
- Steinmaus, C.; Lu, M.; Todd, R.; Smith, A.H. Probability estimates for the unique childhood leukemia cluster in Fallon, Nevada, and risks near other U.S. military aviation facilities. Environ. Health Perspect. 2004, 112, 766–771. [Google Scholar] [CrossRef]
- CDC (2003). Cross-sectional exposure assessment of environmental contaminants of Churchill County, Nevada. Final Report. Available online: http://www.cdc.gov/nceh/clusters/Fallon/study.htm/ (accessed on 5 November 2012).
- Kletzin, A.; Adams, M.W.W. Tungsten in biological systems. FEMS Microbiol. Rev. 1996, 18, 5–63. [Google Scholar]
- Kalinich, J.F.; Emond, C.A.; Dalton, T.K.; Mog, S.R.; Coleman, G.D.; Kordell, J.E.; Miller, A.C.; McClain, D.E. Embedded weapons-grade tungsten alloy shrapnel rapidly induces metastatic high-grade rhabnomyosarcomas in F344 rats. Environ. Health Perspect. 2005, 113, 729–734. [Google Scholar] [CrossRef]
- Strigul, N.; Koutsospyros, A.; Arienti, P.; Christodoulatos, C.; Dermatas, D.; Braida, W. Effects of tungsten on environmental systems. Chemosphere 2005, 61, 248–258. [Google Scholar]
- Ringelberg, D.B.; Reynolds, C.M.; Winfield, L.E.; Inouye, L.S.; Johnson, D.R.; Bednar, A.J. Tungsten effects on microbial community structure and activity in a soil. J. Environ. Qual. 2009, 38, 103–110. [Google Scholar] [CrossRef]
- Osterburg, A.R.; Robinson, C.T.; Schwemberger, S.; Mokashi, V.; Stockelman, M.; Babcock, G.F. Sodium tungstate (Na2WO4) exposure increases apoptosis in human peripheral blood lymphocytes. J. Immunotoxicol. 2010, 7, 174–182. [Google Scholar] [CrossRef]
- Guilbert, C.; Kelly, A.D.R.; Petruccelli, L.A.; Lemaire, M.; Mann, K.K. Exposure to tungsten induces DNA damage and apoptosis in developing B lymphocytes. Leukemia 2011, 25, 1900–1904. [Google Scholar]
- Kennedy, A.J.; Johnson, D.R.; Seiter, J.M.; Lindsay, J.H.; Boyd, R.E.; Bednar, A.J.; Allison, P.G. Tungsten toxicity, bioaccumulation and compartmentalization into organisms representing two trophic levels. Environ. Sci. Technol. 2012. [Google Scholar] [CrossRef]
- Kühnel, D.; Scheffler, K.; Wellner, P.; Meissner, T.; Potthoff, A.; Busch, W.; Springer, A.; Schirmer, K. Comparative evaluation of particle properties, formation of reactive oxygen species and genotoxic potential of tungsten carbide based nanoparticles in vitro. J. Hazard. Mater. 2012, 227–228, 418–426. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Eleftheriou, E.P.; Rost, T.L. Effects of sodium tungstate on the ultrastructure and growth of pea (Pisum sativum) and cotton (Gossypiumhirsutum) seedlings. Environ. Exp. Bot. 2008, 63, 416–425. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten affects the cortical microtubules of Pisum sativum root cells: Experiments on tungsten-molybdenum antagonism. Plant Biol. 2010, 12, 114–124. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. The cortical microtubules are a universal target of tungsten toxicity among land plant taxa. J. Biol. Res. (Thessaloniki) 2010, 13, 59–66. [Google Scholar]
- Adamakis, I-D.S.; Panteris, E.; Eleftheriou, E.P. The fatal effect of tungsten on Pisum sativum L. root cells: Indications for endoplasmic reticulum stress-induced programmed cell death. Planta 2011, 234, 21–34. [Google Scholar] [CrossRef]
- Xiong, J.; Fu, G.; Yang, Y.; Zhu, C.; Tao, L. Tungstate: Is it really a specific nitrate reductase inhibitor in plant nitric oxide research? J. Exp. Bot. 2012, 63, 33–41. [Google Scholar] [CrossRef]
- Hille, R. Molybdenum and tungsten in biology. Trends Biochem. Sci. 2002, 27, 360–367. [Google Scholar]
- Keith, L.S.; Moffett, D.B.; Rosemond, Z.A.; Wohlers, D.W. Agency for Toxic Substances and Disease Registry. ATSDR evaluation of health effects of tungsten and relevance to public health. Toxicol. Ind. Health. 2007, 23, 347–387. [Google Scholar] [CrossRef]
- Jonas, W.; van der Voet, G.B.; Todorov, T.I.; Centeno, J.A.; Ives, J.; Mullick, F.G. Metals and health: A clinical toxicological perspective on tungsten and review of the literature. Mil. Med. 2007, 172, 1002–1005. [Google Scholar]
- Jelikić-Stankov, M.; Uskoković-Marković, S.; Holclajtner-Antunović, I.; Todorović, M.; Djurdjević, P. Compounds of Mo, V and W in biochemistry and their biomedical activity. J. Trace Elem. Med. Biol. 2007, 21, 8–16. [Google Scholar]
- Johnson, D.R.; Inouye, L.S.; Bednar, A.J.; Clarke, J.U.; Winfield, L.E.; Boyd, R.E.; Ang, C.Y.; Goss, J. Tungsten bioavailability and toxicity in sunflowers (Helianthus annuus L.). Land Contam. Reclam. 2009, 17, 141–151. [Google Scholar] [CrossRef]
- Kumar, A.; Aery, N.C. Effect of tungsten on growth, biochemical constituents, molybdenum and tungsten contents in wheat. Plant Soil Environ. 2011, 57, 519–525. [Google Scholar]
- Hale, K.L.; Tufan, H.A.; Pickering, I.J.; George, G.N.; Terry, N.; Pilon, M.; Pilon-Smits, E.A.H. Anthocyanins facilitate tungsten accumulation in Brassica. Physiol. Plant. 2002, 116, 351–358. [Google Scholar] [CrossRef]
- Seidov, I.M. Apropos of the soil method of decontaminating sewage waters containing cobalt and wolfram. Gig. Sanit. 1963, 28, 93–96. [Google Scholar]
- Seidov, I.M. Some data on the effect of vegetables enriched with cobalt and wolfram on the animal body. Vopr. Pitan. 1964, 23, 73–77. [Google Scholar]
- Johnson, M.K.; Rees, D.C.; Adams, M.W.W. Tungstoenzymes. Chem. Rev. 1996, 96, 2817–2839. [Google Scholar] [CrossRef]
- Higgins, E.S.; Richert, D.A.; Westerfeld, W.W. Molybdenum deficiency and tungstate inhibition studies. J. Nutr. 1956, 59, 539–559. [Google Scholar]
- Bittner, F.; Mendel, R.R. Cell biology of molybdenum. In Cell Biology of Metals and Nutrients; Hell, R., Mendel, R.R., Eds.; Springer Press: Heidelberg, Germany, 2010; Volume 17, Plant Cell Monographs. [Google Scholar]
- Omarov, R.T.; Akaba, S.; Koshiba, T.; Lips, S.W. Aldehyde oxidase in roots, leaves and seeds of barley (Hordeum vulgare L.). J. Exp. Bot. 1999, 50, 63–69. [Google Scholar]
- Jiang, F.; Heilmeier, H.; Hartung, W. Abscisic acid relations of plants grown on tungsten enriched substrates. Plant Soil 2007, 301, 37–49. [Google Scholar] [CrossRef]
- Jiang, X.Y.; Omarov, T.; Yesbergenova, S.Z.; Sagi, M. The effect of molybdate and tungstate in the growth medium on abscisic acid content and the Mo-hydroxylases activities in barley (Hordeum vulgare L.). Plant Sci. 2004, 167, 297–300. [Google Scholar] [CrossRef]
- Brychkova, G.; Yarmolinsky, D.; Fluhr, R.; Sagi, M. The determination of sulfite levels and its oxidation in plant leaves. Plant Sci. 2012, 190, 123–130. [Google Scholar] [CrossRef]
- Xu, J.; Yin, H.X.; Liu, X.J.; Yuan, T.; Mi, Q.; Yang, L.L.; Xie, Z.X.; Wang, W.Y. Nitric oxide alleviates Fe deficiency-induced stress in Solanum nigrum. Biol. Plantarum 2009, 53, 784–788. [Google Scholar] [CrossRef]
- Chen, J.; Zhong, Y.M.; Zhang, H.Q.; Shi, Z.Q. Nitrate reductase-dependent nitric oxide production is involved in microcystin-LR-induced oxidative stress in Brassica rapa. Water Air Soil Pollut. 2012, 223, 4141–4152. [Google Scholar] [CrossRef]
- Yu, L.Z.; Wu, X.Q.; Ye, J.R; Zhang, S.N.; Wang, S. NOS-like-mediated nitric oxide is involved in Pinus thunbergii response to the invasion of Bursaphelenchus xylophilu. Plant Cell Reports 2012. [Google Scholar] [CrossRef]
- Kaiser, W.M.; Weine, H.; Kandlbinder, A.; Tsai, C.B.; Rockel, P.; Sonoda, M.; Planchet, E. Modulation of nitrate reductase: Some new insights, an unusual case and a potentially important side reaction. J. Exp. Bot. 2002, 53, 875–882. [Google Scholar] [CrossRef]
- Xiong, J.; Lu, H.; Lu, K.; Duan, Y.; An, L.; Zhu, C. Cadmium decreases crown root number by decreasing endogenous nitric oxide which is indispensable for crown root primordial initiation in rice seedlings. Planta 2009, 230, 599–610. [Google Scholar]
- Wang, B.L.; Tang, X.Y.; Cheng, L.Y.; Zhang, A.Z.; Zhang, W.H.; Zhang, F.S.; Liu, J.Q.; Cao, Y.; Allan, D.L.; Vance, C.P.; et al. Nitric oxide is involved in phosphorus deficiency-induced cluster-root development and citrate exudation in white lupin. New Phytol. 2010, 187, 1112–1123. [Google Scholar]
- Kumar, R.T.; Prommer, J.; Watanabe, M. Endogenous nitric oxide generation in protoplast chloroplasts. Plant Cell Reports 2012. [Google Scholar] [CrossRef]
- Bamford, J.E.; Butler, A.D.; Heim, K.E.; Pittinger, C.A.; Lemus, R.; Staveley, J.P.; Lee, K.B.; Venezia, C.; Pardus, M.J. Toxicity of sodium tungstate to earthworm, oat, radish, and lettuce. Environ. Toxicol. Chem. 2011, 30, 2312–2318. [Google Scholar] [CrossRef]
- Kumar, A.; Aery, N.C. Effect of tungsten on the growth, dry-matter production, and biochemical constituents of cowpea. Commun. Soil Sci. Plant Anal. 2012, 43, 1098–1107. [Google Scholar] [CrossRef]
- Bednar, A.J.; Jones, W.T.; Boyd, R.E.; Ringelberg, D.B.; Larson, S.L. Geochemical parameters influencing tungsten motility in soils. J. Environ. Qual. 2008, 37, 399–402. [Google Scholar]
- Siemann, S.; Schneider, K.; Oley, M.; Mueller, A. Characterization of a tungsten-substituted nitrogenase isolated from Rhodobacter capsulatus. Biochemistry 2003, 42, 3846–3857. [Google Scholar] [CrossRef]
- Johnson, D.R.; Ang, C.; Bednar, A.J.; Inouye, L.S. Tungsten effects on phosphate-dependent biochemical pathways are species and liver cell line dependent. Toxicol. Sci. 2010, 116, 523–532. [Google Scholar] [CrossRef]
- Sanz, L.; James, A.H.M.; Dewitte, W. To divide and to rule; regulating cell division in roots during post-embryonic growth. In Progress in Botany; Ulrich, E.L., Beyschlag, W., Burkhard, B., Francis, D., Eds.; Springer Verlag: Berlin, Germany, 2012; Volume 72, pp. 57–80. [Google Scholar]
- Barceló, J.; Poschenrieder, C. Structural and ultrastructural changes in heavy metal exposed plants. In Heavy Metal Stress in Plants. From Molecules to Ecosystems; Prasad, M.N.V., Hagemeyer, J., Eds.; SpringerVerlag: Berlin, Germany, 1999; pp. 183–205. [Google Scholar]
- Krysiak, C.; Mazuś, B.; Buchowicz, J. Generation of DNA double-strand breaks and inhibition of somatic embryogenesis by tungsten microparticles in wheat. Plant Cell Tissue Organ Cult. 1999, 58, 163–170. [Google Scholar]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Melissa, P. Effects of hexavalent chromium on microtubule organization, ER distribution and callose deposition in root tip cells of Allium cepa L. Protoplasma 2012, 249, 401–416. [Google Scholar] [CrossRef]
- Eleftheriou, E.P.; Adamakis, I.-D.S.; Fatsiou, M.; Panteris, E. Hexavalent chromium disrupts mitosis by stabilizing microtubules in Lens culinaris Moench. root tip cells. Physiol. Plant. 2012. [Google Scholar] [CrossRef]
- Lam, E.; Pontier, D.; del Pozo, O. Die and let live—Programmed cell death in plants. Curr. Opin. Plant Biol. 1999, 2, 502–507. [Google Scholar]
- Gadjev, I.; Stone, J.M.; Gechev, T.S. Programmed cell death in plants: New insights into redox regulation and the role of hydrogen peroxide. In International Review of Cell and Molecular Biology; Elsevier Inc.: Amsterdam, The Netherland, 2008; Volume 270, pp. 87–144. [Google Scholar]
- Jones, A.M. Programmed cell death in development and defence. Plant Physiol. 2001, 125, 94–97. [Google Scholar] [CrossRef]
- Koukalova, B.; Kovarik, A.; Fajkus, J.; Siroky, J. Chromatin fragmentation associated with apoptotic changes in tobacco cells exposed to cold stress. FEBS Lett. 1997, 414, 289–292. [Google Scholar]
- Obara, K.; Kuriyama, H.; Fukuda, H. Direct evidence of active and rapid nuclear degradation triggered by vacuole rupture during programmed cell death in Zinnia. Plant Physiol. 2001, 125, 615–626. [Google Scholar] [CrossRef]
- Lam, E. Vacuolar proteases livening up programmed cell death. Trends Cell Biol. 2005, 15, 124–127. [Google Scholar]
- Kuriyama, H.; Fukuda, H. Developmental programmed cell death in plants. Curr. Opin. Plant Biol. 2002, 5, 568–573. [Google Scholar]
- Kuthanova, A.; Opatrny, Z.; Fischer, L. Is internucleosomal DNA fragmentation an indicator of programmed death in plant cells? J. Exp. Bot. 2008, 59, 2233–2240. [Google Scholar] [CrossRef]
- Reape, T.J.; Molony, E.M.; McCabe, P.F. Programmed cell death in plants: Distinguishing between different modes. J. Exp. Bot. 2008, 59, 435–444. [Google Scholar]
- Fojtová, M.; Kovařík, A. Genotoxic effect of cadmium is associated with apoptotic changes in tobacco cells. Plant Cell Environ. 2000, 23, 531–537. [Google Scholar] [CrossRef]
- Pan, J.W.; Zhu, M.Y.; Chen, H. Aluminum-induced cell death in root-tip cells of barley. Environ. Exp. Bot. 2001, 46, 71–79. [Google Scholar]
- Yakimova, E.T.; Kapchina-Toteva, V.M.; Laarhoven, L.J.; Harren, F.M.; Woltering, E.J. Involvement of ethylene and lipid signalling in cadmium-induced programmed cell death in tomato suspension cells. Plant Physiol. Biochem. 2006, 44, 581–589. [Google Scholar] [CrossRef]
- Yakimova, E.T.; Kapchina-Toteva, V.M.; Woltering, E.J. Signal transduction events in aluminum-induced cell death in tomato suspension cells. J. Plant Physiol. 2007, 164, 702–708. [Google Scholar]
- Iakimova, E.T.; Woltering, E.J.; Kapchina-Toteva, V.M.; Harren, F.J.M.; Cristescu, S.M. Cadmium toxicity in cultured tomato cells—Role of ethylene, proteases and oxidative stress in cell death signaling. Cell Biol. Int. 2008, 32, 1521–1529. [Google Scholar] [CrossRef]
- Suarez, M.F.; Filonova, L.H.; Smertenko, A.; Savenkov, E.I.; Clapham, D.H.; von Arnold, S.; Zhivotovsky, B.; Bozhkov, P.V. Metacaspase-dependent programmed cell death is essential for plant embryogenesis. Curr. Biol. 2004, 14, R339–R340. [Google Scholar]
- Bonneau, L.; Ge, Y.; Drury, G.E.; Gallois, P. What happened to plant caspases? J. Exp. Bot. 2008, 59, 491–499. [Google Scholar] [CrossRef]
- Rotari, V.I.; He, R.; Gallois, P. Death by proteases in plants: Whodunit. Physiol. Plant. 2005, 123, 376–385. [Google Scholar]
- Elbaz, M.; Avni, A.; Weil, M. Constitutive caspase-like machinery executes programmed cell death in plant cells. Cell Death Differ. 2002, 9, 726–733. [Google Scholar] [CrossRef]
- Mlejnek, P.; Procházka, S. Activation of caspase-like proteases and induction of apoptosis by isopentenyladenosine in tobacco BY-2 cells. Planta 2002, 215, 158–166. [Google Scholar]
- Kisselev, A.F.; Garcia-Calvo, M.; Overkleeft, H.S.; Peterson, E.; Pennington, M.W.; Ploegh, H.L.; Thornberry, N.A.; Goldberg, A.L. The caspase-like sites of proteasomes, their substrate specificity, new inhibitors and substrates, and allosteric interactions with the trypsin-like sites. J. Biol. Chem. 2003, 278, 35869–35877. [Google Scholar]
- Hatsugai, N.; Iwasaki, S.; Tamura, K.; Kondo, M.; Fuji, K.; Ogasawara, K.; Nishimura, M.; Nishimura, I.H. A novel membrane fusion-mediated plant immunity against bacterial pathogens. Genes Dev. 2009, 23, 2496–2506. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 1999, 11, 431–443. [Google Scholar]
- Gechev, T.S.; van Breusege, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar]
- Yokouchi, M.; Hiramatsu, N.; Hayakawa, K.; Kasai, A.; Takano, Y.; Yao, J.; Kitamura, M. Atypical, bidirectional regulation of cadmium-induced apoptosis via distinct signaling of unfolded protein response. Cell Death Differ. 2007, 14, 1467–1474. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, W.; Li, B.; Wang, Y.; Li, K.; Sodmergen; Han, C.; Zhang, Y.; Li, X. An endoplasmic reticulum response pathway mediates programmed cell death of root tip induced by water stress in Arabidopsis. New Phytol. 2010, 186, 681–695. [Google Scholar] [CrossRef]
- Cacas, J.L. Devil inside: Does plant programmed cell death involve the endomembrane system? Plant Cell Environ. 2010, 33, 1453–1473. [Google Scholar]
- Schröder, M.; Kaufman, R.J. ER stress and the unfolded protein response. Mutat. Res. 2005, 569, 29–63. [Google Scholar]
- Iwata, Y.; Koizumi, N. Unfolded protein response followed by induction of cell death in cultured tobacco cells treated with tunicamycin. Planta 2005, 220, 804–807. [Google Scholar] [CrossRef]
- Urade, R. The endoplasmic reticulum stress signalling pathways in plants. BioFactors 2009, 35, 326–331. [Google Scholar]
- Mendel, R.R.; Kruse, T. Cell biology of molybdenum in plants and humans. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1568–1579. [Google Scholar] [CrossRef]
- Schwarz, G.; Schulze, J.; Bittner, F.; Eilers, T.; Kuper, J.; Bollmann, G.; Nerlich, A.; Brinkmann, H.; Mendel, R.R. The molybdenum cofactor biosynthetic protein Cnx1 complements molybdate-repairable mutants, transfers molybdenum to the metal binding pterin, and is associated with the cytoskeleton. Plant Cell 2000, 12, 2455–2471. [Google Scholar]
- Kirsch, J.; Betz, H. Widespread expression of gephyrin, a putative glycine receptor-tubulin linker protein, in rat brain. Brain Res. 1993, 621, 301–310. [Google Scholar] [CrossRef]
- Adamakis, I.D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten effects on the actin microfilament network of meristematic cells of Zea mays L. In Proceedings of the 34th Scientific Conference of Hellenic Association for Biological Sciences, Trikala, Greece, 17–19 May, 2012; p. 3.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. Tungsten Toxicity in Plants. Plants 2012, 1, 82-99. https://doi.org/10.3390/plants1020082
Adamakis I-DS, Panteris E, Eleftheriou EP. Tungsten Toxicity in Plants. Plants. 2012; 1(2):82-99. https://doi.org/10.3390/plants1020082
Chicago/Turabian StyleAdamakis, Ioannis-Dimosthenis S., Emmanuel Panteris, and Eleftherios P. Eleftheriou. 2012. "Tungsten Toxicity in Plants" Plants 1, no. 2: 82-99. https://doi.org/10.3390/plants1020082
APA StyleAdamakis, I.-D. S., Panteris, E., & Eleftheriou, E. P. (2012). Tungsten Toxicity in Plants. Plants, 1(2), 82-99. https://doi.org/10.3390/plants1020082