The Epicardium and Coronary Artery Formation

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Cardiac Vascular System
3. The Embryonic Epicardium and the Coronary Vascular System Are a Developmental Unit
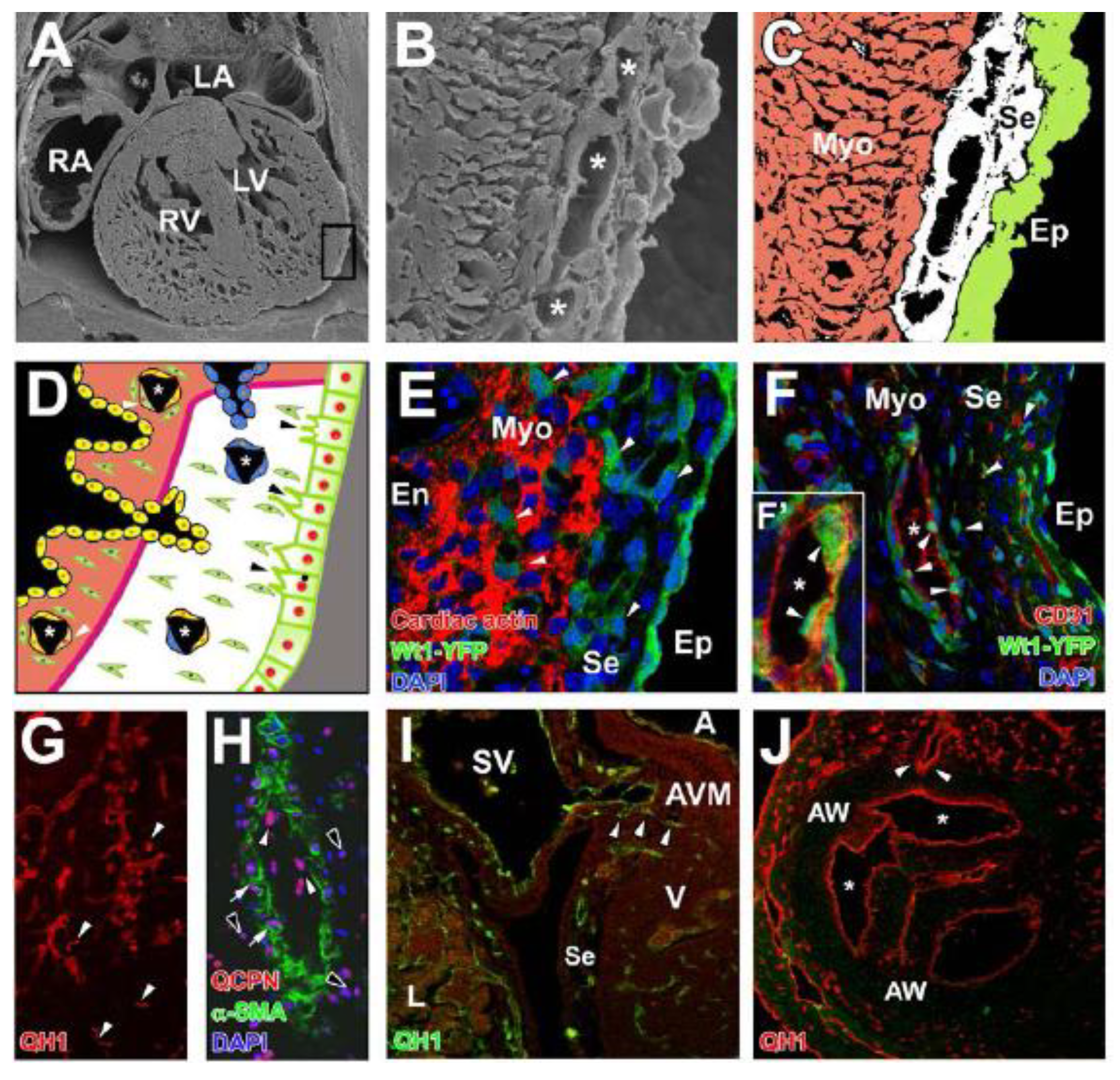
3.1. The Proepicardium and the Formation of the Primitive Epicardium
3.2. Epicardial Epithelial-to-Mesenchymal Transition
4. Vascularizing the Vertebrate Heart
5. The Elements of the Coronary System
5.1. Making the Coronary Tree Grow: Vasculogenesis versus Angiogenesis
5.2. Chicken or Mice?
5.3. Epicardially-Derived Cell (EPDC) Adult Fates
6. Epicardial Cell Contribution to Coronary Blood Vessel
6.1. Sources of Coronary Endothelial Cells
6.2. Sources of Coronary Smooth Muscle Cells

6.3. Sources of Coronary Fibroblasts
7. Building an Arteriovenous Coronary Vascular Tree
Coronary Arteries and Veins

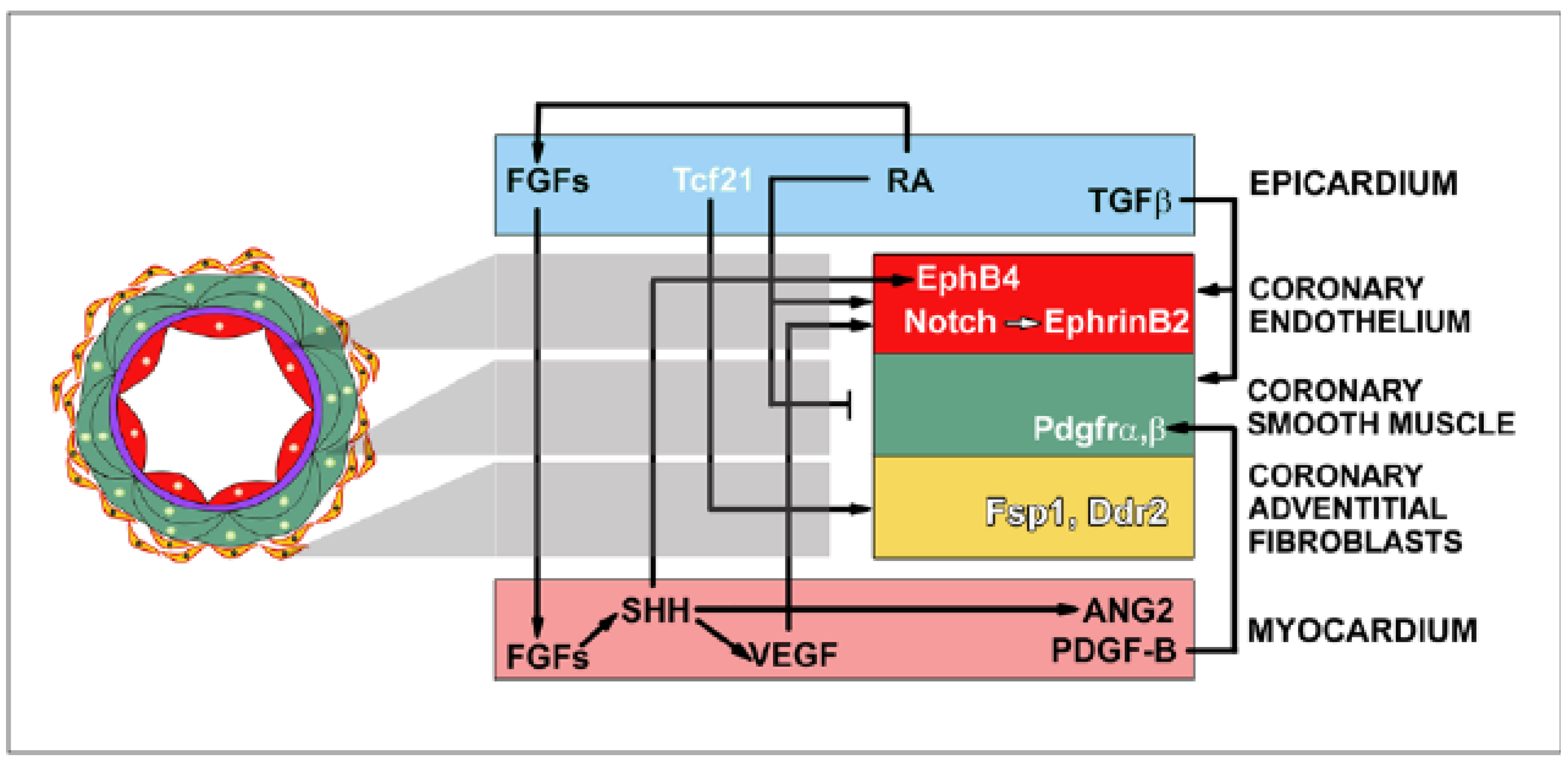
8. Signaling Coordination during Epicardial Development
- (1)
- Regulation of epicardial epithelial-to-mesenchymal transition: The Wt1 gene encodes for a transcription factor reported to control the transcriptional repressor, Snail [17], epicardial retinoic acid signaling [20] and Wnt/beta-catenin-dependent signaling [18]. Wt1 can be thus regarded as a coordinating gene in the regulation of epithelial-to-mesenchymal transition, as it promotes the loss of cell adhesion (via the cadherin repressor Snail), sustains pro-epithelial-to-mesenchymal transition epicardial retinoid synthesis and the associated activation of the canonical Wnt pathway.
- (2)
- Differentiation of coronary cell progenitors: As previously indicated, Wt1 is involved in the regulation of epicardial retinoid signaling, and disruption of retinoic acid signaling in the epicardium downregulates PDGFRα,β expression in epicardial-derived cells, therefore altering coronary progenitor cell differentiation [20]. The Notch/Delta pathway, a key molecule in the activation of endothelial and endocardial signaling [57], is required for the promotion of arterial fate during coronary blood vessel development [58,59]. It is not known how Notch signaling interacts with the crucial function played by growth factors secreted by the embryonic myocardium, like FGFs and VEGF, in the regulation of coronary endothelial vasculogenesis [60]. These two growth factors are closely related, as VEGF has been reported to be dependent on myocardial FGF-induced Hedgehog activity [61]. On the other hand, Notch is also likely to be involved in the control of arteriovenous coronary endothelium differentiation. In this context, the downregulation of the nuclear transcription factor, COUP-TFII (a Notch repressor, [62]), and the upregulation of ephrinB2 tyrosine kinase expression in CA progenitor cells has been suggested to mark the beginning of coronary artery endothelium re-specification [56]. However, this point has not yet been proven, and the genetic regulation of this process remains unknown. Tcf21, another transcription factor, is required for the specification of fibroblast (but not smooth muscle cell) lineages from the epicardium prior to epicardial epithelial-to-mesenchymal transition [25].
- (3)
- Endocardium contribution to embryonic coronary vessels: It is still unknown which signals trigger the outgrowth of the endocardium to participate in coronary blood vessel formation. It has been suggested that the key mechanism is provided by a transmural (endocardial-to-epicardial) increasing gradient of hypoxia that activates myocardial VEGF synthesis, promoting endocardial sprouting [32]. Spatio-temporal patterning of the endocardial sprouts have been proposed to be partially regulated by cell membrane-bound ephrins [56] and semaphorins [47].
- (4)
- Maturation of coronary blood vessels: The stabilization of the embryonic coronary endothelial outline requires perivascular/coronary smooth muscle differentiation and the initiation of endothelial-smooth muscle cell-to-cell interaction. Synergistic retinoic acid and VEGF signaling seems to regulate the physiological delay of coronary smooth muscle differentiation until an extensive coronary capillary network has formed [22]. FGFs seem to have a key developmental function in the regulation of transmural organization of coronary arteries [63,64]. Furthermore, cardiomyocytes also play an important role in the patterning and mural location of coronary arteries, as shown by experiments disrupting myocardial cell polarity [31]. On a final note, we want to emphasize that the definitive cues needed to stabilize the spatial patterning of major coronary vessels seems to be triggered by the activation of an effective blood flow in coronary arteries through unknown mechanisms.
- (5)
- Connection of primitive coronary arteries to the aortic root: Presumptive embryonic coronary arteries grow from the ventricle towards the aortic root, where they eventually connect to the systemic blood flow via the coronary ostia. It is well known that coronary ostia open to the left and right Valsalva sinuses (coronary sinuses) of the aortic valve [36,37,65], but the mechanisms that control the patterning of the two coronary stems remains unknown. Moreover, not many explanations are available on the developmental mechanisms that prevent coronary arteries from connecting to the posterior aortic sinus or to the pulmonary artery, and fewer are the explanations of the anomalous origin of coronary ostia from the so-called “wrong sinus” or from the pulmonary root. A plausible explanation would combine both repulsive signals emanating from the subpulmonary myocardium [66] and some pro-vascular signals especially active at the dorsolateral cardiac outflow tract [67].
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; Fullerton, H.J.; Gillespie, C.; Hailpern, S.M.; Heit, J.A.; Howard, V.J.; Huffman, M.D.; Kissela, B.M.; Kittner, S.J.; Lackland, D.T.; Lichtman, J.H.; Lisabeth, L.D.; Magid, D.; Marcus, G.M.; Marelli, A.; Matchar, D.B.; McGuire, D.K.; Mohler, E.R.; Moy, C.S.; Mussolino, M.E.; Nichol, G.; Paynter, N.P.; Schreiner, P.J.; Sorlie, P.D.; Stein, J.; Turan, T.N.; Virani, S.S.; Wong, N.D.; Woo, D.; Turner, M.B. Heart disease and stroke statistics—2013 update: A report from the American Heart Association. Circulation 2013, 127, e6–e245. [Google Scholar] [CrossRef]
- Eichmann, A.; Yuan, L.; Moyon, D.; Lenoble, F.; Pardanaud, L.; Breant, C. Vascular development: from precursor cells to branched arterial and venous networks. Int. J. Dev. Biol. 2005, 49, 259–267. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; Muñoz-Chápuli, R. Origin of the vertebrate endothelial cell lineage: Ontology and Phylogeny. In Heart Development and Reegeneration; Rosenthal, N., Harvey, R.P., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2010; pp. 465–486. [Google Scholar]
- Leibowitz, J.O. The History of Coronary Heart Disease; University of California Press (Berkley): Ewing, NJ, USA, 1970. [Google Scholar]
- Männer, J. The development of pericardial villi in the chick embryo. Anat. Embryol. (Berl). 1992, 186, 379–385. [Google Scholar] [CrossRef]
- Wessels, A.; Pérez-Pomares, J.M. The epicardium and epicardially derived cells (EPDCs) as cardiac stem cells. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 276, 43–57. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; de la Pompa, J.L. Signaling during epicardium and coronary vessel development. Circ. Res. 2011, 109, 1429–1442. [Google Scholar] [CrossRef]
- Männer, J.; Pérez-Pomares, J.M.; Macías, D.; Muñoz-Chápuli, R. The origin, formation and developmental significance of the epicardium: a review. Cells Tissues Organs 2001, 169, 89–103. [Google Scholar] [CrossRef]
- Bouchey, D.; Drake, C.J.; Wunsch, A.M.; Little, C.D. Distribution of connective tissue proteins during development and neovascularization of the epicardium. 1996, 1, E104–E115. [Google Scholar]
- Pérez-Pomares, J.M.; Macías, D.; García-Garrido, L.; Muñoz-Chápuli, R. Contribution of the primitive epicardium to the subepicardial mesenchyme in hamster and chick embryos. Dev. Dyn. 1997, 210, 96–105. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; Macías, D.; García-Garrido, L.; Muñoz-Chápuli, R. The origin of the subepicardial mesenchyme in the avian embryo: an immunohistochemical and quail-chick chimera study. Dev. Biol. 1998, 200, 57–68. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; Phelps, A.; Sedmerova, M.; Carmona, R.; González-Iriarte, M.; Muñoz-Chápuli, R.; Wessels, A. Experimental Studies on the Spatiotemporal Expression of WT1 and RALDH2 in the Embryonic Avian Heart: A Model for the Regulation of Myocardial and Valvuloseptal Development by Epicardially Derived Cells (EPDCs). Dev. Biol. 2002, 247, 307–326. [Google Scholar] [CrossRef]
- Dettman, R.W.; Denetclaw, W.; Ordahl, C.P.; Bristow, J. Common epicardial origin of coronary vascular smooth muscle, perivascular fibroblasts, and intermyocardial fibroblasts in the avian heart. Dev. Biol. 1998, 193, 169–181. [Google Scholar] [CrossRef]
- Gittenberger-de Groot, A.C.; Vrancken Peeters, M.-P.F.M.; Mentink, M.M.T.; Gourdie, R.G.; Poelmann, R.E. Epicardium-Derived Cells Contribute a Novel Population to the Myocardial Wall and the Atrioventricular Cushions. Circ. Res. 1998, 82, 1043–1052. [Google Scholar] [CrossRef]
- Wessels, A.; van den Hoff, M.J.B.; Adamo, R.F.; Phelps, A.L.; Lockhart, M.M.; Sauls, K.; Briggs, L.E.; Norris, R.A.; van Wijk, B.; Perez-Pomares, J.M.; Dettman, R.W.; Burch, J.B.E. Epicardially derived fibroblasts preferentially contribute to the parietal leaflets of the atrioventricular valves in the murine heart. Dev. Biol. 2012, 366, 111–124. [Google Scholar] [CrossRef]
- Morabito, C.J.; Dettman, R.W.; Kattan, J.; Collier, J.M.; Bristow, J. Positive and negative regulation of epicardial-mesenchymal transformation during avian heart development. Dev. Biol. 2001, 234, 204–215. [Google Scholar] [CrossRef]
- Martínez-estrada, O.M.; Lettice, L.A.; Essafi, A.; Guadix, J.A.; Slight, J.; Velecela, V.V.; Hall, E.; Reichmann, J.; Devenney, P.S.; Hohenstein, P.; Hosen, N.; Hill, R.E.; Muñoz-Chapuli, R.; Hastie, N.D.; Nicholas, D. Wt1 is required for cardiovascular progenitor cell formation through transcriptional control of Snail and E-cadherin. Nat. Genet. 2010, 42, 89–93. [Google Scholar] [CrossRef]
- Von Gise, A.; Zhou, B.; Honor, L.B.; Ma, Q.; Petryk, A.; Pu, W.T. WT1 regulates epicardial epithelial to mesenchymal transition through β-catenin and retinoic acid signaling pathways. Dev. Biol. 2011, 356, 421–431. [Google Scholar]
- Casanova, J.C.; Travisano, S.; de la Pompa, J.L. Epithelial-to-mesenchymal transition in epicardium is independent of Snail1. Genesis 2013, 51, 32–40. [Google Scholar] [CrossRef]
- Guadix, J.A.; Ruiz-Villalba, A.; Lettice, L.; Velecela, V.; Muñoz-Chápuli, R.; Hastie, N.D.; Pérez-Pomares, J.M.; Martínez-Estrada, O.M. Wt1 controls retinoic acid signalling in embryonic epicardium through transcriptional activation of Raldh2. Development 2011, 138, 1093–1097. [Google Scholar] [CrossRef]
- Von Gise, A.; Pu, W.T. Endocardial and epicardial epithelial to mesenchymal transitions in heart development and disease. Circ. Res. 2012, 110, 1628–1645. [Google Scholar] [CrossRef]
- Azambuja, A.P.; Portillo-Sánchez, V.; Rodrigues, M.V.; Omae, S.V.; Schechtman, D.; Strauss, B.E.; Costanzi-Strauss, E.; Krieger, J.E.; Perez-Pomares, J.M.; Xavier-Neto, J. Retinoic acid and VEGF delay smooth muscle relative to endothelial differentiation to coordinate inner and outer coronary vessel wall morphogenesis. Circ. Res. 2010, 107, 204–216. [Google Scholar] [CrossRef]
- Braitsch, C.M.; Combs, M.D.; Quaggin, S.E.; Yutzey, K.E. Pod1/Tcf21 is regulated by retinoic acid signaling and inhibits differentiation of epicardium-derived cells into smooth muscle in the developing heart. Dev. Biol. 2012, 368, 345–357. [Google Scholar] [CrossRef]
- Wu, M.; Smith, C.L.; Hall, J.A.; Lee, I.; Luby-phelps, K.; Tallquist, M.D. Epicardial spindle orientation control cell entry into the myocardium. Dev. Cell 2011, 19, 114–125. [Google Scholar]
- Acharya, A.; Baek, S.T.; Huang, G.; Eskiocak, B.; Goetsch, S.; Sung, C.Y.; Banfi, S.; Sauer, M.F.; Olsen, G.S.; Duffield, J.S.; Olson, E.N.; Tallquist, M.D. The bHLH transcription factor Tcf21 is required for lineage-specific EMT of cardiac fibroblast progenitors. Development 2012, 139, 2139–2149. [Google Scholar] [CrossRef]
- Santer, R.M. Morphology and innervation of the fish heart. Adv. Anat. Embryol. Cell Biol. 1985, 89, 1–102. [Google Scholar] [CrossRef]
- Barnett, P.; Cox, G.K.; Farrell, A.P.; Farrell, N.D.; Galli, G.L.J.; Garofalo, F.; Gillis, T.E.; Hicks, J.W.; Icardo, J.M.; Jourdan, H.; McMahon, B.R.; Moorman, A.F. Ontogeny and Phylogeny of the Vertebrate Heart; Sedmera, D., Wang, T., Eds.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Maria, T.; Rodrigues, D.A.; Palmeira, J.A.O.; Mendonça, J.T.D.; Gomes, O.M. Estudo evolutivo da anatomia das artérias coronárias em espécies de vertebrados com técnica de moldagem em acetato de vinil. Vinilite 1999, 14, 331–339. [Google Scholar]
- Farrell, A.P.; Gamperl, A.K.; Francis, E.T.B. Comparative aspects of heart morphology. In Biology of the Reptilia; Gans, C., Gaunt, A., Eds.; Society for the study of amphibians and reptiles: Atlanta, GA, USA, 1998; pp. 375–424. [Google Scholar]
- Tevosian, S.G.; Deconinck, A.E.; Tanaka, M.; Schinke, M.; Litovsky, S.H.; Izumo, S.; Fujiwara, Y.; Orkin, S.H. FOG-2, a cofactor for GATA transcription factors, is essential for heart morphogenesis and development of coronary vessels from epicardium. Cell 2000, 101, 729–739. [Google Scholar] [CrossRef]
- Phillips, H.M.; Rhee, H.J.; Murdoch, J.N.; Hildreth, V.; Peat, J.D.; Anderson, R.H.; Copp, A.J.; Chaudhry, B.; Henderson, D.J. Disruption of planar cell polarity signaling results in congenital heart defects and cardiomyopathy attributable to early cardiomyocyte disorganization. Circ. Res. 2007, 101, 137–145. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, Z.; Lui, W.; Chen, X.; Wang, Y.; Chamberlain, A.A.; Moreno-Rodriguez, R.A.; Markwald, R.R.; O’Rourke, B.P.; Sharp, D.J.; Zheng, D.; Lenz, J.; Baldwin, H.S.; Chang, C.-P.; Zhou, B. Endocardial cells form the coronary arteries by angiogenesis through myocardial-endocardial VEGF signaling. Cell 2012, 151, 1083–1096. [Google Scholar] [CrossRef]
- Ogden, J.A. The Origin of Coronary Arteries. Circulation 1998, 38, 150. [Google Scholar]
- Hertig, A. Angiogenesis in the Early Human Chorion and in the Primary Placenta of the Macaque Monkey; Carnegie Institution of Washington: Washington, DC, USA, 1935. [Google Scholar]
- Folkman, J.; Haudenschild, C. Angiogenesis in vitro. Nature 1980, 288, 551–556. [Google Scholar] [CrossRef]
- Waldo, K.L.; Willner, W.; Kirby, M.L. Origin of the proximal coronary artery stems and a review of ventricular vascularization in the chick embryo. Am. J. Anat. 1990, 188, 109–120. [Google Scholar] [CrossRef]
- Bogers, A.J.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; Péault, B.M.; Huysmans, H.A. Development of the origin of the coronary arteries, a matter of ingrowth or outgrowth? Anat. Embryol. (Berl). 1989, 180, 437–441. [Google Scholar] [CrossRef]
- Kattan, J.; Dettman, R.W.; Bristow, J. Formation and remodeling of the coronary vascular bed in the embryonic avian heart. Dev. Dyn. 2004, 230, 34–43. [Google Scholar]
- Risau, W.; Flamme, I. Vasculogenesis. Annu. Rev. Cell Biol. 1995, 11, 73–91. [Google Scholar] [CrossRef]
- Männer, J. Does the subepicardial mesenchyme contribute myocardioblasts to the myocardium of the chick embryo heart? A quail-chick chimera study tracing the fate of the epicardial primordium. Anat. Rec. 1999, 255, 212–226. [Google Scholar] [CrossRef]
- Moore, A.W.; McInnes, L.; Kreidberg, J.; Hastie, N.D.; Schedl, A. YAC complementation shows a requirement for Wt1 in the development of epicardium, adrenal gland and throughout nephrogenesis. Development 1999, 126, 1845–1857. [Google Scholar]
- Kraus, F.; Haenig, B.; Kispert, A. Cloning and expression analysis of the mouse T-box gene tbx18. Mech. Dev. 2001, 100, 87–91. [Google Scholar] [CrossRef]
- Mikawa, T.; Gourdie, R.G. Pericardial mesoderm generates a population of coronary smooth muscle cells migrating into the heart along with ingrowth of the epicardial organ. Dev. Biol. 1996, 174, 221–232. [Google Scholar] [CrossRef]
- Vrancken Peeters, M.P.; Gittenberger-de Groot, A.C.; Mentink, M.M.; Poelmann, R.E. Smooth muscle cells and fibroblasts of the coronary arteries derive from epithelial-mesenchymal transformation of the epicardium. Anat. Embryol. (Berl). 1999, 199, 367–378. [Google Scholar] [CrossRef]
- Ruiz-Villalba, A.; Ziogas, A.; Ehrbar, M.; Pérez-Pomares, J.M. Characterization of epicardial-derived cardiac interstitial cells: Differentiation and mobilization of heart fibroblast progenitors. PLoS One 2013, 8, e53694. [Google Scholar]
- Guadix, J.A.; Carmona, R.; Muñoz-Chápuli, R.; Pérez-Pomares, J.M. In vivo and in vitro analysis of the vasculogenic potential of avian proepicardial and epicardial cells. Dev. Dyn. 2006, 235, 1014–1026. [Google Scholar] [CrossRef]
- Katz, T.C.; Singh, M.K.; Degenhardt, K.; Rivera-feliciano, J.; Johnson, R.L.; Epstein, J.A.; Tabin, C.J. Distinct Compartments of the Proepicardial Organ Give Rise to Coronary Vascular Endothelial Cells. Dev. Cell 2012, 22, 639–650. [Google Scholar] [CrossRef]
- Waldo, K.L.; Kumiski, D.H.; Kirby, M.L. Association of the cardiac neural crest with development of the coronary arteries in the chick embryo. Anat. Rec. 1994, 239, 315–331. [Google Scholar] [CrossRef]
- Smith, C.L.; Baek, S.T.; Sung, C.Y.; Tallquist, M.D. Epicardial-derived cell epithelial-to-mesenchymal transition and fate specification require PDGF receptor signaling. Circ. Res. 2011, 108, e15–e26. [Google Scholar] [CrossRef]
- Grego-Bessa, J.; Pérez-Pomares, J.M.; de la Pompa, J.L. Signaling Pathways in Valve Formation: The Origin of Congenital Defects. In Heart Development and Reegeneration; Rosenthal, N., Harvey, R.P., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 389–413. [Google Scholar]
- Le Noble, F.; Moyon, D.; Pardanaud, L.; Yuan, L.; Djonov, V.; Matthijsen, R.; Bréant, C.; Fleury, V.; Eichmann, A. Flow regulates arterial-venous differentiation in the chick embryo yolk sac. Development 2004, 131, 361–375. [Google Scholar]
- Wang, H.U.; Chen, Z.F.; Anderson, D.J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef]
- Zhong, T.P.; Childs, S.; Leu, J.P.; Fishman, M.C. Gridlock signalling pathway fashions the first embryonic artery. Nature 2001, 414, 216–220. [Google Scholar] [CrossRef]
- Herzog, Y.; Kalcheim, C.; Kahane, N.; Reshef, R.; Neufeld, G. Differential expression of neuropilin-1 and neuropilin-2 in arteries and veins. Mech. Dev. 2001, 109, 115–119. [Google Scholar] [CrossRef]
- Garcia-Cardeña, G.; Comander, J.; Anderson, K.R.; Blackman, B.R.; Gimbrone, M.A. Biomechanical activation of vascular endothelium as a determinant of its functional phenotype. Proc. Natl. Acad. Sci. USA 2001, 98, 4478–4485. [Google Scholar] [CrossRef]
- Red-Horse, K.; Ueno, H.; Weissman, I.L.; Krasnow, M.A. Coronary arteries form by developmental reprogramming of venous cells. Nature 2010, 464, 549–553. [Google Scholar] [CrossRef]
- Grego-Bessa, J.; Luna-Zurita, L.; del Monte, G.; Bolós, V.; Melgar, P.; Arandilla, A.; Garratt, A.N.; Zang, H.; Mukouyama, Y.; Chen, H.; Shou, W.; Ballestar, E.; Esteller, M.; Rojas, A.; Pérez-Pomares, J.M.; de la Pompa, J.L. Notch Signaling Is Essential for Ventricular Chamber Development. Dev. Cell 2007, 12, 415–429. [Google Scholar] [CrossRef]
- Del Monte, G.; Casanova, J.C.; Guadix, J.A.; MacGrogan, D.; Burch, J.B.E.; Pérez-Pomares, J.M.; de la Pompa, J.L. Differential Notch signaling in the epicardium is required for cardiac inflow development and coronary vessel morphogenesis. Circ. Res. 2011, 108, 824–836. [Google Scholar] [CrossRef]
- Grieskamp, T.; Rudat, C.; Lüdtke, T.H.-W.; Norden, J.; Kispert, A. Notch signaling regulates smooth muscle differentiation of epicardium-derived cells. Circ. Res. 2011, 108, 813–823. [Google Scholar] [CrossRef]
- Tomanek, R.J.; Ishii, Y.; Holifield, J.S.; Sjogren, C.L.; Hansen, H.K.; Mikawa, T. VEGF family members regulate myocardial tubulogenesis and coronary artery formation in the embryo. Circ. Res. 2006, 98, 947–953. [Google Scholar] [CrossRef]
- Lavine, K.J.; White, A.C.; Park, C.; Smith, C.S.; Choi, K.; Long, F.; Hui, C.; Ornitz, D.M. Fibroblast growth factor signals regulate a wave of Hedgehog activation that is essential for coronary vascular development. Genes Dev. 2006, 20, 1651–1666. [Google Scholar] [CrossRef]
- You, L.; Lin, F.; Lee, C.; DeMayo, F.; Tsai, M.; Tsai, S. Suppression of Notch signalling by the COUP-TFII transcription factor regulates vein identity. Nature 2005, 435, 1–6. [Google Scholar]
- Pennisi, D.J.; Mikawa, T. Normal patterning of the coronary capillary plexus is dependent on the correct transmural gradient of FGF expression in the myocardium. Dev. Biol. 2005, 279, 378–390. [Google Scholar] [CrossRef]
- Pennissi, D.J.; Mikawa, T.; Pennisi, D.J.; Mikawa, T.; Pennissi, D.J.; Mikawa, T. FGFR-1 is required by epicardium-derived cells for myocardial invasion and correct coronary vascular lineage differentiation. Development 2009, 328, 148–159. [Google Scholar]
- Ando, K.; Nakajima, Y.; Yamagishi, T.; Yamamoto, S.; Nakamura, H. Development of proximal coronary arteries in quail embryonic heart: multiple capillaries penetrating the aortic sinus fuse to form main coronary trunk. Circ. Res. 2004, 94, 346–352. [Google Scholar] [CrossRef]
- Théveniau-Ruissy, M.; Dandonneau, M.; Mesbah, K.; Ghez, O.; Mattei, M.-G.; Miquerol, L.; Kelly, R.G. The del22q11.2 candidate gene Tbx1 controls regional outflow tract identity and coronary artery patterning. Circ. Res. 2008, 103, 142–148. [Google Scholar] [CrossRef]
- Bernanke, D.H.; Velkey, J.M. Development of the coronary blood supply: Changing concepts and current ideas. Anat. Rec. 2002, 269, 198–208. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pires-Gomes, A.A.S.; Pérez-Pomares, J.M. The Epicardium and Coronary Artery Formation. J. Dev. Biol. 2013, 1, 186-202. https://doi.org/10.3390/jdb1030186
Pires-Gomes AAS, Pérez-Pomares JM. The Epicardium and Coronary Artery Formation. Journal of Developmental Biology. 2013; 1(3):186-202. https://doi.org/10.3390/jdb1030186
Chicago/Turabian StylePires-Gomes, Adriana A. S., and José M. Pérez-Pomares. 2013. "The Epicardium and Coronary Artery Formation" Journal of Developmental Biology 1, no. 3: 186-202. https://doi.org/10.3390/jdb1030186