1. Introduction
Dengue fever, endemic in tropical and subtropical regions, is a significant public health problem in Southeast Asia [
1], and no effective vaccine or specific medication is available [
2]. Over 33 million cases of dengue fever with several death cases are reported every year [
3]. Dengue fever is caused by the mosquito-borne dengue virus (DENV), which is an enveloped, single-stranded, positive-sense RNA flavivirus. Four distinct, but closely related, dengue serotypes have been identified. Infection by a single serotype provokes a mild to high fever for a few days, after which the patient usually recovers steadily and gains a long-lasting immunity against the infecting serotype. However, subsequent heterotypic infections increase the risk of developing dengue haemorrhagic fever (DHF) and shock syndrome (DHS) with a ~5% mortality rate and severe complications [
4,
5].
DENV’s genome comprises ten genes encoding for the capsid (C), premembrane (prM), envelope (E), and seven nonstructural (NS) proteins [
6]. The envelope is formed by the association of 180 E-protein units and constitutes the outer surface of the dengue virus particle. The E-protein mediates virus–host attachment and also contains the binding site for neutralizing antibodies [
7,
8]. It is composed of three domains: (ED1–3) [
9].
The ED3 domain of E-protein is a ∼100-residue globular ß-sheet protein that adopts the fold of an Ig-like domain. It contains two putative dominant epitopes recognized by antibodies, and a region that can affect the binding to the heparin sulfate receptor, which drives the virus entry into the host cell [
9,
10]. As such, ED3 plays a critical role in viral infection. Phylogenetic studies show that DEN4 is the oldest of the four DEN viruses [
11]. Surprisingly, only eight amino acid mutations in ED3 separate the two most divergent DEN4 strains, among which five have been implicated in the emergence of the human DEN4 virus [
11,
12] (
Supplementary Materials, Figure S1 and Table S1). Recent X-ray crystallography (DEN3-ED3, DEN4-ED3) [
13,
14] and nuclear magnetic resonance (NMR) (DEN4-ED3) [
15] analysis demonstrated that dengue ED3 remains natively folded even when it is isolated from the rest of the E-protein. Moreover, previous studies indicated that residues in the hydrophobic core of ED3 were not fully conserved among the four serotypes [
13] and that a rearrangement of the side-chains interdigitization ensured the stabilization of the protein core and thus of the natively folded structure of DEN-ED3 across serotypes [
14]. More generally, the Ig-fold is a classical fold found in several proteins with unrelated functions, and it would be of interest to determine whether proteins with Ig-fold but different core packing exhibit similar folding properties at the residue level [
16].
The combination of high hydrostatic pressure with NMR has proven to be a powerful tool to obtain high-resolution structural information about protein folding [
17,
18,
19,
20,
21,
22,
23,
24,
25]. Indeed, multidimensional NMR intrinsically provides multiple residue-specific probes, while the reversibility of high-pressure folding/unfolding experiments ensures the proper characterization of thermodynamic parameters. In addition, contrary to chemical denaturants or temperature, pressure does not affect the protein uniformly: it acts by reducing volume, primarily by eliminating solvent-excluded voids. Since these voids are heterogeneously distributed throughout the protein 3D structure, certain regions are more pressure-dependent than others, thus favoring the characterization of folding intermediates [
17,
26].
In the present study, we used high-pressure NMR spectroscopy to monitor the unfolding of ED3 from dengue-4 virus (DEN4-ED3) and shed insight into the structural details of its folding pathway. In particular, we found that the folding pathway of DEN4-ED3 shares similarities with that of the I27 module from the giant multimodular protein Titin found in the striated muscle cells [
27], which also adopts an Ig-like fold. In both proteins, unfolding starts with the disruption of the N- and C-terminal strands on the edge of the ß-sandwich, but the Transition State Ensemble (TSE) of DEN4-ED3 was found to be significantly more hydrated, and hence less structured, than that of Titin I27.
2. Results
2.1. Setting the Physical–Chemical Conditions for ED3 Unfolding
The DEN4-ED3 domain did not completely unfold in the NMR buffer, at pH 7, in a 5–40 °C temperature range, and in the pressure range allowed by the experimental set-up (1–2500 bar). Thus, we decided to destabilize the protein slightly by adding a subdenaturing concentration of a chaotropic reagent (guanidinium chloride).
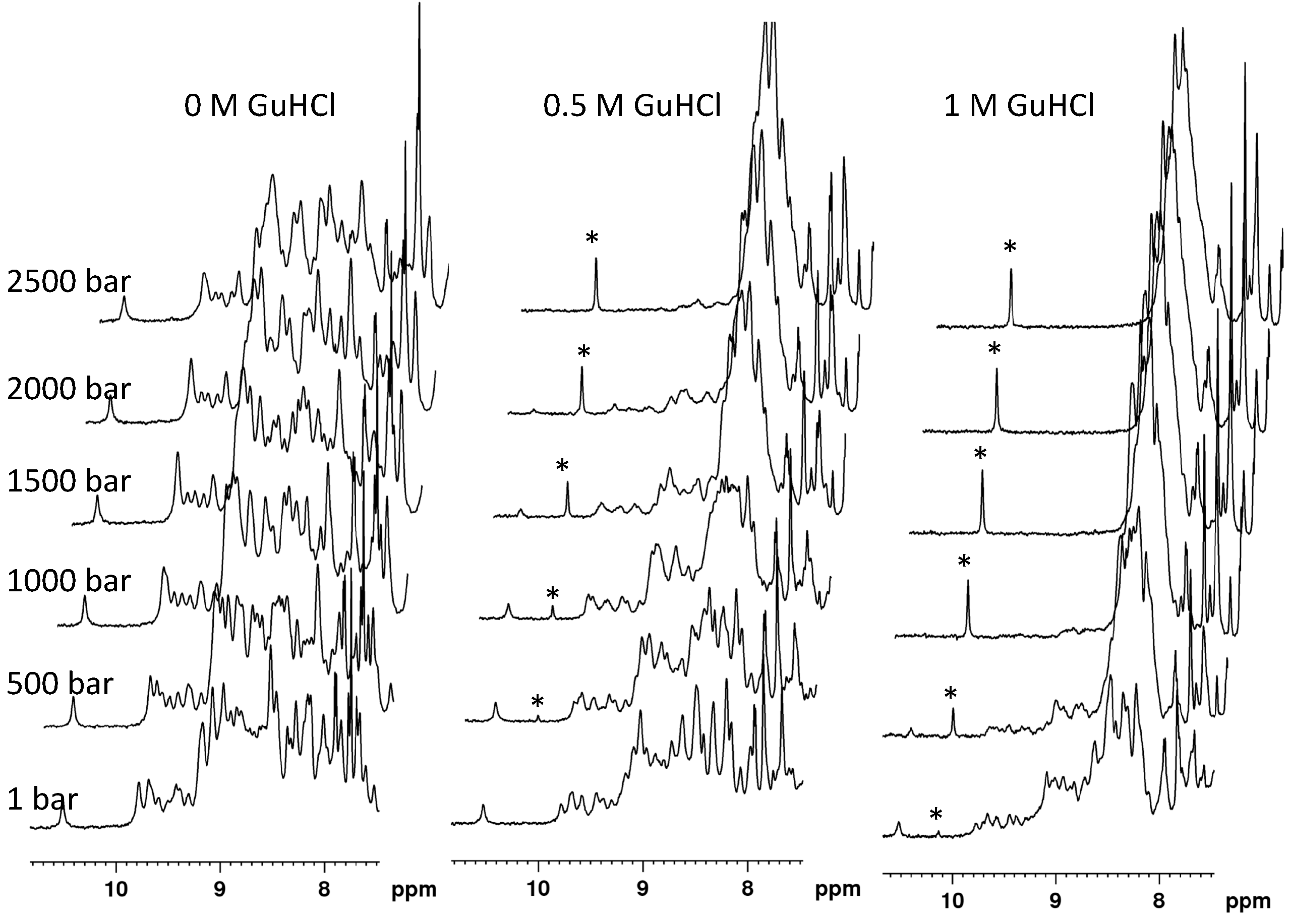
The indolic proton HN resonances of the native and unfolded protein species were used to monitor DEN4-ED3 unfolding with pressure, using 1D NMR spectroscopy (
Figure 1): they are well individualized and do not overlap with HN amide resonances in the 1D spectrum of DEN4-ED3, and thus easy to follow. After different trial-and-error assays, adjusting the concentration of GuHCl and the temperature, we found that the protein completely unfolds in the 1–2500 bar pressure range at pH 7 (Tris 10 mM), 10 °C, and at a concentration of GuHCl of 0.5 M (
Figure 1).
Note that the concomitant decrease of the folded-state indole HN resonance of Trp103 and the increase of the corresponding resonance in the unfolded state strongly support a slow exchange between the chemical environments this residue experiences in the folded and unfolded states: the fit of the pressure dependence of these two resonances with a two-state equilibrium (Equation (1)) yields ∆Vf0 and ∆G0 values within experimental uncertainty.
2.2. Assignment of the Amide Protons
The solution structure of DEN4-ED3 was solved previously by NMR (Protein Data Bank (PDB) entry: 2h0p) using a complete set of {
1H,
13C,
15N} triple-resonance experiments for chemical-shifts assignment (deposited at BRMB: 7087) [
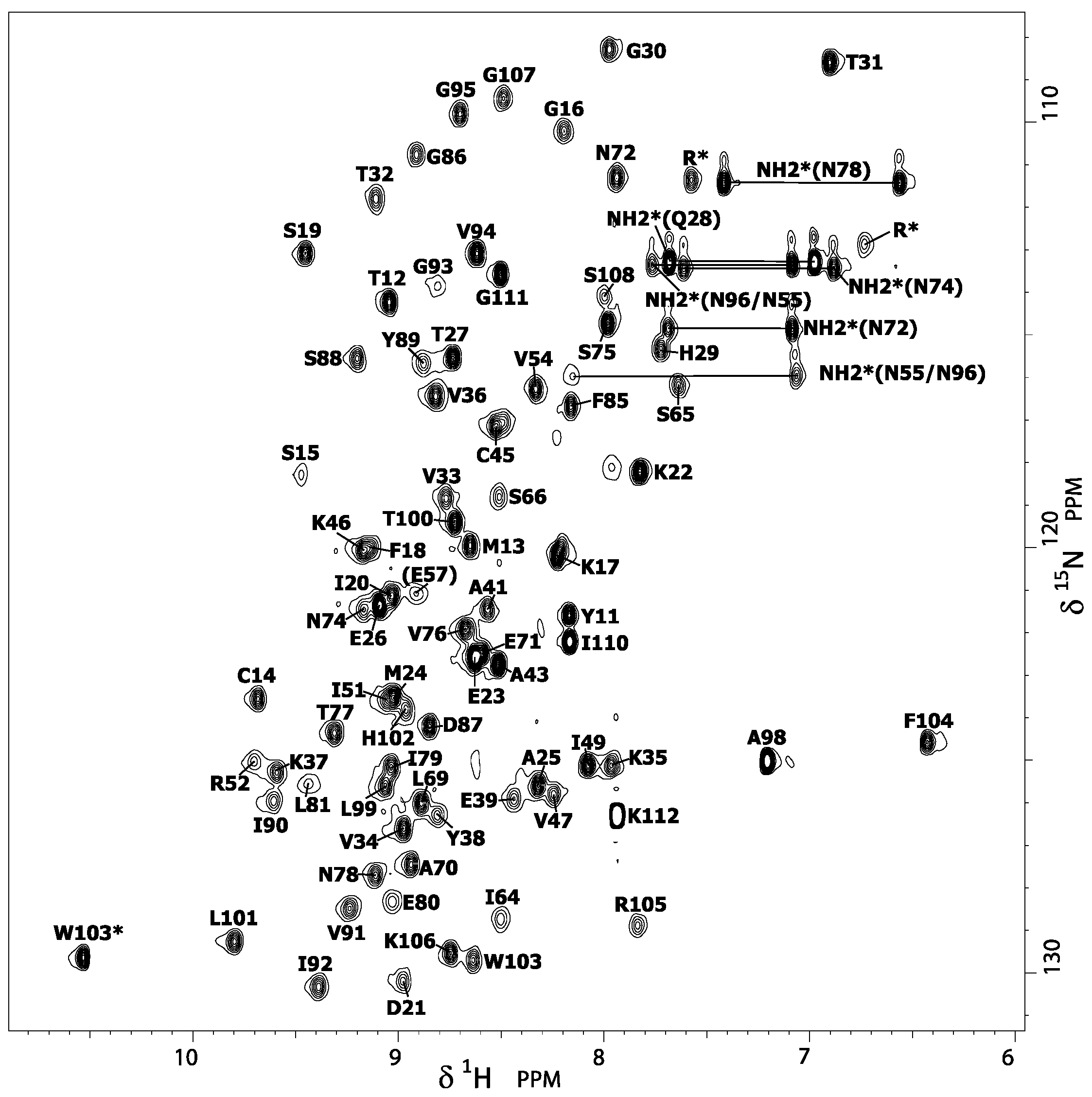
15]. This structure was determined at 25 °C and pH 7.5 (50 mM Tris buffer, 50 mM NaCl), under physical and chemical conditions slightly different than those used in the present denaturation study (pH 7, 10 °C, no salt, and 0.5 M GuHCl), justifying the reassignment of the amide group resonances. Thus, from
15N-Edited nuclear Overhauser effect spectroscopy (NOESY) and total correlation spectroscopy (TOCSY) three-dimensional (3D) experiments, the classical sequential assignment strategy was used to reassign the amide proton and nitrogen resonances (and, besides, the Hα resonances) displayed in the {
1H,
15N} HSQC spectra further used for the denaturation study (
Figure 2). These minimal changes in chemical and physical conditions result in very slight differences in chemical shifts (
Supplementary Materials, Figure S2), suggesting that the Ig-like 3D structure of DEN4-ED3 (
Figure 3A) is not altered by the presence of guanidinium chloride. Thus, the 3D structure previously determined by Volk et al. [
15] was used as a template for displaying the results obtained in the present study.
2.3. Local Stability Probed by Proton/Deuteron Exchange Measurements.
Local stability is important information when studying protein unfolding, since it is often related to partial unfolding of the protein. H/D exchange measurements can provide such information since they give access to information on the strength of the H-bonds, hence on the stability of the secondary structure elements. H/D exchange for DEN4-ED3 was investigated in our experimental conditions (pH 7 and T = 10 °C), but in the absence of chemical denaturant, to explore the local stability of the protein. To this aim, {
1H,
15N} HSQC spectra were recorded with time on a protein sample freshly dissolved in D
2O (see Materials and Methods for details). After protein dissolution, 25 amide cross-peaks were still apparent in the first HSQC recorded, but accurate fit with an exponential decay can be obtained for the HSQC cross-peak intensity of only 22 residues (18 involved in the H-bonds stabilizing the ß-sheets) over 107 nonproline residues. Their corresponding protecting factors (PFs) were calculated from the exchange rate constants obtained from the fit [
28] (
Supplementary Materials, Table S2). The local stability of the different ß-sheets was then estimated by the average value of the protecting factors (<PF>) calculated over the amide protons involved in the main-chain H-bond stabilizing the ß-sheet (
Figure 3).
These results reveal that the ß4-ß8 and ß8-ß9 sheets, which form a triple-stranded ß-sheet on one face of the ß-sandwich, appear significantly more stable (4.4 < average log(PF) < 4.6) than the ß1-ß2 and ß2-ß7 sheets on the other face of the ß-sandwich (3.4 < average log(PF) < 4). Although amide cross-peaks of residues involved in the main-chain stabilizing H-bonds were present in the first HSQC spectrum recorded on the freshly dissolved protein sample, the corresponding PF cannot be measured for the short ß5-ß7 sheet (the ß5 strand consists only of two residues: Arg62-Ile63) due to a rapid decrease of the cross-peak intensities with time. This means that this sheet is extremely unstable and probably only transiently formed. Note that the ß3-ß6 sheet, also extremely short since it involves only two residues in each strand (Cys45-Lys46 on ß3 strand, Leu69-Ala70 on ß6 strand), shows a stability only slightly weaker than observed for the ß-sandwich (aver. log(PF) ≈ 3), even if it does not contribute to its structure by itself.
2.4. High-Pressure Unfolding Monitored with Nuclear Magnetic Resonance Spectroscopy: Steady-State Studies
To further probe the structural details of DEN4-ED3 folding, two-dimensional (2D) {
1H,
15N} HSQC spectra of
15N uniformly labeled DEN4-ED3 in the presence of 0.5 M guanidinium chloride were recorded as a function of pressure (
Figure 4). As typically observed, the overall intensity of each native state peaks decreased as a function of pressure, while the intensity of the corresponding unfolded state peaks, centered around 8.5 ppm in the proton dimension, concomitantly increased. This strongly suggests a slow equilibrium between the chemical environment of each residue in the native and unfolded state on the NMR timescale, as well as a two-state transition between each native/unfolded pairwise cross-peak during the unfolding process. Thus, locally, the loss of intensity for each native state cross-peak can be interpreted as a two-state transition, even though the global protein unfolding does not conform to such a model [
17]. Since resonance assignments were available only for the native state structure of DEN4-ED3, the decrease in intensity with pressure for each corresponding cross-peak was fitted to a two-state, pressure-induced unfolding model as described in the Materials and Methods (Equation (1),
Figure 4), yielding residue-specific values for the apparent volume change (∆
Vf0) and apparent free energy (∆
G0) of unfolding [
17] (
Figure 5). Accurate fitting (
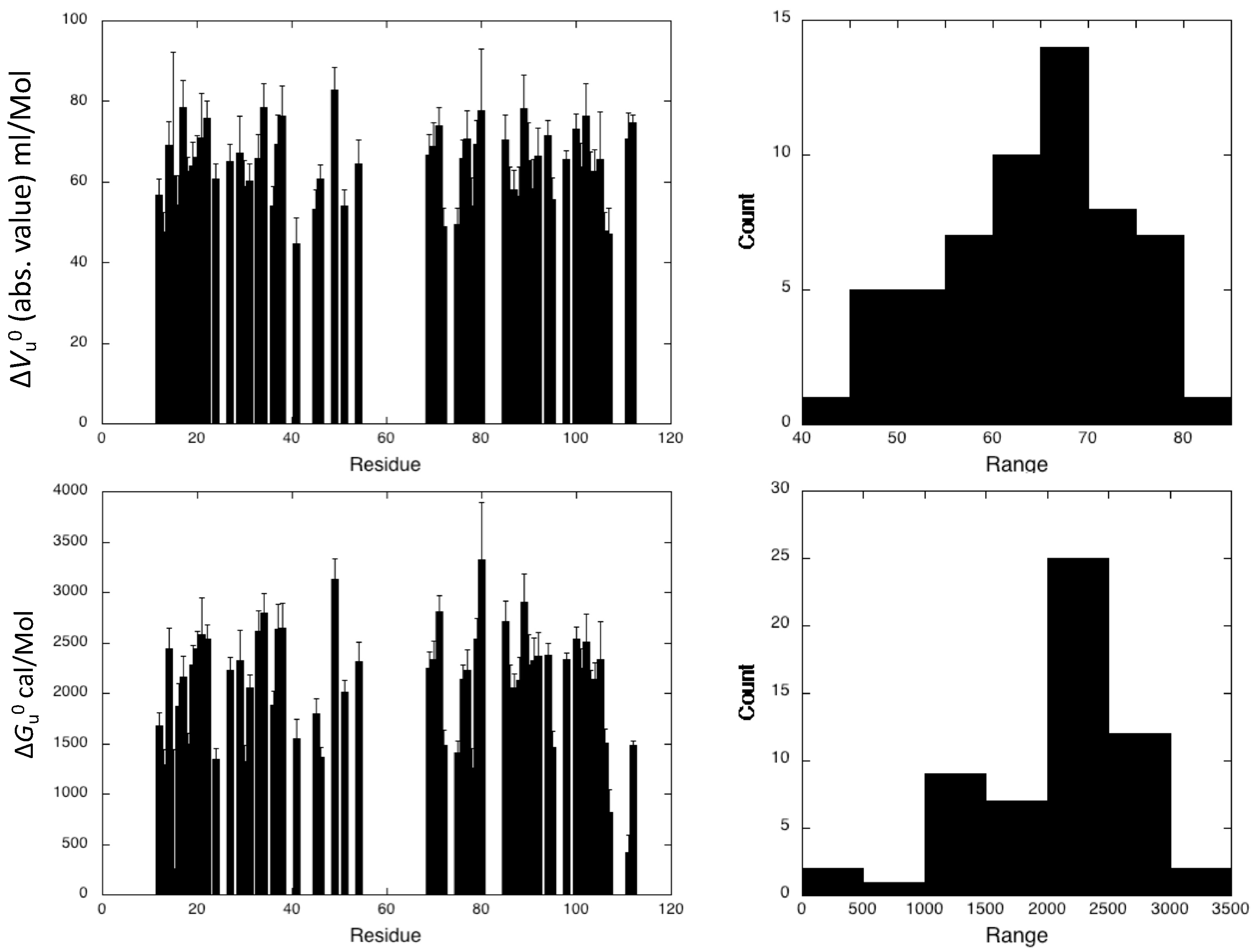
Supplementary Material, Figure S3) was obtained for 58 residues of the 107 nonproline residues, providing a substantial number of local probes for the description of the DEN4-ED3 folding pathway. The fitted curves correspond to residues for which the cross-peaks do not overlap either in the folded state of the protein or in between the folded and unfolded states. Residues that exhibit cross-peaks with very low intensity at atmospheric pressure, probably due to slow or intermediate exchange between different populations, cannot be fitted accurately and were discarded from the analysis.
The DEN4-ED3 exhibits moderate stability at 0.5 M guanidinium chloride, with an average value <∆
Gu0> = 2.23 ± 0.08 kcal/mol. Besides, an average <∆
Vu0> value of −64 ± 9 mL/mol was obtained for the construct. While the two-state model was adequate for all individual residue-specific unfolding curves, different profiles were observed for different residues, demonstrating significant deviation from two-state behavior of the global unfolding for the protein. The asymmetric distributions of apparent ∆
Vu0 and apparent ∆
Gu0 (
Figure 5) are indicative of partial unfolding of the ED3 domain at intermediate pressures.
Fractional contact maps were constructed for DEN4-ED3 to visualize which regions of the protein become disordered at intermediate pressures [
17]. The probability of contact for any pair of residues,
Pi,j, at a given pressure was defined as the geometric mean of the fractional intensity of the HN resonances of each of the two residues in the folded state at the same pressure (
Pij = (
Pi ×
Pj)
1/2), extracted from the normalized residue-specific denaturation curves [
21].
The pressure dependence of the contact maps shows that the first region affected by an increase of pressure implicates one edge of the ß-sandwich, namely the junction made by the ß-strands 1 and 9, corresponding to the N- and C-termini of the molecule. A significant (
Pij ≤ 50%) loss of contacts is already apparent at 600 bar (
Figure 6), and up to 1000 bar, this partial unfolding concerns mainly this region. Above this pressure, partial unfolding extends throughout the ß-sandwich, probably as a result of water penetration in the hydrophobic core of the protein. Interestingly, unfolding propagates first on one face of the ß-sandwich (strands 1, 2, 7), while the other face (strands 4, 8, 9) remains almost unaffected until 1400 bar. Note that, from proton/deuteron exchange experiments, the ß1-ß2-ß7 sheet is less stable than the ß4-ß8-ß9 sheet; thus, partial unfolding seems to map to the zone of fragility in the ß-sandwich.
2.5. High-Pressure Unfolding Monitored with Nuclear Magnetic Resonance Spectroscopy: Kinetics Studies
To further probe the folding reaction, we performed a kinetics study by recording series of 2D {
1H,
15N} SOFAST-HMQC experiments (2 min measuring time each) [
29] over 2 h after 200 bar positive pressure jumps. From the exponential decrease in intensity with time of each native state cross-peaks in these real-time 2D NMR experiments, it is in principle possible to extract the folding/unfolding residue-specific relaxation time τ. This measurement can be repeated after similar P-jumps between different pressures, and the different values of τ obtained further plotted versus pressure, yielding the so-called residue-specific “chevron plots” that allow extracting residue-specific rate constants of folding
kf and unfolding
ku, as well as residue-specific activation volumes of folding (∆
Vf‡) and unfolding (∆
Vu‡) [
24]. Among these results, the residue-specific values of ∆
Vf‡ are of special importance, since they provide an interesting description of the hydration state of the transition state ensemble (TSE), and more precisely, which residue is hydrated (volumetrically close to the unfolded state) or dehydrated (volumetrically close to the folded state) at the TSE. Unfortunately, in the case of DEN4-ED3, the weak exponential decrease in intensity with time observed for most of the native state cross-peaks after P-jumps in the real-time 2D NMR experiments precluded an accurate determination of the residue-specific relaxation times
τ, and, consequently, an accurate determination of the residue-specific values of ∆
Vf‡.
Nevertheless, the NMR measurement of global folding kinetic parameters remains possible. For this, we recorded series of 1D proton NMR spectra after P-jumps [
25,
27]. The proton resonance at 0.96 ppm, corresponding to unfolded-state methyl proton resonances, was used as a probe to measure the global relaxation times τ (
Figure 7A). Indeed, it provides a much more sensitive probe to monitor the folding/unfolding reaction than the amide cross-peaks that correspond each to a single proton, but at the expense of the sequence specificity. Thus, global values of
τ can be accurately determined from the exponential growth of this resonance after P-jumps (
Figure 7B,C), yielding reliable global values for ∆
Vf‡, ∆
Vu‡,
ku, and
kf.
A value of ∆
Vf‡ = 20.7 ± 1.1 mL/mol for the global apparent activation volumes of folding was obtained for DEN4-ED3 from the chevron plot (
Figure 7D). This corresponds to about 40% of the value of ∆
Vf0 at equilibrium when referring to the same methyl resonance in 1D proton NMR steady-state experiments recorded under the same conditions (∆
Vf0 = 50.3 ± 2.6 mL/mol,
Supplementary Materials, Figure S4). This result shows that less than half of the void volumes present in the folded state of DEN4-ED3 are formed already in the TSE, implying that the TSE is substantially hydrated. Besides, we obtained from the same chevron plot a value of
kf0 = 1.14 (± 0.08) × 10
−3 s
−1 for the average kinetic constant of folding at ambient pressure. Note that, because amide and methyl groups probe different chemical environments, the apparent global value of ∆
Vf0 obtained from the unfolded-state methyl band differs significantly from the apparent residue-specific values of ∆
Vf0 measured from the native state amide resonances.
3. Discussion
In this study, we monitored the pressure-induced unfolding of DEN4-ED3 with high-resolution NMR spectroscopy. From the analysis of the residue-specific denaturation curves, the first event in DEN4-ED3 unfolding was found to be the rupture of the junction between the N- and C-terminal ß-strands 1 and 9, on one edge of the ß-sandwich. Interestingly, this behavior is rather similar to what has been recently shown for the I27 domain of the multimodular giant protein Titin from the striated muscle cell, which also adopts an Ig-like fold, but with a significantly different sequence (sequence identity <7%) [
27]. In Titin I27, the N- and C-termini are involved in ß-sheet structures: a small, distorted antiparallel (AB) ß-sheet, and a longer parallel (A’G) ß-sheet (where the capital letters stand for the ß-strands numbering, following the usual convention for Ig-like modules). The local pressure disruption of these sheets starts around 600 bar and concerns mainly these secondary structures at pressures up to 1000 bar. The local unfolding of these short ß-sheets has been proposed as the starting event of Titin Ig-like domain unfolding, yielding an intermediate folding state where the N-terminal strand is detached from the ß-sandwich. This partial unfolding of Ig-like modules of Titin in the I-band of the sarcomere has been proposed to contribute to the passive elasticity of striated muscle [
30,
31]. Beyond 1000 bar, the entire protein unfolds, due to water penetration into the hydrophobic core of the molecule. In the case of DEN4-ED3, the N- and C-terminal strands adopt a similar parallel orientation but do not form a parallel ß-sheet on one edge of the ß-sandwich, as for Titin I27. This might explain in part the difference in stability between the two proteins: addition of 1.7 M guanidine was necessary to decrease the stability of I27 sufficiently to ensure complete denaturation in the pressure range allowed by our experimental set-up (1–2500 bar), versus only 0.5 M in the case of DEN4-ED3. Despite the difference in stability, the starting event of unfolding remains strikingly similar for the two proteins: the disruption of the edge of the ß-sandwich bounded by the N- and C-terminal strands. As for Titin I27, the partial unfolding of DEN4-ED3 concerns only this region for a significant pressure range (600 to 1000 bar), suggesting that a reasonably stable intermediate state exists in the folding landscape of DEN4-ED3, where the N- and C-terminal strands are only loosely bound to the ß-sandwich.
Above 1000 bar, unfolding extends to the entire molecule, but in a different manner than for Titin I27. In the case of the I27 Ig-like module, the folded structure is completely destroyed above 1000 bar. In contrast, DEN4-ED3 unfolding progresses in a more stepwise manner, first affecting one sheet (ß1-ß2-ß7, or A-B-E, following the Ig-fold convention) of the ß-sandwich, with the structure of the second one (ß4-ß8-ß9, or C-F-G, following the Ig-fold convention) remaining preserved until a pressure of 1400 bar. This suggests that under hydrostatic pressure, water does not progress in a similar way within the hydrophobic core of the two proteins. DEN4-ED3 presents the characteristic core organization described by Clarke et al. [
32] for Ig-like domains; the buried hydrophobic residues borne by strands 2 (B, in the usual Ig-fold convention) and 8 (F) are closely entangled and form the “spinal column” of the core, whereas hydrophobic residues from strands 4, 7, and 9 (C,E, and G) pack onto this central scaffold (
Figure 8). Even if a similar hydrophobic core has been reported for Titin I27 [
27], closer inspection reveals some differences: strand C contributes also to the spinal column of the hydrophobic core, with the indole ring of Trp34 being inserted between the buried hydrophobic residues from strands B and F. The remaining hydrophobic residues from strands E and G make a dense network of hydrophobic interactions around this compact core, mainly organized around Trp34 (
Figure 8). It is tempting to speculate that the more concerted unfolding transition for Titin I27 is a consequence of the more integrated packing patterns of its core. Similarly, the difference in stability between the two proteins probably originates from the different organization of their hydrophobic core. Nevertheless, only extensive molecular dynamic simulations could bring precise insights on the side-chain reorganization in the core of the molecule under water progression.
This distinct core organization is likely responsible for the different hydration states of the TSEs for the two proteins as well. Indeed, kinetics experiments show that the TSE is considerably more hydrated, and hence less organized, in the case of DEN4-ED3 (∆
Vf‡ ≈ 40% of ∆
Vf0) than for Titin I27 (∆
Vf‡ ≈ 95% of ∆
Vf0). For both proteins, residue-specific values of activation volumes cannot be obtained, thus preventing definition of which part of the molecule is disordered and in interaction with solvent, and which part is ordered and dehydrated at the folding barrier. Nevertheless, based on ø value analysis, Fowler and Clarke [
16] showed that the structure of I27 at the TSE was close to that of the folded state: only the A’ and G strands were completely unstructured, with ø values lower than 0.2, while all other strands of the ß sandwich were structured to some extent. This means that the structure of I27 at the TSE corresponds more or less to the structure of the intermediate state, as defined by HP-NMR analysis. Similarly, it is tempting to think that the 40% of dehydrated structures in the TSE might correspond to the ß4-ß8-ß9 (C-F-G) triple-stranded ß-sheet, the hydrated part consisting of the rest of the molecule, including the ß1-ß2-ß7 (A-B-E) sheet that unfolds at lower pressure.
Besides, our P-jump experiments yield a value of
kf0 = 1.14 (± 0.08) × 10
−3 s
−1 for the rate constant for folding of DEN4-ED3, more than one order of magnitude lower than the value previously measured for Titin I27 (
kf0 = 2.24 (± 0.11) × 10
−2 s
−1) [
27]. This seems to corroborate the observation made by Clarke et al. [
32], who found a positive linear correlation between the logarithm of the rate constant for folding and the stability of Ig-like protein.
5. Concluding Remarks
In this study, high-hydrostatic-pressure NMR was used to analyze at a residue-level resolution the folding pathway of DE4-ED3, an Ig-like protein domain from the dengue virus envelope. Interestingly, we found some similarity with the folding pathway of I27 module, another Ig-like domain found in Titin, a protein from the sarcomere of striated muscle cell. Notably, they share a common folding intermediate where the two N- and C-terminal strands are detached from the ß-sandwich. This finding represents an experimental confirmation of previous theoretical studies suggesting that folding pathways are conserved in protein families [
41,
42]. On the other hand, the kinetic studies performed on the two proteins show that, whereas the two proteins fold in a similar way, the structure of the Transition State Ensemble can be very different: dehydrated, with a structure close to the folded state in the case of Titin I27, or relatively hydrated, with an intermediary structure between the folded and unfolded state for DEN4-ED3.
In conclusion, it is of interest to see that two proteins with unrelated function that adopt a similar topology and a similar 3D fold (an Ig-like fold, in the present case) might have similar folding energy landscape, including similar folding intermediates, even though their sequences are completely different. This difference in primary structure might be responsible for the different folding kinetics observed for the two proteins, resulting in an earlier folding barrier for DEN4-ED3, and hence different structures at the Transition State Ensemble.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







