Thimet Oligopeptidase (EC 3.4.24.15) Key Functions Suggested by Knockout Mice Phenotype Characterization

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
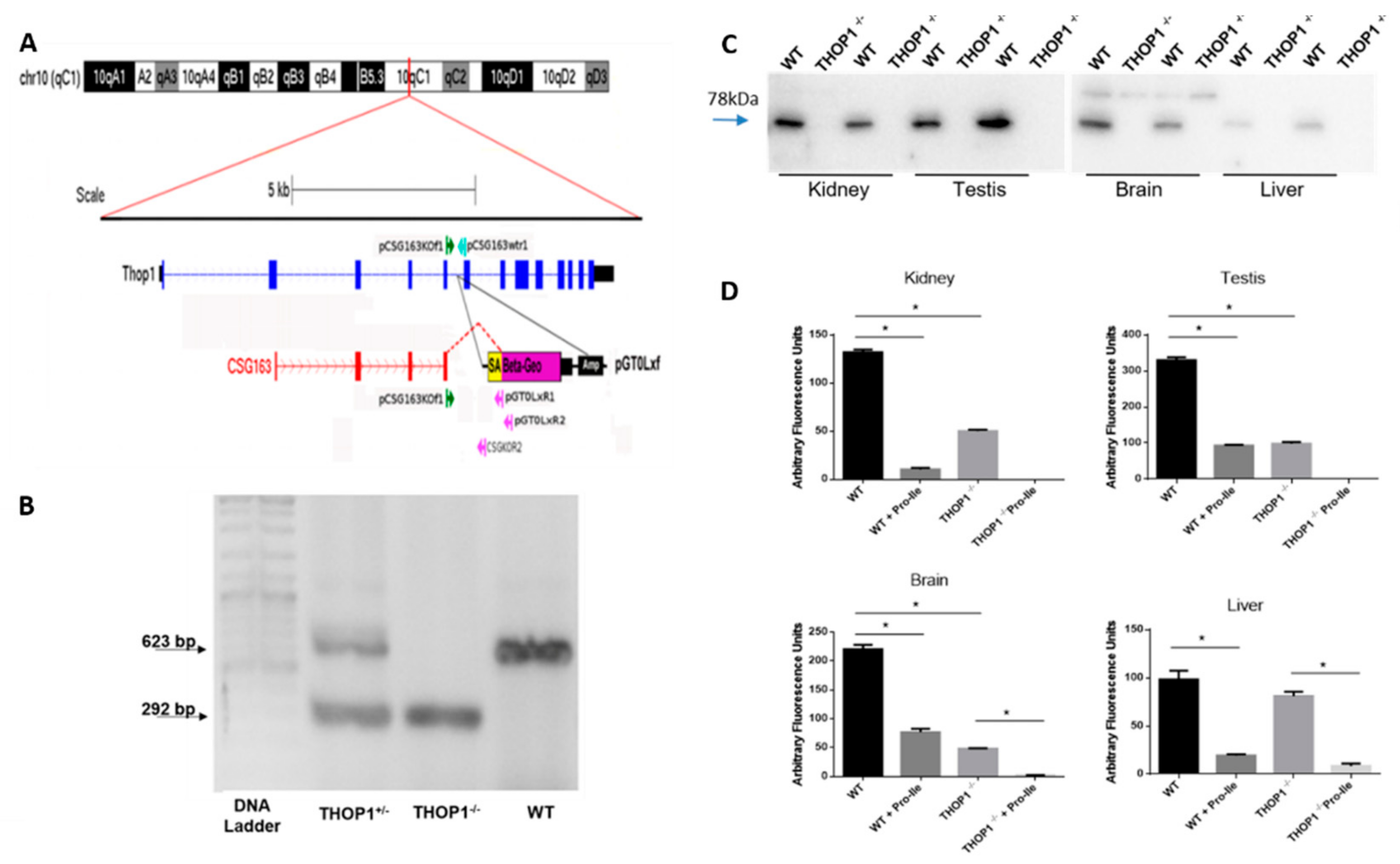
2.1. THOP1-/- Generation
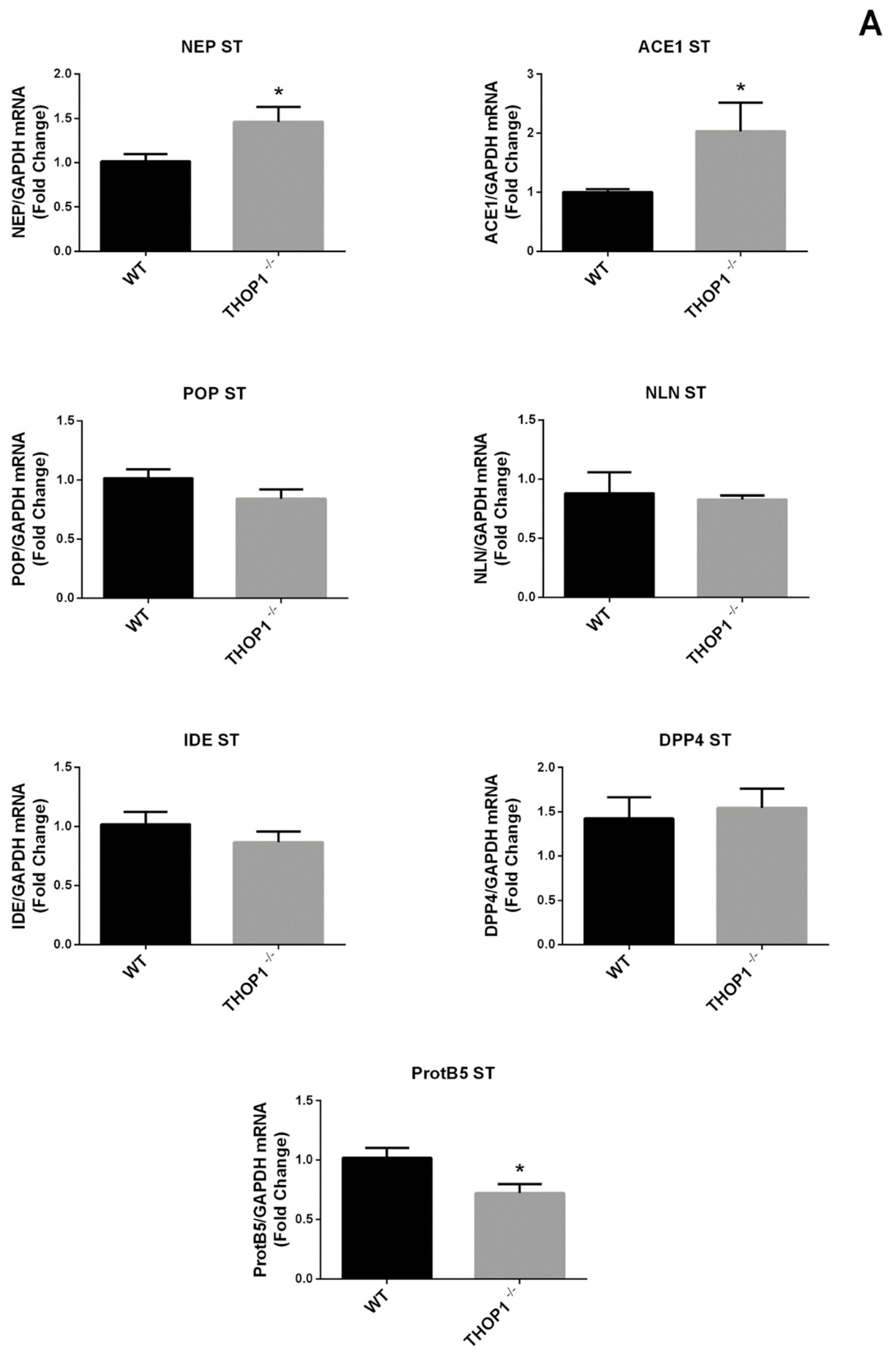
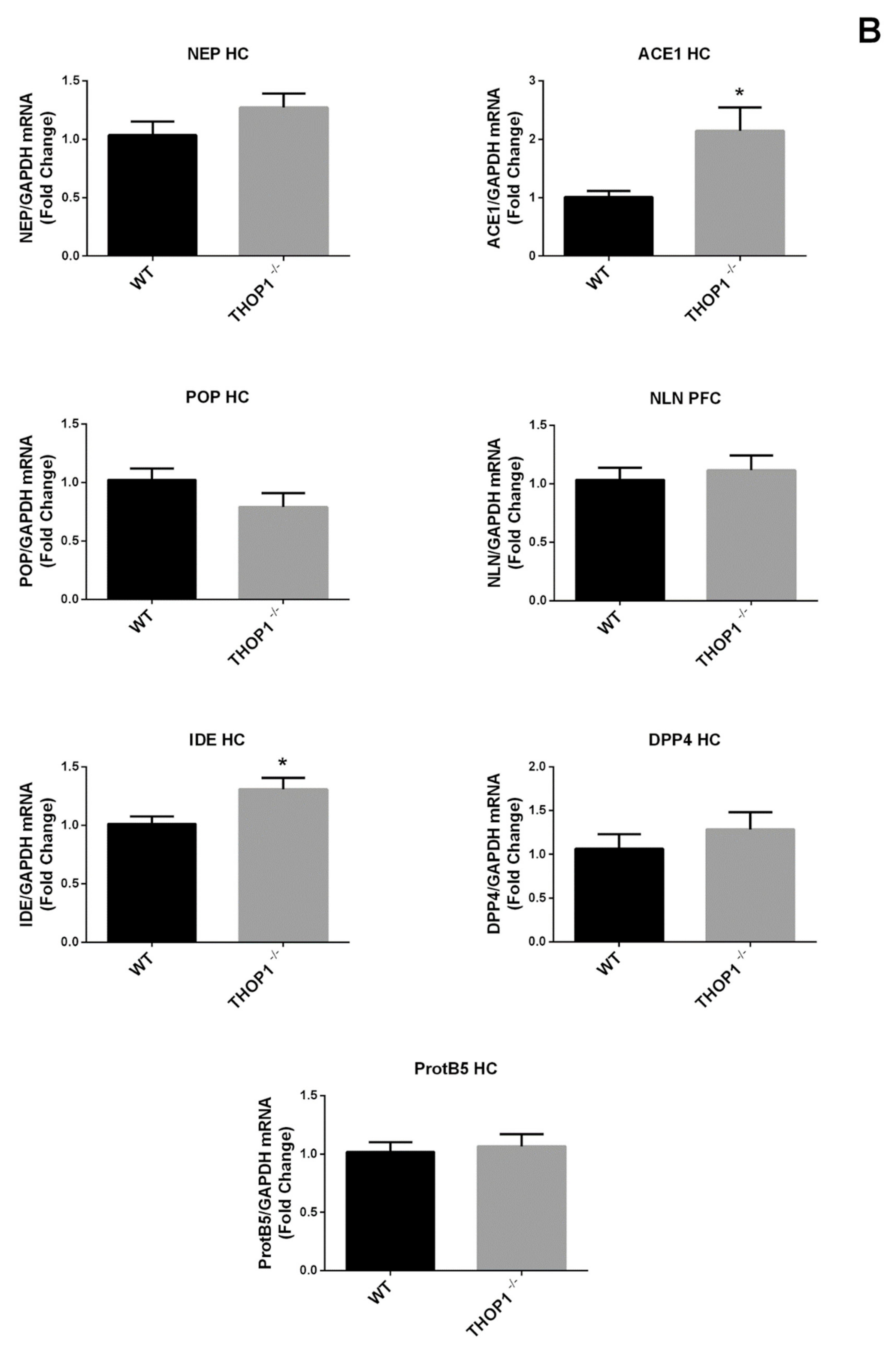
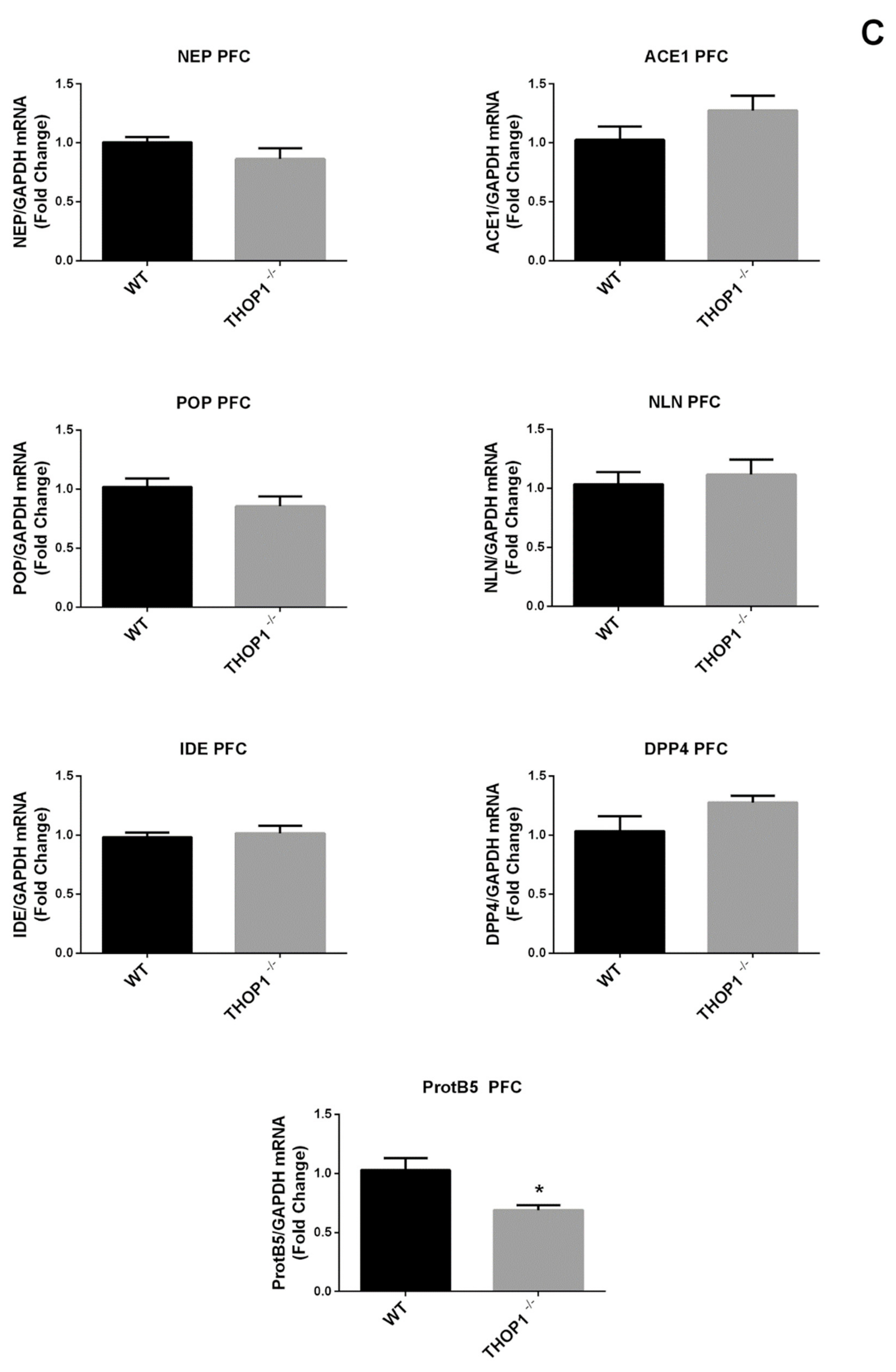
2.2. Global Gene Expression Analyses
2.3. Peptidomics
2.4. Complete Blood Count and Coagulation Analysis
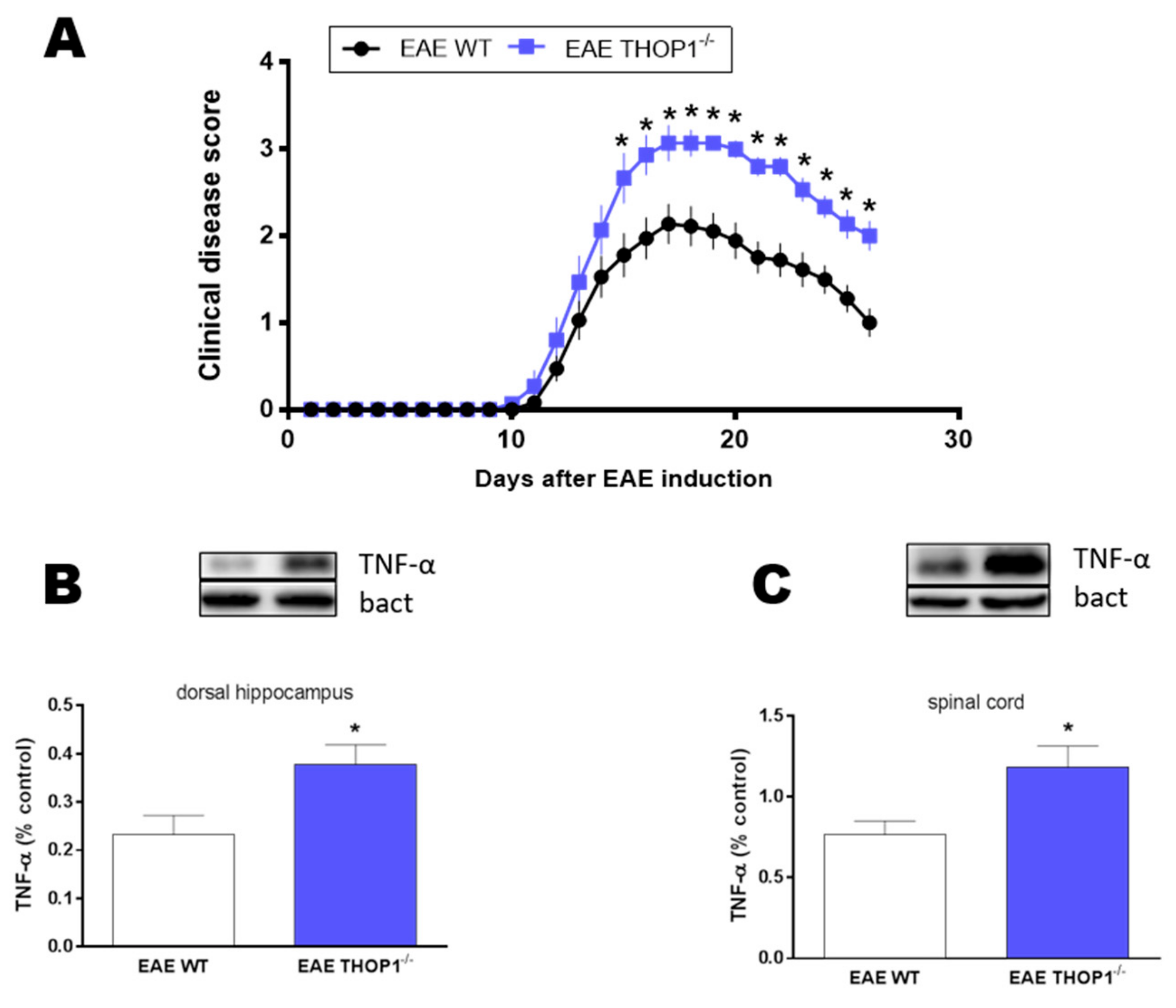
2.5. Autoimmune Encephalomyelitis Neurodegeneration Model
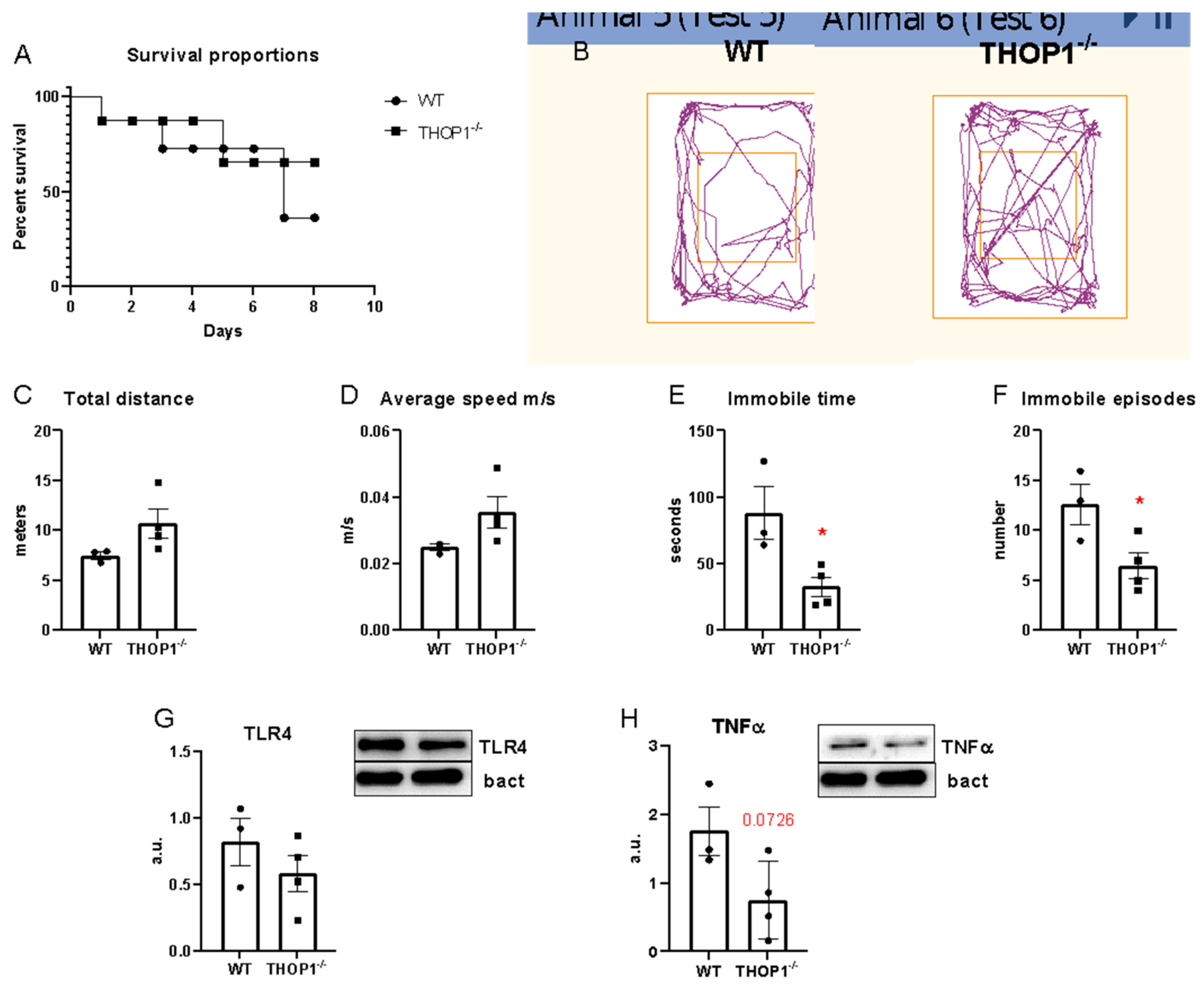
2.6. Sepsis
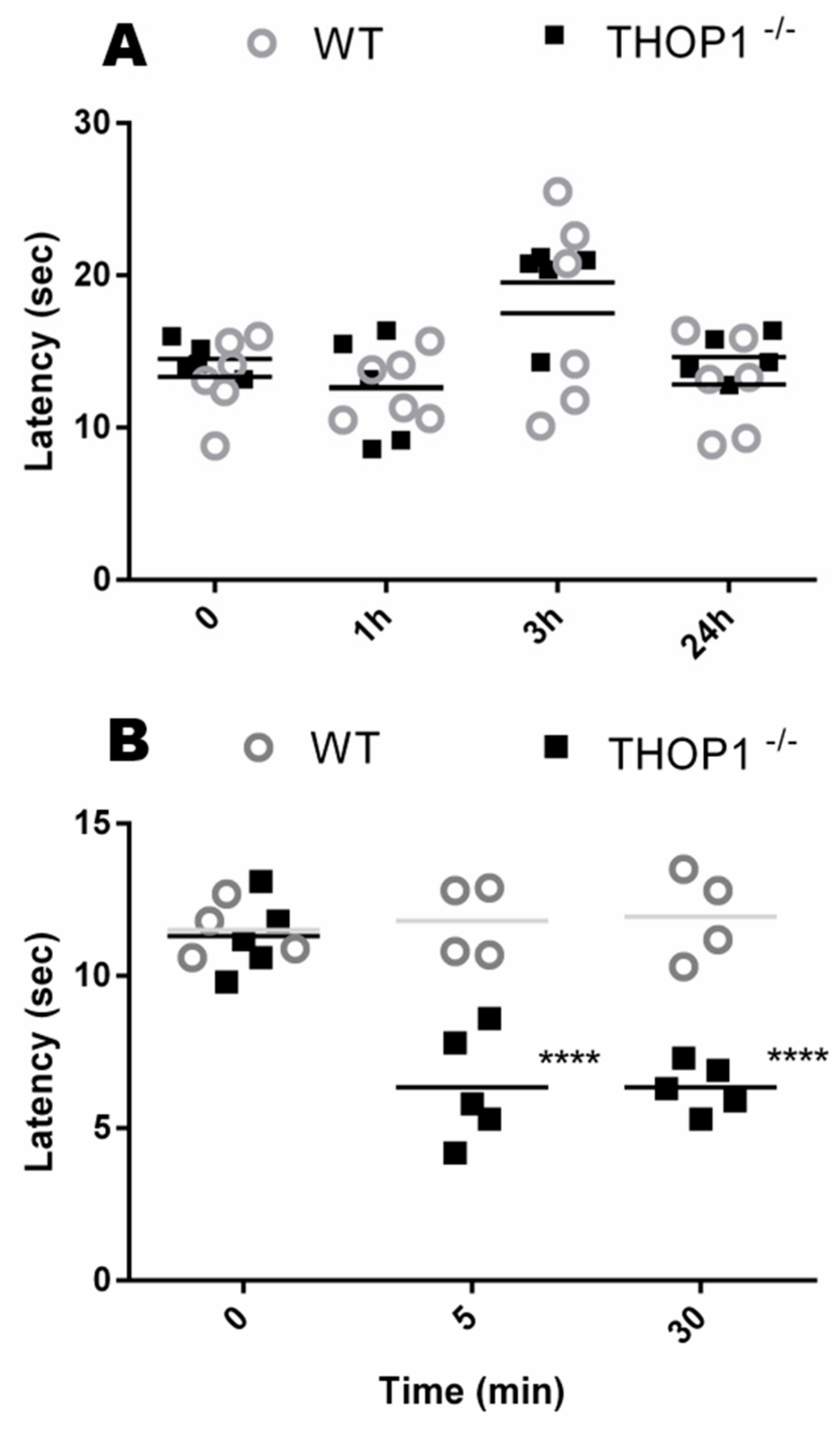
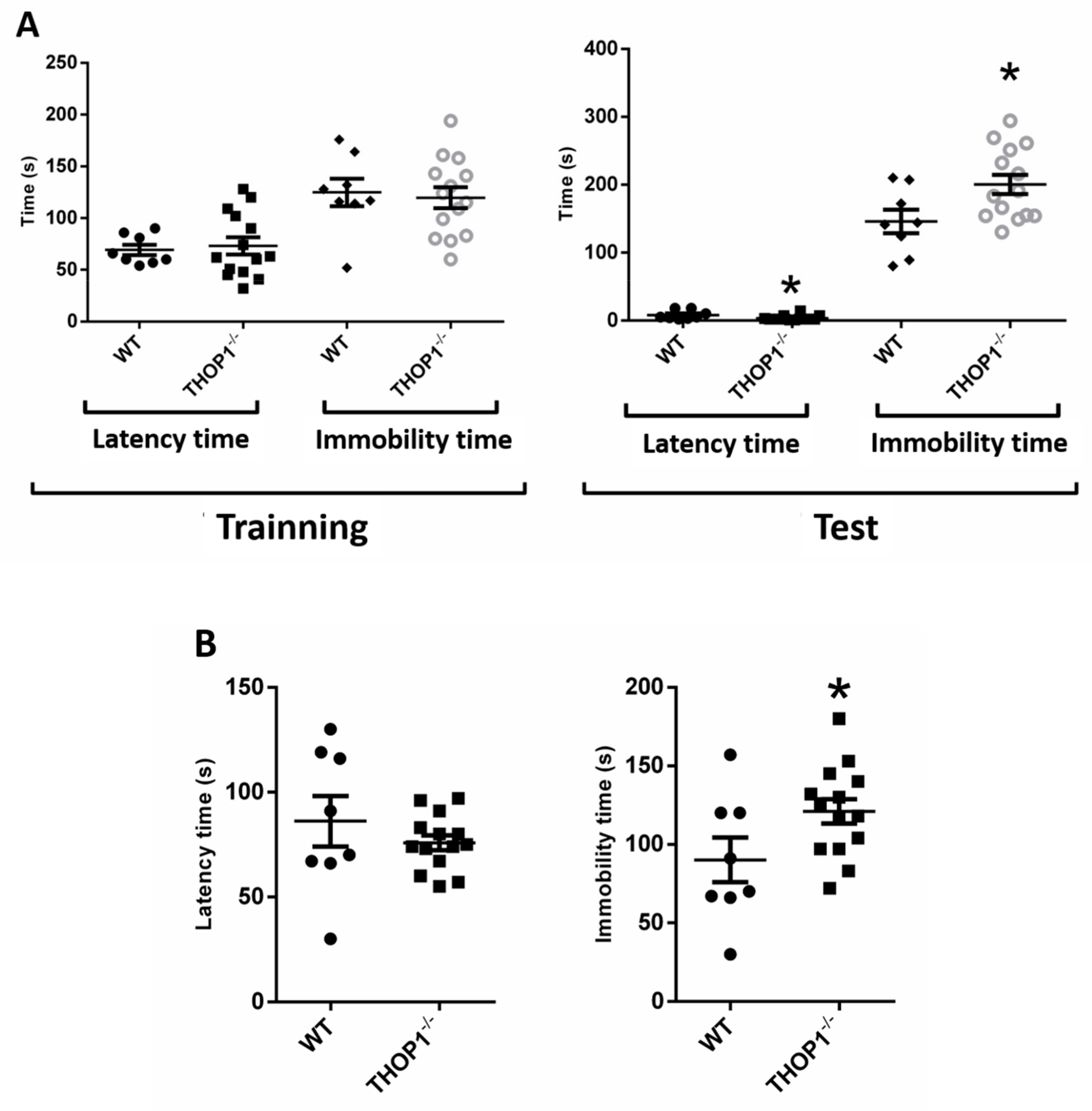
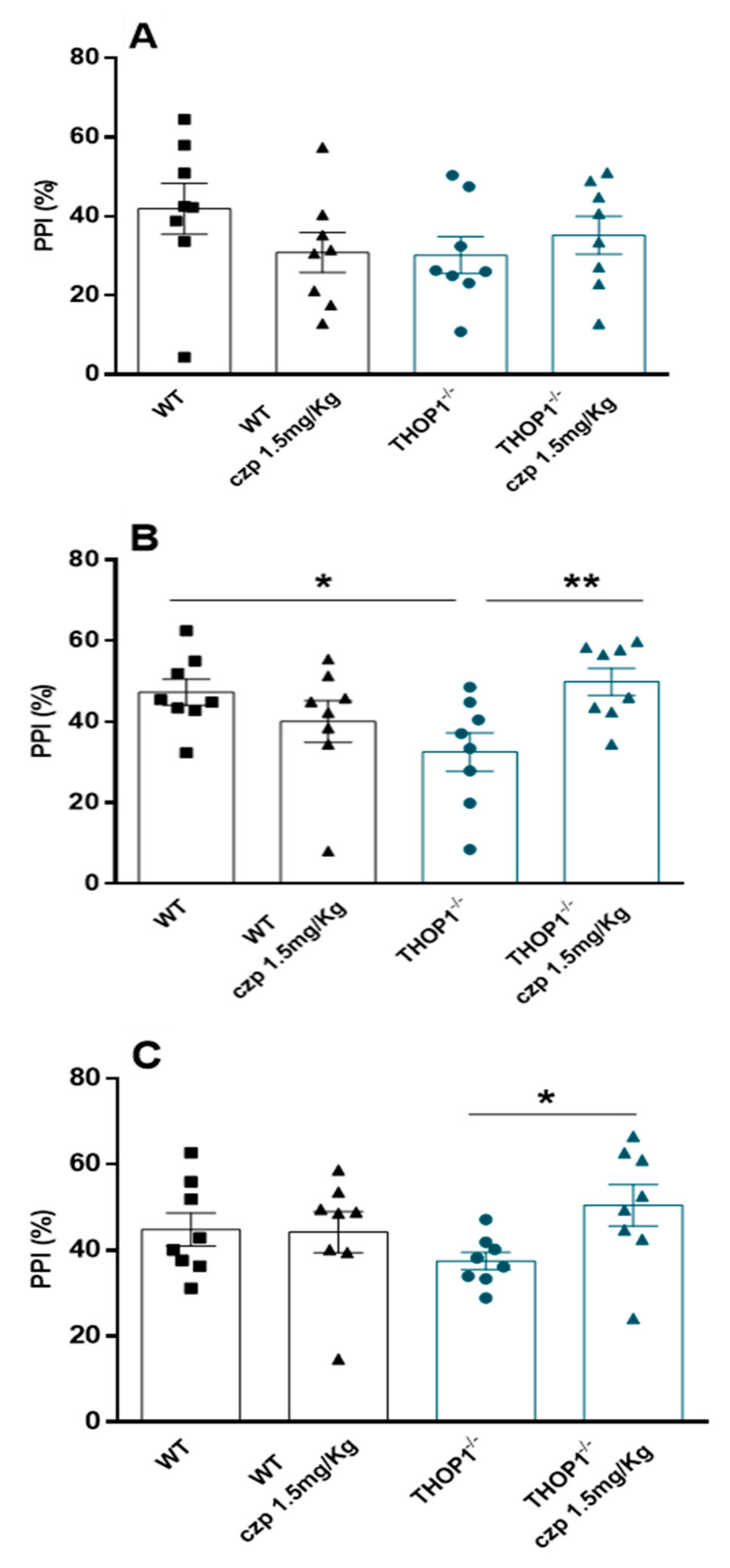
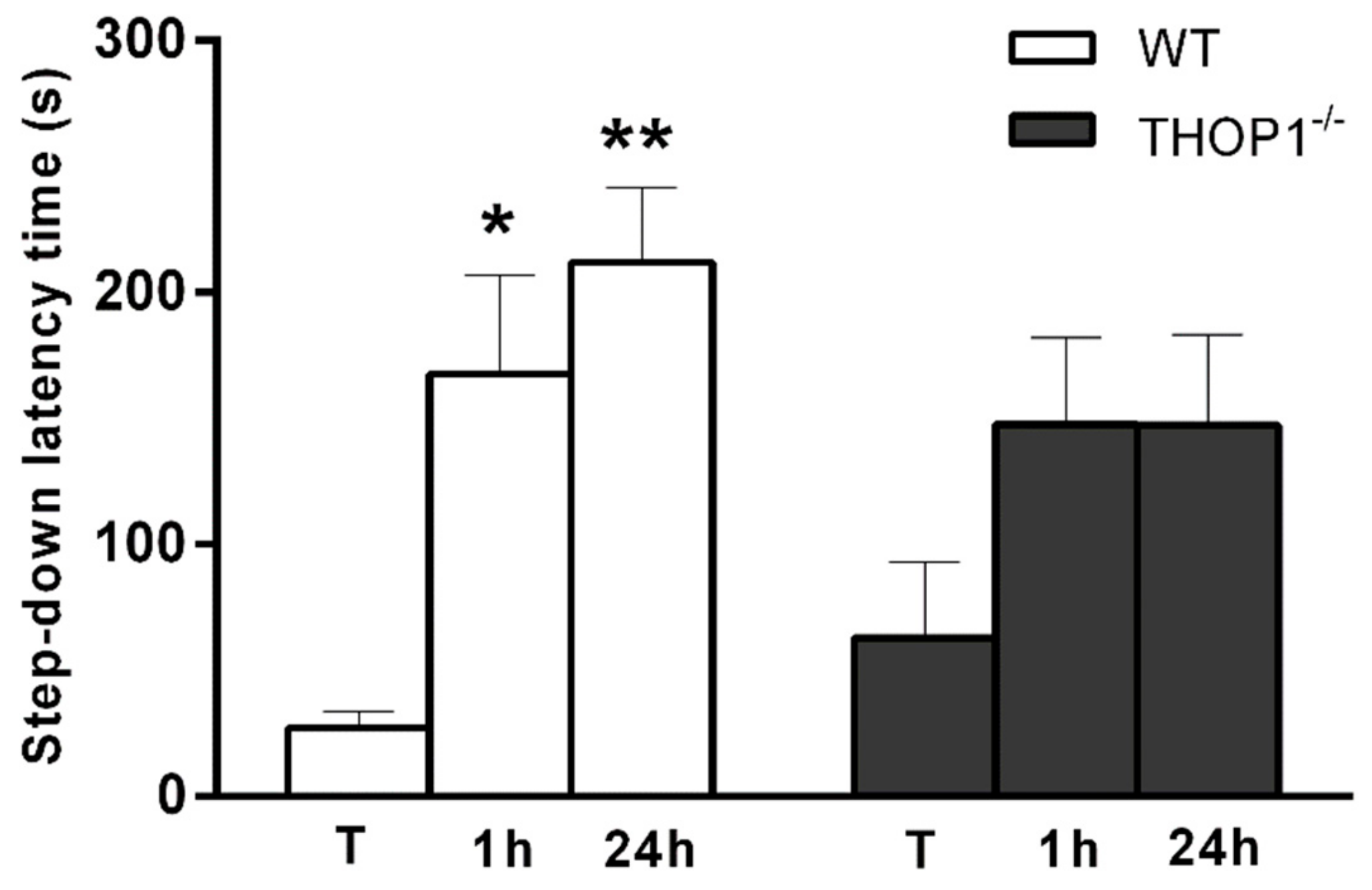
2.7. Behavior Analysis
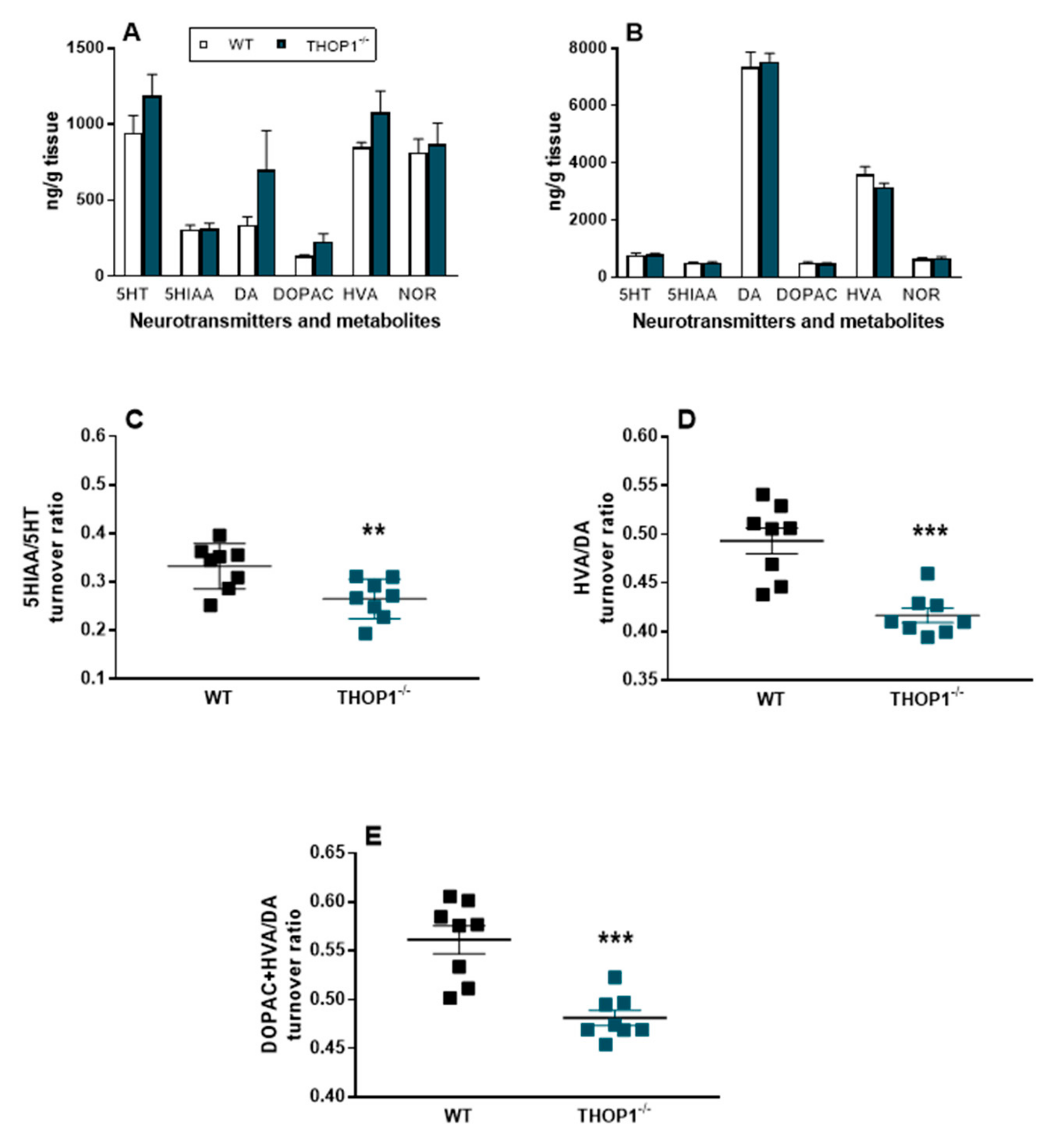
2.8. Neurotransmitter Levels and Turnover
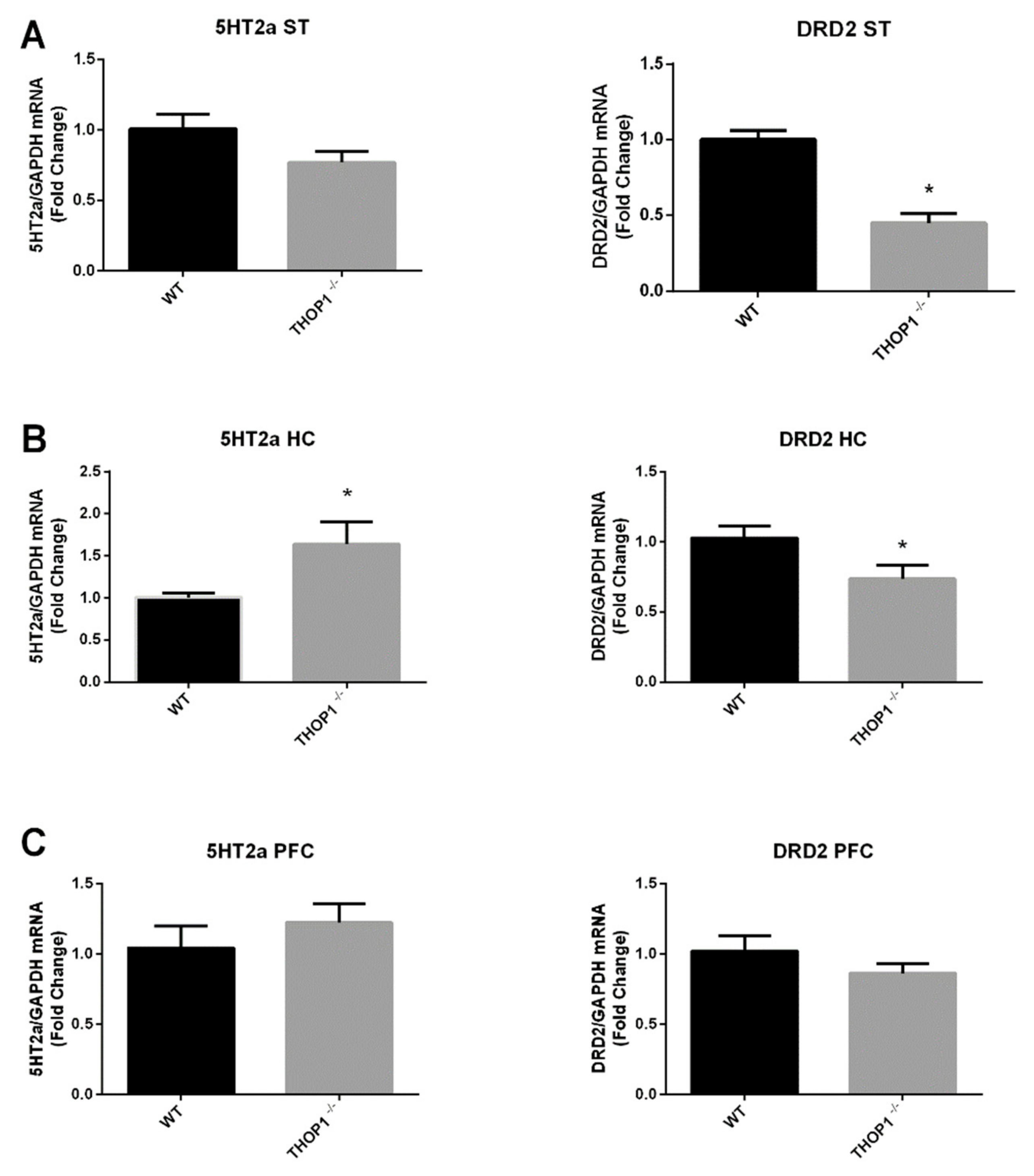
2.9. mRNA Levels of 5HT 2a and DA D2 Receptors
3. Discussion
4. Experimental Procedures
4.1. Animals
4.2. Generation of THOP1-/-
4.3. Genotyping
4.4. Enzymatic Activity
4.5. Blood Count Analysis and Coagulation
4.6. EAE Induction
4.7. Sepsis Induction
4.8. Western Blot Assays
4.9. Quantitative Real-Time PCR (qRT-PCR)
4.10. Global Gene Expression Analysis by Affymetrix GeneChip Mouse Gene 2.0 ST Array Platform
4.11. Peptide Extraction
4.12. Peptide Quantification
4.13. Dimethyl Isotopic Labeling for Peptide Semi-Quantitative Analyses
4.14. Liquid Chromatography (LC) and Mass Spectrometry (MS)
4.15. MS/MS Data Analyses
4.16. Animal Behavior Tests
4.17. Hot Plate
4.18. Forced Swim Test
4.19. Tail Suspension Test
4.20. Passive Avoidance Test (Step-Down)
4.21. Prepulse Inhibition Test (PPI)
4.22. Open Field
4.23. Elevated Plus Maze
4.24. Barnes Maze
4.25. Novel Object Recognition Test
4.26. Quantitation of Neurotransmitters and Their Metabolites in Specific Areas of Mice Brain
4.27. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Acker, G.R.; Molineaux, C.; Orlowski, M. Synaptosomal membrane-bound form of endopeptidase-24.15 generates Leu-enkephalin from dynorphin1-8, alpha- and beta-neoendorphin, and Met-enkephalin from Met-enkephalin-Arg6-Gly7-Leu8. J. Neurochem. 1987, 48, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, M.; Michaud, C.; Chu, T.G. A soluble metalloendopeptidase from rat brain. Purification of the enzyme and determination of specificity with synthetic and natural peptides. Eur. J. Biochem. 1983, 135, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kest, B.; Orlowski, M.; Molineaux, C.J.; Bodnar, R.J. Antinociceptive properties of inhibitors of endopeptidase 24.15. Int. J. Neurosci. 1991, 56, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Lasdun, A.; Orlowski, M. Inhibition of endopeptidase 24.15 greatly increases the release of luteinizing hormone and follicle stimulating hormone in response to luteinizing hormone/releasing hormone. J. Pharmacol. Exp. Ther. 1990, 253, 1265–1271. [Google Scholar] [PubMed]
- Rioli, V.; Kato, A.; Portaro, F.C.V.; Cury, G.K.; Kaat, K.T.; Vincent, B.; Checler, F.; Camargo, A.C.M.; Glucksman, M.J.; Roberts, J.L.; et al. Neuropeptide specificity and inhibition of recombinant isoforms of the endopeptidase 3.4.24.16 family: comparison with the related recombinant endopeptidase 3.4.24.15. Biochem. Biophys. Res. Commun. 1998, 250, 11–255. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.M.; Graeff, F.G. Subcellular distribution and properties of the bradykinin inactivation system in rabbit brain homogenates. Biochem. Pharmacol. 1969, 18, 548–549. [Google Scholar] [CrossRef]
- Oliveira, E.B.; Martins, A.R.; Camargo, A.C. Isolation of brain endopeptidases: influence of size and sequence of substrates structurally related to bradykinin. Biochemistry 1976, 15, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.; Campos, M.; Hemerly, J.P.; Ferro, E.S.; Camargo, A.C.M.; Juliano, M.A.; Juliano, L. Selective Neurotensin-Derived Internally Quenched Fluorogenic Substrates for Neurolysin (EC 3.4.24.16): Comparison with Thimet Oligopeptidase (EC 3.4.24.15) and Neprilysin (EC 3.4.24.11). Anal. Biochem. 2001, 292, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Checler, F.; Ferro, E.S. Neurolysin: From Initial Detection to Latest Advances. Neurochem. Res. 2018, 43, 2017–2024. [Google Scholar] [CrossRef] [PubMed]
- Checler, F.; Vincent, J.P.; Kitabgi, P. Degradation of neurotensin by rat brain synaptic membranes: involvement of a thermolysin-like metalloendopeptidase (enkephalinase), angiotensin-converting enzyme, and other unidentified peptidases. J. Neurochem. 1983, 41, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.; Oliveira, E.B.; Toffoletto, O.; Metters, K.M.; Rossier, J. Brain endo-oligopeptidase A, a putative enkephalin converting enzyme. J. Neurochem. 1987, 48, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, M.; Reznik, S.; Ayala, J.; Pierotti, A.R. Endopeptidase 24.15 from rat testes. Isolation of the enzyme and its specificity toward synthetic and natural peptides, including enkephalin-containing peptides. Biochem. J. 1989, 261, 951–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villar-Cheda, B.; Dominguez-Meijide, A.; Valenzuela, R.; Granado, N.; Moratalla, R.; Labandeira-Garcia, J.L. Aging-related dysregulation of dopamine and angiotensin receptor interaction. Neurobiol. Aging 2014, 35, 1726–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasdun, A.; Reznik, S.; Molineaux, C.J.; Orlowski, M. Inhibition of endopeptidase 24.15 slows the in vivo degradation of luteinizing hormone-releasing hormone. J. Pharmacol. Exp. Ther. 1989, 251, 439–447. [Google Scholar] [PubMed]
- Wu, T.J.; Pierotti, A.R.; Jakubowski, M.; Sheward, W.J.; Glucksman, M.J.; Smith, A.I.; King, J.C.; Fink, G.; Roberts, J.L. Endopeptidase EC 3.4.24.15 presence in the rat median eminence and hypophysial portal blood and its modulation of the luteinizing hormone surge. J. Neuroendocrinol. 1997, 9, 813–822. [Google Scholar] [CrossRef]
- Wu, T.J.; Pagano, E.; Mani, S.K. A biological role for the gonadotrophin-releasing hormone (GnRH) metabolite, GnRH-(1-5). J. Neuroendocrinol. 2009, 21, 293–298. [Google Scholar]
- Ferro, E.S.; Tambourgi, D.V.; Gobersztejn, F.; Gomes, M.D.; Sucupira, M.; Armelin, M.C.; Kipnis, T.L.; Camargo, A.C. Secretion of a neuropeptide-metabolizing enzyme similar to endopeptidase 22.19 by glioma C6 cells. Biochem. Biophys. Res. Commun. 1993, 191, 275–281. [Google Scholar] [CrossRef]
- Crack, P.J.; Wu, T.J.; Cummins, P.M.; Ferro, E.S.; Tullai, J.W.; Glucksman, M.J.; Roberts, J.L. The association of metalloendopeptidase EC 3.4.24.15 at the extracellular surface of the AtT-20 cell plasma membrane. Brain Res. 1999, 835, 113–124. [Google Scholar] [CrossRef]
- Ferro, E.S.; Tullai, J.W.; Glucksman, M.J.; Roberts, J.L. Secretion of Metalloendopeptidase 24.15 (EC 3.4.24.15). DNA Cell Biol. 1999, 18, 781–789. [Google Scholar] [CrossRef]
- Oliveira, V.; Garrido, P.A.G.; Rodrigues, C.C.; Colquhoun, A.; Castro, L.M.; Almeida, P.C.; Shida, C.S.; Juliano, M.A.; Juliano, L.; Camargo, A.C.M.; et al. Calcium modulates endopeptidase 24.15 (EC 3.4.24.15) membrane association, secondary structure and substrate specificity. FEBS J. 2005, 272, 2078–2992. [Google Scholar] [CrossRef]
- Carreno, F.R.; Goni, C.N.; Castro, L.M.; Ferro, E.S. 14-3-3 epsilon modulates the stimulated secretion of endopeptidase 24.15. J. Neurochem. 2005, 93, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Tullai, J.W.W.; Cummins, P.M.M.; Pabon, A.; Roberts, J.L.L.; Lopingco, M.C.C.; Shrimpton, C.N.N.; Smith, A.I.I.; Martignetti, J.A.A.; Ferro, E.S.; Glucksman, M.J.J. The neuropeptide processing enzyme EC 3.4.24.15 is modulated by protein kinase A phosphorylation. J. Biol. Chem. 2000, 275, 36514–36522. [Google Scholar] [CrossRef] [PubMed]
- Berti, D.A.; Morano, C.; Russo, L.C.; Castro, L.M.; Cunha, F.M.; Zhang, X.; Sironi, J.; Klitzke, C.F.; Ferro, E.S.; Fricker, L.D. Analysis of intracellular substrates and products of thimet oligopeptidase in human embryonic kidney 293 cells. J. Biol. Chem. 2009, 284, 14105–14116. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.; Campos, M.; Melo, R.L.; Ferro, E.S.; Camargo, A.C.M.; Juliano, M.A.; Juliano, L. Substrate specificity characterization of recombinant metallo oligo-peptidases thimet oligopeptidase and neurolysin. Biochemistry 2001, 40, 4417–4425. [Google Scholar] [CrossRef] [PubMed]
- Portaro, F.C.V.; Gomes, M.D.; Cabrera, A.; Fernandes, B.L.; Silva, C.L.; Ferro, E.S.; Juliano, L.; De Camargo, A.C.M. Thimet oligopeptidase and the stability of MHC class I epitopes in macrophage cytosol. Biochem. Biophys. Res. Commun. 1999, 255, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.; Hines, C.S.; Coll-Rodriguez, J.; Rodgers, D.W. Crystal Structure of Human Thimet Oligopeptidase Provides Insight into Substrate Recognition, Regulation, and Localization. J. Biol. Chem. 2004, 279, 20480–20489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierotti, A.; Dong, K.W.; Glucksman, M.J.; Orlowski, M.; Roberts, J.L. Molecular cloning and primary structure of rat testes metalloendopeptidase EC 3.4.24.15. Biochemistry 1990, 29, 10323–10329. [Google Scholar] [CrossRef]
- McKie, N.; Dando, P.M.; Rawlings, N.D.; Barrett, A.J. Thimet oligopeptidase: similarity to “soluble angiotensin II-binding protein” and some corrections to the published amino acid sequence of the rat testis enzyme. Biochem. J. 1993, 295, 57–60. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. Evolutionary families of peptidases. Biochem. J. 1993, 290, 205–218. [Google Scholar] [Green Version]
- Shrimpton, C.N.; Glucksman, M.J.; Lew, R.A.; Tullai, J.W.; Margulies, E.H.; Roberts, J.L.; Smith, A.I. Thiol activation of endopeptidase EC 3.4.24.15. A novel mechanism for the regulation of catalytic activity. J. Biol. Chem. 1997, 272, 17395–17399. [Google Scholar] [CrossRef]
- Demasi, M.; Piassa Filho, G.M.; Castro, L.M.; Ferreira, J.C.; Rioli, V.; Ferro, E.S. Oligomerization of the cysteinyl-rich oligopeptidase EP24.15 is triggered by S-glutathionylation. Free Radic. Biol. Med. 2008, 44, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Malvezzi, A.; Higa, P.M.; T-do Amaral, A.; Silva, G.M.; Gozzo, F.C.; Ferro, E.S.; Castro, L.M.; de Rezende, L.; Monteiro, G.; Demasi, M.; et al. The cysteine-rich protein thimet oligopeptidase as a model of the structural requirements for S-glutathiolation and oxidative oligomerization. PLoS One 2012, 7, e39408. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Fontenele-Neto, J.D.; Massarelli, E.E.; Garrido, P.A.G.; Beaudet, A.; Ferro, E.S. Comparative fine structural distribution of endopeptidase 24.15 (EC3.4.24.15) and 24.16 (EC3.4.24.16) in rat brain. J. Comp. Neurol. 2001, 438, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Massarelli, E.E.; Casatti, C.A.; Kato, A.; Camargo, A.C.M.; Bauer, J.A.; Glucksman, M.J.; Roberts, J.L.; Hirose, S.; Ferro, E.S. Differential subcellular distribution of neurolysin (EC 3.4.24.16) and thimet oligopeptidase (EC 3.4.24.15) in the rat brain. Brain Res. 1999, 851, 261–265. [Google Scholar] [CrossRef]
- Silva, C.L.; Portaro, F.C.V.; Bonato, V.L.D.; De Camargo, A.C.M.; Ferro, E.S. Thimet oligopeptidase (EC 3.4.24.15), a novel protein on the route of MHC class I antigen presentation. Biochem. Biophys. Res. Commun. 1999, 255, 591–595. [Google Scholar] [CrossRef] [PubMed]
- KIM, S.I.; PABON, A.; Swanson, T.A.; GLUCKSMAN, M.J. Regulation of cell-surface major histocompatibility complex class I expression by the endopeptidase EC3.4.24.15 (thimet oligopeptidase). Biochem. J. 2003, 375, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.H.; Khan, S.; Seifert, U.; Le Gall, S.; Chow, K.M.; Paschen, A.; Bres-Vloemans, S.A.; de Ru, A.; van Montfoort, N.; Franken, K.L.M.C.; et al. Antigen processing by nardilysin and thimet oligopeptidase generates cytotoxic T cell epitopes. Nat. Immunol. 2011, 12, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Grum-Tokars, V.; Swanson, T.A.; Cotter, E.J.; Cahill, P.A.; Roberts, J.L.; Cummins, P.M.; Glucksman, M.J. Novel roles of neuropeptide processing enzymes: EC3.4.24.15 in the neurome. J. Neurosci. Res. 2003, 74, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Rioli, V.; Gozzo, F.C.; Heimann, A.S.; Linardi, A.; Krieger, J.E.; Shida, C.S.; Almeida, P.C.; Hyslop, S.; Eberlin, M.N.; Ferro, E.S. Novel natural peptide substrates for endopeptidase 24.15, neurolysin, and angiotensin-converting enzyme. J. Biol. Chem. 2003, 278, 8547–8555. [Google Scholar] [CrossRef] [PubMed]
- Ferro, E.S.; Hyslop, S.; Camargo, A.C.M. Intracellullar peptides as putative natural regulators of protein interactions. J. Neurochem. 2004, 91, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Rioli, V.; Ferro, E.S. Substrate Capture Assay Using Inactive Oligopeptidases to Identify Novel Peptides. Methods Mol. Biol. 2018, 1719, 97–105. [Google Scholar] [PubMed]
- Gelman, J.S.; Sironi, J.; Castro, L.M.; Ferro, E.S.; Fricker, L.D. Peptidomic analysis of human cell lines. J. Proteome Res. 2011, 10, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Fesenko, I.A.; Arapidi, G.P.; Skripnikov, A.Y.; Alexeev, D.G.; Kostryukova, E.S.; Manolov, A.I.; Altukhov, I.A.; Khazigaleeva, R.A.; Seredina, A.V.; Kovalchuk, S.I.; et al. Specific pools of endogenous peptides are present in gametophore, protonema, and protoplast cells of the moss Physcomitrella patens. BMC Plant Biol. 2015, 15, 87. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.M.M.; Correa, C.N.; Iwai, L.K.; Ferro, E.S.; de Castro, L.M. Characterization of Intracellular Peptides from Zebrafish (Danio rerio) Brain. Zebrafish 2019, 16. [Google Scholar] [CrossRef]
- Berezniuk, I.; Sironi, J.J.; Wardman, J.; Pasek, R.C.; Berbari, N.F.; Yoder, B.K.; Fricker, L.D. Quantitative peptidomics of Purkinje cell degeneration mice. PLoS One 2013, 8, e60981. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Yang, C.; Castro, L.M.; Tashima, A.K.; Ferro, E.S.; Moir, R.D.; Willis, I.M.; Fricker, L.D. Analysis of the Yeast Peptidome and Comparison with the Human Peptidome. PLoS One 2016, 11, e0163312. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.C.; Castro, L.M.; Gozzo, F.C.; Ferro, E.S. Inhibition of thimet oligopeptidase by siRNA alters specific intracellular peptides and potentiates isoproterenol signal transduction. FEBS Lett. 2012, 586, 3287–3292. [Google Scholar] [CrossRef] [PubMed]
- Cunha, F.M.; Berti, D.A.; Ferreira, Z.S.; Klitzke, C.F.; Markus, R.P.; Ferro, E.S. Intracellular peptides as natural regulators of cell signaling. J. Biol. Chem. 2008, 283, 24448–24459. [Google Scholar] [CrossRef] [PubMed]
- Pollio, G.; Hoozemans, J.J.M.M.; Andersen, C.A.; Roncarati, R.; Rosi, M.C.; van Haastert, E.S.; Seredenina, T.; Diamanti, D.; Gotta, S.; Fiorentini, A.; et al. Increased expression of the oligopeptidase THOP1 is a neuroprotective response to Abeta toxicity. Neurobiol. Dis. 2008, 31, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Sundstrom, J.M.; Hernández, C.; Weber, S.R.; Zhao, Y.; Dunklebarger, M.; Tiberti, N.; Laremore, T.; Simó-Servat, O.; Garcia-Ramirez, M.; Barber, A.J.; et al. Proteomic Analysis of Early Diabetic Retinopathy Reveals Mediators of Neurodegenerative Brain Diseases. Invest. Ophthalmol. Vis. Sci. 2018, 59, 2264–2274. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H. Prognosis related miRNAs, DNA methylation, and epigenetic interactions in lung adenocarcinoma. Neoplasma 2019, 66, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Barghash, A.; Arslan, T. Robust Detection of Outlier Samples and Genes in Expression Datasets. J. Proteomics Bioinform. 2016, 09, 38–48. [Google Scholar] [CrossRef]
- Castro, L.M.; Cavalcanti, D.M.L.P.; Araujo, C.B.; Rioli, V.; Icimoto, M.Y.; Gozzo, F.C.; Juliano, M.; Juliano, L.; Oliveira, V.; Ferro, E.S. Peptidomic analysis of the neurolysin-knockout mouse brain. J. Proteomics 2014, 111, 238–248. [Google Scholar] [CrossRef]
- Crochemore, T.; de Piza, F.M.T.; dos Rodrigues, R.R.; de Guerra, J.C.C.; Ferraz, L.J.R.; Corrêa, T.D. A new era of thromboelastometry. Einstein (São Paulo) 2017, 15, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Bittner, S.; Afzali, A.M.; Wiendl, H.; Meuth, S.G. Myelin Oligodendrocyte Glycoprotein (MOG35-55) Induced Experimental Autoimmune Encephalomyelitis (EAE) in C57BL/6 Mice. J. Vis. Exp. 2014. [CrossRef]
- Sun, D.; Whitaker, J.N.; Huang, Z.; Liu, D.; Coleclough, C.; Wekerle, H.; Raine, C.S. Myelin antigen-specific CD8+ T cells are encephalitogenic and produce severe disease in C57BL/6 mice. J. Immunol. 2001, 166, 7579–7587. [Google Scholar] [CrossRef]
- Smith, A.I.; Lew, R.A.; Shrimpton, C.N.; Evans, R.G.; Abbenante, G. A novel stable inhibitor of endopeptidases EC 3.4.24.15 and 3.4.24.16 potentiates bradykinin-induced hypotension. Hypertens. 2000, 35, 626–630. [Google Scholar] [CrossRef]
- Novaira, H.J.; Yates, M.; Diaczok, D.; Kim, H.; Wolfe, A.; Radovick, S. The gonadotropin-releasing hormone cell-specific element is required for normal puberty and estrous cyclicity. J. Neurosci. 2011, 31, 3336–3343. [Google Scholar] [CrossRef]
- Yamanaka, C.; Lebrethon, M.C.; Vandersmissen, E.; Gerard, A.; Purnelle, G.; Lemaitre, M.; Wilk, S.; Bourguignon, J.P. Early prepubertal ontogeny of pulsatile gonadotropin-releasing hormone (GnRH) secretion: I. Inhibitory autofeedback control through prolyl endopeptidase degradation of GnRH. Endocrinology 1999, 140, 4609–4615. [Google Scholar] [CrossRef]
- Lew, R.A.; Tetaz, T.J.; Glucksman, M.J.; Roberts, J.L.; Smith, A.I. Evidence for a two-step mechanism of gonadotropin-releasing hormone metabolism by prolyl endopeptidase and metalloendopeptidase EC 3.4.24.15 in ovine hypothalamic extracts. J. Biol. Chem. 1994, 269, 12626–12632. [Google Scholar] [PubMed]
- Cardozo, C.; Orlowski, M. Evidence that enzymatic conversion of N-[1(R,S)-carboxy-3-phenylpropyl]-Ala-Ala-Phe-p-aminobenzoate, a specific inhibitor of endopeptidase 24.15, to N-[1(R,S)-carboxy-3-phenylpropyl]-Ala-Ala is necessary for inhibition of angiotensin converting enzyme. Peptides 1993, 14, 1259–1262. [Google Scholar] [CrossRef]
- Telford, S.E.; Smith, A.I.; Lew, R.A.; Perich, R.B.; Madden, A.C.; Evans, R.G. Role of angiotensin converting enzyme in the vascular effects of an endopeptidase 24.15 inhibitor. Br. J. Pharmacol. 1995, 114, 1185–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojaghi, M.; Kastelic, J.; Thundathil, J. Testis-specific isoform of angiotensin-converting enzyme (tACE) is involved in the regulation of bovine sperm capacitation. Mol. Reprod. Dev. 2017, 84, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Pai, J.; Yoon, T.; Kim, N.D.; Lee, I.-S.; Yu, J.; Shin, I. High-throughput profiling of peptide-RNA interactions using peptide microarrays. J. Am. Chem. Soc. 2012, 134, 19287–19296. [Google Scholar] [CrossRef] [PubMed]
- Pai, J.; Hyun, S.; Hyun, J.Y.; Park, S.-H.; Kim, W.-J.; Bae, S.-H.; Kim, N.-K.; Yu, J.; Shin, I. Screening of Pre-miRNA-155 Binding Peptides for Apoptosis Inducing Activity Using Peptide Microarrays. J. Am. Chem. Soc. 2016, 138, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Sharp, P.A. Tat-SF1: cofactor for stimulation of transcriptional elongation by HIV-1 Tat. Science 1996, 274, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Monte, E.R.; Rossato, C.; Llanos, R.P.; Russo, L.C.; de Castro, L.M.; Gozzo, F.C.; de Araujo, C.B.; Peron, J.P.S.; Sant’Anna, O.A.; Ferro, E.S.; et al. Interferon-gamma activity is potentiated by an intracellular peptide derived from the human 19S ATPase regulatory subunit 4 of the proteasome. J. Proteomics 2017, 151, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Wu, J.Q.; Ye, M.J.; Zheng, K.; He, J.C.; Zhang, X.; Liu, J.H.; Tian, H.J.; Gong, B.H.; Chen, D.C.; et al. Association of angiotensin-converting enzyme gene polymorphism with schizophrenia and depressive symptom severity in a Chinese population. Hum. Psychopharmacol. 2015, 30, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.J.; Matsas, R.; Kenny, A.J. Are there neuropeptide-specific peptidases? Biochem. Pharmacol. 1985, 34, 1347–1356. [Google Scholar] [CrossRef]
- Oliveira, V.; Araújo, M.C.; Rioli, V.; de Camargo, A.C.M.; Tersariol, I.L.S.; Juliano, M.A.; Juliano, L.; Ferro, E.S. A structure-based site-directed mutagenesis study on the neurolysin (EC 3.4.24.16) and thimet oligopeptidase (EC 3.4.24.15) catalysis. FEBS Lett. 2003, 541, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Fricker, L.D.; Gelman, J.S.; Castro, L.M.; Gozzo, F.C.; Ferro, E.S. Peptidomic analysis of HEK293T cells: effect of the proteasome inhibitor epoxomicin on intracellular peptides. J. Proteome Res. 2012, 11, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Castro, L.M.; Dulman, R.; Yang, C.; Schmidt, M.; Ferro, E.S.; Fricker, L.D. Proteasome inhibitors alter levels of intracellular peptides in HEK293T and SH-SY5Y cells. PLoS One 2014, 9, e103604. [Google Scholar] [CrossRef] [PubMed]
- Koudriavtseva, T.; Mainero, C. Neuroinflammation, neurodegeneration and regeneration in multiple sclerosis: intercorrelated manifestations of the immune response. Neural Regen. Res. 2016, 11, 1727. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Castelli, L.; Goverman, J.M. MHC class I-restricted myelin epitopes are cross-presented by Tip-DCs that promote determinant spreading to CD8+ T cells. Nat. Immunol. 2013, 14, 254–261. [Google Scholar] [CrossRef]
- York, I.A.; Mo, A.X.Y.; Lemerise, K.; Zeng, W.; Shen, Y.; Abraham, C.R.; Saric, T.; Goldberg, A.L.; Rock, K.L. The cytosolic endopeptidase, thimet oligopeptidase, destroys antigenic peptides and limits the extent of MHC class I antigen presentation. Immunity 2003, 18, 429–440. [Google Scholar] [CrossRef]
- Mars, L.T.; Saikali, P.; Liblau, R.S.; Arbour, N. Contribution of CD8 T lymphocytes to the immuno-pathogenesis of multiple sclerosis and its animal models. Biochim. Biophys. Acta 2011, 1812, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Shchetynsky, K.; Diaz-Gallo, L.-M.; Folkersen, L.; Hensvold, A.H.; Catrina, A.I.; Berg, L.; Klareskog, L.; Padyukov, L. Discovery of new candidate genes for rheumatoid arthritis through integration of genetic association data with expression pathway analysis. Arthritis Res. Ther. 2017, 19, 19. [Google Scholar] [CrossRef] [Green Version]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Benjamim, C.F.; Ferreira, S.H.; Cunha, F.D.Q. Role of Nitric Oxide in the Failure of Neutrophil Migration in Sepsis. J. Infect. Dis. 2000, 182, 214–223. [Google Scholar] [CrossRef]
- Delano, M.J.; Ward, P.A. Sepsis-induced immune dysfunction: can immune therapies reduce mortality? J. Clin. Invest. 2016, 126, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Moldawer, L.L.; Opal, S.M.; Reinhart, K.; Turnbull, I.R.; Vincent, J.-L. Sepsis and septic shock. Nat. Rev. Dis. Prim. 2016, 2, 16045. [Google Scholar] [CrossRef] [PubMed]
- Battle, C.E.; Davies, G.; Evans, P.A. Long term health-related quality of life in survivors of sepsis in South West Wales: an epidemiological study. PLoS One 2014, 9, e116304. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, N.A.; Unsinger, J.; Hotchkiss, R.S.; Ayala, A. The new normal: immunomodulatory agents against sepsis immune suppression. Trends Mol. Med. 2014, 20, 224–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Sepsis-induced immunosuppression: from cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013, 13, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.D.; Juliano, L.; Ferro, E.S.; Matsueda, R.; Camargo, A.C.M. Dynorphin-derived peptides reveal the presence of a critical cysteine for the activity of brain endo-oligopeptidase A. Biochem. Biophys. Res. Commun. 1993, 197. [Google Scholar] [CrossRef] [PubMed]
- Shrimpton, C.N.; Smith, A.I.; Lew, R.A. Soluble metalloendopeptidases and neuroendocrine signaling. Endocr. Rev. 2002, 23, 647–664. [Google Scholar] [CrossRef] [PubMed]
- Peitl, V.; Štefanović, M.; Karlović, D. Depressive symptoms in schizophrenia and dopamine and serotonin gene polymorphisms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 77, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Park, I.H.; Lee, B.C.; Kim, J.J.; Kim, J.I.; Koo, M.-S. Effort-Based Reinforcement Processing and Functional Connectivity Underlying Amotivation in Medicated Patients with Depression and Schizophrenia. J. Neurosci. 2017, 37, 4370–4380. [Google Scholar] [CrossRef]
- Ray, K.; Hines, C.S.; Rodgers, D.W. Mapping sequence differences between thimet oligopeptidase and neurolysin implicates key residues in substrate recognition. Protein Sci. 2002, 11, 2237–2246. [Google Scholar] [CrossRef]
- Kuzman, M.R.; Medved, V.; Terzic, J.; Krainc, D. Genome-wide expression analysis of peripheral blood identifies candidate biomarkers for schizophrenia. J. Psychiatr. Res. 2009, 43, 1073–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, D.M.L.P.; Castro, L.M.; Rosa Neto, J.C.; Seelaender, M.; Neves, R.X.; Oliveira, V.; Forti, F.L.; Iwai, L.K.; Gozzo, F.C.; Todiras, M.; et al. Neurolysin knockout mice generation and initial phenotype characterization. J. Biol. Chem. 2014, 289, 15426–15440. [Google Scholar] [CrossRef] [PubMed]
- Acharjee, S.; Nayani, N.; Tsutsui, M.; Hill, M.N.; Ousman, S.S.; Pittman, Q.J. Altered cognitive-emotional behavior in early experimental autoimmune encephalitis--cytokine and hormonal correlates. Brain. Behav. Immun. 2013, 33, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Hamasato, E.K.; de Lima, A.P.N.; de Oliveira, A.P.L.; dos Santos Franco, A.L.; de Lima, W.T.; Palermo-Neto, J. Cohabitation with a sick partner increases allergic lung inflammatory response in mice. Brain. Behav. Immun. 2014, 42, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, N.; Novaes, L.S.; Dragunas, G.; Rodrigues, J.R.; Brandão, W.; Camarini, R.; Peron, J.P.S.; Munhoz, C.D. High dose of dexamethasone protects against EAE-induced motor deficits but impairs learning/memory in C57BL/6 mice. Sci. Rep. 2019, 9, 6673. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Huber-Lang, M.S.; Flierl, M.A.; Ward, P.A. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat. Protoc. 2009, 4, 31–36. [Google Scholar] [CrossRef]
- Reckziegel, P.; Festuccia, W.T.; Britto, L.R.G.; Jang, K.L.L.; Romão, C.M.; Heimann, J.C.; Fogaça, M.V.; Rodrigues, N.S.; Silva, N.R.; Guimarães, F.S.; et al. A novel peptide that improves metabolic parameters without adverse central nervous system effects. Sci. Rep. 2017, 7, 14781. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 2012; ISBN 0125476361. [Google Scholar]
- Chartoumpekis, D.V.; Zaravinos, A.; Ziros, P.G.; Iskrenova, R.P.; Psyrogiannis, A.I.; Kyriazopoulou, V.E.; Habeos, I.G. Differential expression of microRNAs in adipose tissue after long-term high-fat diet-induced obesity in mice. PLoS One 2012, 7, e34872. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45. [Google Scholar] [CrossRef]
- Gomes, M.D.; Lecker, S.H.; Jagoe, R.T.; Navon, A.; Goldberg, A.L. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc. Natl. Acad. Sci. USA 2001, 98, 14440–14445. [Google Scholar] [CrossRef] [Green Version]
- Fricker, L.D.; Lim, J.; Pan, H.; Che, F.-Y. Peptidomics: identification and quantification of endogenous peptides in neuroendocrine tissues. Mass Spectrom. Rev. 2006, 25, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Udenfriend, S.; Stein, S.; Böhlen, P.; Dairman, W.; Leimgruber, W.; Weigele, M. Fluorescamine: a reagent for assay of amino acids, peptides, proteins, and primary amines in the picomole range. Science 1972, 178, 871–872. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.M.; Berti, D.A.; Russo, L.C.; Coelho, V.; Gozzo, F.C.; Oliveira, V.; Ferro, E.S. Similar Intracellular Peptide Profile of TAP1/β2 Microglobulin Double-Knockout Mice and C57BL/6 Wild-Type Mice as Revealed by Peptidomic Analysis. AAPS J. 2010, 12, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.; Peres, F.F.; Almeida, V.; Calzavara, M.B.; Zuardi, A.W.; Hallak, J.E.C.; Crippa, J.A.S.; Abílio, V.C. Effects of cannabinoid drugs on the deficit of prepulse inhibition of startle in an animal model of schizophrenia: the SHR strain. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoglio, L.J.; Carobrez, A.P. Previous maze experience required to increase open arms avoidance in rats submitted to the elevated plus-maze model of anxiety. Behav. Brain Res. 2000, 108, 197–203. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: a primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar] [PubMed]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl). 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Bertorelli, R.; Ongini, E.; Costentin, J.; Vaugeois, J.M. Adenosine A2A receptor antagonists are potential antidepressants: evidence based on pharmacology and A2A receptor knockout mice. Br. J. Pharmacol. 2001, 134, 68–77. [Google Scholar] [CrossRef]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Fujioka, R.; Nii, T.; Iwaki, A.; Shibata, A.; Ito, I.; Kitaichi, K.; Nomura, M.; Hattori, S.; Takao, K.; Miyakawa, T.; et al. Comprehensive behavioral study of mGluR3 knockout mice: implication in schizophrenia related endophenotypes. Mol. Brain 2014, 7, 31. [Google Scholar] [CrossRef]
- Geyer, M.A.; Ellenbroek, B. Animal behavior models of the mechanisms underlying antipsychotic atypicality. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.S.; Bevilaqua, L.R.M.; Bonini, J.S.; Rossato, J.I.; Köhler, C.A.; Medina, J.H.; Izquierdo, I.; Cammarota, M. Angiotensin II blocks memory consolidation through an AT2 receptor-dependent mechanism. Psychopharmacology (Berl). 2005, 179, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Nasehi, M.; Piri, M.; Nouri, M.; Farzin, D.; Nayer-Nouri, T.; Zarrindast, M.R. Involvement of dopamine D1/D2 receptors on harmane-induced amnesia in the step-down passive avoidance test. Eur. J. Pharmacol. 2010, 634, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Reckziegel, P.; Boufleur, N.; Barcelos, R.C.S.; Benvegnú, D.M.; Pase, C.S.; Muller, L.G.; Teixeira, A.M.; Zanella, R.; Prado, A.C.P.; Fett, R.; et al. Oxidative stress and anxiety-like symptoms related to withdrawal of passive cigarette smoke in mice: beneficial effects of pecan nut shells extract, a by-product of the nut industry. Ecotoxicol. Environ. Saf. 2011, 74, 1770–1778. [Google Scholar] [CrossRef] [PubMed]
- Singer, P.; Yee, B.K. Reversal of scopolamine-induced disruption of prepulse inhibition by clozapine in mice. Pharmacol. Biochem. Behav. 2012, 101, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, G.G.; Botton, P.H.; Moreira, J.D.; Ardais, A.P.; Oses, J.P.; Souza, D.O. Influence of environmental enrichment on an object recognition task in CF1 mice. Physiol. Behav. 2010, 99, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.J.; Ramabadran, K. Enhancement of a nociceptive reaction by opioid antagonists in mice. Br. J. Pharmacol. 1978, 64, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, M.; Arqué, G.; McDonald, J.; Andreu, N.; Martínez-Cué, C.; Flórez, J.; Fillat, C. Behavioral Characterization of a Mouse Model Overexpressing DSCR1/ RCAN1. PLoS One 2011, 6, e17010. [Google Scholar] [CrossRef]
- Braff, D.; Stone, C.; Callaway, E.; Geyer, M.; Glick, I.; Bali, L. Prestimulus effects on human startle reflex in normals and schizophrenics. Psychophysiology 1978, 15, 339–343. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Geyer, M.A. Using an animal model of deficient sensorimotor gating to study the pathophysiology and new treatments of schizophrenia. Schizophr. Bull. 1998, 24, 285–301. [Google Scholar] [CrossRef]
- Nunes, E.A. Schizophrenia: Progress in multidisciplinary treatment. Rev. Bras. Psiquiatr. 2012, 34, 521–522. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, e52434. [Google Scholar] [CrossRef] [PubMed]
- Carobrez, A.P.; Bertoglio, L.J. Ethological and temporal analyses of anxiety-like behavior: the elevated plus-maze model 20 years on. Neurosci. Biobehav. Rev. 2005, 29, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S.; Ferguson, S.A. Barnes maze testing strategies with small and large rodent models. J. Vis. Exp. 2014, e51194. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.S.; Sunyer, B.; Höger, H.; Luber, G. Evaluation of spatial memory of C57BL/6J and CD1 mice in the Barnes maze, the Multiple T-maze and in the Morris water maze. Behav. Brain Res. 2009, 198, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Bevins, R.A.; Besheer, J. Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study “recognition memory”. Nat. Protoc. 2006, 1, 1306–1311. [Google Scholar] [CrossRef]
- Margatho, R.O.; de Massoco, C.O.; Calefi, A.S.; Cruz, D.S.G.; Sandini, T.M.; Alves, G.J.; Florio, J.C.; Palermo-Neto, J. Beta-Adrenergic Blockade Decreases the Neuroimmune Changes in Mice Induced by Cohabitation with an Ehrlich Tumor-Bearing Cage Mate. Neuroimmunomodulation 2017, 24, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Sandini, T.M.; Reis-Silva, T.M.; Navas-Suáresz, P.; Auada, A.V.V.; Lebrun, I.; Flório, J.C.; Bernardi, M.M.; Spinosa, H.S. Ivermectin reduces motor coordination, serum testosterone, and central neurotransmitter levels but does not affect sexual motivation in male rats. Reprod. Toxicol. 2017, 74, 195–203. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Peptide Sequence | Average Ratio | Average Ratio | Average Ratio | |
|---|---|---|---|---|---|
| Location | THOP1/WT PFC | THOP1/WT ST | THOP1/WT HC | ||
| ±SE (n) | ±SE (n) | ±SE (n) | |||
| Acyl-CoA-binding protein | ATVGDVNTDRPGLLDL | C | 1.04 ± 0.32 (3) | 1.80 ± 0.66 (3) | |
| Acyl-CoA-binding protein | GDVNTDRPGLLDL | C | 1.90 ± 0.58 (3) | ||
| Acyl-CoA-binding protein | KQATVGDVNTDRPGLLDL 2+ | C | 1.11 ± 0.26 (3) | 2.50 ± 1.17 (3) | |
| Acyl-CoA-binding protein | KQATVGDVNTDRPGLLDL 3+ | C | 0.46 ± 0.12 (3) | 1.03 ± 0.27 (3) | 2.20 ± 0.96 (3) |
| Acyl-CoA-binding protein | SHFKQATVGDVNTDRPGLLDL | C | 1.36 ± 0.39 (3) | ||
| Acyl-CoA-binding protein | TVGDVNTDRPGLLDL | C | 1.07 ± 0.31 (3) | 2.60 ± 1.33 (3) | |
| Acyl-CoA-binding protein | VEKVDELKKKYGI 2+ | C | 0.4 ± 0.01 (2) | 2.3 ± 0.90 (3) | |
| Acyl-CoA-binding protein | VEKVDELKKKYGI 3+ | C | 0.4 ± 0.10 (2) | 0.91 ± 0.14 (3) | 2.07 ± 0.73 (3) |
| ATP synthase-coupling factor 6 | KFDDPKFEVIDKPQS | M | 1.12 ± 0.26 (2) | ||
| Cerebellin-4 | AANSKVAFSAVRSTN | V | 1.12 ± 0.11 (3) | ||
| Cerebellin-4 | SKVAFSAVRSTN | V | 0.89 ± 0.21 (3) | ||
| Clathrin coat assembly protein AP180 | ASKGLGSDLDSSLAS (1.52 ± 0.12) | M/C | 0.53 ± 0.11 (3) | 1.08 ± 0.12 (3) | 1.05 ± 0.04 (3) |
| Clathrin coat assembly protein AP180 | KGLGSDLDSSLASL (1.54 ± 0.16) | M/C | 0.56 ± 0.14 (3) | ||
| Clathrin coat assembly protein AP180 | SKGLGSDLDSSLASL | M/C | 0.55 ± 0.12 (3) | ||
| Clathrin coat assembly protein AP180 | SPSPTPATQSPKKPPAKDPLADLNIKDFL | M/C | 0.51 ± 0.12 (3) | 1.17 ± 0.18 (3) | 0.99 ± 0.06 (3) |
| Cysteine and glycine-rich protein 1 | GQGAGALVHSE | N | 1.08 ± 0.02 (3) | 2.17 ± 0.64 (3) | |
| Cytochrome b-c1 complex subunit Rieske | ARSGPFAPVLSAT | M | 0.81 ± 0.19 (3) | ||
| Cytochrome b-c1 complex subunit Rieske | GLNVPASVRF | M | 0.94 ± 0.21 (3) | ||
| Cytochrome b-c1 complex subunit Rieske | SGQAAARPLVA | M | 0.86 ± 0.21 (3) | 1.01 ± 0.10 (3) | |
| Cytochrome b-c1 complex subunit Rieske | SGQAAARPLVATV | M | 0.75 ± 0.14 (3) | 1.25 ± 0.20 (3) | 0.94 ± 0.08 (3) |
| Cytochrome b-c1 complex subunit Rieske | TVGLNVPASVRF | M | 0.95 ± 0.17 (3) | 1.17 ± 0.16 (3) | 0.98 ± 0.13 (3) |
| Cytochrome c oxidase subunit 5A | GISTPEELGLDKV | M | 0.49 ± 0.06 (3) | 1.05 ± 0.19 (3) | 1.51 ± 0.34 (3) |
| Cytochrome c oxidase subunit 5A | NDFASAVRILEV (1.43 ± 0.62) | M | 0.36 ± 0.12 (2) | 1.70 ± 0.38 (3) | |
| Cytochrome c oxidase subunit 5A | RPTLNELGISTPEELGLDKV | M | 0.42 ± 0.12 (3) | 1.20 ± 0.25 (3) | 1.53 ± 0.37 (3) |
| Cytochrome c oxidase subunit 6A1 | FHNPHVNPLPTGYEDE | M | 1.09 ± 0.13 (3) | 1.12 ± 0.14 (3) | |
| Cytochrome c oxidase subunit 6A1 | HNPHVNPLPTGYEDE | M | 0.86 ± 0.13 (3) | 1.13 ± 0.36 (3) | 1.30 ± 0.20 (3) |
| Cytochrome c oxidase subunit 6A1 | NPHVNPLPTGYEDE | M | 1.07 ± 0.18 (3) | ||
| Gamma-enolase | AGNSDLILPVPAFNVINGGSHAGNKL | C | 0.76 ± 0.20 (3) | ||
| Glyceraldehyde-3-phosphate dehyd. | AFRVPTPNVSVVDL | C/N | 1.51 ± 0.09 (3) | 0.99 ± 0.09 (3) | |
| LIM zinc-binding domain-containing Nebulette | TQVVSDAAYKGVQPHVV | C | 0.59 ± 0.06 (3) | 0.93 ± 0.22 (3) | |
| Macrophage migration inhibitory factor | AQATGKPAQYIA | C | 0.92 ± 0.12 (3) | ||
| Macrophage migration inhibitory factor | AQATGKPAQYIA | C | 1.17 ± 0.02 (2) | ||
| Macrophage migration inhibitory factor | AQATGKPAQYIAVHVVPDQL 2+ | C | 0.65 ± 0.03 (3) | 0.87 ± 0.07 (3) | |
| Macrophage migration inhibitory factor | AQATGKPAQYIAVHVVPDQL 3+ | C | 0.59 ± 0.12 (3) | 1.17 ± 0.07 (3) | 0.75 ± 0.15 (3) |
| Microtubule-associated protein 2 | AEDVTAALAKQGL | C | 0.87 ± 0.06 (3) | ||
| Microtubule-associated protein tau | ADEVSASLAKQGL | C | 0.78 ± 0.24 (3) | 1.17 ± 0.16 (3) | 0.95 ± 0.12 (3) |
| Neurogranin | GRKGPGPGGPGGAGGARGGAGGGPSGD | C | 0.60 ± 0.10 (3) | 0.84 ± 0.03 (3) | 1.08 ± 0.30 (3) |
| Neurogranin | KGPGPGGPGGAGGARGGAGGGPSGD 2+ | C | 0.55 ± 0.20 (3) | 0.64 ± 0.02 (3) | 1.58 ± 0.37 (3) |
| Neurogranin | KGPGPGGPGGAGGARGGAGGGPSGD 3+ | C | 0.78 ± 0.01 (3) | 1.63 ± 0.37 (3) | |
| Neurogranin | RKGPGPGGPGGAGGARGGAGGGPSGD | C | 0.94 ± 0.20 (3) | 1.28 ± 0.55 (3) | 1.57 ± 0.38 (3) |
| Peptidyl-prolyl cis-trans isomerase A | ADDEPLGRVSF | C | 0.85 ± 0.14 (3) | 1.10 ± 0.02 (3) | |
| Peptidyl-prolyl cis-trans isomerase A | ADKVPKTAENF (1.59 ± 0.21) | C | 0.71 ± 0.14 (3) | ||
| Peptidyl-prolyl cis-trans isomerase A | ADKVPKTAENFR | C | 1.36 ± 0.30 (2) | ||
| Peptidyl-prolyl cis-trans isomerase A | ADKVPKTAENFRAL | C | 0.77 ± 0.08 (3) | 1.26 ± 0.20 (3) | |
| Peptidyl-prolyl cis-trans isomerase A | EDENFILKHTGPGILSM | C | 0.80 ± 0.10 (3) | ||
| Peptidyl-prolyl cis-trans isomerase A | ELFADKVPKTAENF | C | 1.19 ± 0.23 (2) | ||
| Peptidyl-prolyl cis-trans isomerase A | ELFADKVPKTAENFRAL | C | 0.98 ± 0.35 (3) | ||
| Peptidyl-prolyl cis-trans isomerase A | ITADDEPLGRVSF | C | 0.66 ± 0.16 (3) | 0.88 ± 0.13 (3) | |
| Peptidyl-prolyl cis-trans isomerase A | LFADKVPKTAENFRAL | C | 0.93 ± 0.15 (3) | ||
| Peptidyl-prolyl cis-trans isomerase A | FDITADDEPLGRVSF | C | 1.03 ± 0.22 (3) | ||
| Peptidyl-prolyl cis-trans isomerase A | LFADKVPKTAENF | C | 0.96 ± 0.22 (3) | ||
| Peptidyl-prolyl cis-trans isomerase FKBP1A | VFDVELLKLE | C | 0.84 ± 0.13 (3) | 1.03 ± 0.22 (3) | |
| Phosphatidylethanolamine-binding protein 1 | AGVTVDELGKVL | C | 1.11 ± 0.24 (2) | ||
| Phosphatidylethanolamine-binding protein 1 | AGVTVDELGKVLTPTQV | C | 1.40 ± 0.04 (3) | 0.81 ± 0.11 (3) | |
| Phosphatidylethanolamine-binding protein 1 | DDYVPKLYEQLSGK | C | 0.67 ± 0.08 (3) | 1.11 ± 0.17 (3) | 0.79 ± 0.06 (3) |
| Phosphatidylethanolamine-binding protein 1 | DGLDPGKLYTL | C | 1.06 ± 0.20 (3) | ||
| Phosphatidylethanolamine-binding protein 1 | GLDPGKLYTL | C | 0.77 ± 0.15 (3) | ||
| Phosphatidylethanolamine-binding protein 1 | GVTVDELGKVLTPTQV | C | 1.22 ± 0.20 (3) | ||
| Phosphatidylethanolamine-binding protein 1 | KGNDISSGTVL | C | 1.10 ± 0.12 (3) | ||
| Phosphatidylethanolamine-binding protein 1 | KGNDISSGTVLSDYVGSGPPSGTGL | C | 1.38 ± 0.04 (2) | ||
| Proenkephalin-A | SPQLEDEAKELQ | V | 0.69 ± 0.25 (3) | 0.83 ± 0.20 (3) | |
| ProSAAS | ASAPLVETSTPLRL | V | 1.20 ± 0.22 (2) | ||
| ProSAAS | SLSAASAPLVETSTPLRL | V | 0.92 ± 0.08 (3) | 1.20 ± 0.32 (3) | 0.84 ± 0.16 (2) |
| Secretogranin-1 | LLDEGHYPVRESPIDTA | V | 1.71 ± 0.15 (3) | ||
| Secretogranin-1 | SFARAPQLDL | V | 1.33 ± 0.33 (3) | 0.85 ± 0.31 (3) | |
| Somatostatin | ANSNPAMAPRE (methionine oxidation) | V | 1.73 ± 0.13 (3) | ||
| Somatostatin | SANSNPAMAPRE | V | 1.65 ± 0.13 (3) | 2.03 ± 0.65 (3) | 1.10 ± 0.31 (3) |
| Somatostatin | SANSNPAMAPRE (methionine oxidation) | V | 2.03 ± 0.47 (2) | 1.84 ± 0.52 (3) | |
| Synapsin-1 | AGGPPHPQLNKS | C | 0.97 ± 0.29 (3) | ||
| Tubulin beta-2A chain | SGPFGQIFRPDNF | C | 1.07 ± 0.22 (3) | 0.80 ± 0.02 (3) | |
| Tubulin beta-3 chain | DDEESEAQGPK | C | 2.23 ± 0.57 (2) | 1.60 ± 0.24 (2) |
| WT | THOP-/- | |
|---|---|---|
| Granulocytes (%) | 21.02 ± 0.63 | 24.22 ± 0.55 ** |
| Monocytes (%) | 6.62 ± 0.17 | 7.47 ± 0.27 * |
| Lymphocytes (%) | 72.46 ± 0.62 | 68.32 ± 0.80 ** |
| Platelets 109/L | 1509.00 ± 73.12 | 1159.00 ± 108.50 * |
| Hemoglobin g/L | 140.20 ± 1.74 | 135.30 ± 3.57 |
| WBC 109/L | 10.09 ± 1.00 | 8.64 ± 0.71 |
| RBC 1012/L | 9.62 ± 0.10 | 9.25 ± 0.28 |
| RDW | 12.80 ± 0.21 | 13.12 ± 0.29 |
| MCV fL | 46.78 ± 0.23 | 47.37 ± 0.26 |
| MCHC g/L | 311.22 ± 0.89 | 308.50 ± 1.54 |
| Gene | Sequence | Amplicon (bp) | Accession Number |
|---|---|---|---|
| Neurolysin (Nln) | Fwd: CACCCTATCACAGACGAGCT Rev: GGGACTGGTCAACTTTGCTC | 106 | NM_029447 |
| Neprilysin (NEP) | Fwd: CCTGAACTTTGCCCAGGTGT Rev: GCGGCAATGAAAGGCATCTG | 148 | NM_001289462.1 |
| Angiotensin I Converting Enzyme 1 (ACE1) | Fwd: ACCCTAGGACCTGCCAATCT Rev: CGTGAGGAAGCCAGGATGTT | 164 | NM_207624.5 |
| Prolyl oligopeptidase (POP) | Fwd: GGGTGCTCCGACACTAAACA Rev: GACGGGTACTGGATGTCGTC | 98 | NM_011156.3 |
| Insulin Degrading enzyme (IDE) | Fwd: GTCCATGTTCTTGCCAGGGA Rev: TTCACGAGGGGAAACAGTGG | 161 | NM_031156.3 |
| Dipeptidyl peptidase 4 (DPP4) | Fwd: GACGGCAGAGGAAGTGGTT Rev: CGCTTGCTATCCACAAATCCC | 134 | NM_010074.3 |
| Proteasome subunit beta 5 (ProtB5) | Fwd: CCAAACTGCTCGCTAACATGG Rev: GTTCCCCTCGCTGTCTACG | 119 | NM_011186.1 |
| 5-Hydroxytryptamine (serotonin) receptor 2A (5-HT2A) | Fwd: GTTTCCTTGTCATGCCCGTG Rev: CCAGGTAAATCCAGACGGCA | 93 | NM_172812.2 |
| Dopamine D2receptor (DRD2) | Fwd: ATGGGAGTTTCCCAGTGAACA Rev: ATGGGGCTATACCGGGTCC | 115 | NM_010077.2 |
| Hypoxanthine guanine phosphoribosyl transferase (HPRT) |
Fwd: TGCTGACCTGCTGGATTACA Rev: TTTATGTCCCCCGTTGACTGA | 120 | NM_013556.2 |
| Ribosomal protein L19 (RPL19) | Fwd: CAATGCCAACTCCCGTCA Rev: GTGTTTTTCCGGCAACGAG | 102 | NM_009078.2 |
| Glyceraldehyde-3-phosphatedehydrogenase (GAPDH) | Fwd: GTGCAGTGCCAGCCTCGTCC Rev: CAGGCGCCCAATACGGCCAA | 75 | BC085275.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, N.B.; Franco, R.D.; Camarini, R.; Munhoz, C.D.; Eichler, R.A.S.; Gewehr, M.C.F.; Reckziegel, P.; Llanos, R.P.; Dale, C.S.; da Silva, V.R.O.; et al. Thimet Oligopeptidase (EC 3.4.24.15) Key Functions Suggested by Knockout Mice Phenotype Characterization. Biomolecules 2019, 9, 382. https://doi.org/10.3390/biom9080382
dos Santos NB, Franco RD, Camarini R, Munhoz CD, Eichler RAS, Gewehr MCF, Reckziegel P, Llanos RP, Dale CS, da Silva VRO, et al. Thimet Oligopeptidase (EC 3.4.24.15) Key Functions Suggested by Knockout Mice Phenotype Characterization. Biomolecules. 2019; 9(8):382. https://doi.org/10.3390/biom9080382
Chicago/Turabian Styledos Santos, Nilton B., Roseane D. Franco, Rosana Camarini, Carolina D. Munhoz, Rosangela A. S. Eichler, Mayara C. F. Gewehr, Patricia Reckziegel, Ricardo P. Llanos, Camila S. Dale, Victoria R. O. da Silva, and et al. 2019. "Thimet Oligopeptidase (EC 3.4.24.15) Key Functions Suggested by Knockout Mice Phenotype Characterization" Biomolecules 9, no. 8: 382. https://doi.org/10.3390/biom9080382