HMGB1-triggered inflammation inhibition of notoginseng leaf triterpenes against cerebral ischemia and reperfusion injury via MAPK and NF-κB signaling pathways

 ,
,
Abstract
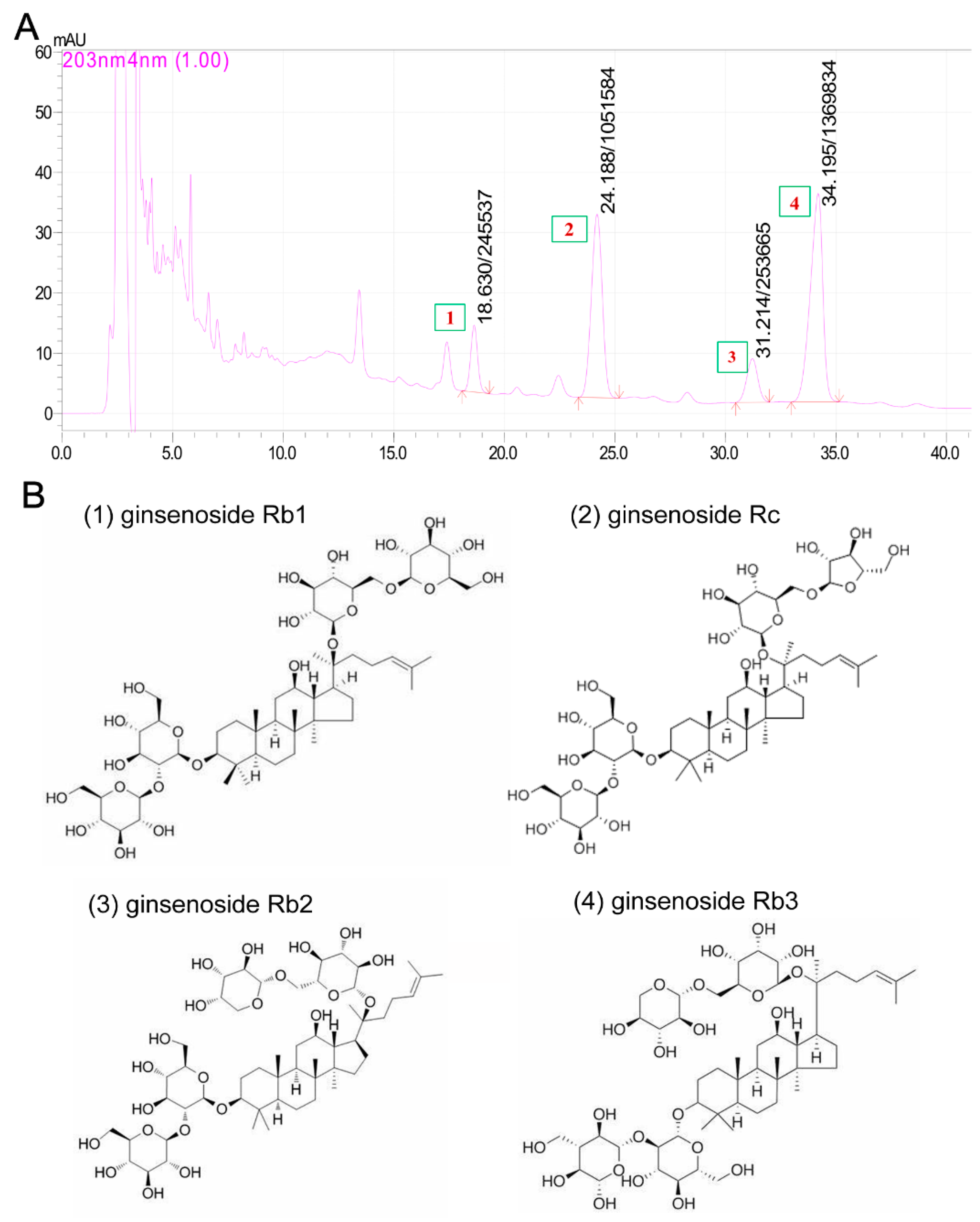
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods and Materials
2.1. Animals
2.2. Experimental Groups and Drug Administration
2.3. Transient Focal Brain Ischemia/Reperfusion Mode
2.4. Measurement of Neurological Deficit
2.5. TTC Staining
2.6. Detection of Brain Water Content
2.7. Measurement of BBB Permeability
2.8. Detection of Inflammatory Cytokines in Serums and Brain Tissues from Rats
2.9. Histopathological Examination
2.10. TUNEL Staining Assay
2.11. Immunofluorescence
2.12. Western Blot Analysis
2.13. Data and Statistical Analyses
3. Results
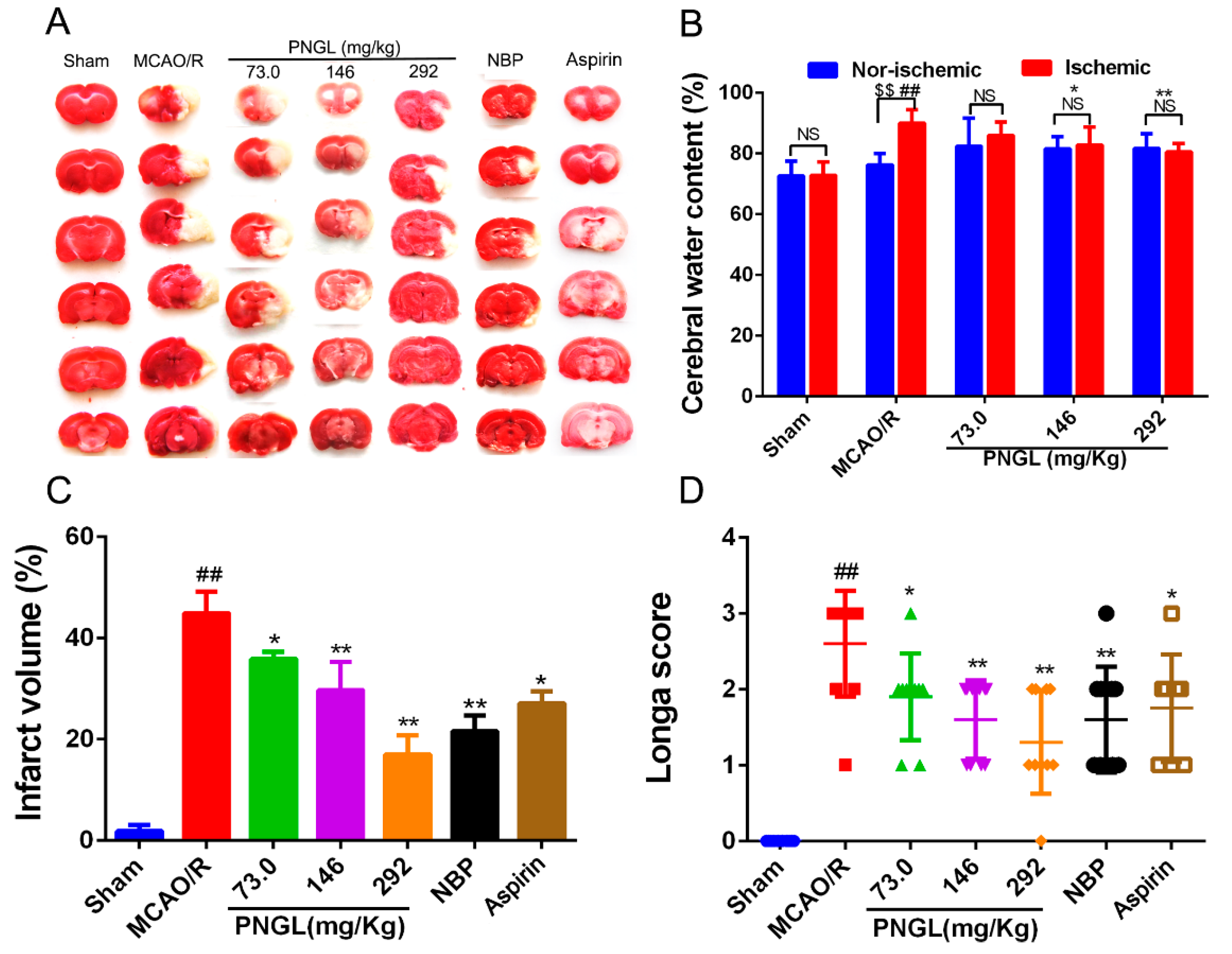
3.1. PNGL Improves Neurological Functions and Attenuate Brain Swellings and Infarcts in MCAO/R Rats
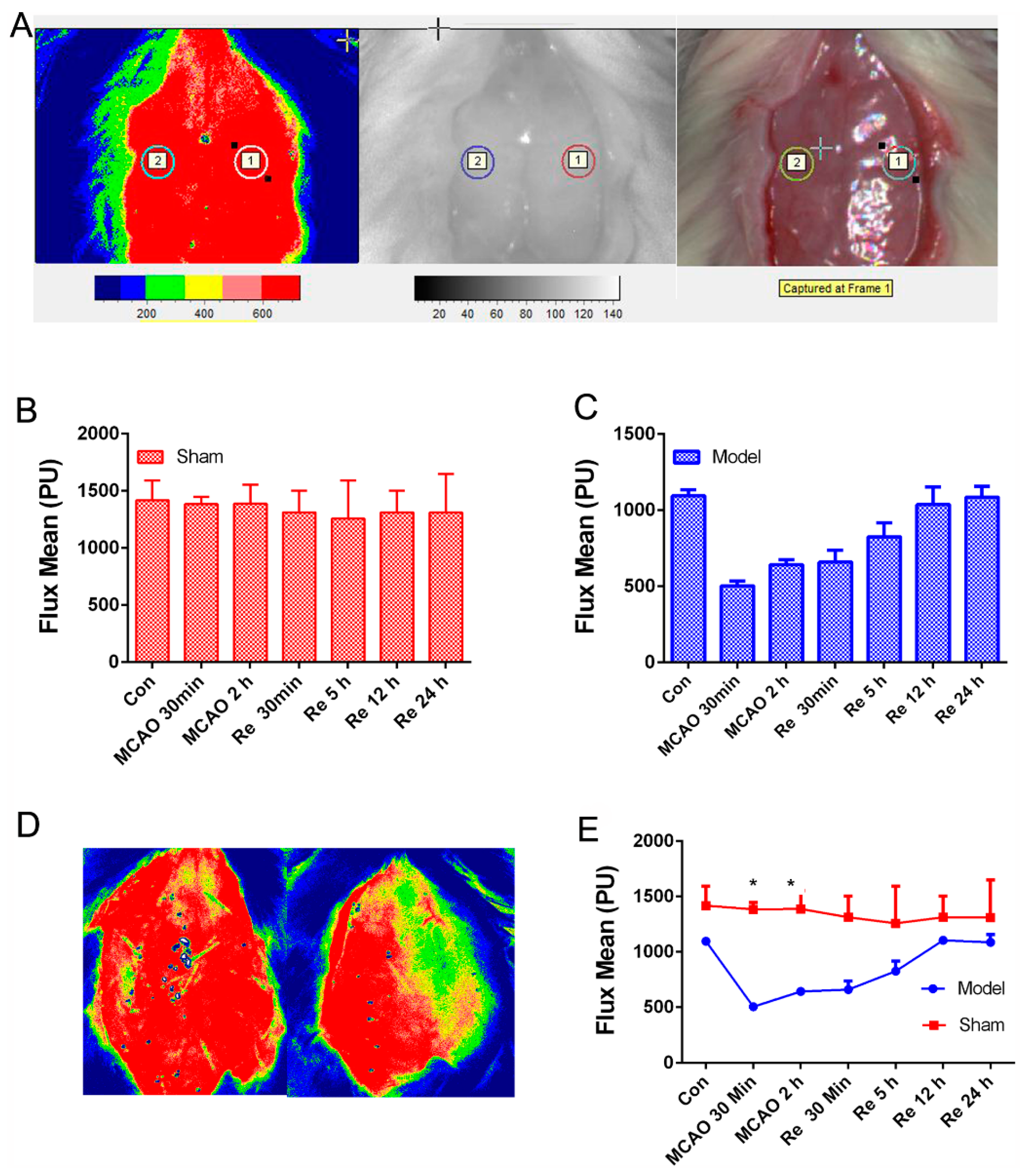
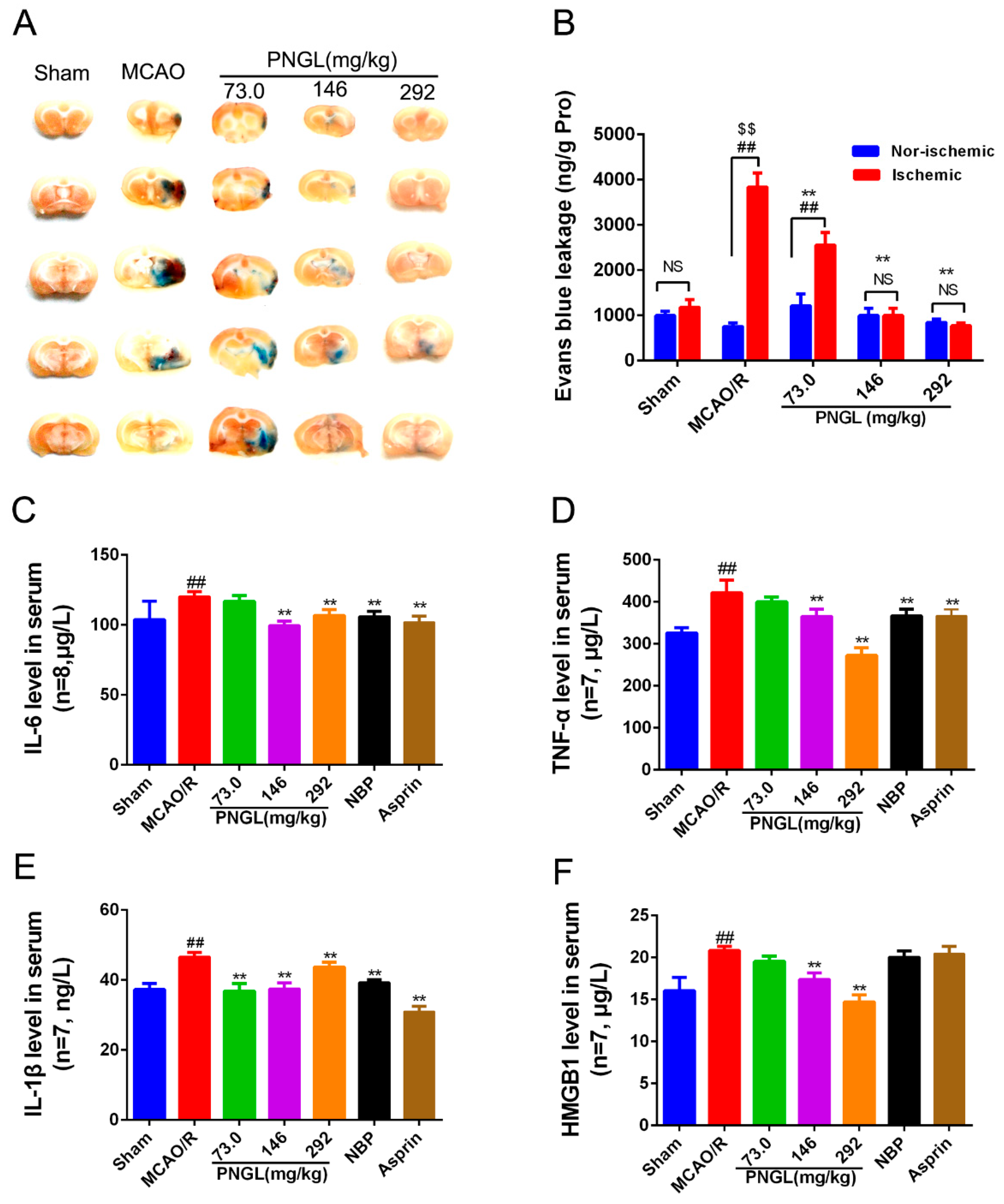
3.2. PNGL Alleviates BBB Disruption and Inflammatory Cytokines in MCAO/R Rats
3.3. PNGL Decreases Neuronal Apoptosis and Loss Caused By Ischemia
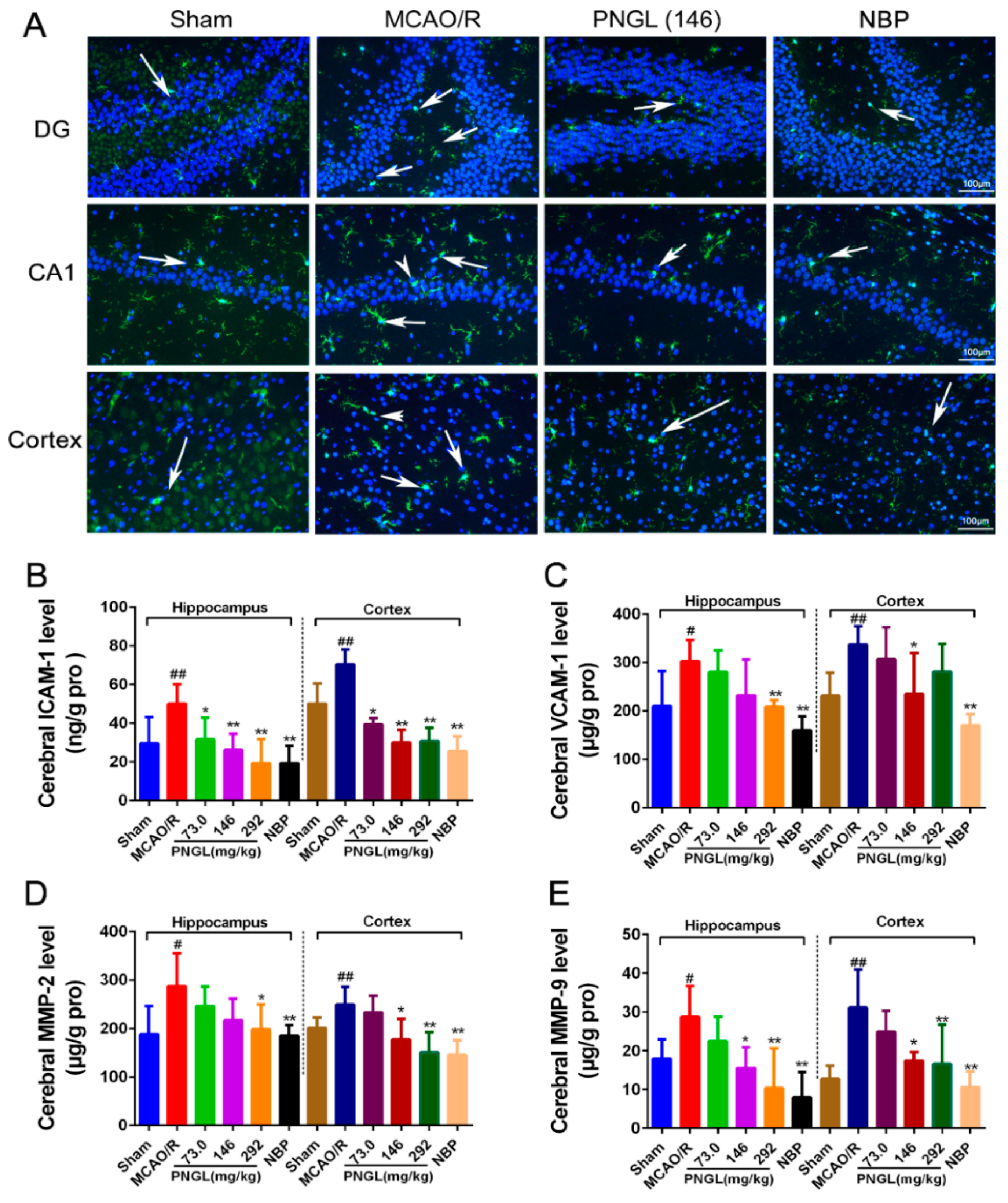
3.4. PNGL Downregulates Inflammatory Cytokines and Inhibits Microglia Activation in Ischemic Brains
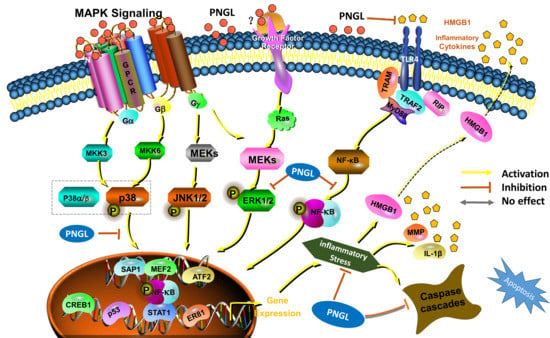
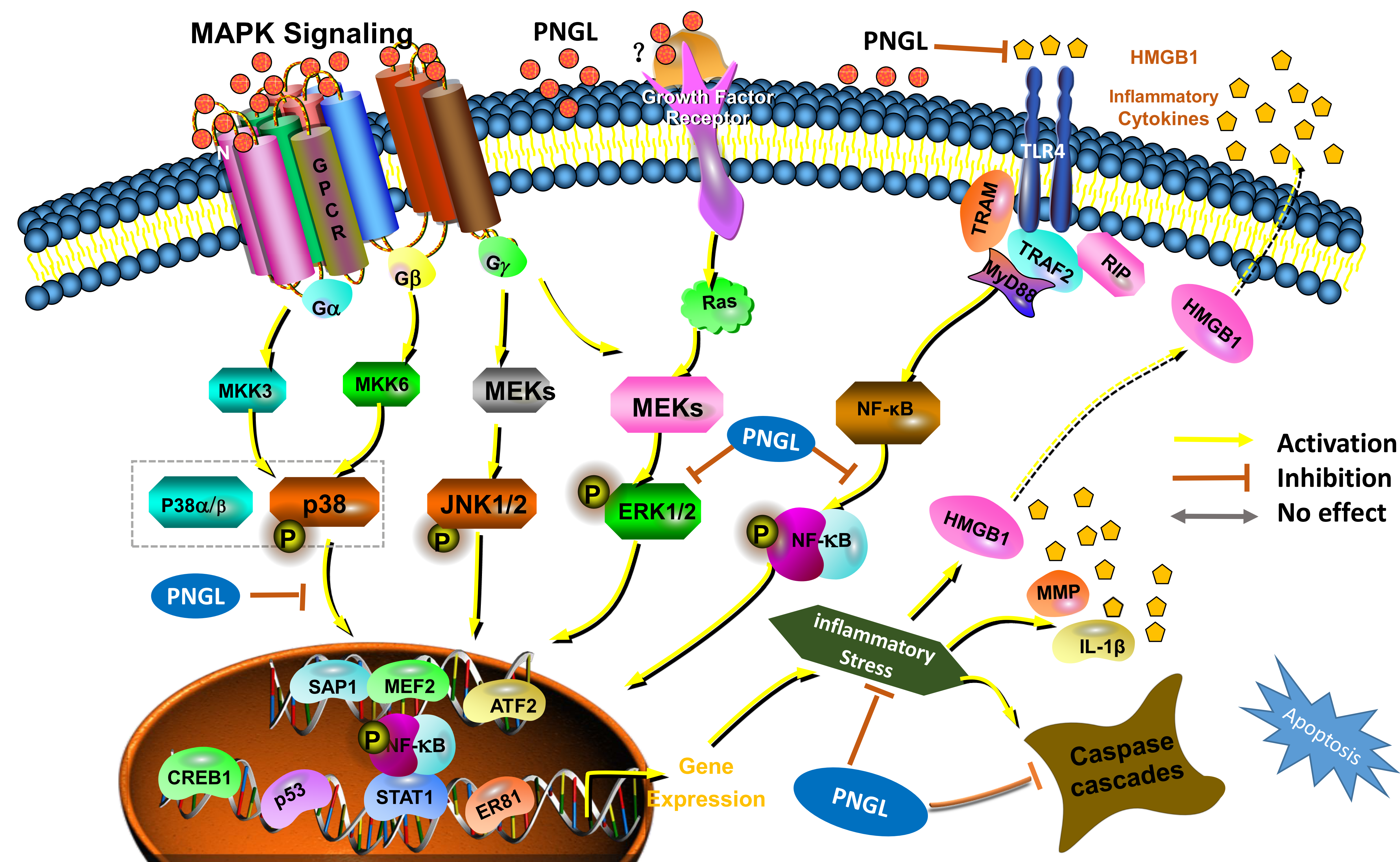
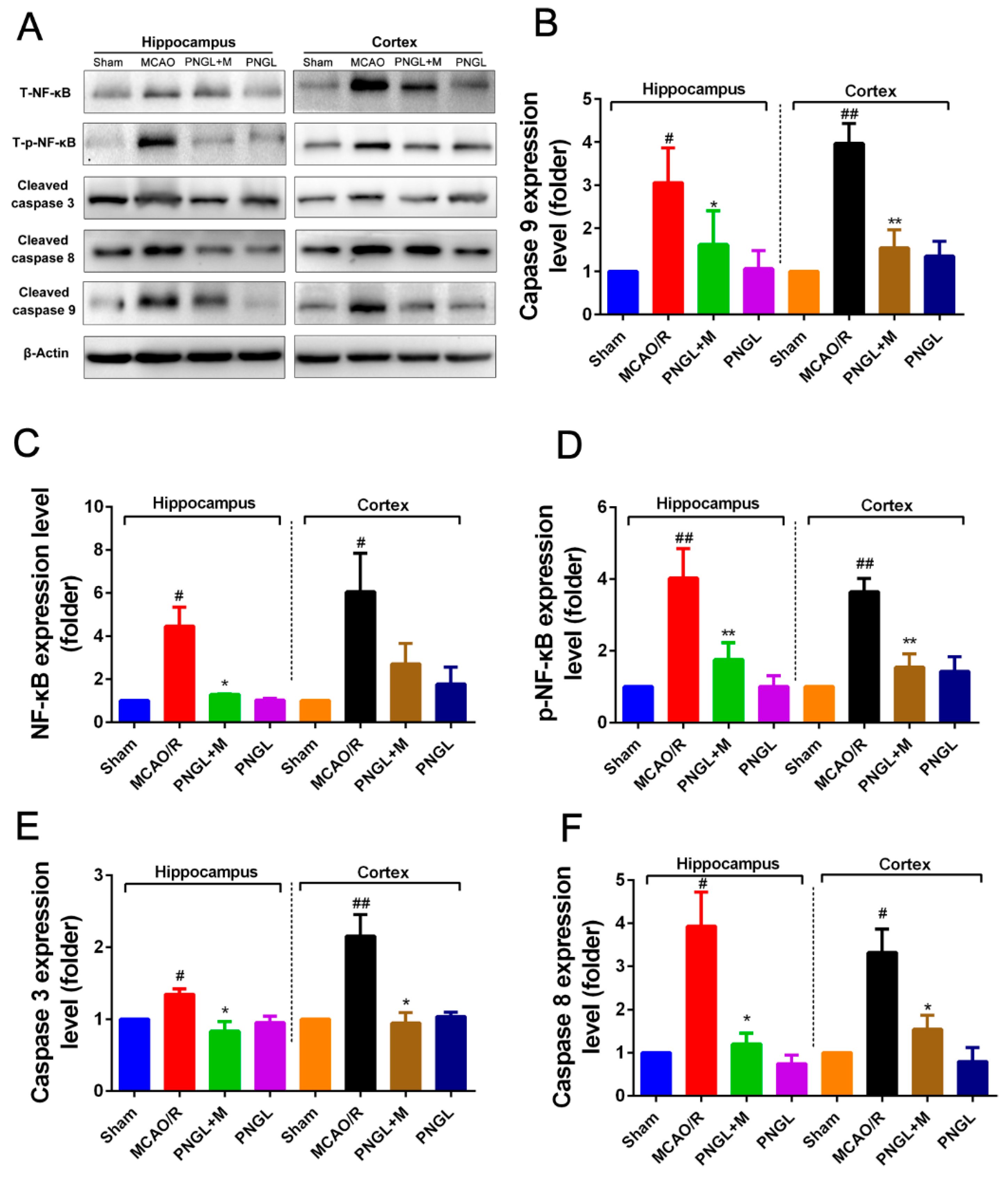
3.5. PNGL Inhibits NF-Κb Signaling Pathway
3.6. PNGL Regulates Mapks Signaling Pathway
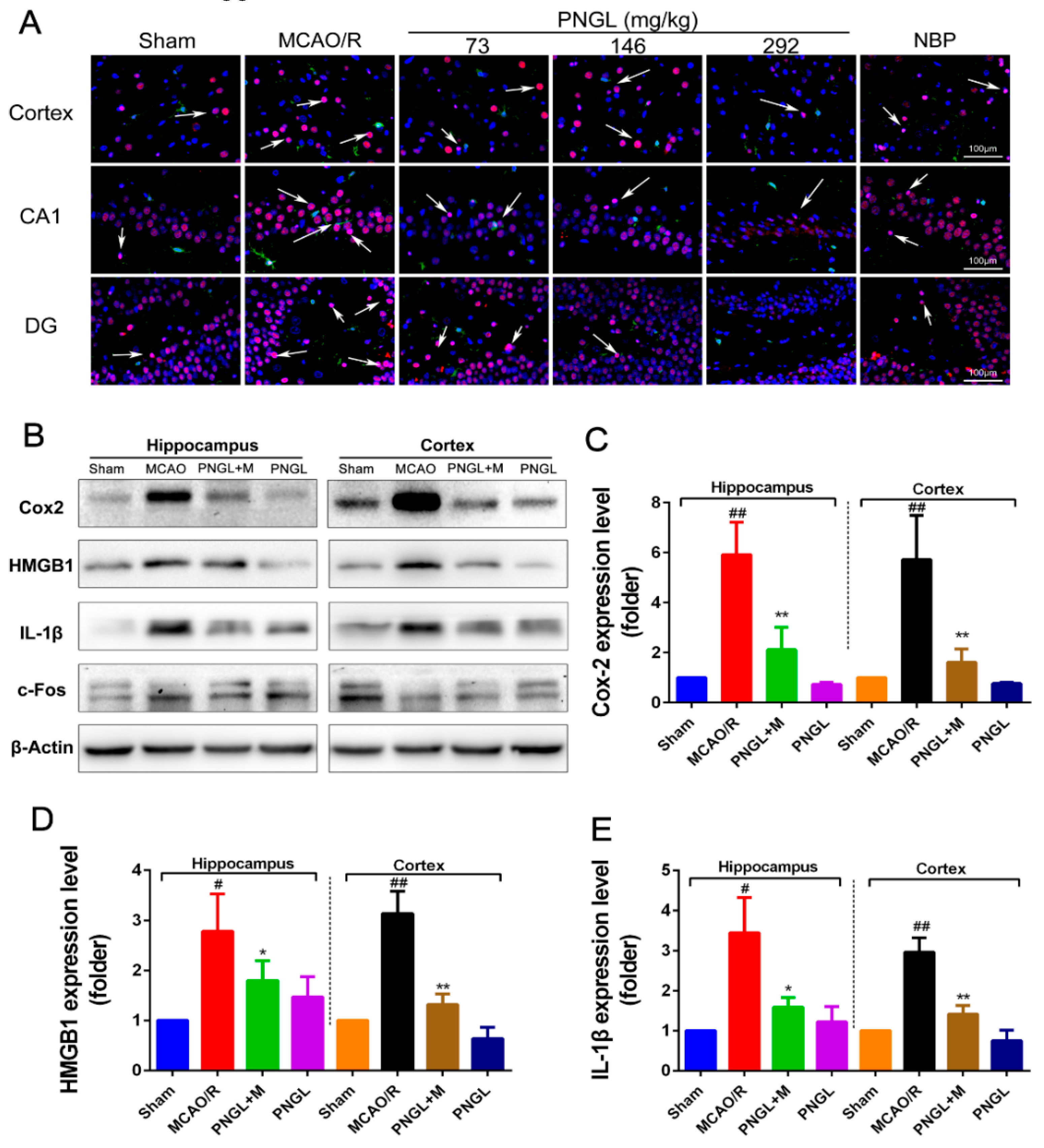
3.7. PNGL Inhibits HMGB1 Expression and Its HMGB1-Triggered Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jamison, J.T. Mechanisms of persistent translation arrest following global brain ischemia and reperfusion. Dissertations Theses, Wayne State University, Detroit, MI, USA, January 2011. [Google Scholar]
- Kamel, H.; Iadecola, C. Brain-Immune Interactions and Ischemic Stroke: Clinical Implications. JAMA Neurol. 2012, 69, 576–581. [Google Scholar]
- Xie, W.; Zhou, P.; Sun, Y.; Meng, X.; Dai, Z.; Sun, G.; Sun, X. Protective Effects and Target Network Analysis of Ginsenoside Rg1 in Cerebral Ischemia and Reperfusion Injury: A Comprehensive Overview of Experimental Studies. Cells 2018, 7, 270. [Google Scholar] [CrossRef] [PubMed]
- Antonino, T.; Riccardo, D.S.; Domenico, D.R.; Claudio, P.; Sergio, L.P.; Antonio, P.; Giuseppe, L. Effects of clinical and laboratory variables and of pretreatment with cardiovascular drugs in acute ischaemic stroke: A retrospective chart review from the GIFA study. Int. J. Cardiol. 2011, 151, 318–322. [Google Scholar]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—from mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, J.; Peng, Q.; Liu, Y.; Liu, W.; Luo, C.; Peng, J.; Li, J.; Yung, K.K.L.; Mo, Z. Neuroprotective effects of bilobalide on cerebral ischemia and reperfusion injury are associated with inhibition of pro-inflammatory mediator production and down-regulation of JNK1/2 and p38 MAPK activation. J. Neuroinflammation 2014, 11, 167. [Google Scholar] [CrossRef] [PubMed]
- Jianhua, Q.; Masaki, N.; Yumei, W.; Sims, J.R.; Sumei, Q.; Savitz, S.I.; Salvatore, S.; Moskowitz, M.A. Early release of HMGB-1 from neurons after the onset of brain ischemia. J. Cereb. Blood Flow Metab. 2008, 28, 927–938. [Google Scholar]
- Tao, X.; Sun, X.; Yin, L.; Han, X.; Xu, L.; Qi, Y.; Xu, Y.; Li, H.; Lin, Y.; Liu, K.; et al. Dioscin ameliorates cerebral ischemia/reperfusion injury through the downregulation of TLR4 signaling via HMGB-1 inhibition. Free. Radic. Boil. Med. 2015, 84, 103–115. [Google Scholar] [CrossRef]
- Yi, L.; Chang-Pei, G.; Shuang, Z.; Xi-Kun, Z.; Xue-Feng, L.; Yu-Quan, W.; Jin-Liang, Y.; Min, W. FIP200 is involved in murine pseudomonas infection by regulating HMGB1 intracellular translocation. Cell. Physiol. Biochem. Pharmacol. 2014, 33, 1733–1744. [Google Scholar]
- Cheng, Y.; Wang, D.; Wang, B.; Li, H.; Xiong, J.; Xu, S.; Chen, Q.; Tao, K.; Yang, X.; Zhu, Y.; et al. HMGB1 translocation and release mediate cigarette smoke–induced pulmonary inflammation in mice through a TLR4/MyD88-dependent signaling pathway. Mol. Boil. Cell 2017, 28, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.-H.; Hsieh, S.-Y.; Yen, C.-C.; Wu, H.-C.; Chen, K.-L.; Hung, D.-Z.; Chen, C.-H.; Wu, C.-C.; Su, Y.-C.; Chen, Y.-W.; et al. Involvement of oxidative stress-mediated ERK1/2 and p38 activation regulated mitochondria-dependent apoptotic signals in methylmercury-induced neuronal cell injury. Toxicol. Lett. 2011, 204, 71–80. [Google Scholar] [CrossRef]
- Pietri, M.; Caprini, A.; Mouillet-Richard, S.; Pradines, E.; Ermonval, M.; Grassi, J.; Kellermann, O.; Schneider, B. Overstimulation of PrPCSignaling Pathways by Prion Peptide 106-126 Causes Oxidative Injury of Bioaminergic Neuronal Cells. J. Boil. Chem. 2006, 281, 28470–28479. [Google Scholar] [CrossRef] [PubMed]
- Procaccio, V.; Bris, C.; De La Barca, J.C.; Oca, F.; Chevrollier, A.; Amati-Bonneau, P.; Bonneau, D.; Reynier, P. Perspectives of drug-based neuroprotection targeting mitochondria. Rev. Neurol. 2014, 170, 390–400. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Nan, G. The Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway as a Discovery Target in Stroke. J. Mol. Neurosci. 2016, 59, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Eto, M.; Kozaki, K.; Akishita, M.; Okabe, T.; Ouchi, Y. Raloxifene analogue LY117018 suppresses oxidative stress-induced endothelial cell apoptosis through activation of ERK1/2 signaling pathway. Eur. J. Pharmacol. 2008, 589, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Kovalská, M.; Kovalska, L.; Pavlíková, M.; Janickova, M.; Mikušková, K.; Adamkov, M.; Kaplan, P.; Tatarkova, Z.; Lehotský, J. Intracellular Signaling MAPK Pathway After Cerebral Ischemia–Reperfusion Injury. Neurochem. Res. 2012, 37, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Lu, S.; Luo, Y.; Wang, S.; Yang, K.; Zhai, Y.; Sun, G.; Sun, X. Attenuation of TNF-α-Induced Inflammatory Injury in Endothelial Cells by Ginsenoside Rb1 via Inhibiting NF-κB, JNK and p38 Signaling Pathways. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Sung-Chun, T.; Arumugam, T.V.; Xiangru, X.; Aiwu, C.; Mughal, M.R.; Gyu, J.D.; Lathia, J.D.; Siler, D.A.; Srinivasulu, C.; Xin, O. Pivotal role for neuronal Toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci USA 2007, 104, 13798–13803. [Google Scholar] [Green Version]
- Wang, X.; Wang, C.; Wang, J.; Zhao, S.; Zhang, K.; Wang, J.; Zhang, W.; Wu, C.; Yang, J. Pseudoginsenoside-F11 (PF11) exerts anti-neuroinflammatory effects on LPS-activated microglial cells by inhibiting TLR4-mediated TAK1/IKK/NF-κB, MAPKs and Akt signaling pathways. Neuropharmacology 2014, 79, 642–656. [Google Scholar] [CrossRef]
- Liesz, A.; Dalpke, A.; Mracsko, E.; Antoine, D.J.; Roth, S.; Zhou, W.; Yang, H.; Na, S.-Y.; Akhisaroglu, M.; Fleming, T.; et al. DAMP Signaling is a Key Pathway Inducing Immune Modulation after Brain Injury. J. Neurosci. 2015, 35, 583–598. [Google Scholar] [CrossRef] [Green Version]
- Eckert, B. Acute Stroke Therapy 1981–2009*. Clin. Neuroradiol. 2009, 19, 8. [Google Scholar] [CrossRef]
- Yang, X.; Xiong, X.; Wang, H.; Wang, J. Protective Effects of Panax Notoginseng Saponins on Cardiovascular Diseases: A Comprehensive Overview of Experimental Studies. Evidence-Based Complement. Altern. Med. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bao-Ying, H.; Xian-Jin, L.; Ren, Q.; Zheng-Lin, J.; Li-Hua, X.; Guo-Hua, W.; Xia, L.; Bin, P. Treatment with ginseng total saponins improves the neurorestoration of rat after traumatic brain injury. J. Ethnopharmacol. 2014, 155, 1243–1255. [Google Scholar]
- Xie, W.; Meng, X.; Zhai, Y.; Zhou, P.; Ye, T.; Wang, Z.; Sun, G.; Sun, X. Panax Notoginseng Saponins: A Review of Its Mechanisms of Antidepressant or Anxiolytic Effects and Network Analysis on Phytochemistry and Pharmacology. Molecules 2018, 23, 940. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Fan, J.; Han, J. Ameliorating effects of traditional Chinese medicine preparation, Chinese materia medica and active compounds on ischemia/reperfusion-induced cerebral microcirculatory disturbances and neuron damage. Acta Pharm. Sin. B 2015, 5, 8–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, H.; Zhang, C.; Shi, Z.-B.; Yang, H.-Q.; Wang, K.-Z. Protective effects and mechanism of Panax Notoginseng saponins on oxidative stress-induced damage and apoptosis of rabbit bone marrow stromal cells. Chin. J. Integr. Med. 2010, 16, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; He, S.; Sun, Y.; Meng, X.; Sun, G.; Sun, X. Ginsenoside Rb1 as an Anti-Diabetic Agent and Its Underlying Mechanism Analysis. Cells 2019, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; Meng, X.; Zhai, Y.; Dong, X.; Zhang, X.; Sun, G.; Sun, X. Notoginsenoside R1 Ameliorates Diabetic Retinopathy through PINK1-Dependent Activation of Mitophagy. Cells 2019, 8, 213. [Google Scholar] [CrossRef]
- Zhou, P.; Xie, W.; Sun, Y.; Dai, Z.; Li, G.; Sun, G.; Sun, X. Ginsenoside Rb1 and mitochondria: A short review of the literature. Mol. Cell. Probes 2019, 43, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; Luo, Y.; Lu, S.; Dai, Z.; Wang, R.; Zhang, X.; Li, G.; Sun, G.; Sun, X. Inhibitory Effects of Ginsenoside Rb1 on Early Atherosclerosis in ApoE-/- Mice via Inhibition of Apoptosis and Enhancing Autophagy. Molecules 2018, 23, 2912. [Google Scholar] [CrossRef]
- Zhou, N.; Tang, Y.; Keep, R.F.; Ma, X.; Xiang, J. Antioxidative effects of Panax notoginseng saponins in brain cells. Phytomedicine 2014, 21, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Y.; Wang, S.; Li, C.-J.; Ma, J.; Chen, F.-Y.; Peng, Y.; Wang, X.-L.; Zhang, D.-M. Dammarane-type saponins from the leaves of Panax notoginseng and their neuroprotective effects on damaged SH-SY5Y cells. Phytochem. 2018, 145, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.-L.; Pan, H.-Q.; Wang, H.; Yao, S.; Yang, W.-Z.; Hou, J.-J.; Jin, Q.-H.; Wu, W.-Y.; Guo, D.-A. Global profiling combined with predicted metabolites screening for discovery of natural compounds: Characterization of ginsenosides in the leaves of Panax notoginseng as a case study. J. Chromatogr. A 2018, 1538, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.H.; Li, X.Z.; Shang, X.H.; Liu, J.X. Protective effects of saponines of stem and leaf of Panax notoginseng on acute myocardial ischemia in anaesthetic dogs. Chin. Mater. Med. 2006, 31, 62–65. (In Chinese) [Google Scholar]
- Chen, Z.H.; Wang, D.C.; Li, H.L.; Wei, J.X.; Wang, J.F.; Du, Y.C. Hemodynamic effects of san chi (Panax notoginseng) root, leaf, flower and saponins on anesthetized dogs. Acta Pharm. Sin. 1983, 18, 818–822. (In Chinese) [Google Scholar]
- Meng, X.; Xie, W.; Xu, Q.; Liang, T.; Xu, X.; Sun, G.; Sun, X. Neuroprotective Effects of Radix Scrophulariae on Cerebral Ischemia and Reperfusion Injury via MAPK Pathways. Molecules 2018, 23, 2401. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.M.; Jung, B.J.; Lee, J.I.; Choi, S.K.; Kang, S.K. Middle Cerebral Artery Occlusion in Rats: Evaluation of the Model With or Without Craniectomy. J. Korean Neurosurg Soc. 1994, 23, 3–11. [Google Scholar]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84. [Google Scholar] [CrossRef] [PubMed]
- Bederson, J.B.; Pitts, L.H.; Tsuji, M.; Nishimura, M.C.; Davis, R.L.; Bartkowski, H. Rat middle cerebral artery occlusion: evaluation of the model and development of a neurologic examination. Stroke 1986, 17, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Meng, X.; Luo, Y.; Wu, Y.; Ye, T.; Zhou, P.; Ding, S.; Wang, M.; Lu, S.; Zhu, L. Notoginsenoside R1 ameliorates diabetic encephalopathy by activating the Nrf2 pathway and inhibiting NLRP3 inflammasome activation. Oncotarget 2018, 9, 9344–9363. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Yang, Y.; Zhang, P. Neuroprotective effects of penehyclidine hydrochloride against cerebral ischemia/reperfusion injury in mice. Brain Res. Bull. 2016, 121, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Muir, K.W.; Pippa, T.; Naveed, S.; Elizabeth, W. Inflammation and ischaemic stroke. Curr. Opin. Neurol. 2007, 20, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, P.; Zhang, X.; Li, D.; Wang, R.; Ji, C.; Li, G.; Holscher, C. The diabetes drug semaglutide reduces infarct size, inflammation, and apoptosis, and normalizes neurogenesis in a rat model of stroke. Neuropharmacology 2019, 158, 107748. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.E.; Dong, Y.; Lu, Y.; Tucker, D.; Wang, R.; Zhang, Q. Beneficial Effects of a CaMKIIα Inhibitor TatCN21 Peptide in Global Cerebral Ischemia. J. Mol. Neurosci. 2017, 61, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Park, B.-K.; Kim, Y.H.; Shim, I.; Kang, I.-C.; Lee, M.Y. Antidepressant Effect of Fraxinus rhynchophylla Hance Extract in a Mouse Model of Chronic Stress-Induced Depression. BioMed Res. Int. 2018, 2018, 8249563. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; Luo, Y.; Lu, S.; Dai, Z.; Wang, R.; Sun, G.; Sun, X. Protective Effects of Total Saponins of Aralia elata (Miq.) on Endothelial Cell Injury Induced by TNF-alpha via Modulation of the PI3K/Akt and NF-kappaB Signalling Pathways. Int. J. Mol. Sci. 2018, 20. [Google Scholar] [CrossRef]
- Cheng, M.; Yang, L.; Dong, Z.; Wang, M.; Sun, Y.; Liu, H.; Wang, X.; Sai, N.; Huang, G.; Zhang, X. Folic acid deficiency enhanced microglial immune response via the Notch1/nuclear factor kappa B p65 pathway in hippocampus following rat brain I/R injury and BV2 cells. J. Cell. Mol. Med. 2019, 23, 4795–4807. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, X.B.; Xu, Y.; Hou, X.Y. A Brief Ischemic Postconditioning Protects Against Amyloid-beta Peptide Neurotoxicity by Downregulating MLK3-MKK3/6-P38MAPK Signal in Rat Hippocampus. J. Alzheimer’s Dis. 2019, 1–14. [Google Scholar]
- Singh, R.; Kulikowicz, E.; Santos, P.T.; Koehler, R.C.; Martin, L.J.; Lee, J.K. Spatial T-maze identifies cognitive deficits in piglets 1 month after hypoxia-ischemia in a model of hippocampal pyramidal neuron loss and interneuron attrition. Behav. Brain Res. 2019, 369, 111921. [Google Scholar] [CrossRef]
- Yin, H.Z.; Wang, H.L.; Ji, S.G.; Medvedeva, Y.V.; Tian, G.; Bazrafkan, A.K.; Maki, N.Z.; Akbari, Y.; Weiss, J.H. Rapid Intramitochondrial Zn2+ Accumulation in CA1 Hippocampal Pyramidal Neurons After Transient Global Ischemia: A Possible Contributor to Mitochondrial Disruption and Cell Death. J. Neuropathol. Exp. Neurol. 2019, 78, 655–664. [Google Scholar] [CrossRef]
- Ramirez-Sanchez, J.; Pires, E.N.S.; Meneghetti, A.; Hansel, G.; Nunez-Figueredo, Y.; Pardo-Andreu, G.L.; Ochoa-Rodriguez, E.; Verdecia-Reyes, Y.; Delgado-Hernandez, R.; Salbego, C.; et al. JM-20 Treatment After MCAO Reduced Astrocyte Reactivity and Neuronal Death on Peri-infarct Regions of the Rat Brain. Mol. Neurobiol. 2019, 56, 502–512. [Google Scholar] [CrossRef]
- Xiong, T.Q.; Chen, L.M.; Gui, Y.; Jiang, T.; Tan, B.H.; Li, S.L.; Li, Y.C. The effects of epothilone D on microtubule degradation and delayed neuronal death in the hippocampus following transient global ischemia. J. Chem. Neuroanat. 2019, 98, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhou, H.; Qu, Z.; Hou, J.; Chen, W.; Cai, W.; Cheng, Q.; Chuang, D.Y.; Chen, S.; Li, S.; et al. From Analysis of Ischemic Mouse Brain Proteome to Identification of Human Serum Clusterin as a Potential Biomarker for Severity of Acute Ischemic Stroke. Transl. Stroke Res. 2019, 10, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, Z.; Cui, J.; Mccrary, M.L.; Song, H.; Mobashery, S.; Chang, M.; Gu, Z. Early Abrogation of Gelatinase Activity Extends the Time Window for tPA Thrombolysis after Embolic Focal Cerebral Ischemia in Mice. Eneuro 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H. Systemic Inflammation and Cardiovascular Comorbidity in Psoriasis Patients: Causes and Consequences. Front. Immunol. 2018, 9, 579. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, G.C.; Chantler, P.D.; Barr, T.L. Stroke-associated pattern of gene expression previously identified by machine-learning is diagnostically robust in an independent patient population. Genom. Data 2017, 14, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Qin, W.; Hu, X.; Mu, S.; Zhu, J.; Lu, W.; Luo, Y. Lentivirus-mediated overexpression of OTULIN ameliorates microglia activation and neuroinflammation by depressing the activation of the NF-κB signaling pathway in cerebral ischemia/reperfusion rats. J. Neuroinflammation 2018, 15, 83. [Google Scholar] [CrossRef]
- Laird, M.D.; Shields, J.S.; Sangeetha, S.R.; Kimbler, D.E.; R David, F.; Basheer, S.; Patrick, Y.; Nathan, Y.; Vender, J.R.; Dhandapani, K.M. High mobility group box protein-1 promotes cerebral edema after traumatic brain injury via activation of toll-like receptor 4. Glia 2013, 62, 26–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao-Juan, F.; Shu-Xia, L.; Chao, W.; Peng-Peng, K.; Qing-Juan, L.; Jun, H.; Hong-Bo, L.; Fan, L.; Yu-Jun, Z.; Xiao-Hui, F. The PTEN/PI3K/Akt signaling pathway mediates HMGB1-induced cell proliferation by regulating the NF-κB/cyclin D1 pathway in mouse mesangial cells. Am. J. Physiology Cell Physiol. 2014, 306, 1119–1128. [Google Scholar]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, W.; Zhu, T.; Dong, X.; Nan, F.; Meng, X.; Zhou, P.; Sun, G.; Sun, X. HMGB1-triggered inflammation inhibition of notoginseng leaf triterpenes against cerebral ischemia and reperfusion injury via MAPK and NF-κB signaling pathways. Biomolecules 2019, 9, 512. https://doi.org/10.3390/biom9100512
Xie W, Zhu T, Dong X, Nan F, Meng X, Zhou P, Sun G, Sun X. HMGB1-triggered inflammation inhibition of notoginseng leaf triterpenes against cerebral ischemia and reperfusion injury via MAPK and NF-κB signaling pathways. Biomolecules. 2019; 9(10):512. https://doi.org/10.3390/biom9100512
Chicago/Turabian StyleXie, Weijie, Ting Zhu, Xi Dong, Fengwei Nan, Xiangbao Meng, Ping Zhou, Guibo Sun, and Xiaobo Sun. 2019. "HMGB1-triggered inflammation inhibition of notoginseng leaf triterpenes against cerebral ischemia and reperfusion injury via MAPK and NF-κB signaling pathways" Biomolecules 9, no. 10: 512. https://doi.org/10.3390/biom9100512
APA StyleXie, W., Zhu, T., Dong, X., Nan, F., Meng, X., Zhou, P., Sun, G., & Sun, X. (2019). HMGB1-triggered inflammation inhibition of notoginseng leaf triterpenes against cerebral ischemia and reperfusion injury via MAPK and NF-κB signaling pathways. Biomolecules, 9(10), 512. https://doi.org/10.3390/biom9100512