Modelling of Molasses Fermentation for Bioethanol Production: A Comparative Investigation of Monod and Andrews Models Accuracy Assessment

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism and Fermentation Medium
2.2. Fermentation
2.3. Analytical Methods
2.4. Models Theory
2.4.1. Growth Models
2.4.2. Substrate and Product Models
2.5. Models Simulations and Validation
3. Results and Discussion
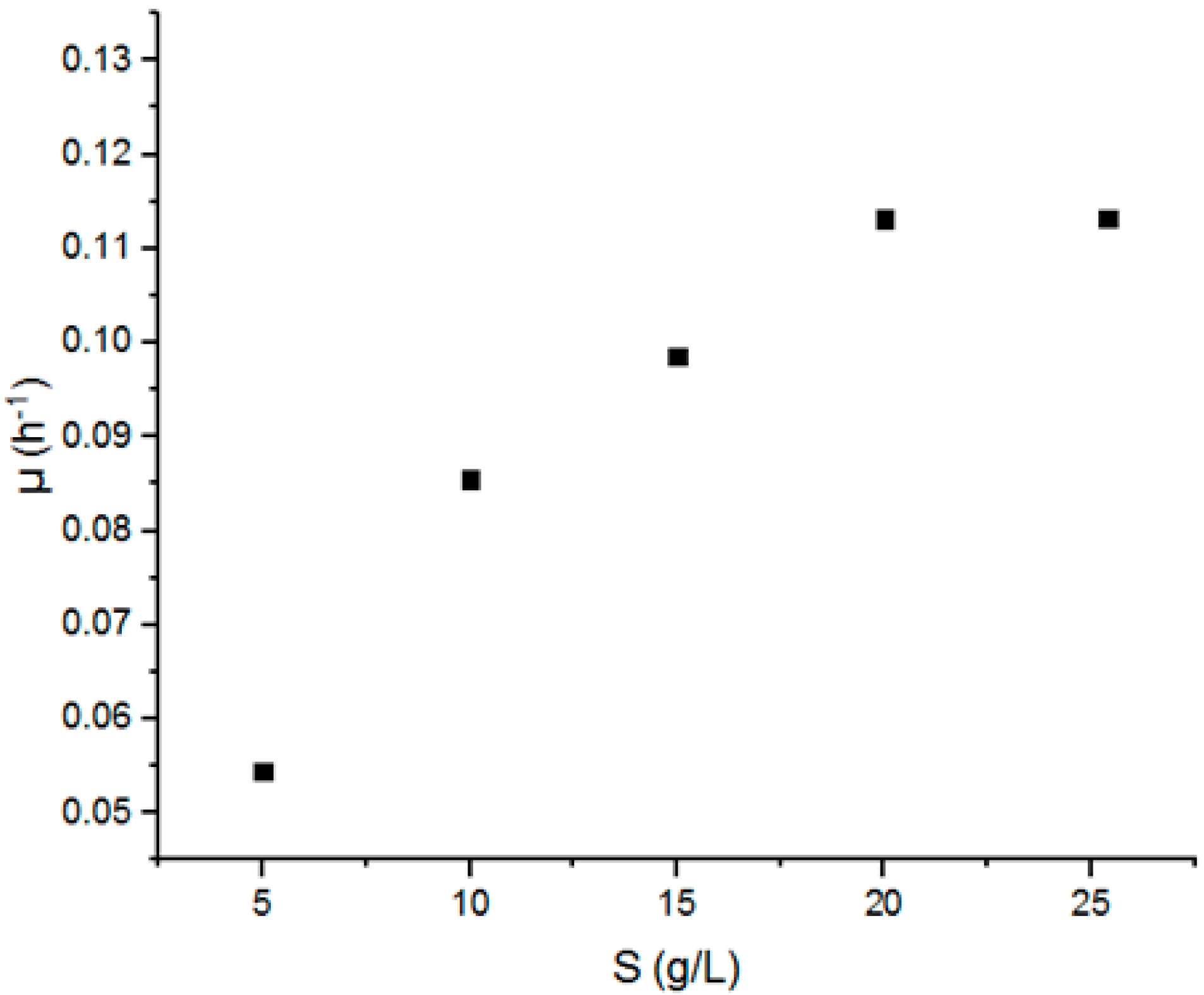
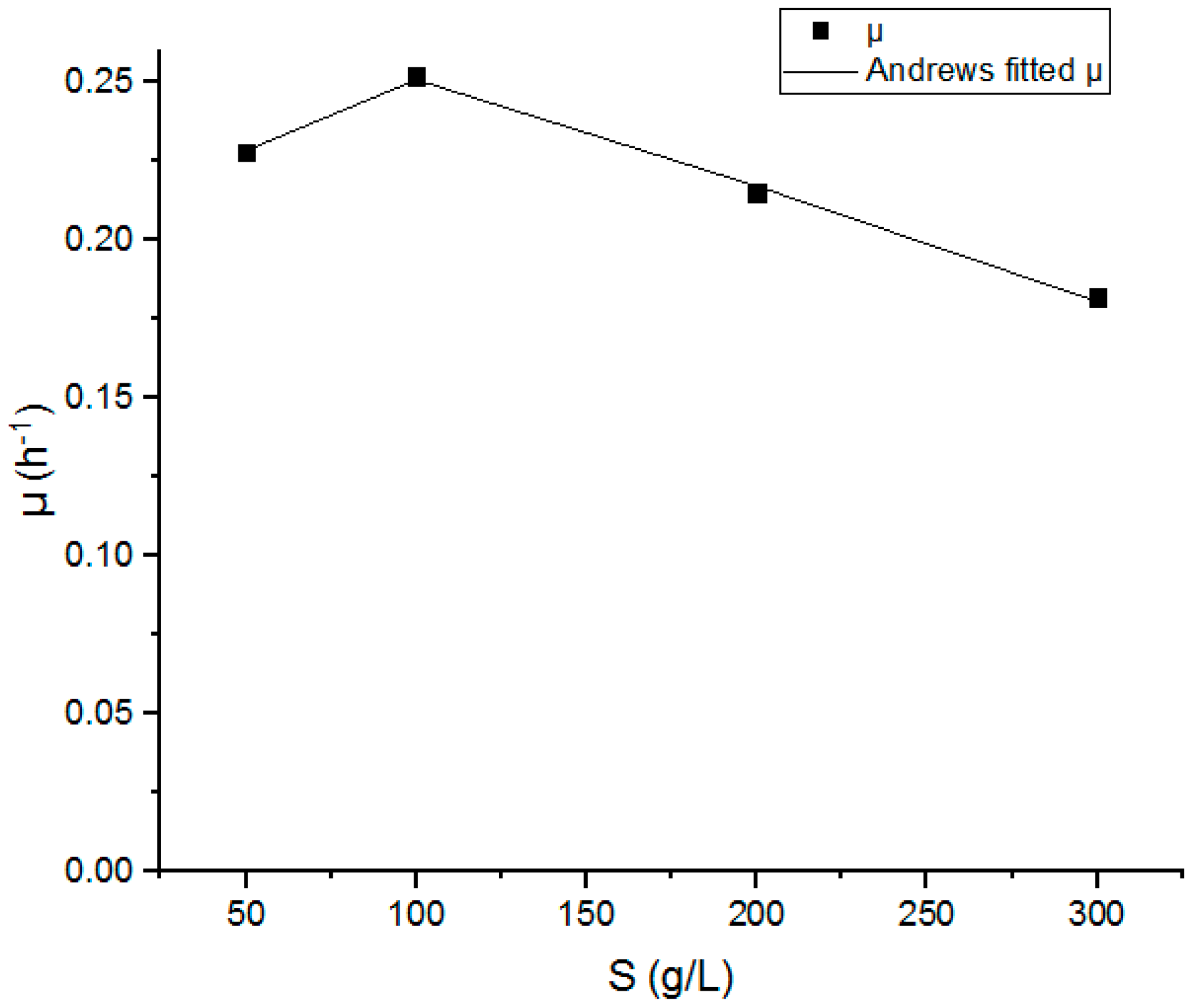
3.1. Calculation of Kinetics Parameters
3.2. Calculation of Yield Coefficients
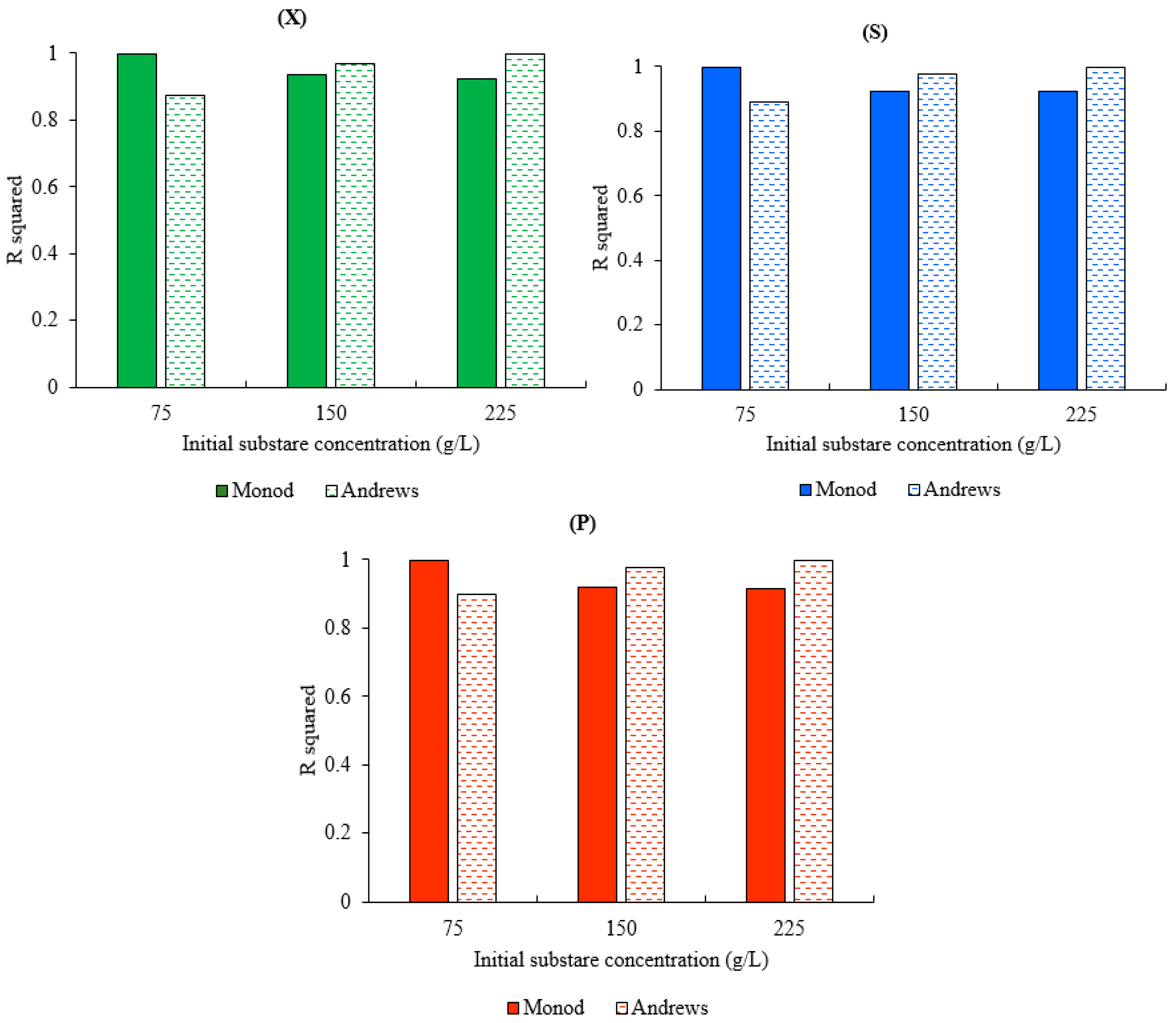
3.3. Model Validation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Rahim, N.A.; Hosenuzzaman, M.; Saidur, R.; Mahbubul, I.M.; Rashid, M.M. Energy savings in the combustion based process heating in industrial sector. Renew. Sustain. Energy Rev. 2012, 16, 4527–4536. [Google Scholar] [CrossRef]
- Neto, M.R.B.; Carvalho, P.C.M.; Carioca, J.O.B.; Canafístula, F.J.F. Biogas/photovoltaic hybrid power system for decentralized energy supply of rural areas. Energy Policy 2010, 38, 4497–4506. [Google Scholar] [CrossRef]
- Nel, W.P.; Cooper, C.J. Implications of fossil fuel constraints on economic growth and global warming. Energy Policy 2009, 37, 166–180. [Google Scholar] [CrossRef]
- Hoel, M.; Kverndokk, S. Depletion of fossil fuels and the impacts of global warming. Resour. Energy Econ. 1996, 18, 115–136. [Google Scholar] [CrossRef]
- Goldemberg, J. The promise of clean energy. Energy Policy 2006, 34, 2185–2190. [Google Scholar] [CrossRef]
- Jatoi, A.S.; Parkash, A.; Aziz, S.; Soomro, S.A.; Shah, S.F. Mathematical modeling for ethanol production from molasses using thermotolerant kluyeromyces marxians. Sci. Int. 2016, 28, 319–322. [Google Scholar]
- Demirbas, A. Progress and recent trends in biofuels. Prog. Energy Combust. Sci. 2007, 33, 1–18. [Google Scholar] [CrossRef]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.J.; Negro, M.J. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour. Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef]
- Zentou, H.; Abidin, Z.Z.; Zouanti, M.; Greetham, D. Effect of operating conditions on molasses fermentation for bioethanol production. Int. J. Appl. Eng. Res. 2015, 12, 1–6. [Google Scholar]
- Gasmalla, M.A.A.; Yang, R.; Nikoo, M.; Man, S. Production of ethanol from sudanese sugar cane molasses and evaluation of its quality. J. Food Process Technol. 2012, 3, 163–165. [Google Scholar]
- Elena, P.; Gabriela, R.; Camelia, B.; Traian, H. Bioethanol production from molasses by different strains of Saccharomyces cerevisiae. Ann. Univ. Dunarea JOS Galati Fascicle VI Food Technol. 2009, 33, 49. [Google Scholar]
- Periyasamy, S.; Venkatachalam, S.; Ramasamy, S.; Srinivasan, V. Production of bio-ethanol from sugar molasses using Saccharomyces cerevisiae. Mod. Appl. Sci. 2009, 3, 32. [Google Scholar] [CrossRef]
- Dodić, J.M.; Vučurović, D.G.; Dodić, S.N.; Grahovac, J.A.; Popov, S.D.; Nedeljković, N.M. Kinetic modelling of batch ethanol production from sugar beet raw juice. Appl. Energy 2012, 99, 192–197. [Google Scholar] [CrossRef]
- Garhyan, P.; Elnashaie, S. Utilization of mathematical models to investigate the bifurcation and chaotic behavior of ethanol fermentors. Math. Comput. Model. 2004, 39, 381–427. [Google Scholar] [CrossRef]
- Oliveira, S.C.; Oliveira, R.C.; Tacin, M.V.; Gattás, E.A.L. Kinetic modeling and optimization of a batch ethanol fermentation process. J. Bioprocess. Biotech. 2016, 6, 266. [Google Scholar]
- Monod, J. La technique de culture continue, théorie et applications. Ann. Inst. Pasteur 1950, 79, 390–410. [Google Scholar]
- Rorke, D.; Gueguim Kana, E.B. Kinetics of bioethanol production from waste sorghum leaves using Saccharomyces cerevisiae BY4743. Fermentation 2017, 3, 19. [Google Scholar] [CrossRef]
- Srimachai, T.; Nuithitikul, K.; Sompong, O.; Kongjan, P.; Panpong, K. Optimization and Kinetic Modeling of Ethanol Production from Oil Palm Frond Juice in Batch Fermentation. Energy Procedia 2015, 79, 111–118. [Google Scholar] [CrossRef]
- Ariyajaroenwong, P.; Laopaiboon, P.; Salakkam, A.; Srinophakun, P.; Laopaiboon, L. Kinetic models for batch and continuous ethanol fermentation from sweet sorghum juice by yeast immobilized on sweet sorghum stalks. J. Taiwan Inst. Chem. Eng. 2016, 66, 210–216. [Google Scholar] [CrossRef]
- Manikandan, K.; Saravanan, V.; Viruthagiri, T. Kinetics studies on ethanol production from banana peel waste using mutant strain of Saccharomyces cerevisiae. Ind. J. Biotechnol. 2008, 7, 83–88. [Google Scholar]
- Ahmad, F.; Jameel, A.T.; Kamarudin, M.H.; Mel, M. Study of growth kinetic and modeling of ethanol production by Saccharomyces cerevisae. Afr. J. Biotechnol. 2011, 10, 18842–18846. [Google Scholar]
- Shafaghat, H.; Najafjour, G.D.; Rezaei, P.S.; Sharifzadeh, M. Growth kinetics and ethanol productivity of Saccharomyces cerevisiae PTCC 24860 on various carbon sources. World Appl. Sci. J. 2009, 7, 140–144. [Google Scholar]
- Raposo, S.; Pardão, J.M.; Diaz, I.; Lima-Costa, M.E. Kinetic modelling of bioethanol production using agro-industrial by-products. Int. J. Energy Environ. 2009, 3, 8. [Google Scholar]
- Singh, J.; Sharma, R. Growth kinetic and modeling of ethanol production by wilds and mutant Saccharomyces cerevisiae MTCC 170. Eur. J. Exp. Biol 2015, 5, 1–6. [Google Scholar]
- Comelli, R.N.; Seluy, L.G.; Isla, M.A. Performance of several Saccharomyces strains for the alcoholic fermentation of sugar-sweetened high-strength wastewaters: Comparative analysis and kinetic modelling. New Biotechnol. 2016, 33, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Guidini, C.Z.; Marquez, L.D.S.; de Almeida Silva, H.; de Resende, M.M.; Cardoso, V.L.; Ribeiro, E.J. Alcoholic fermentation with flocculant Saccharomyces cerevisiae in fed-batch process. Appl. Biochem. Biotechnol. 2014, 172, 1623–1638. [Google Scholar] [CrossRef] [PubMed]
- Kostov, G.; Popova, S.; Gochev, V.; Koprinkova-Hristova, P.; Angelov, M.; Georgieva, A. Modeling of batch alcohol fermentation with free and immobilized yeasts Saccharomyces cerevisiae 46 EVD. Biotechnol. Biotechnol. Equip. 2012, 26, 3021–3030. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of sugars, byproducts, and degradation products in liquid fraction process samples. Golden Natl. Renew. Energy Lab. 2006, 11. [Google Scholar]
- Lee, J.-W.; Rodrigues, R.C.L.B.; Kim, H.J.; Choi, I.-G.; Jeffries, T.W. The roles of xylan and lignin in oxalic acid pretreated corncob during separate enzymatic hydrolysis and ethanol fermentation. Bioresour. Technol. 2010, 101, 4379–4385. [Google Scholar] [CrossRef]
- Lee, J.-W.; Rodrigues, R.C.L.B.; Jeffries, T.W. Simultaneous saccharification and ethanol fermentation of oxalic acid pretreated corncob assessed with response surface methodology. Bioresour. Technol. 2009, 100, 6307–6311. [Google Scholar] [CrossRef]
- Sen, R.; Swaminathan, T. Response surface modeling and optimization to elucidate and analyze the effects of inoculum age and size on surfactin production. Biochem. Eng. J. 2004, 21, 141–148. [Google Scholar] [CrossRef]
- Kovárová-Kovar, K.; Egli, T. Growth kinetics of suspended microbial cells: from single-substrate-controlled growth to mixed-substrate kinetics. Microbiol. Mol. Biol. Rev. 1998, 62, 646–666. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Model | µmax (h−1) | Ks (g.L−1) | Ki (g.L−1) | Reference |
|---|---|---|---|---|---|
| Sorghum leaves | Monod | 0.176 | 10.11 | ---- | [17] |
| Oil palm frond juice | Monod | 0.150 | 10.21 | ---- | [18] |
| Sweet sorghum juice | Monod | 0.313 | 47.51 | ---- | [19] |
| Banana peels | Monod | 1.500 | 25.00 | ---- | [20] |
| Glucose | Monod | 0.084 | 213.60 | ---- | [21] |
| Glucose | Monod | 0.650 | 11.39 | ---- | [22] |
| Citrus waste pulp | Monod | 0.350 | 10.69 | ---- | [23] |
| Glucose | Monod | 0.133 | 3.70 | ---- | [24] |
| Beet molasses | Monod | 0.355 | 6.65 | ---- | [23] |
| Soft drinks mixture | Andrews | 0.606 | 65.53 | 0.029 | [25] |
| Sucrose | Andrews | 0.103 | 30.24 | 109.8 | [26] |
| Sugar cane juice | Andrews | 0.500 | 0.006 | 139.7 | [15] |
| Glucose | Andrews | 0.088 | 700 | 3.730 | [27] |
| Reduced Chi Squared | 7.90971 × 10−6 |
| Adjusted R-Squared | 0.99071 |
| μmax (h−1) | 0.5086 ± 0.04 |
| Ks (g.L−1) | 47.53789 ± 9.27 |
| Ki (g.L−1) | 181.01639 ± 29.14 |
| Initial Substrate Concentration (g.L−1) | Yx/s | Yp/s |
|---|---|---|
| 5 | 0.267 | 0.384 |
| 10 | 0.282 | 0.397 |
| 15 | 0.290 | 0.446 |
| 20 | 0.278 | 0.435 |
| 25 | 0.283 | 0.439 |
| Average | 0.280 ± 0.0084 | 0.420 ± 0.0028 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zentou, H.; Zainal Abidin, Z.; Yunus, R.; Awang Biak, D.R.; Zouanti, M.; Hassani, A. Modelling of Molasses Fermentation for Bioethanol Production: A Comparative Investigation of Monod and Andrews Models Accuracy Assessment. Biomolecules 2019, 9, 308. https://doi.org/10.3390/biom9080308
Zentou H, Zainal Abidin Z, Yunus R, Awang Biak DR, Zouanti M, Hassani A. Modelling of Molasses Fermentation for Bioethanol Production: A Comparative Investigation of Monod and Andrews Models Accuracy Assessment. Biomolecules. 2019; 9(8):308. https://doi.org/10.3390/biom9080308
Chicago/Turabian StyleZentou, Hamid, Zurina Zainal Abidin, Robiah Yunus, Dayang Radiah Awang Biak, Mustapha Zouanti, and Abdelkader Hassani. 2019. "Modelling of Molasses Fermentation for Bioethanol Production: A Comparative Investigation of Monod and Andrews Models Accuracy Assessment" Biomolecules 9, no. 8: 308. https://doi.org/10.3390/biom9080308
APA StyleZentou, H., Zainal Abidin, Z., Yunus, R., Awang Biak, D. R., Zouanti, M., & Hassani, A. (2019). Modelling of Molasses Fermentation for Bioethanol Production: A Comparative Investigation of Monod and Andrews Models Accuracy Assessment. Biomolecules, 9(8), 308. https://doi.org/10.3390/biom9080308