Evolutionary Origin of the Proepicardium



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Comparative Anatomy of the Proepicardium

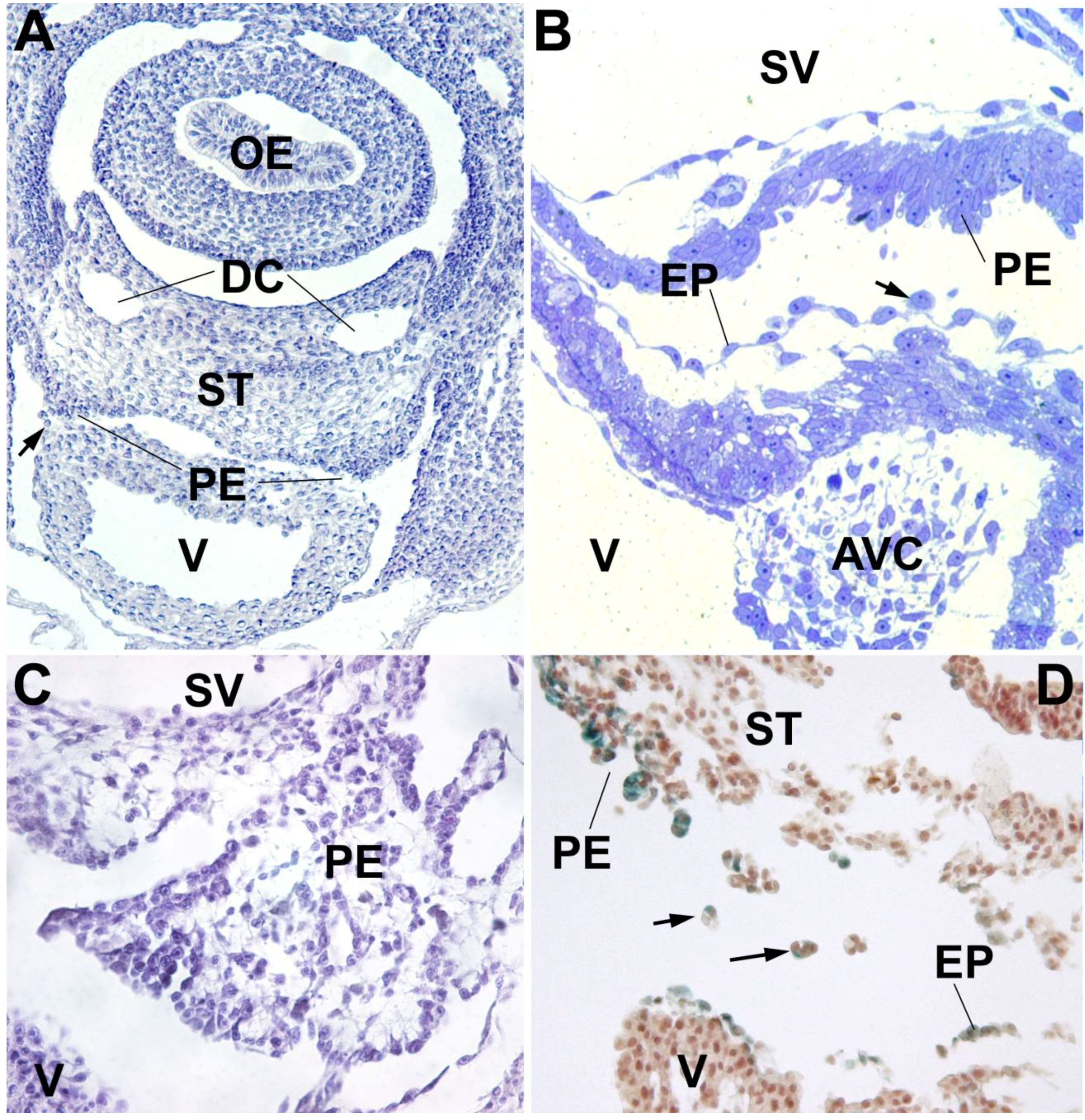
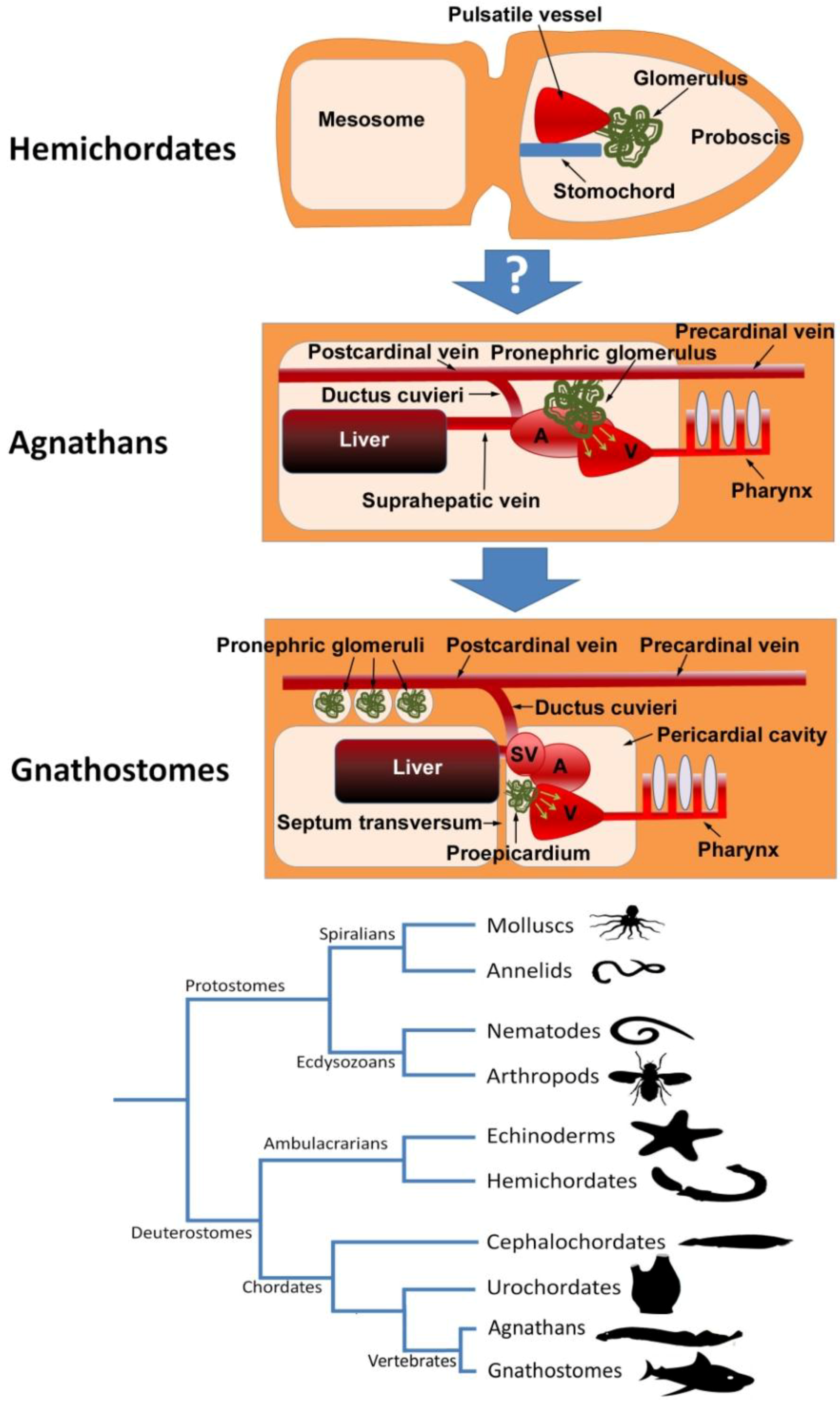
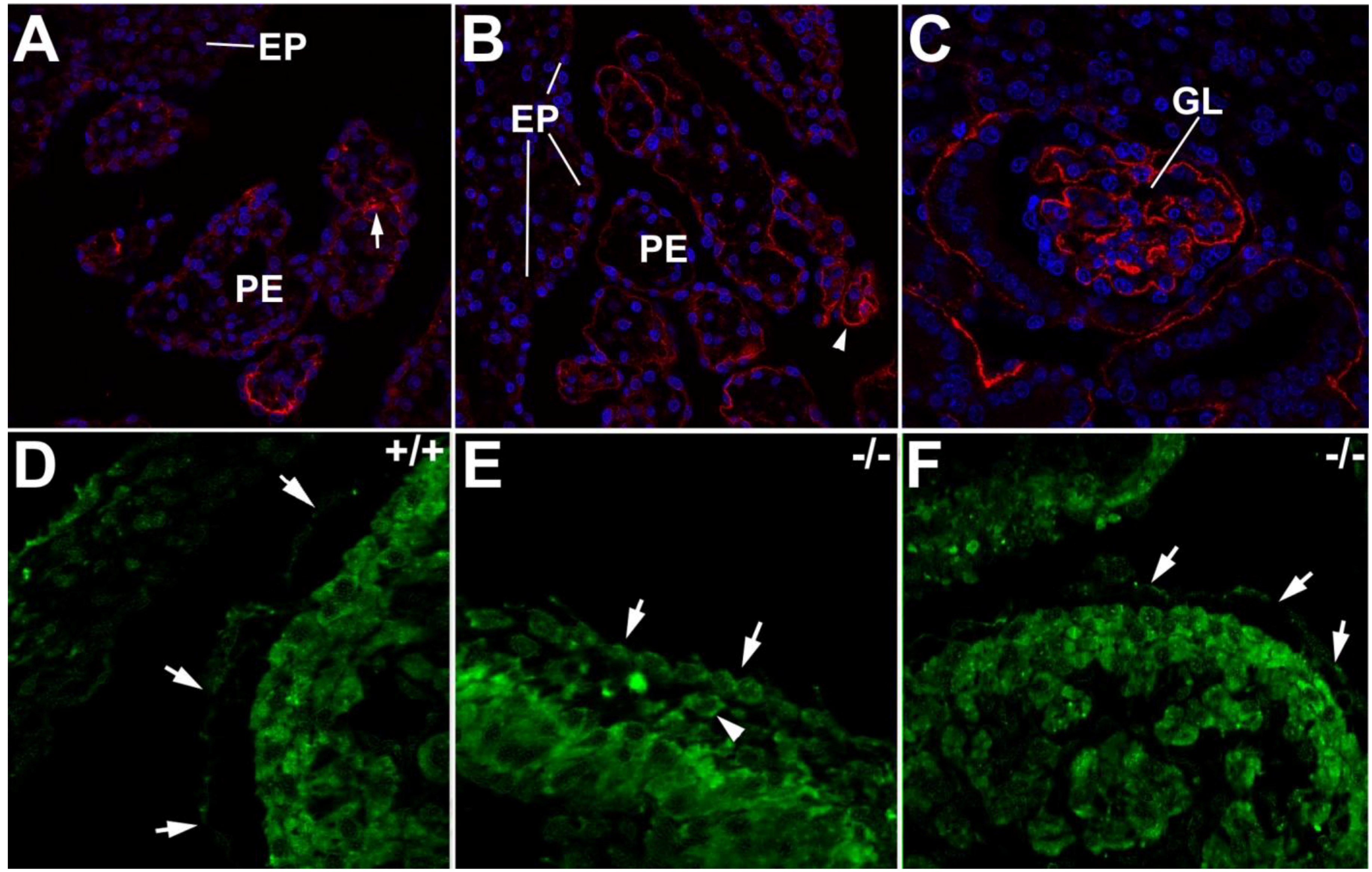
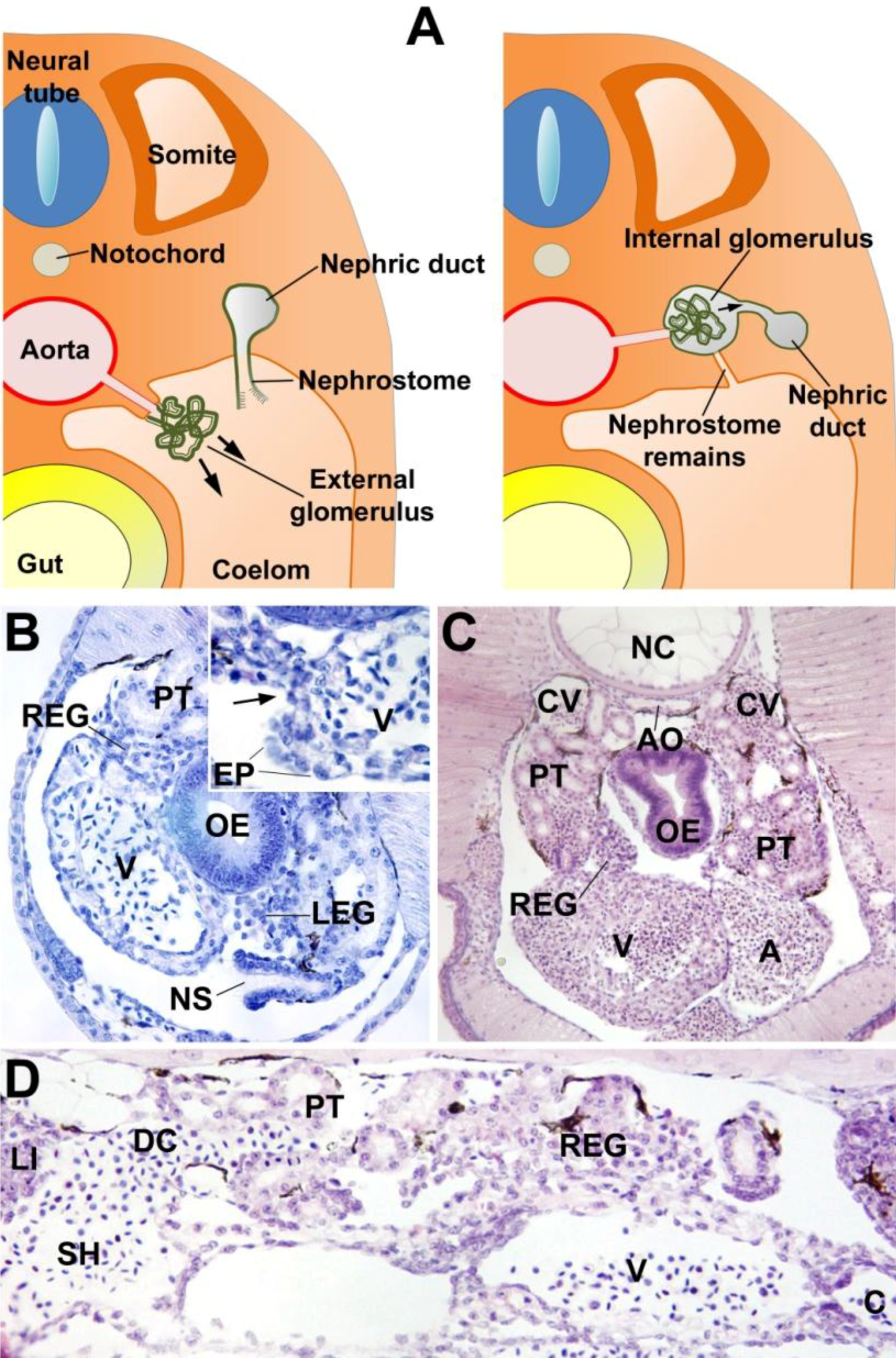
3. The Proepicardium as the Evolutionary Remain of a Pronephric External Glomerulus
4. Gene Expression in the Proepicardium. Comparison with Kidney Development
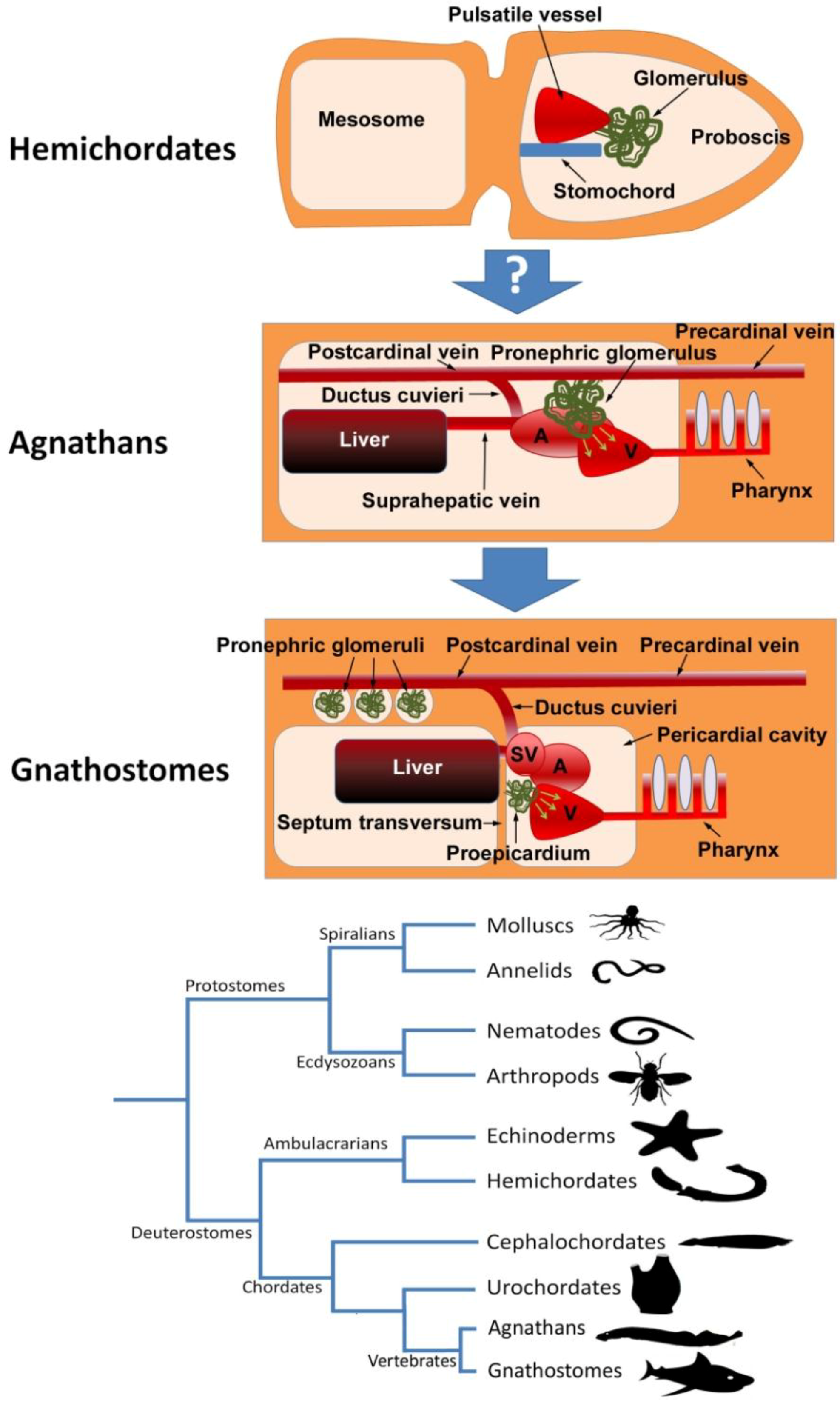
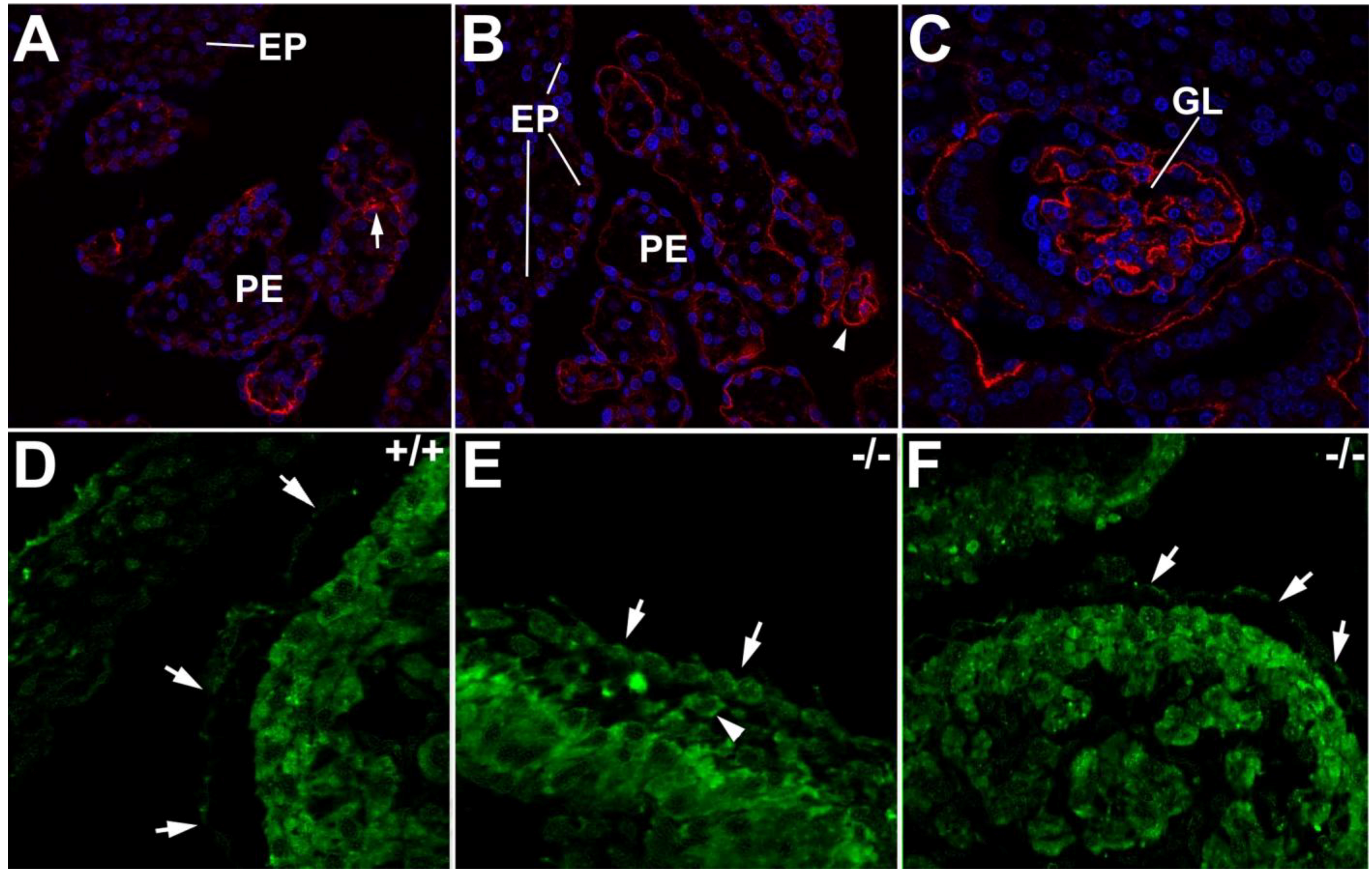
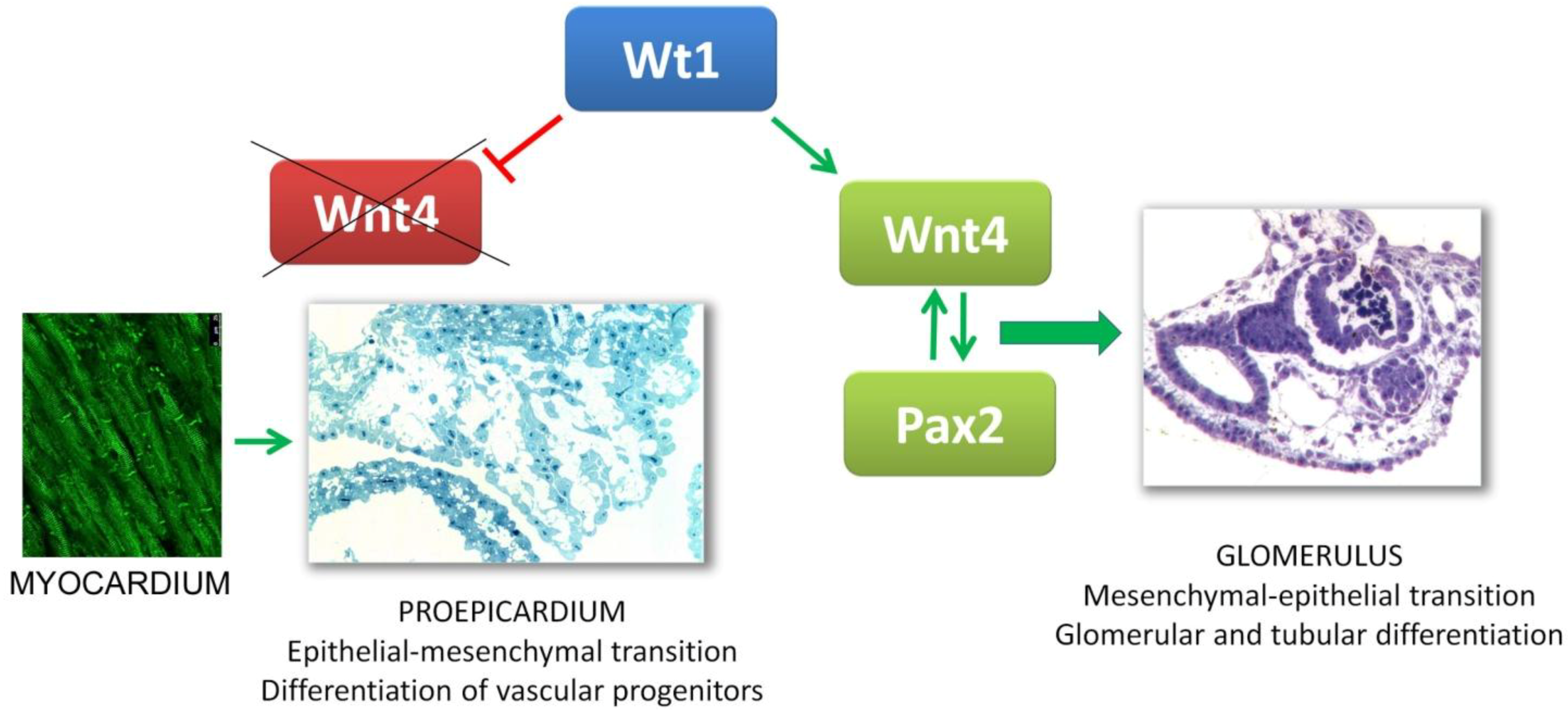
5. The Proepicardium and the Heart-Kidney Complex
6. Proepicardium, Kidney and Hematopoiesis
7. Concluding Remarks
Acknowledgments
Conflict of Interest
References and Notes
- Manasek, F.J. Embryonic development of the heart. I. A light and electron microscopic study of myocardial development in the early chick embryo. J. Morphol. 1968, 125, 329–365. [Google Scholar] [CrossRef]
- Manasek, F.J. Embryonic development of the heart. II. Formation of the epicardium. J. Embryol. Exp. Morphol. 1969, 22, 333–348. [Google Scholar]
- His, W. Anatomie menschlicher Embryonen. Teil III Zur Geschichte der Organe; Vogel: Leipzig, Germany, 1885. [Google Scholar]
- Kurkiewicz, T. O histogenezie miesna sercowego zwierzat kregowych - Zur Histogenese des Herzmuskels der Wirbeltiere. Bul. Int. Acad. Sci. Cracovie 1909, 177, 148–191. [Google Scholar]
- Männer, J.; Pérez-Pomares, J.M.; Macías, D.; Muñoz-Chápuli, R. The origin, formation, and developmental significance of the epicardium: a review. Cell Tiss. Org. 2001, 169, 89–103. [Google Scholar] [CrossRef]
- Budelmann, B.U.; Schipp, R.; Von Boletzky, S. Cephalopoda. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Kohn, A.J., Eds.; Wiley-Liss: New York, NY, USA, 1997; Volume 6A, pp. 119–414. [Google Scholar]
- Pombal, M.A.; Carmona, R.; Megías, M.; Ruiz, A.; Pérez-Pomares, J.M.; Muñoz-Chápuli, R. Epicardial development in lamprey supports an evolutionary origin of the vertebrate epicardium from an ancestral pronephric external glomerulus. Evol. Dev. 2008, 10, 210–216. [Google Scholar] [CrossRef]
- Jahr, M.; Schlueter, J.; Brand, T.; Männer, J. Development of the proepicardium in Xenopus laevis. Dev. Dyn. 2008, 237, 3088–3096. [Google Scholar] [CrossRef]
- Schlueter, J.; Brand, T. A right-sided pathway involving FGF8/Snai1 controls asymmetric development of the proepicardium in the chick embryo. Proc. Natl. Acad. Sci. USA 2009, 106, 7485–7490. [Google Scholar] [CrossRef]
- Schulte, I.; Schlueter, J.; Abu-Issa, R.; Brand, T.; Manner, J. Morphological and molecular left-right asymmetries in the development of the proepicardium: A comparative analysis on mouse and chick embryos. Dev. Dyn. 2007, 236, 684–695. [Google Scholar] [CrossRef]
- Muñoz-Chápuli, R.; Macías, D.; Ramos, C.; De Andrés, A.V.; Gallego, A.; Navarro, P. Heart development in the dogfish (Scyliorhinus canicula): A model for the study of the basic vertebrate cardiogenesis. Cardioscience 1994, 5, 245–253. [Google Scholar]
- Muñoz-Chápuli, R.; Macías, D.; Ramos, C.; Fernández, B.; Sans-Coma, V. Development of the epicardium in the dogfish (Scyliorhinus canicula). Acta Zool. 1997, 78, 39–46. [Google Scholar] [CrossRef]
- Icardo, J.M.; Guerrero, A.; Durán, A.C.; Colvee, E.; Domezain, A.; Sans-Coma, V. The development of the epicardium in the sturgeon Acipenser naccarii. Anat. Rec. 2009, 292, 1593–1601. [Google Scholar] [CrossRef]
- Serluca, F.C. Development of the proepicardial organ in the zebrafish. Dev. Biol. 2008, 315, 18–27. [Google Scholar] [CrossRef]
- Fransen, M.E.; Lemanski, L.F. Epicardial development in the axolotl, Ambystoma mexicanum. Anat. Rec. 1990, 226, 228–236. [Google Scholar] [CrossRef]
- Sejima, H.; Isokawa, K.; Shimizu, O.; Morikawa, T.; Ootsu, H.; Numata, K.; Fukai, M.; Kubota, S.; Toda, Y. Possible participation of isolated epicardial cell clusters in the formation of chick embryonic epicardium. J. Oral Sci. 2001, 43, 109–106. [Google Scholar] [CrossRef]
- Komiyama, M.; Ito, K.; Shimada, Y. Origin and development of the epicardium in the mouse embryo. Anat. Embryol. 1987, 176, 183–189. [Google Scholar] [CrossRef]
- Kuhn, H.J.; Liebherr, G. The early development of the epicardium in Tupaja belangerie. Anat. Embryol. 1988, 177, 225–234. [Google Scholar] [CrossRef]
- Männer, J. The development of pericardial villi in the chick embryo. Anat. Embryol. 1992, 186, 379–385. [Google Scholar] [CrossRef]
- Männer, J. Experimental study on the formation of the epicardium in chick embryos. Anat Embryol. 1993, 187, 281–289. [Google Scholar] [CrossRef]
- MacKinnon, M.R.; Heatwole, H. Comparative cardiac anatomy of the reptilia. IV. The coronary arterial circulation. J. Morphol. 1981, 170, 1–27. [Google Scholar] [CrossRef]
- Davies, F.; Francis, E.T.B.; King, T.S. The conducting (connecting) system of the crocodilian heart. J. Anat. 1952, 86, 152–161. [Google Scholar]
- Buchanan, J.G. The Gross and Minute Anatomy of the Heart of the Lizard, Leiolopisma grande (Gray). Trans. Proc. Royal Soc. N. Z. 1956, 84, 103–119. [Google Scholar]
- Viragh, S.; Gittenberger-de Groot, A.C.; Poelmann, R.E.; Kalman, F. Early development of quail heart epicardium and associated vascular and glandular structures. Anat. Embryol. 1993, 188, 381–393. [Google Scholar]
- Virágh, S.; Challice, C.E. The origin of the epicardium and the embryonic myocardial circulation in the mouse. Anat. Rec. 1981, 201, 157–168. [Google Scholar] [CrossRef]
- Brandli, A.W. Towards a molecular anatomy of the Xenopus pronephric kidney. Int. J. Dev. Biol. 1999, 43, 381–395. [Google Scholar]
- Jones, E.A. Xenopus: A prince among models for pronephric kidney development. J. Am. Soc. Nephrol. 2005, 16, 313–321. [Google Scholar] [CrossRef]
- Hiruma, T.; Nakamura, H. Origin and development of the pronephros in the chick embryo. J. Anat. 2003, 203, 539–552. [Google Scholar] [CrossRef]
- Stuckmann, I.; Evans, S.; Lassar, A.B. Erythropoietin and retinoic acid, secreted from the epicardium, are required for cardiac myocyte proliferation. Dev. Biol. 2003, 255, 334–349. [Google Scholar] [CrossRef]
- Lavine, K.J.; Yu, K.; White, A.C.; Zhang, X.; Smith, C.; Partanen, J.; Ornitz, D.M. Endocardial and epicardial derived FGF signals regulate myocardial proliferation and differentiation in vivo. Dev. Cell. 2005, 8, 85–95. [Google Scholar] [CrossRef]
- Merki, E.; Zamora, M.; Raya, A.; Kawakami, Y.; Wang, J.; Zhang, X.; Burch, J.; Kubalak, S.W.; Kaliman, P.; Izpisúa-Belmonte, J.C.; Chien, K.R.; Ruiz-Lozano, P. Epicardial retinoid X receptor alpha is required for myocardial growth and coronary artery formation. Proc. Natl. Acad. Sci. USA 2005, 102, 18455–18460. [Google Scholar] [CrossRef]
- Martínez-Estrada, O.M.; Lettice, L.A.; Essafi, A.; Guadix, J.A.; Slight, J.; Velecela, V.; Hall, E.; Reichmann, J.; Devenney, P.S.; Hohenstein, P.; Hosen, N.; Hill, R.E.; Muñoz-Chapuli, R.; Hastie, N.D. Wt1 is required for mesenchymal cardiovascular progenitor cell formation in epicardium and ES cells through direct transcriptional control of Snail and E-cadherin. Nat. Genet. 2010, 42, 89–93. [Google Scholar] [CrossRef]
- Drummond, I.A.; Majumdar, A.; Hentschel, H.; Elger, M.; Solnica-Krezel, L.; Schier, A.F.; Neuhauss, S.C.; Stemple, D.L.; Zwartkruis, F.; Rangini, Z.; Driever, W.; Fishman, M.C. Early development of the zebrafish pronephros and analysis of mutations affecting pronephric function. Development 1998, 125, 4655–4667. [Google Scholar]
- Moore, A.W.; McInnes, L.; Kreidberg, J.; Hastie, N.D.; Schedl, A. YAC complementation shows a requirement for Wt1 in the development of epicardium, adrenal gland and throughout nephrogenesis. Development 1999, 126, 1845–1857. [Google Scholar]
- Quaggin, S.E.; Schwartz, L.; Cui, S.; Igarashi, P.; Deimling, J.; Post, M.; Rossant, J. The basic-helix-loop-helix protein pod1 is critically important for kidney and lung organogenesis. Development 1999, 126, 5771–5783. [Google Scholar]
- Cui, S.; Schwartz, L.; Quaggin, S.E. Pod1 is required in stromal cells for glomerulogenesis. Dev. Dyn. 2003, 226, 512–522. [Google Scholar] [CrossRef]
- Acharya, A.; Baek, S.T.; Huang, G.; Eskiocak, B.; Goetsch, S.; Sung, C.Y.; Banfi, S.; Sauer, M.F.; Olsen, G.S.; Duffield, J.S.; Olson, E.N.; Tallquist, M.D. The bHLH transcription factor Tcf21 is required for lineage-specific EMT of cardiac fibroblast progenitors. Development 2012, 139, 2139–2149. [Google Scholar] [CrossRef]
- Christoffels, V.M.; Mommersteeg, M.T.; Trowe, M.O.; Prall, O.W.; de Gier-de Vries, C.; Soufan, A.T.; Bussen, M.; Schuster-Gossler, K.; Harvey, R.P.; Moorman, A.F.; Kispert, A. Formation of the venous pole of the heart from an Nkx2–5-negative precursor population requires Tbx18. Circ. Res. 2006, 98, 1555–1563. [Google Scholar] [CrossRef]
- Takeichi, M.; Nimura, K.; Mori, M.; Nakagami, H.; Kaneda, Y. The transcription factors Tbx18 and Wt1 control the epicardial epithelial-mesenchymal transition through bi-directional regulation of Slug in murine primary epicardial cells. PLoS One 2013, 8, e57829. [Google Scholar]
- Putaala, H.; Soininen, R.; Kilpelainen, P.; Wartiovaara, J.; Tryggvason, K. The murine nephrin gene is specifically expressed in kidney, brain and pancreas: inactivation of the gene leads to massive proteinuria and neonatal death. Hum. Mol. Genet. 2001, 10, 1–8. [Google Scholar] [CrossRef]
- Wagner, N.; Morrison, H.; Pagnotta, S.; Michiels, J.F.; Schwab, Y.; Tryggvason, K.; Schedl, A.; Wagner, K.D. The podocyte protein nephrin is required for cardiac vessel formation. Hum. Mol. Genet. 2011, 20, 2182–2194. [Google Scholar] [CrossRef]
- Wagner, N.; Wagner, K.D.; Scholz, H.; Kirschner, K.M.; Schedl, A. Intermediate filament protein nestin is expressed in developing kidney and heart and might be regulated by the Wilms' tumor suppressor Wt1. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R779–R787. [Google Scholar] [CrossRef]
- Wharram, B.L.; Goyal, M.; Gillespie, P.J.; Wiggins, J.E.; Kershaw, D.B.; Holzman, L.B.; Dysko, R.C.; Saunders, T.L.; Samuelson, L.C.; Wiggins, R.C. Altered podocyte structure in GLEPP1 (Ptpro)-deficient mice associated with hypertension and low glomerular filtration rate. J. Clin. Invest. 2000, 106, 1281–1290. [Google Scholar] [CrossRef]
- Osafune, K.; Nishinakamura, R.; Komazaki, S.; Asashima, M. In vitro induction of the pronephric duct in Xenopus explants. Develop. Growth Differ. 2002, 44, 161–167. [Google Scholar] [CrossRef]
- Olivey, H.E.; Mundell, N.A.; Austin, A.F.; Barnett, J.V. Transforming growth factor-β stimulates epithelial-mesenchymal transformation in the proepicardium. Dev. Dyn. 2006, 235, 50–59. [Google Scholar] [CrossRef]
- Jenkins, S.J.; Hutson, D.R.; Kubalak, S.W. Analysis of the proepicardium-epicardium transition during the malformation of the RXRalpha-/- epicardium. Dev. Dyn. 2005, 233, 1091–1101. [Google Scholar] [CrossRef]
- Vaughan, M.R.; Pippin, J.W.; Griffin, S.V.; Krofft, R.; Fleet, M.; Haseley, L.; Shankland, S.J. ATRA induces podocyte differentiation and alters nephrin and podocin expression in vitro and in vivo. Kidney Int. 2005, 68, 133–144. [Google Scholar] [CrossRef]
- Essafi, A.; Webb, A.; Berry, R.L.; Slight, J.; Burn, S.F.; Spraggon, L.; Velecela, V.; Martinez-Estrada, O.M.; Wiltshire, J.H.; Roberts, S.G.; Brownstein, D.; Davies, J.A.; Hastie, N.D.; Hohenstein, P. A wt1-controlled chromatin switching mechanism underpins tissue-specific wnt4 activation and repression. Dev. Cell. 2011, 21, 559–574. [Google Scholar] [CrossRef]
- Bouchard, M.; Souabni, A.; Mandler, M.; Neubüser, A.; Busslinger, M. Nephric lineage specification by Pax2 and Pax8. Genes Dev. 2002, 16, 2958–2970. [Google Scholar] [CrossRef]
- Torban, E.; Dziarmaga, A.; Iglesias, D.; Chu, L.L.; Vassilieva, T.; Little, M.; Eccles, M.; Discenza, M.; Pelletier, J.; Goodyer, P. PAX2 activates WNT4 expression during mammalian kidney development. J. Biol. Chem. 2006, 281, 12705–12712. [Google Scholar] [CrossRef]
- Dressler, G.R. Patterning and early cell lineage decisions in the developing kidney: The role of Pax genes. Pediatr. Nephrol. 2011, 26, 1387–1394. [Google Scholar] [CrossRef]
- Phelps, D.E.; Dressler, G.R. Aberrant expression of Pax2 in Danforth's short tail (Sd) mice. Dev. Biol. 1993, 157, 251–258. [Google Scholar] [CrossRef]
- Ryan, G.; Steele-Perkins, V.; Morris, J.F.; Rauscher, F.J., 3rd; Dressler, G.R. Repression of Pax2 by WT1 during normal kidney development. Development 1995, 121, 867–875. [Google Scholar]
- Watt, A.J.; Battle, M.A.; Li, J.; Duncan, S.A. GATA4 is essential for formation of the proepicardium and regulates cardiogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 12573–12578. [Google Scholar]
- Dame, C.; Sola, M.C.; Lim, K.C.; Leach, K.M.; Fandrey, J.; Ma, Y.; Knöpfle, G.; Engel, J.D.; Bungert, J. Hepatic erythropoietin gene regulation by GATA-4. J. Biol. Chem. 2004, 279, 2955–2961. [Google Scholar]
- Zhou, B.; von Gise, A.; Ma, Q.; Rivera-Feliciano, J.; Pu, W.T. Nkx2–5- and Isl1-expressing cardiac progenitors contribute to proepicardium. Biochem. Biophys. Res. Commun. 2008, 375, 450–453. [Google Scholar] [CrossRef]
- Holmgren, N. On the pronephros and the blood in Myxine glutinosa. Acta Zool. 1950, 31, 233–348. [Google Scholar] [CrossRef]
- Fänge, R. Structure and function of the excretory organs of Myxinoids. In The Biology of Myxine; Brodal, A., Fänge, R., Eds.; Universitetforlaget: Oslo, Norway, 1963; pp. 516–529. [Google Scholar]
- Lowe, C.J.; Terasaki, M.; Wu, M.; Freeman, R.M., Jr.; Runft, L.; Kwan, K.; Haigo, S.; Aronowicz, J.; Lander, E.; Gruber, C.; Smith, M.; Kirschner, M.; Gerhart, J. Dorsoventral patterning in hemichordates: insights into early chordate evolution. PLoS Biol. 2006, 4, e291. [Google Scholar] [CrossRef]
- Pascual-Anaya, J.; Albuixech-Crespo, B.; Somorjai, I.M.; Carmona, R.; Oisi, Y.; Alvarez, S.; Kuratani, S.; Muñoz-Chápuli, R.; Garcia-Fernández, J. The evolutionary origins of chordate hematopoiesis and vertebrate endothelia. Dev. Biol. 2013, 375, 182–192. [Google Scholar] [CrossRef]
- Hirakow, R. Epicardial formation in staged human embryos. Acta Anat. Nippon. 1992, 67, 616–622. [Google Scholar]
- Kattan, J.; Dettman, R.W.; Bristow, J. Formation and remodeling of the coronary vascular bed in the embryonic avian heart. Dev. Dyn. 2004, 230, 34–43. [Google Scholar] [CrossRef]
- Tomanek, R.J.; Ishii, Y.; Holifield, J.S.; Sjogren, C.L.; Hansen, H.K.; Mikawa, T. VEGF family members regulate myocardial tubulogenesis and coronary artery formation in the embryo. Circ. Res. 2006, 98, 947–953. [Google Scholar] [CrossRef]
- Wilting, J.; Buttler, K.; Schulte, I.; Papoutsi, M.; Schweigerer, L.; Manner, J. The proepicardium delivers hemangioblasts but not lymphangioblasts to the developing heart. Dev. Biol. 2007, 305, 451–459. [Google Scholar] [CrossRef]
- Carpenter, K.L.; Turpen, J.B. Experimental studies on hemopoiesis in the pronephros of Rana pipiens. Differentiation 1979, 14, 167–174. [Google Scholar] [CrossRef]
- Willett, C.E.; Cortés, A.; Zuasti, A.; Zapata, A.G. Early hematopoiesis and developing lymphoid organs in the zebrafish. Dev. Dyn. 1999, 214, 323–336. [Google Scholar] [CrossRef]
- Luchtel, L.L.; Martin, A.W.; Deyrup-Olsen, I.; Boer, H.H. Gastropoda: Pulmonata. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Kohn, A.J., Eds.; Wiley-Liss: New York, NY, USA, 1997; Volume 6B, pp. 459–718. [Google Scholar]
- Økland, S. The heart ultrastructure of Lepidopleurus asellus (Spengler) and Tonicella marmorea (Fabricius) (Mollusca: Polyplacophora). Zoomorphology 1980, 96, 1–19. [Google Scholar] [CrossRef]
- Fahrner, A.; Haszprunar, G. Microanatomy, ultrastructure, and systematic significance of the excretory system and mantle cavity of an acochlidian gastropod (Opisthobranchia). J. Moll. Stud. 2002, 68, 87–94. [Google Scholar] [CrossRef]
- Denholm, B.; Skaer, H. Bringing together components of the fly renal system. Curr. Opin. Genet. Dev. 2009, 19, 526–532. [Google Scholar] [CrossRef]
- Mandal, L.; Banerjee, U.; Hartenstein, V. Evidence for a fruit fly hemangioblast and similarities between lymph-gland hematopoiesis in fruit fly and mammal aorta-gonadal-mesonephros mesoderm. Nat. Genet. 2004, 36, 1019–1023. [Google Scholar] [CrossRef]
- Ishii, Y.; Langberg, J.D.; Hurtado, R.; Lee, S.; Mikawa, T. Induction of proepicardial marker gene expression by the liver bud. Development 2007, 134, 3627–3637. [Google Scholar] [CrossRef]
- Liu, J.; Stainier, D.Y. Tbx5 and Bmp signaling are essential for proepicardium specification in zebrafish. Circ. Res. 2010, 106, 1818–1828. [Google Scholar] [CrossRef]
- Wagner, K.D.; Wagner, N.; Bondke, A.; Nafz, B.; Flemming, B.; Theres, H.; Scholz, H. The Wilms' tumor suppressor Wt1 is expressed in the coronary vasculature after myocardial infarction. FASEB J. 2002, 16, 1117–1119. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cano, E.; Carmona, R.; Muñoz-Chápuli, R. Evolutionary Origin of the Proepicardium. J. Dev. Biol. 2013, 1, 3-19. https://doi.org/10.3390/jdb1010003
Cano E, Carmona R, Muñoz-Chápuli R. Evolutionary Origin of the Proepicardium. Journal of Developmental Biology. 2013; 1(1):3-19. https://doi.org/10.3390/jdb1010003
Chicago/Turabian StyleCano, Elena, Rita Carmona, and Ramón Muñoz-Chápuli. 2013. "Evolutionary Origin of the Proepicardium" Journal of Developmental Biology 1, no. 1: 3-19. https://doi.org/10.3390/jdb1010003