
Developmental Mechanism of Limb Field Specification along the Anterior–Posterior Axis during Vertebrate Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
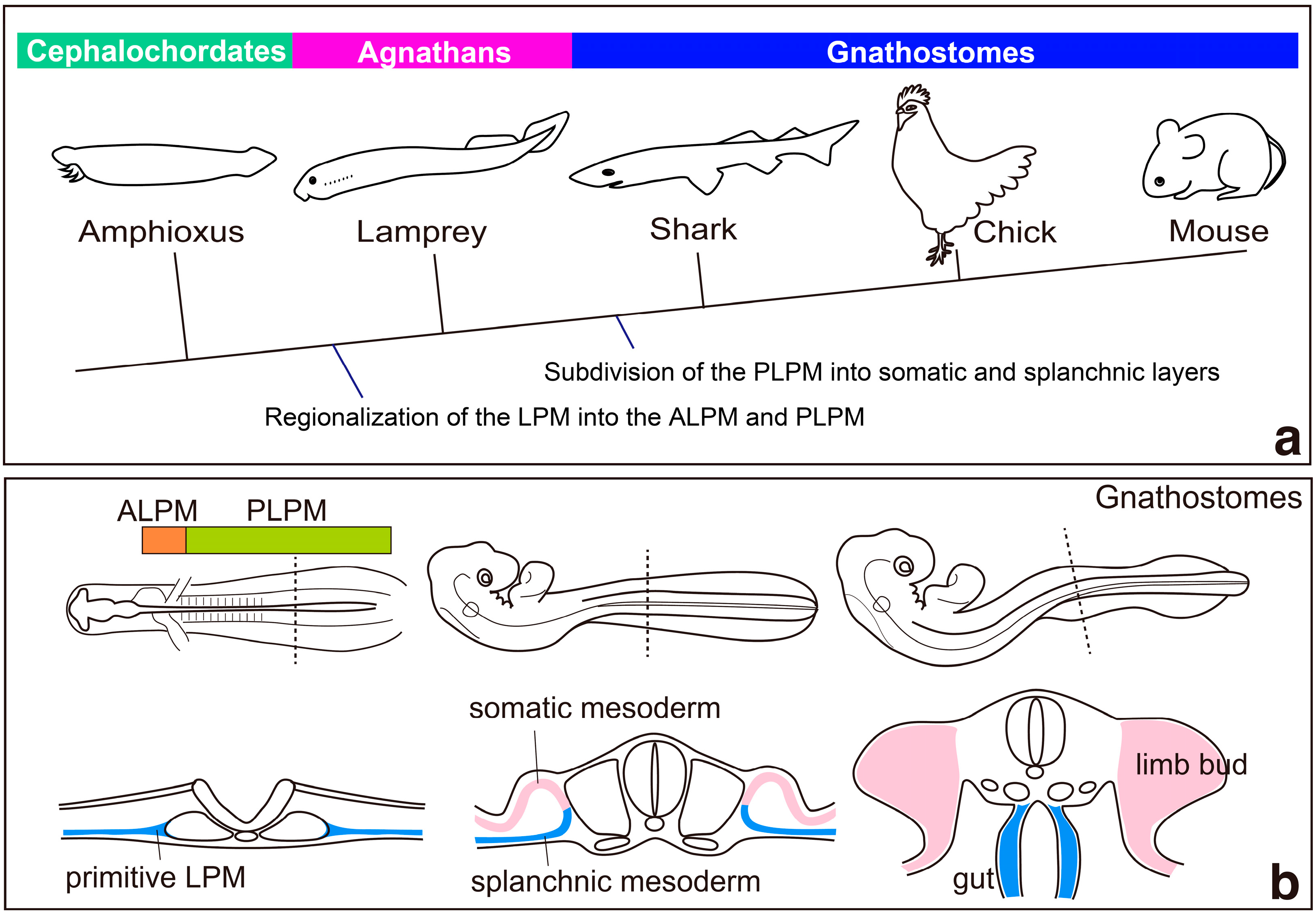
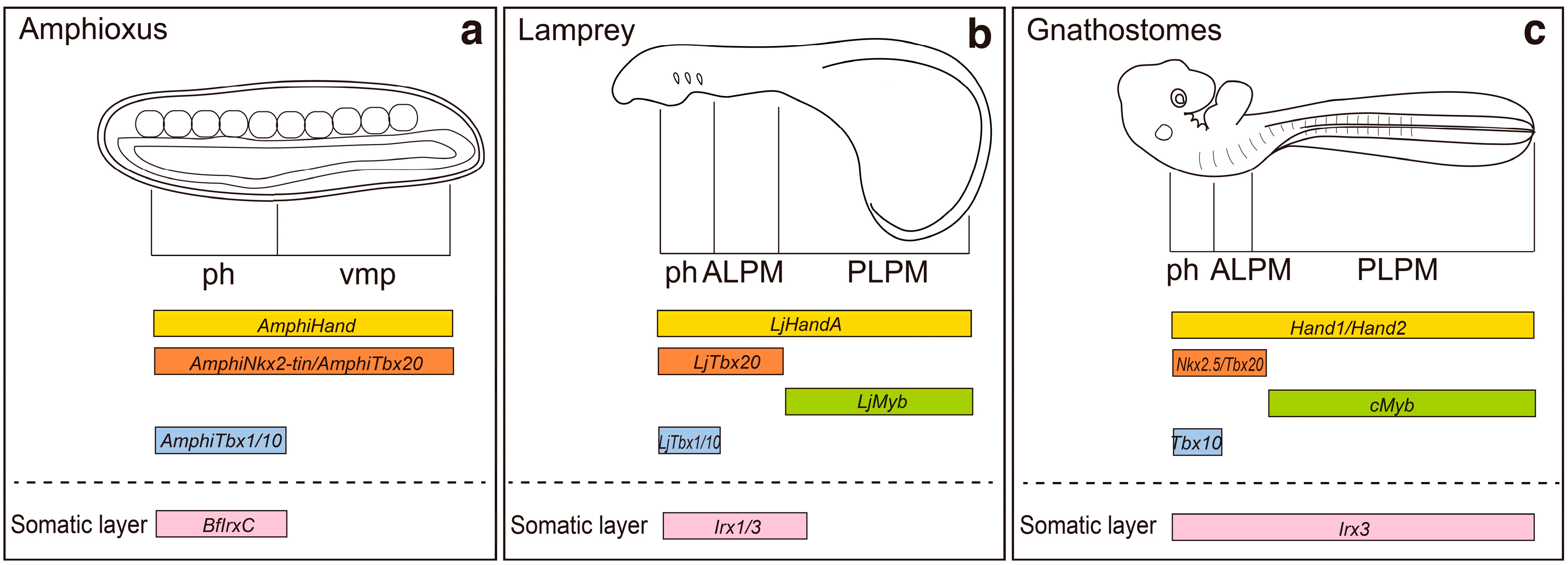
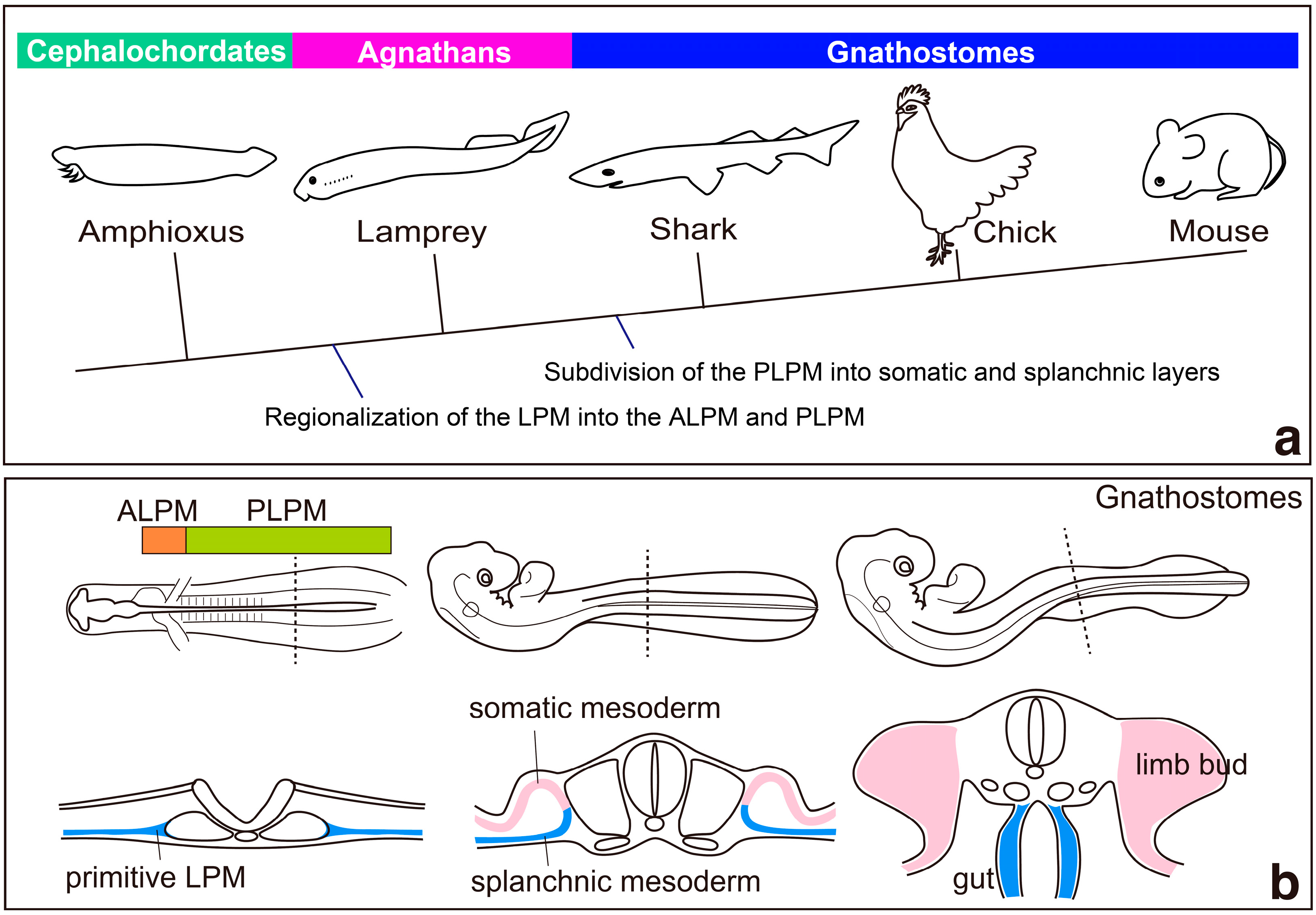
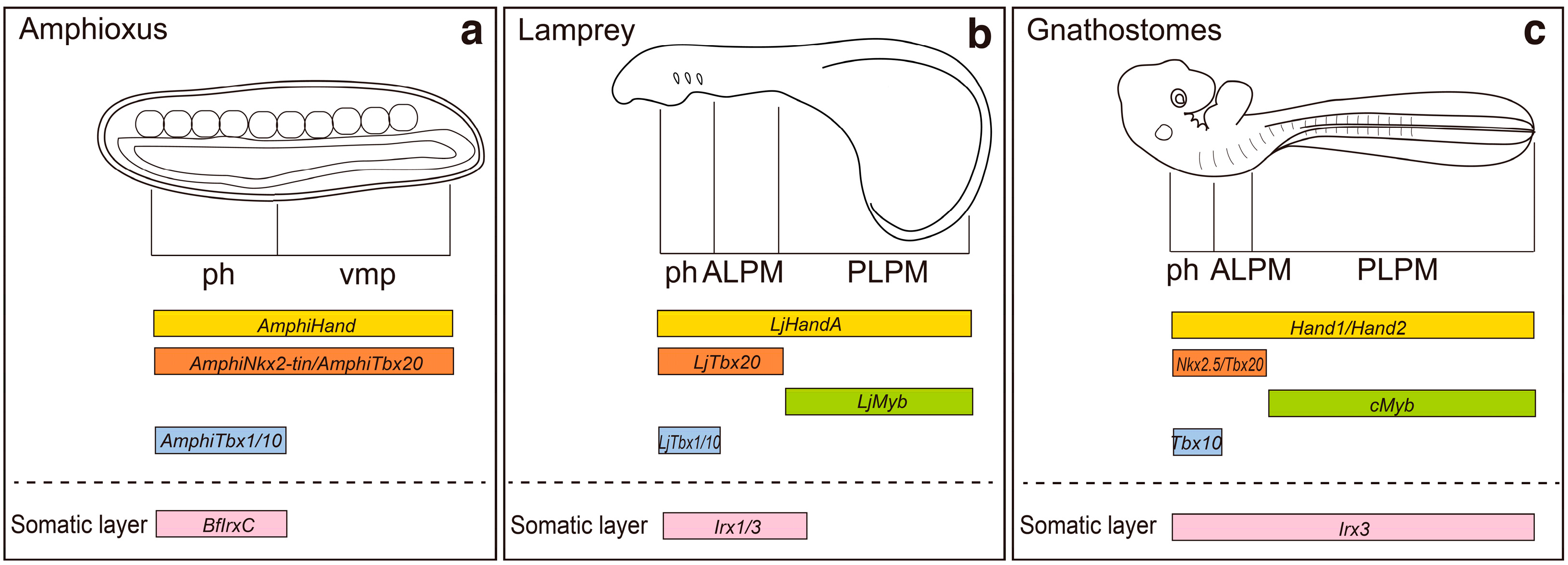
2. Regionalization of the Lateral Plate Mesoderm into the ALPM and the PLPM: Retinoic Acid and Hox Genes
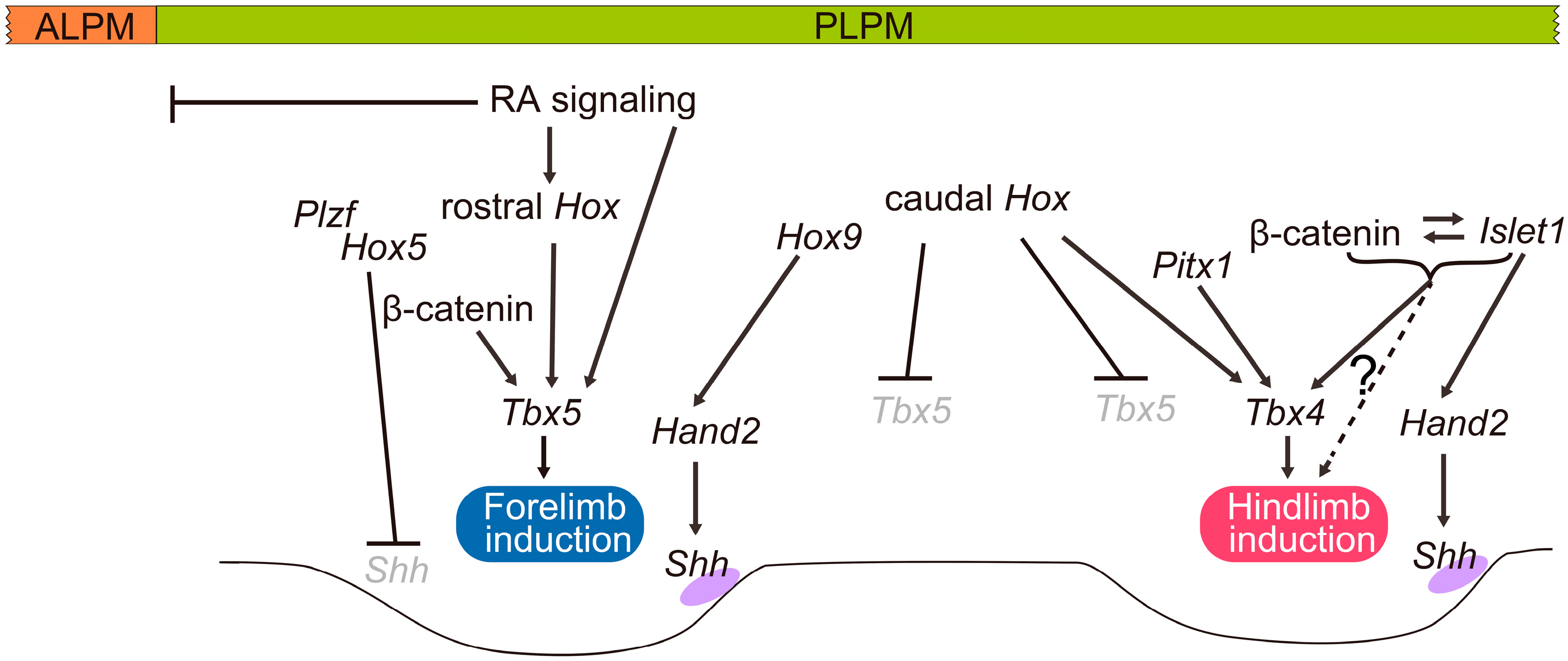
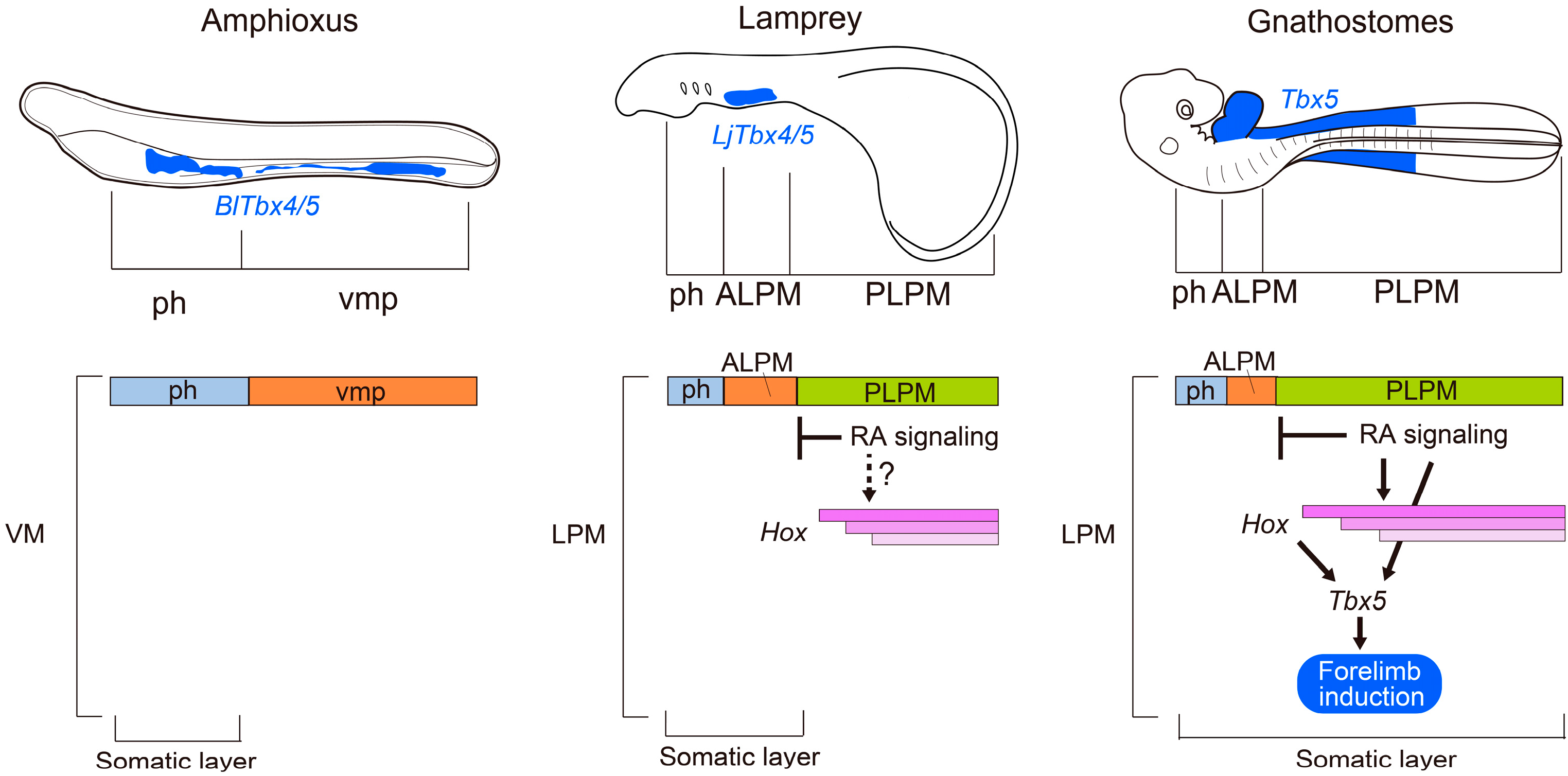
3. Specification of Limb-Forming Fields: Tbx4, Tbx5 and Hox Genes
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Coates, M.I. The origin of vertebrate limbs. Dev. Suppl. 1994, 169–180. [Google Scholar]
- Janvier, P. Early Vertebrates; Clarendon Press: Oxford, UK, 1996. [Google Scholar]
- Donoghue, P.C.; Forey, P.L.; Aldridge, R.J. Conodont affinity and chordate phylogeny. Biol. Rev. Camb. Philos. Soc. 2000, 75, 191–251. [Google Scholar] [CrossRef] [PubMed]
- Onimaru, K.; Shoguchi, E.; Kuratani, S.; Tanaka, M. Development and evolution of the lateral plate mesoderm: Comparative analysis of amphioxus and lamprey with implications for the acquisition of paired fins. Dev. Biol. 2011, 359, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Waxman, J.S.; Keegan, B.R.; Roberts, R.W.; Poss, K.D.; Yelon, D. Hoxb5b acts downstream of retinoic acid signaling in the forelimb field to restrict heart field potential in zebrafish. Dev. Cell 2008, 15, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sirbu, I.O.; Mic, F.A.; Molotkova, N.; Molotkov, A.; Kumar, S.; Duester, G. Retinoic acid promotes limb induction through effects on body axis extension but is unnecessary for limb patterning. Curr. Biol. 2009, 19, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Cohn, M.J.; Patel, K.; Krumlauf, R.; Wilkinson, D.G.; Clarke, J.D.; Tickle, C. Hox9 genes and vertebrate limb specification. Nature 1997, 387, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, J.L.; Burke, A.C. Hox genes and morphological identity: Axial versus lateral patterning in the vertebrate mesoderm. Development 2000, 127, 4265–4275. [Google Scholar] [PubMed]
- Funayama, N.; Sato, Y.; Matsumoto, K.; Ogura, T.; Takahashi, Y. Coelom formation: Binary decision of the lateral plate mesoderm is controlled by the ectoderm. Development 1999, 126, 4129–4138. [Google Scholar] [PubMed]
- Logan, M. Finger or toe: The molecular basis of limb identity. Development 2003, 130, 6401–6410. [Google Scholar] [CrossRef] [PubMed]
- Minguillon, C.; Nishimoto, S.; Wood, S.; Vendrell, E.; Gibson-Brown, J.J.; Logan, M.P. Hox genes regulate the onset of Tbx5 expression in the forelimb. Development 2012, 139, 3180–3188. [Google Scholar] [CrossRef] [PubMed]
- Minguillon, C.; del Buono, J.; Logan, M.P. Tbx5 and Tbx4 are not sufficient to determine limb-specific morphologies but have common roles in initiating limb outgrowth. Dev. Cell 2005, 8, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Lun, K.; Rauch, G.J.; Rhinn, M.; Piotrowski, T.; Houart, C.; Sordino, P.; Kuchler, A.M.; Schulte-Merker, S.; Geisler, R.; et al. Retinoic acid signalling in the zebrafish embryo is necessary during pre-segmentation stages to pattern the anterior-posterior axis of the CNS and to induce a pectoral fin bud. Development 2002, 129, 2851–2865. [Google Scholar] [PubMed]
- Gibert, Y.; Gajewski, A.; Meyer, A.; Begemann, G. Induction and prepatterning of the zebrafish pectoral fin bud requires axial retinoic acid signaling. Development 2006, 133, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Bruce, A.E.; Oates, A.C.; Prince, V.E.; Ho, R.K. Additional hox clusters in the zebrafish: Divergent expression patterns belie equivalent activities of duplicate hoxB5 genes. Evol. Dev. 2001, 3, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Oosterveen, T.; Niederreither, K.; Dolle, P.; Chambon, P.; Meijlink, F.; Deschamps, J. Retinoids regulate the anterior expression boundaries of 5′ Hoxb genes in posterior hindbrain. EMBO J. 2003, 22, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, J.; Nonchev, S.; Gould, A.; Whiting, J.; Krumlauf, R. Selectivity, sharing and competitive interactions in the regulation of hoxb genes. EMBO J. 1998, 17, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Rancourt, D.E.; Tsuzuki, T.; Capecchi, M.R. Genetic interaction between hoxb-5 and hoxb-6 is revealed by nonallelic noncomplementation. Genes Dev. 1995, 9, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Ryckebusch, L.; Wang, Z.; Bertrand, N.; Lin, S.C.; Chi, X.; Schwartz, R.; Zaffran, S.; Niederreither, K. Retinoic acid deficiency alters second heart field formation. Proc. Natl. Acad. Sci. USA 2008, 105, 2913–2918. [Google Scholar] [CrossRef] [PubMed]
- Searcy, R.D.; Yutzey, K.E. Analysis of Hox gene expression during early avian heart development. Dev. Dyn. 1998, 213, 82–91. [Google Scholar] [CrossRef]
- Lo, P.C.; Frasch, M. Establishing A-P polarity in the embryonic heart tube: A conserved function of hox genes in Drosophila and vertebrates? Trends Cardiovasc. Med. 2003, 13, 182–187. [Google Scholar] [CrossRef]
- Chisaka, O.; Capecchi, M.R. Regionally restricted developmental defects resulting from targeted disruption of the mouse homeobox gene hox-1.5. Nature 1991, 350, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, N.; Xavier-Neto, J. From the bottom of the heart: Anteroposterior decisions in cardiac muscle differentiation. Curr. Opin. Cell Biol. 2000, 12, 742–746. [Google Scholar] [CrossRef]
- Bertrand, N.; Roux, M.; Ryckebusch, L.; Niederreither, K.; Dolle, P.; Moon, A.; Capecchi, M.; Zaffran, S. Hox genes define distinct progenitor sub-domains within the second heart field. Dev. Biol. 2011, 353, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Stratford, T.; Horton, C.; Maden, M. Retinoic acid is required for the initiation of outgrowth in the chick limb bud. Curr. Biol. 1996, 6, 1124–1133. [Google Scholar] [CrossRef]
- Mic, F.A.; Haselbeck, R.J.; Cuenca, A.E.; Duester, G. Novel retinoic acid generating activities in the neural tube and heart identified by conditional rescue of Raldh2 null mutant mice. Development 2002, 129, 2271–2282. [Google Scholar] [PubMed]
- Niederreither, K.; Subbarayan, V.; Dolle, P.; Chambon, P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 1999, 21, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, T.J.; Zhao, X.; Sandell, L.L.; Evans, S.M.; Trainor, P.A.; Duester, G. Antagonism between retinoic acid and fibroblast growth factor signaling during limb development. Cell Rep. 2013, 3, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.R.; Lee, Y.; Poss, K.D.; Yelon, D. Reiterative roles for fgf signaling in the establishment of size and proportion of the zebrafish heart. Dev. Biol. 2008, 321, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.D.; Venkatesh, T.V.; Holland, L.Z.; Jacobs, D.K.; Bodmer, R. Amphink2-tin, an amphioxus homeobox gene expressed in myocardial progenitors: Insights into evolution of the vertebrate heart. Dev. Biol. 2003, 255, 128–137. [Google Scholar] [CrossRef]
- Charite, J.; McFadden, D.G.; Olson, E.N. The bHLH transcription factor dHand controls Sonic hedgehog expression and establishment of the zone of polarizing activity during limb development. Development 2000, 127, 2461–2470. [Google Scholar] [PubMed]
- Deimling, S.J.; Drysdale, T.A. Retinoic acid regulates anterior-posterior patterning within the lateral plate mesoderm of Xenopus. Mech. Dev. 2009, 126, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Yelon, D.; Ticho, B.; Halpern, M.E.; Ruvinsky, I.; Ho, R.K.; Silver, L.M.; Stainier, D.Y. The bHLH transcription factor hand2 plays parallel roles in zebrafish heart and pectoral fin development. Development 2000, 127, 2573–2582. [Google Scholar] [PubMed]
- Buchberger, A.; Pabst, O.; Brand, T.; Seidl, K.; Arnold, H.H. Chick NKx-2.3 represents a novel family member of vertebrate homologues to the Drosophila homeobox gene tinman: Differential expression of cNKx-2.3 and cNKkx-2.5 during heart and gut development. Mech. Dev. 1996, 56, 151–163. [Google Scholar] [CrossRef]
- Kraus, F.; Haenig, B.; Kispert, A. Cloning and expression analysis of the mouse T-box gene tbx20. Mech. Dev. 2001, 100, 87–91. [Google Scholar] [CrossRef]
- Yamagishi, T.; Nakajima, Y.; Nishimatsu, S.; Nohno, T.; Ando, K.; Nakamura, H. Expression of tbx20 rna during chick heart development. Dev. Dyn. 2004, 230, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Kokubo, N.; Matsuura, M.; Onimaru, K.; Tiecke, E.; Kuraku, S.; Kuratani, S.; Tanaka, M. Mechanisms of heart development in the japanese lamprey, lethenteron japonicum. Evol. Dev. 2010, 12, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. Molecular and evolutionary basis of limb field specification and limb initiation. Dev. Growth Differ. 2013, 55, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Garg, V.; Yamagishi, C.; Hu, T.; Kathiriya, I.S.; Yamagishi, H.; Srivastava, D. Tbx1, a digeorge syndrome candidate gene, is regulated by sonic hedgehog during pharyngeal arch development. Dev. Biol. 2001, 235, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Tiecke, E.; Matsuura, M.; Kokubo, N.; Kuraku, S.; Kusakabe, R.; Kuratani, S.; Tanaka, M. Identification and developmental expression of two Tbx1/10-related genes in the agnathan lethenteron japonicum. Dev. Genes Evol. 2007, 217, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Meulemans, D.; Bronner-Fraser, M. Insights from amphioxus into the evolution of vertebrate cartilage. PLoS ONE 2007, 2, e787. [Google Scholar] [CrossRef] [PubMed]
- Escriva, H.; Bertrand, S.; Germain, P.; Robinson-Rechavi, M.; Umbhauer, M.; Cartry, J.; Duffraisse, M.; Holland, L.; Gronemeyer, H.; Laudet, V. Neofunctionalization in vertebrates: The example of retinoic acid receptors. PLoS Genet. 2006, 2, e102. [Google Scholar] [CrossRef] [PubMed]
- Castillo, H.A.; Cravo, R.M.; Azambuja, A.P.; Simoes-Costa, M.S.; Sura-Trueba, S.; Gonzalez, J.; Slonimsky, E.; Almeida, K.; Abreu, J.G.; de Almeida, M.A.; et al. Insights into the organization of dorsal spinal cord pathways from an evolutionarily conserved raldh2 intronic enhancer. Development 2010, 137, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Koop, D.; Chen, J.; Theodosiou, M.; Carvalho, J.E.; Alvarez, S.; de Lera, A.R.; Holland, L.Z.; Schubert, M. Roles of retinoic acid and Tbx1/10 in pharyngeal segmentation: Amphioxus and the ancestral chordate condition. EvoDevo 2014, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, M.; Holland, N.D.; Laudet, V.; Holland, L.Z. A retinoic acid-Hox hierarchy controls both anterior/posterior patterning and neuronal specification in the developing central nervous system of the cephalochordate amphioxus. Dev. Biol. 2006, 296, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Pasqualetti, M.; Takio, Y.; Hirano, S.; Rijli, F.M.; Kuratani, S. Segmental development of reticulospinal and branchiomotor neurons in lamprey: Insights into the evolution of the vertebrate hindbrain. Development 2004, 131, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, S.; Ueki, T.; Hirano, S.; Aizawa, S. Rostral truncation of a cyclostome, lampetra japonica, induced by all-trans retinoic acid defines the head/trunk interface of the vertebrate body. Dev. Dyn. 1998, 211, 35–51. [Google Scholar] [CrossRef]
- Bertrand, S.; Aldea, D.; Oulion, S.; Subirana, L.; de Lera, A.R.; Somorjai, I.; Escriva, H. Evolution of the role of RA and FGF signals in the control of somitogenesis in chordates. PLoS ONE 2015, 10, e0136587. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, S.; Minguillon, C.; Wood, S.; Logan, M.P. A combination of activation and repression by a colinear Hox code controls forelimb-restricted expression of Tbx5 and reveals hox protein specificity. PLoS Genet. 2014, 10, e1004245. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, S.; Wilde, S.M.; Wood, S.; Logan, M.P. RA acts in a coherent feed-forward mechanism with Tbx5 to control limb bud induction and initiation. Cell Rep. 2015, 12, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.W.; Holland, L.Z.; Williams, N.A.; Holland, N.D. An amphioxus homeobox gene: Sequence conservation, spatial expression during development and insights into vertebrate evolution. Development 1992, 116, 653–661. [Google Scholar] [PubMed]
- Wada, H.; Garcia-Fernandez, J.; Holland, P.W. Colinear and segmental expression of amphioxus Hox genes. Dev. Biol. 1999, 213, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Yu, J.K.; Holland, N.D.; Escriva, H.; Laudet, V.; Holland, L.Z. Retinoic acid signaling acts via Hox1 to establish the posterior limit of the pharynx in the chordate amphioxus. Development 2005, 132, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Holland, N.D.; Escriva, H.; Holland, L.Z.; Laudet, V. Retinoic acid influences anteroposterior positioning of epidermal sensory neurons and their gene expression in a developing chordate (amphioxus). Proc. Nat. Acad. Sci. USA 2004, 101, 10320–10325. [Google Scholar] [CrossRef] [PubMed]
- Koop, D.; Holland, N.D.; Semon, M.; Alvarez, S.; de Lera, A.R.; Laudet, V.; Holland, L.Z.; Schubert, M. Retinoic acid signaling targets Hox genes during the amphioxus gastrula stage: Insights into early anterior-posterior patterning of the chordate body plan. Dev. Biol. 2010, 338, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Yu, R.; Kurokawa, D. Anterior migration of lateral plate mesodermal cells during embryogenesis of the pufferfish Takifugu niphobles: Insight into the rostral positioning of pelvic fins. J. Anat. 2015, 227, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Tulenko, F.J.; McCauley, D.W.; Mackenzie, E.L.; Mazan, S.; Kuratani, S.; Sugahara, F.; Kusakabe, R.; Burke, A.C. Body wall development in lamprey and a new perspective on the origin of vertebrate paired fins. Proc. Nat. Acad. Sci. USA 2013, 110, 11899–11904. [Google Scholar] [CrossRef] [PubMed]
- Kaltenbach, S.L.; Holland, L.Z.; Holland, N.D.; Koop, D. Developmental expression of the three iroquois genes of amphioxus (BfIrxa, BfIrxb, and BfIrxc) with special attention to the gastrula organizer and anteroposterior boundaries in the central nervous system. Gene Expr. Patterns 2009, 9, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Wright, C.V.; Hardwicke, J.; de Robertis, E.M. A gradient of homeodomain protein in developing forelimbs of Xenopus and mouse embryos. Cell 1988, 55, 1017–1024. [Google Scholar] [CrossRef]
- Becker, D.; Eid, R.; Schughart, K. The limb/lpm enhancer of the murine Hoxb6 gene: Reporter gene analysis in transgenic embryos and studies of DNA-protein interactions. Pharm. Acta Helv. 1996, 71, 29–35. [Google Scholar] [CrossRef]
- Nelson, C.E.; Morgan, B.A.; Burke, A.C.; Laufer, E.; DiMambro, E.; Murtaugh, L.C.; Gonzales, E.; Tessarollo, L.; Parada, L.F.; Tabin, C. Analysis of Hox gene expression in the chick limb bud. Development 1996, 122, 1449–1466. [Google Scholar] [PubMed]
- Tamura, K.; Yonei-Tamura, S.; Belmonte, J.C. Differential expression of Tbx4 and Tbx5 in zebrafish fin buds. Mech. Dev. 1999, 87, 181–184. [Google Scholar] [CrossRef]
- Gibson-Brown, J.J.; Agulnik, S.I.; Chapman, D.L.; Alexiou, M.; Garvey, N.; Silver, L.M.; Papaioannou, V.E. Evidence of a role for T-box genes in the evolution of limb morphogenesis and the specification of forelimb/hindlimb identity. Mech. Dev. 1996, 56, 93–101. [Google Scholar] [CrossRef]
- Isaac, A.; Rodriguez-Esteban, C.; Ryan, A.; Altabef, M.; Tsukui, T.; Patel, K.; Tickle, C.; Izpisua-Belmonte, J.C. Tbx genes and limb identity in chick embryo development. Development 1998, 125, 1867–1875. [Google Scholar] [PubMed]
- Ahn, D.G.; Kourakis, M.J.; Rohde, L.A.; Silver, L.M.; Ho, R.K. T-box gene tbx5 is essential for formation of the pectoral limb bud. Nature 2002, 417, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Garrity, D.M.; Childs, S.; Fishman, M.C. The heartstrings mutation in zebrafish causes heart/fin Tbx5 deficiency syndrome. Development 2002, 129, 4635–4645. [Google Scholar] [PubMed]
- Ng, J.K.; Kawakami, Y.; Buscher, D.; Raya, A.; Itoh, T.; Koth, C.M.; Rodriguez Esteban, C.; Rodriguez-Leon, J.; Garrity, D.M.; Fishman, M.C.; et al. The limb identity gene Tbx5 promotes limb initiation by interacting with Wnt2b and Fgf10. Development 2002, 129, 5161–5170. [Google Scholar] [PubMed]
- Agarwal, P.; Wylie, J.N.; Galceran, J.; Arkhitko, O.; Li, C.; Deng, C.; Grosschedl, R.; Bruneau, B.G. Tbx5 is essential for forelimb bud initiation following patterning of the limb field in the mouse embryo. Development 2003, 130, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Rallis, C.; Bruneau, B.G.; del Buono, J.; Seidman, C.E.; Seidman, J.G.; Nissim, S.; Tabin, C.J.; Logan, M.P. Tbx5 is required for forelimb bud formation and continued outgrowth. Development 2003, 130, 2741–2751. [Google Scholar] [CrossRef] [PubMed]
- Basson, C.T.; Bachinsky, D.R.; Lin, R.C.; Levi, T.; Elkins, J.A.; Soults, J.; Grayzel, D.; Kroumpouzou, E.; Traill, T.A.; Leblanc-Straceski, J.; et al. Mutations in human TBX5 [corrected] cause limb and cardiac malformation in holt-oram syndrome. Nat. Genet. 1997, 15, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Newbury-Ecob, R.A.; Terrett, J.A.; Wilson, D.I.; Curtis, A.R.; Yi, C.H.; Gebuhr, T.; Bullen, P.J.; Robson, S.C.; Strachan, T.; et al. Holt-oram syndrome is caused by mutations in TBX5, a member of the Brachyury (T) gene family. Nat. Genet. 1997, 15, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Popperl, H.; Rikhof, H.; Chang, H.; Haffter, P.; Kimmel, C.B.; Moens, C.B. Lazarus is a novel pbx gene that globally mediates hox gene function in zebrafish. Mol. Cell 2000, 6, 255–267. [Google Scholar] [CrossRef]
- Xu, B.; Hrycaj, S.M.; McIntyre, D.C.; Baker, N.C.; Takeuchi, J.K.; Jeannotte, L.; Gaber, Z.B.; Novitch, B.G.; Wellik, D.M. Hox5 interacts with Plzf to restrict Shh expression in the developing forelimb. Proc. Nat. Acad. Sci. USA 2013, 110, 19438–19443. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Wellik, D.M. Axial Hox9 activity establishes the posterior field in the developing forelimb. Proc. Nat. Acad. Sci. USA 2011, 108, 4888–4891. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Capdevila, J.; Buscher, D.; Itoh, T.; Rodriguez Esteban, C.; Izpisua Belmonte, J.C. Wnt signals control FGF-dependent limb initiation and aer induction in the chick embryo. Cell 2001, 104, 891–900. [Google Scholar] [CrossRef]
- Galceran, J.; Hsu, S.C.; Grosschedl, R. Rescue of a wnt mutation by an activated form of LEF-1: Regulation of maintenance but not initiation of brachyury expression. Proc. Nat. Acad. Sci. USA 2001, 98, 8668–8673. [Google Scholar] [CrossRef] [PubMed]
- Cohn, M.J.; Tickle, C. Developmental basis of limblessness and axial patterning in snakes. Nature 1999, 399, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Munsterberg, A.; Anderson, W.G.; Prescott, A.R.; Hazon, N.; Tickle, C. Fin development in a cartilaginous fish and the origin of vertebrate limbs. Nature 2002, 416, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Hale, L.A.; Amores, A.; Yan, Y.L.; Cresko, W.A.; Suzuki, T.; Postlethwait, J.H. Developmental genetic basis for the evolution of pelvic fin loss in the pufferfish takifugu rubripes. Dev. Biol. 2005, 281, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Marcil, A.; Dumontier, E.; Chamberland, M.; Camper, S.A.; Drouin, J. Pitx1 and Pitx2 are required for development of hindlimb buds. Development 2003, 130, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Naiche, L.A.; Papaioannou, V.E. Loss of Tbx4 blocks hindlimb development and affects vascularization and fusion of the allantois. Development 2003, 130, 2681–2693. [Google Scholar] [CrossRef] [PubMed]
- Naiche, L.A.; Papaioannou, V.E. Tbx4 is not required for hindlimb identity or post-bud hindlimb outgrowth. Development 2007, 134, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cai, C.L.; Lin, L.; Qyang, Y.; Chung, C.; Monteiro, R.M.; Mummery, C.L.; Fishman, G.I.; Cogen, A.; Evans, S. Isl1cre reveals a common bmp pathway in heart and limb development. Development 2006, 133, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Marti, M.; Kawakami, H.; Itou, J.; Quach, T.; Johnson, A.; Sahara, S.; O’Leary, D.D.; Nakagawa, Y.; Lewandoski, M.; et al. Islet1-mediated activation of the beta-catenin pathway is necessary for hindlimb initiation in mice. Development 2011, 138, 4465–4473. [Google Scholar] [CrossRef] [PubMed]
- Lanctot, C.; Moreau, A.; Chamberland, M.; Tremblay, M.L.; Drouin, J. Hindlimb patterning and mandible development require the Ptx1 gene. Development 1999, 126, 1805–1810. [Google Scholar] [PubMed]
- Logan, M.; Tabin, C.J. Role of Pitx1 upstream of tbx4 in specification of hindlimb identity. Science 1999, 283, 1736–1739. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.J.; Tanaka, M.; Prescott, A.; Tickle, C. Expression of limb initiation genes and clues to the morphological diversification of threespine stickleback. Curr. Biol. 2003, 13, R951–R952. [Google Scholar] [CrossRef] [PubMed]
- Szeto, D.P.; Rodriguez-Esteban, C.; Ryan, A.K.; O’Connell, S.M.; Liu, F.; Kioussi, C.; Gleiberman, A.S.; Izpisua-Belmonte, J.C.; Rosenfeld, M.G. Role of the bicoid-related homeodomain factor Pitx1 in specifying hindlimb morphogenesis and pituitary development. Genes Dev. 1999, 13, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Logan, M.P. Pitx1 is necessary for normal initiation of hindlimb outgrowth through regulation of Tbx4 expression and shapes hindlimb morphologies via targeted growth control. Development 2011, 138, 5301–5309. [Google Scholar] [CrossRef] [PubMed]
- Liebenberg, F. A pedigree with unusual anomalies of the elbows, wrists and hands in five generations. S. Afr. Med. J. 1973, 47, 745–748. [Google Scholar] [PubMed]
- Spielmann, M.; Brancati, F.; Krawitz, P.M.; Robinson, P.N.; Ibrahim, D.M.; Franke, M.; Hecht, J.; Lohan, S.; Dathe, K.; Nardone, A.M.; et al. Homeotic arm-to-leg transformation associated with genomic rearrangements at the pitx1 locus. Am. J. Hum. Genet. 2012, 91, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Don, E.K.; de Jong-Curtain, T.A.; Doggett, K.; Hall, T.E.; Heng, B.; Badrock, A.P.; Winnick, C.; Nicholson, G.A.; Guillemin, G.J.; Currie, P.D.; et al. Genetic basis of hindlimb loss in a naturally occurring vertebrate model. Biol. Open 2016, 5, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.D.; Marks, M.E.; Peichel, C.L.; Blackman, B.K.; Nereng, K.S.; Jonsson, B.; Schluter, D.; Kingsley, D.M. Genetic and developmental basis of evolutionary pelvic reduction in threespine sticklebacks. Nature 2004, 428, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.F.; Marks, M.E.; Jones, F.C.; Villarreal, G., Jr.; Shapiro, M.D.; Brady, S.D.; Southwick, A.M.; Absher, D.M.; Grimwood, J.; Schmutz, J.; et al. Adaptive evolution of pelvic reduction in sticklebacks by recurrent deletion of a Pitx1 enhancer. Science 2010, 327, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M. Revealing the mechanisms of the rostral shift of pelvic fins among teleost fishes. Evol. Dev. 2011, 13, 382–390. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of anterior/posterior patterning of the axial skeleton by growth/differentiation factor 11. Nat. Genet. 1999, 22, 260–264. [Google Scholar] [PubMed]
- Liu, J.P. The function of growth/differentiation factor 11 (gdf11) in rostrocaudal patterning of the developing spinal cord. Development 2006, 133, 2865–2874. [Google Scholar] [CrossRef] [PubMed]
- Dasen, J.S.; Liu, J.P.; Jessell, T.M. Motor neuron columnar fate imposed by sequential phases of Hox-c activity. Nature 2003, 425, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Drill, E.; Lance-Jones, C. Ectopic expression of Hoxd10 in thoracic spinal segments induces motoneurons with a lumbosacral molecular profile and axon projections to the limb. Dev. Dyn. 2004, 231, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, G.; Scott, S.A.; Capecchi, M.R. Hoxc10 and Hoxd10 regulate mouse columnar, divisional and motor pool identity of lumbar motoneurons. Development 2008, 135, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Toyono, T.; Akamine, A.; Joyner, A. Expression of growth/differentiation factor 11, a new member of the BMP/TGFbeta superfamily during mouse embryogenesis. Mech. Dev. 1999, 80, 185–189. [Google Scholar] [CrossRef]
- Liu, J.P.; Laufer, E.; Jessell, T.M. Assigning the positional identity of spinal motor neurons: Rostrocaudal patterning of Hox-c expression by fgfs, gdf11, and retinoids. Neuron 2001, 32, 997–1012. [Google Scholar] [CrossRef]
- Murata, Y.; Tamura, M.; Aita, Y.; Fujimura, K.; Murakami, Y.; Okabe, M.; Okada, N.; Tanaka, M. Allometric growth of the trunk leads to the rostral shift of the pelvic fin in teleost fishes. Dev. Biol. 2010, 347, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Tanaka, M. Evolution of motor innervation to vertebrate fins and limbs. Dev. Biol. 2011, 355, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Jurberg, A.D.; Aires, R.; Varela-Lasheras, I.; Novoa, A.; Mallo, M. Switching axial progenitors from producing trunk to tail tissues in vertebrate embryos. Dev. Cell 2013, 25, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Itou, J.; Kawakami, H.; Quach, T.; Osterwalder, M.; Evans, S.M.; Zeller, R.; Kawakami, Y. Islet1 regulates establishment of the posterior hindlimb field upstream of the Hand2-Shh morphoregulatory gene network in mouse embryos. Development 2012, 139, 1620–1629. [Google Scholar] [CrossRef] [PubMed]
- Coates, M.I.; Cohn, M. Vertebrate axial and appendicular patterning: The early development of paired appendages. Amer. Zool. 1999, 39, 676–685. [Google Scholar] [CrossRef]
- Horton, A.C.; Mahadevan, N.R.; Minguillon, C.; Osoegawa, K.; Rokhsar, D.S.; Ruvinsky, I.; de Jong, P.J.; Logan, M.P.; Gibson-Brown, J.J. Conservation of linkage and evolution of developmental function within the Tbx2/3/4/5 subfamily of T-box genes: Implications for the origin of vertebrate limbs. Dev. Genes Evol. 2008, 218, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Anaya, J.; Albuixech-Crespo, B.; Somorjai, I.M.; Carmona, R.; Oisi, Y.; Alvarez, S.; Kuratani, S.; Munoz-Chapuli, R.; Garcia-Fernandez, J. The evolutionary origins of chordate hematopoiesis and vertebrate endothelia. Dev. Biol. 2013, 375, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Minguillon, C.; Gibson-Brown, J.J.; Logan, M.P. Tbx4/5 gene duplication and the origin of vertebrate paired appendages. Proc. Nat. Acad. Sci. USA 2009, 106, 21726–21730. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, M. Developmental Mechanism of Limb Field Specification along the Anterior–Posterior Axis during Vertebrate Evolution. J. Dev. Biol. 2016, 4, 18. https://doi.org/10.3390/jdb4020018
Tanaka M. Developmental Mechanism of Limb Field Specification along the Anterior–Posterior Axis during Vertebrate Evolution. Journal of Developmental Biology. 2016; 4(2):18. https://doi.org/10.3390/jdb4020018
Chicago/Turabian StyleTanaka, Mikiko. 2016. "Developmental Mechanism of Limb Field Specification along the Anterior–Posterior Axis during Vertebrate Evolution" Journal of Developmental Biology 4, no. 2: 18. https://doi.org/10.3390/jdb4020018